
Protective Effect against Neosporosis Induced by Intranasal Immunization with Neospora caninum Membrane Antigens Plus Carbomer-Based Adjuvant

 , , and
, , and 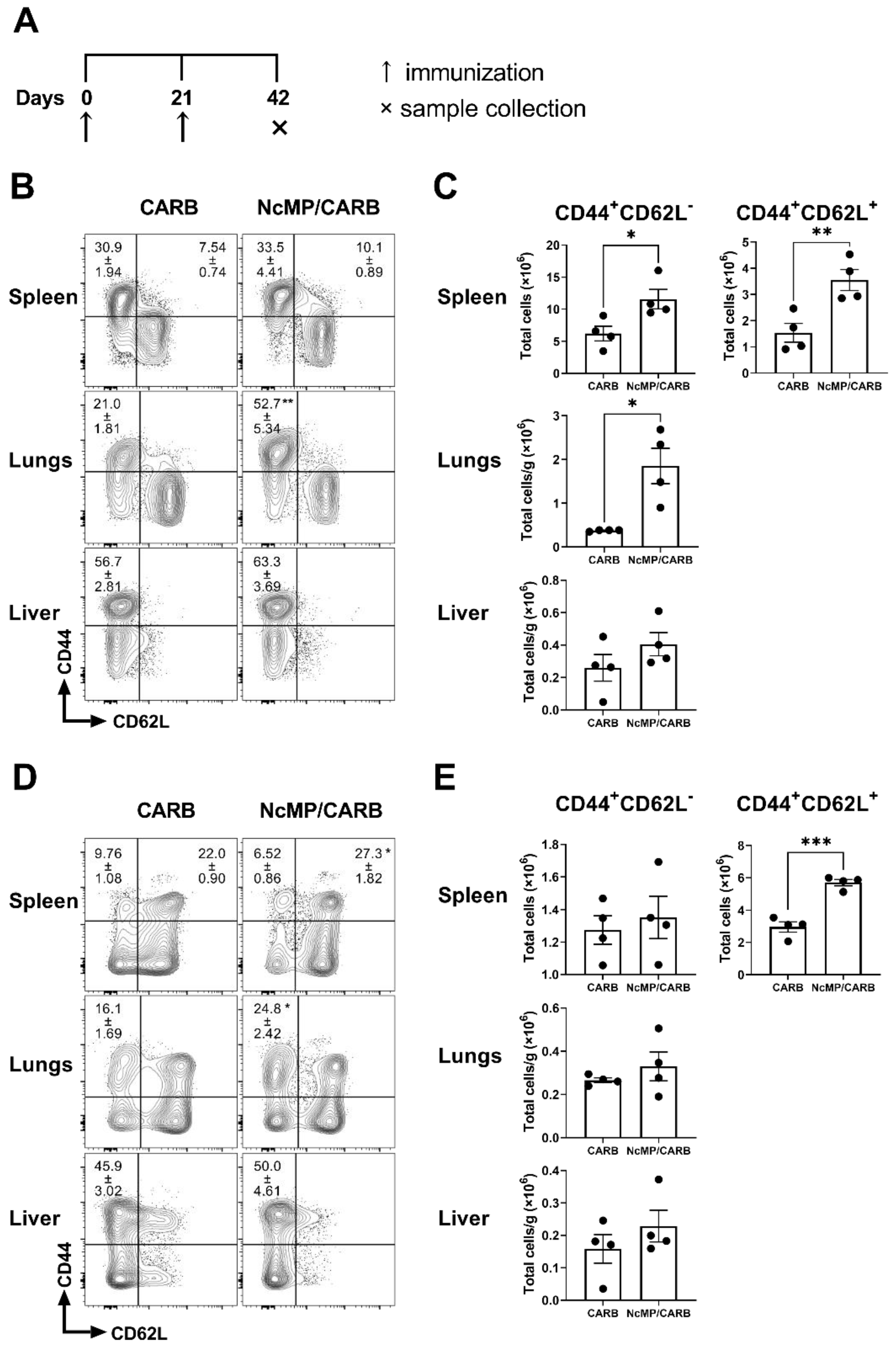{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Growth of Parasites and Preparation of Tachyzoite Lysates and Cell Membrane Extracts
2.3. Immunizations and Tissue Sample Collection
2.4. Antibody Detection
2.5. In Vitro Cell Cultures and Cytokine Detection
2.6. Flow Cytometry Analysis
2.7. DNA Extraction and Real-Time PCR Analysis
2.8. Statistical Analysis
3. Results
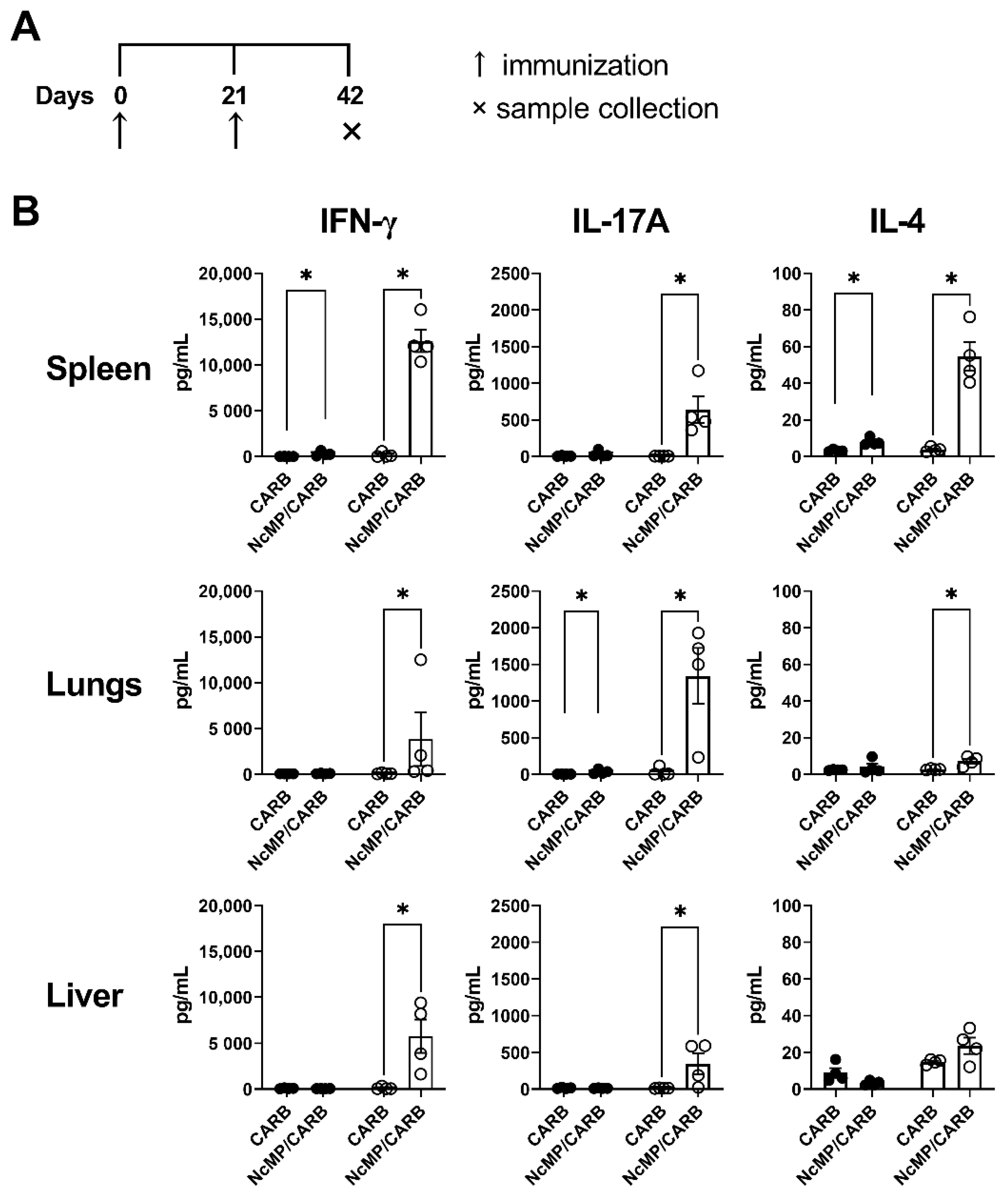
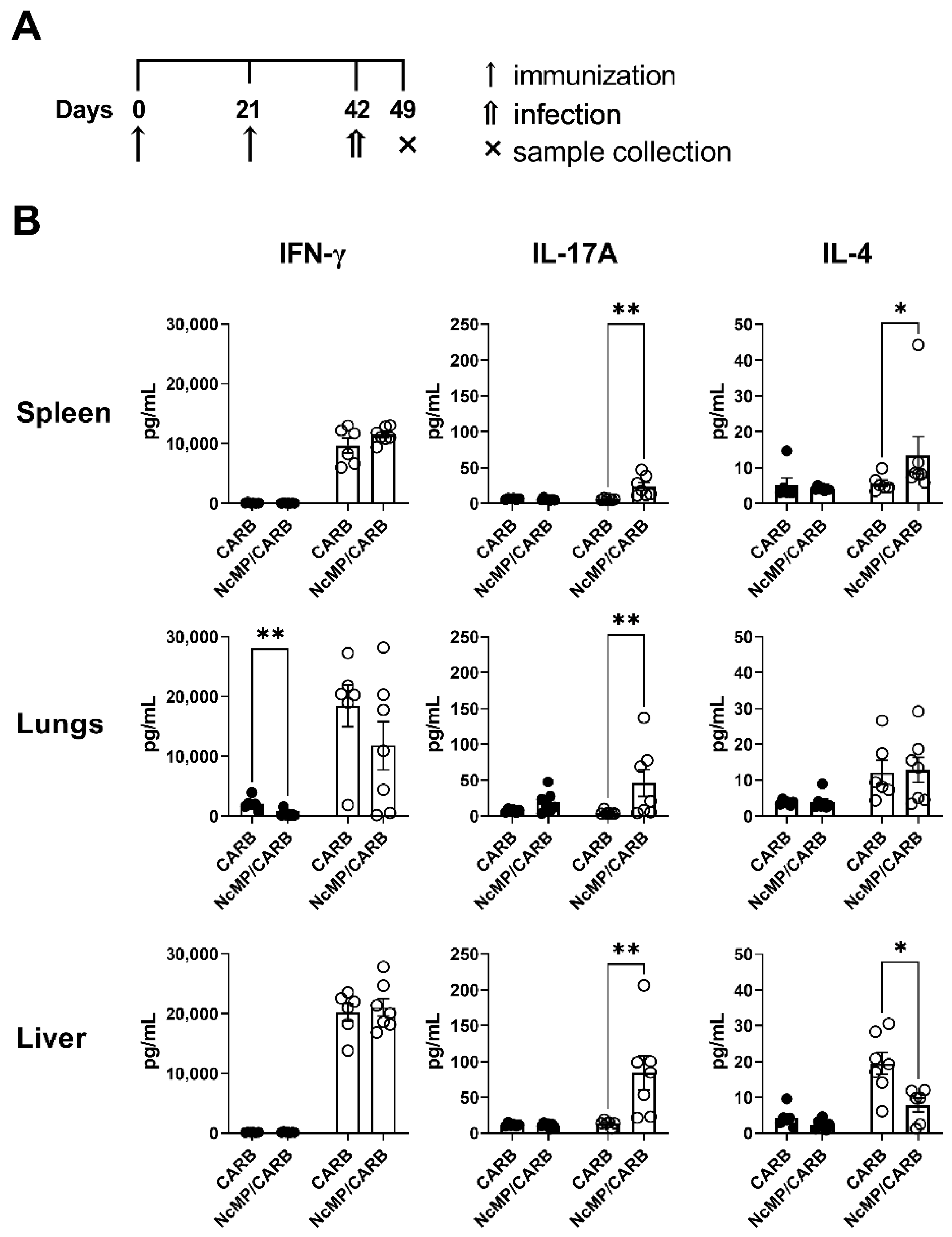
3.1. T Cell Response to N. caninum Antigens Induced by Immunization
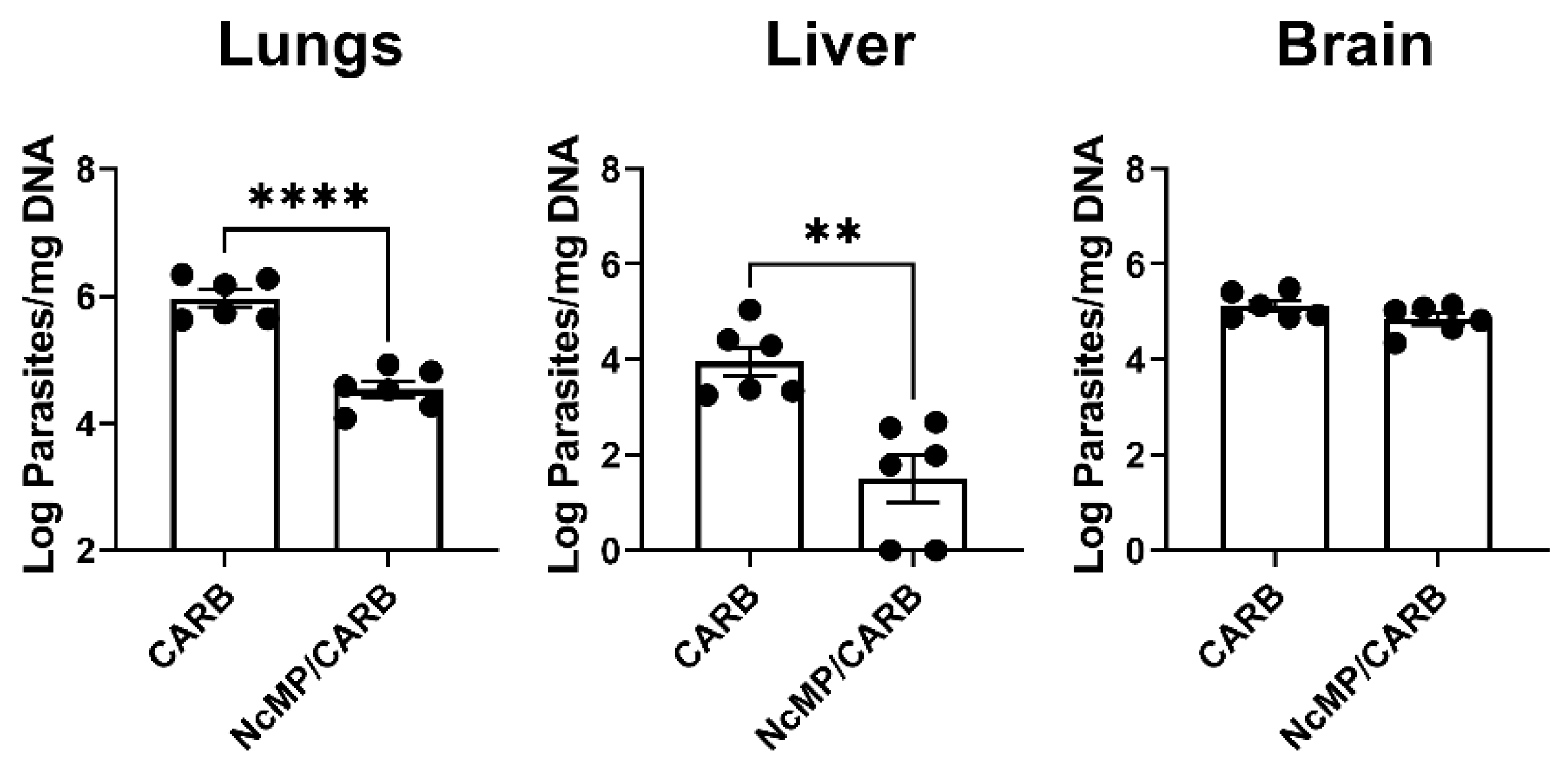
3.2. Parasitic Burden in Immunized Mice
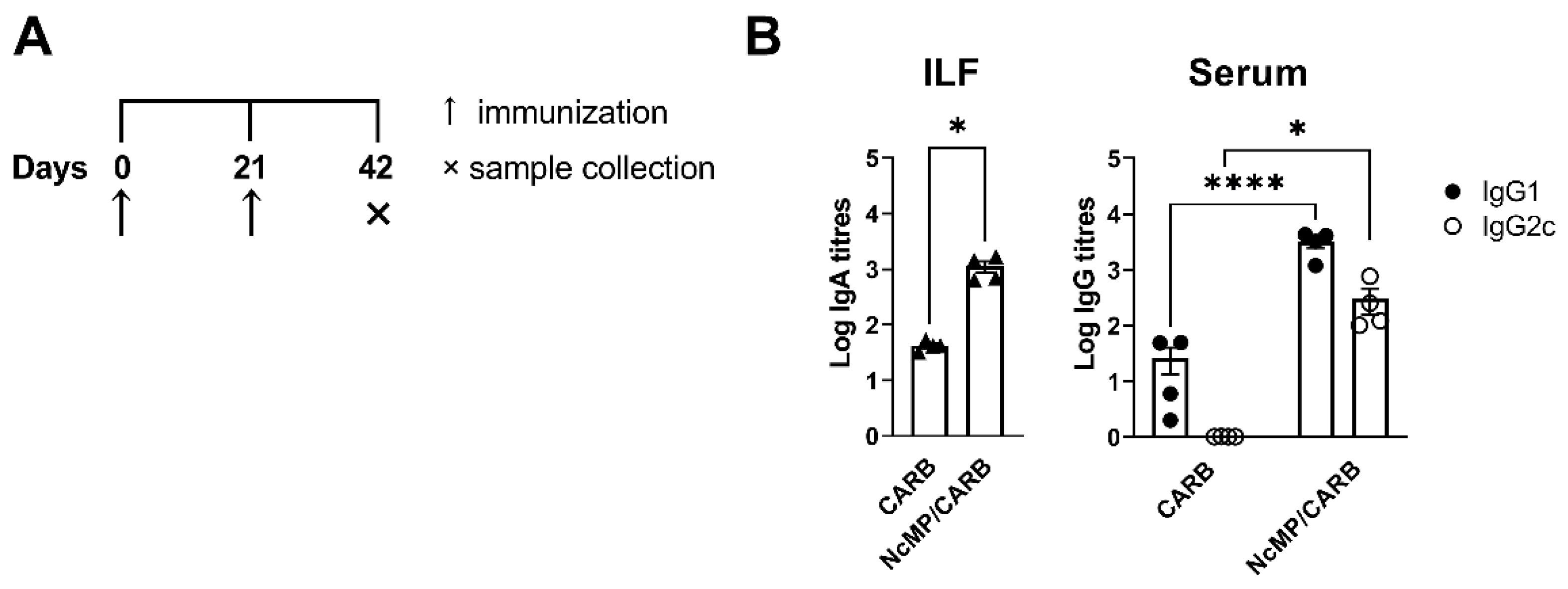
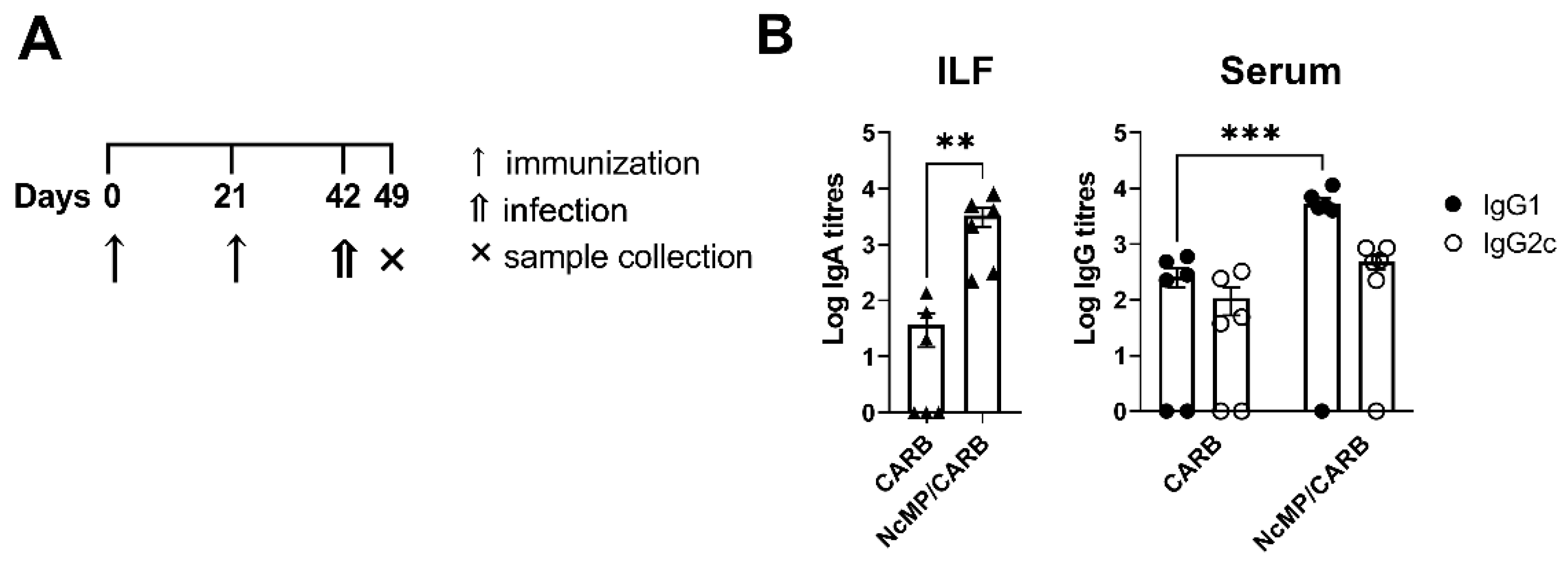
3.3. Parasite-Specific Antibody Levels in Infected Immunized and Nonimmunized Mice
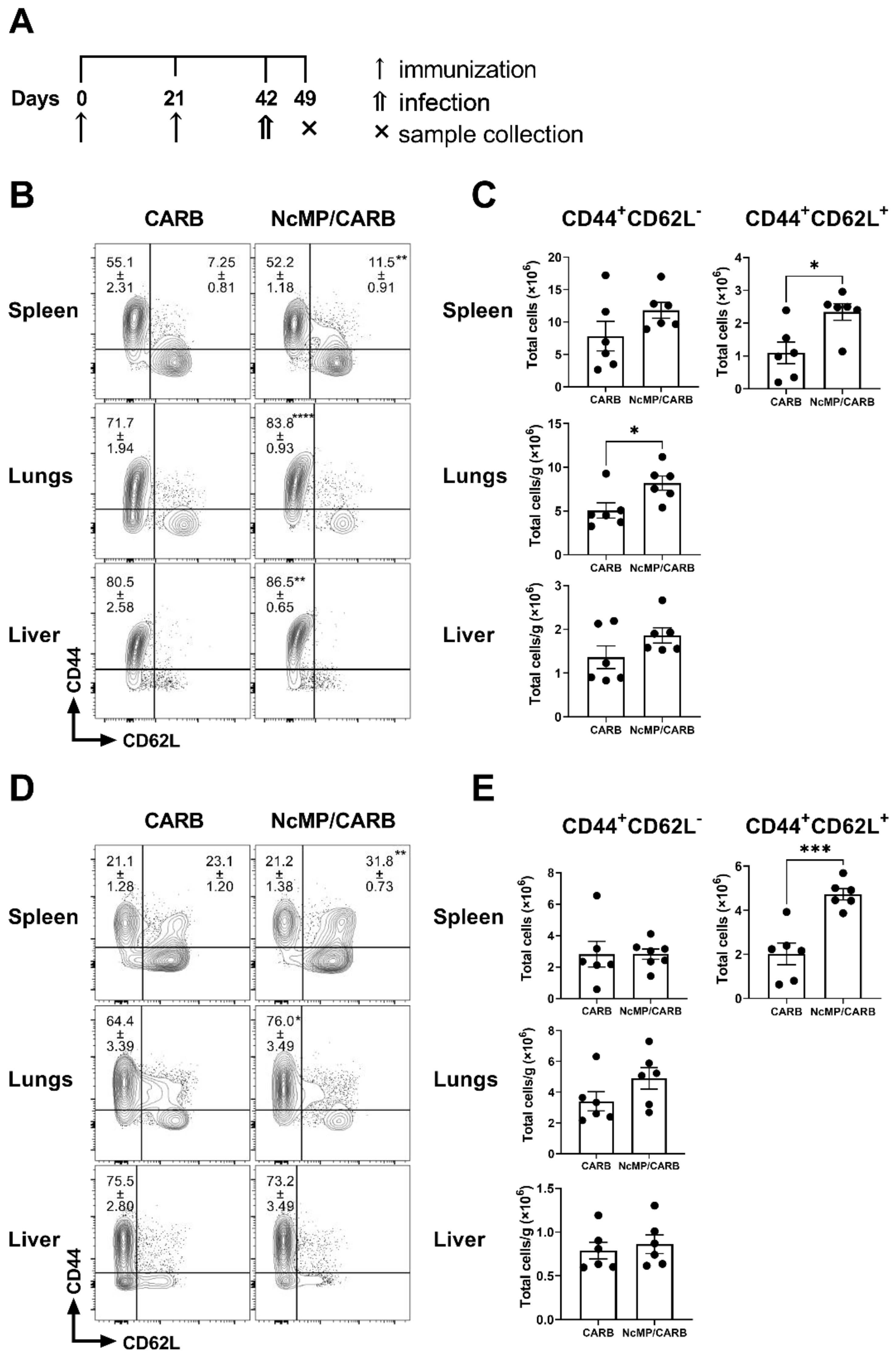
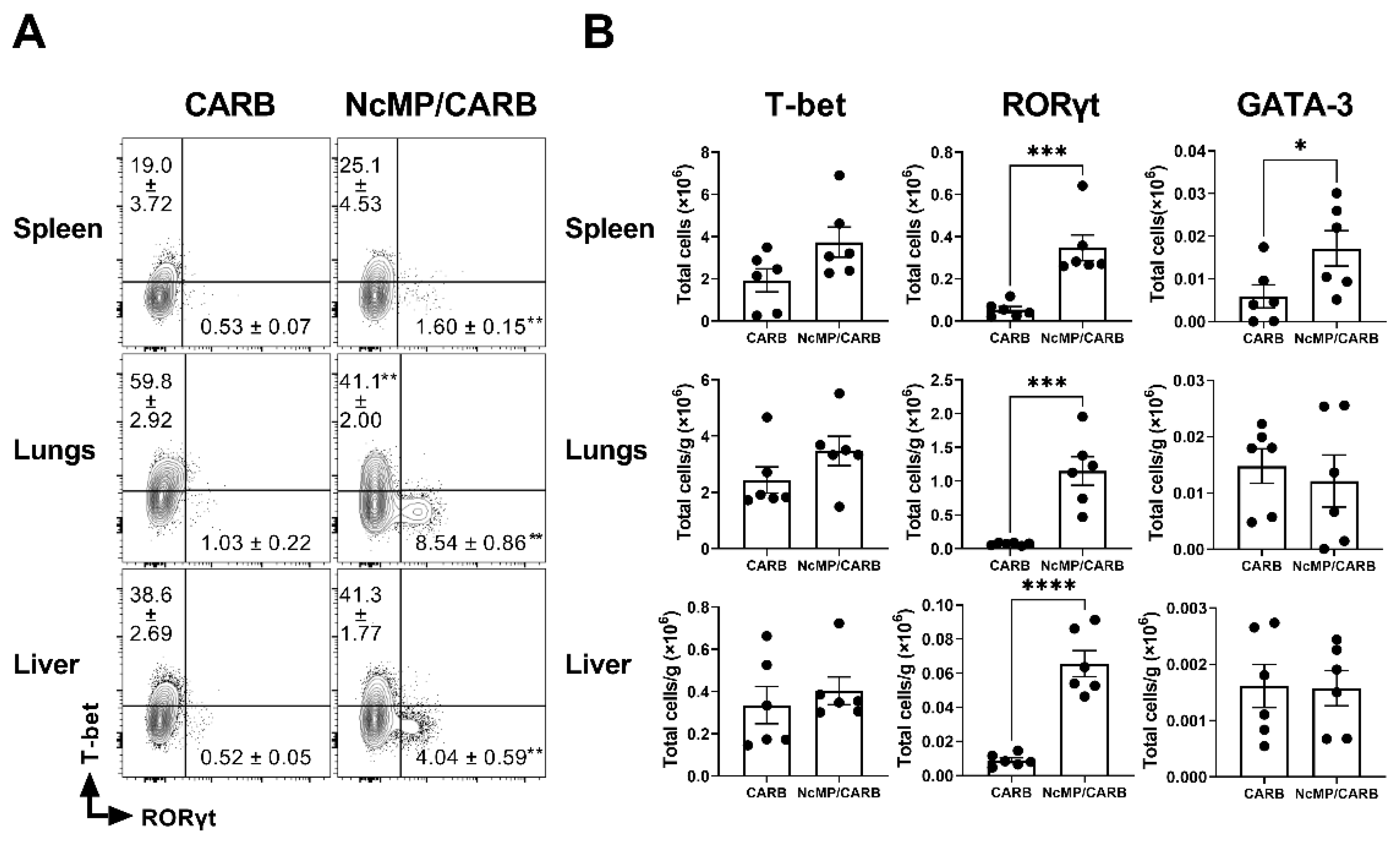
3.4. Differentiation of T cells Induced in Immunized Mice Infected with N. caninum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.; Schares, G.R.M. Neosporosis in animals—The last five years. Vet. Parasitol. 2011, 180, 90–108. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, D.S.; Dubey, J. Neosporosis, Toxoplasmosis, and Sarcocystosis in Ruminants: An Update. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, P.; Shaw, A.P.; Muñoz, V.; Bruce, M.; Torgerson, P.R. Estimating the burden of multiple endemic diseases and health conditions using Bayes’ Theorem: A conditional probability model applied to UK dairy cattle. Prev. Vet. Med. 2022, 203, 105617. [Google Scholar] [CrossRef] [PubMed]
- Demir, P.A.; Eşki, F.; ÜTÜK, A.E. Estimating the total economic costs of Neospora caninum infections in dairy cows in Turkey. Trop. Anim. Health Prod. 2020, 52, 3251–3258. [Google Scholar] [CrossRef] [PubMed]
- Reichel, M.P.; Ayanegui-Alcérreca, M.A.; Gondim, L.; Ellis, J. What is the global economic impact of Neospora caninum in cattle—The billion dollar question. Int. J. Parasitol. 2013, 43, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, C.M.; Soares, I.R.; Mendes, R.G.; Bastos, P.A.D.S.; Katagiri, S.; Zavilenski, R.B.; de Abreu, H.F.P.; Afreixo, V. Meta-analysis of the prevalence and risk factors associated with bovine neosporosis. Trop. Anim. Health Prod. 2019, 51, 1783–1800. [Google Scholar] [CrossRef]
- Rojas-Pirela, M.; Medina, L.; Rojas, M.V.; Liempi, A.I.; Castillo, C.; Pérez-Pérez, E.; Guerrero-Muñoz, J.; Araneda, S.; Kemmerling, U. Congenital Transmission of Apicomplexan Parasites: A Review. Front. Microbiol. 2021, 12, 751648. [Google Scholar] [CrossRef]
- Dubey, J.P.; Schares, G.; Ortega-Mora, L.M. Epidemiology and Control of Neosporosis and Neospora caninum. Clin. Microbiol. Rev. 2007, 20, 323–367. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, R.S.; Vazquez, P.; Ferre, I.; Ortega-Mora, L.-M. Treatment of Toxoplasmosis and Neosporosis in Farm Ruminants: State of Knowledge and Future Trends. Curr. Top. Med. Chem. 2018, 18, 1304–1323. [Google Scholar] [CrossRef]
- Reichel, M.P.; McAllister, M.M.; Pomroy, W.E.; Campero, C.; Ortega-Mora, L.-M.; Ellis, J.T. Control options for Neospora caninum—Is there anything new or are we going backwards? Parasitology 2014, 141, 1455–1470. [Google Scholar] [CrossRef]
- Liu, Y.; Reichel, M.P.; Lo, W.-C. Combined control evaluation for Neospora caninum infection in dairy: Economic point of view coupled with population dynamics. Vet. Parasitol. 2020, 277, 108967. [Google Scholar] [CrossRef] [PubMed]
- Monney, T.; Hemphill, A. Vaccines against neosporosis: What can we learn from the past studies? Exp. Parasitol. 2014, 140, 52–70. [Google Scholar] [CrossRef] [PubMed]
- Fereig, R.M.; Nishikawa, Y. From Signaling Pathways to Distinct Immune Responses: Key Factors for Establishing or Combating Neospora caninum Infection in Different Susceptible Hosts. Pathogens 2020, 9, 384. [Google Scholar] [CrossRef] [PubMed]
- Almería, S.; Serrano-Pérez, B.; López-Gatius, F. Immune response in bovine neosporosis: Protection or contribution to the pathogenesis of abortion. Microb. Pathog. 2017, 109, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Hamada, T.; Inoue, N.; Nagasawa, H.; Fujisaki, K.; Suzuki, N.; Mikami, T. The role of CD4+ or CD8+ T cells in the protective immune response of BALB/c mice to Neospora caninum infection. Vet. Parasitol. 2000, 90, 183–191. [Google Scholar] [CrossRef]
- Correia, A.; Ferreirinha, P.; Botelho, S.; Belinha, A.; Leitão, C.; Caramalho, Í.; Teixeira, L.; González-Fernandéz, Á.; Appelberg, R.; Vilanova, M. Predominant role of interferon-γ in the host protective effect of CD8+ T cells against Neospora caninum infection. Sci. Rep. 2015, 5, 14913. [Google Scholar] [CrossRef]
- Yamane, I.; Kitani, H.; Kokuho, T.; Shibahara, T.; Haritani, M.; Hamaoka, T.; Shimizu, S.; Koiwai, M.; Shimura, K.; Yokomizo, Y. The inhibitory Effect of Interferon Gamma and Tumor Necrosis Factor Alpha on Intracellular Multiplication of Neospora caninum in Primary Bovine Brain Cells. J. Vet. Med. Sci. 2000, 62, 347–351. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Nagasawa, H.; Fujisaki, K.; Suzuki, N.; Mikami, T. Growth-inhibitory effects of interferon-γ on Neospora caninum in murine macrophages by a nitric oxide mechanism. Parasitol. Res. 2000, 86, 768–771. [Google Scholar] [CrossRef]
- Nishikawa, Y.; Tragoolpua, K.; Inoue, N.; Makala, L.; Nagasawa, H.; Otsuka, H.; Mikami, T. In the Absence of Endogenous Gamma Interferon, Mice Acutely Infected with Neospora caninum Succumb to a Lethal Immune Response Characterized by Inactivation of Peritoneal Macrophages. Clin. Diagn. Lab. Immunol. 2001, 8, 811–816. [Google Scholar] [CrossRef] [Green Version]
- Barros, P.D.S.C.; Mota, C.M.; Miranda, V.D.S.; Ferreira, F.B.; Ramos, E.L.P.; Santana, S.S.; Costa, L.F.; Pajuaba, A.C.A.M.; Mineo, J.R.; Mineo, T.W.P. Inducible Nitric Oxide Synthase is required for parasite restriction and inflammatory modulation during Neospora caninum infection. Vet. Parasitol. 2019, 276, 108990. [Google Scholar] [CrossRef]
- Spekker, K.; Leineweber, M.; Degrandi, D.; Ince, V.; Brunder, S.; Schmidt, S.K.; Stuhlsatz, S.; Howard, J.C.; Schares, G.; Degistirici, O.; et al. Antimicrobial effects of murine mesenchymal stromal cells directed against Toxoplasma gondii and Neospora caninum: Role of immunity-related GTPases (IRGs) and guanylate-binding proteins (GBPs). Med. Microbiol. Immunol. 2012, 202, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.J.; Vermont, S.J.; Cotton, J.A.; Harris, D.; Hill-Cawthorne, G.A.; Könen-Waisman, S.; Latham, S.M.; Mourier, T.; Norton, R.; Quail, M.A.; et al. Comparative Genomics of the Apicomplexan Parasites Toxoplasma gondii and Neospora caninum: Coccidia Differing in Host Range and Transmission Strategy. PLoS Pathog. 2012, 8, e1002567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Expósito, D.; González-Warleta, M.; Espinosa, J.; Vallejo-García, R.; Castro-Hermida, J.A.; Calvo, C.; Ferreras, M.C.; Pérez, V.; Benavides, J.; Mezo, M. Maternal immune response in the placenta of sheep during recrudescence of natural congenital infection of Neospora caninum. Vet. Parasitol. 2020, 285, 109204. [Google Scholar] [CrossRef] [PubMed]
- Peckham, R.K.; Brill, R.; Foster, D.; Bowen, A.L.; Leigh, J.; Coffey, T.; Flynn, R.J. Two distinct populations of Bovine IL-17+ T-cells can be induced and WC1+IL-17+γδ T-cells are effective killers of protozoan parasites. Sci. Rep. 2014, 4, 5431. [Google Scholar] [CrossRef] [Green Version]
- Debache, K.; Hemphill, A. Differential effects of intranasal vaccination with recombinant NcPDI in different mouse models of Neospora caninum infection. Parasite Immunol. 2012, 35, 11–20. [Google Scholar] [CrossRef]
- Flynn, R.J.; Marshall, E.S. Parasite limiting macrophages promote IL-17 secretion in naive bovine CD4+ T-cells during Neospora caninum infection. Vet. Immunol. Immunopathol. 2011, 144, 423–429. [Google Scholar] [CrossRef]
- Uchida, M.; Nagashima, K.; Akatsuka, Y.; Murakami, T.; Ito, A.; Imai, S.; Ike, K. Comparative study of protective activities of Neospora caninum bradyzoite antigens, NcBAG1, NcBSR4, NcMAG1, and NcSAG4, in a mouse model of acute parasitic infection. Parasitol. Res. 2012, 112, 655–663. [Google Scholar] [CrossRef]
- Mansilla, F.C.; Franco-Mahecha, O.L.; Lavoria, M.; Moore, D.; Giraldez, A.N.; Iglesias, M.E.; Wilda, M.; Capozzo, A.V. The immune enhancement of a novel soy lecithin/β-glucans based adjuvant on native Neospora caninum tachyzoite extract vaccine in mice. Vaccine 2012, 30, 1124–1131. [Google Scholar] [CrossRef]
- Monney, T.; Rütti, D.; Schorer, M.; Debache, K.; Grandgirard, D.; Leib, S.L.; Hemphill, A. RecNcMIC3-1-R is a microneme- and rhoptry-based chimeric antigen that protects against acute neosporosis and limits cerebral parasite load in the mouse model for Neospora caninum infection. Vaccine 2011, 29, 6967–6975. [Google Scholar] [CrossRef]
- Williams, D.J.L.; Guy, C.S.; Smith, R.F.; Ellis, J.; Björkman, C.; Reichel, M.P.; Trees, A.J. Immunization of Cattle with Live Tachyzoites of Neospora caninum Confers Protection against Fetal Death. Infect. Immun. 2007, 75, 1343–1348. [Google Scholar] [CrossRef] [Green Version]
- Staska, L.M.; Davies, C.J.; Brown, W.C.; McGuire, T.C.; Suarez, C.E.; Park, J.Y.; Mathison, B.A.; Abbott, J.R.; Baszler, T.V. Identification of Vaccine Candidate Peptides in the NcSRS2 Surface Protein of Neospora caninum by Using CD4 + Cytotoxic T Lymphocytes and Gamma Interferon-Secreting T Lymphocytes of Infected Holstein Cattle. Infect. Immun. 2005, 73, 1321–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrianarivo, A.; Choromanski, L.; McDonough, S.; Packham, A.; Conrad, P. Immunogenicity of a killed whole Neospora caninum tachyzoite preparation formulated with different adjuvants. Int. J. Parasitol. 1999, 29, 1613–1625. [Google Scholar] [CrossRef]
- Ferreirinha, P.; Dias, J.; Correia, A.; Pérez-Cabezas, B.; Santos, C.; Teixeira, L.; Ribeiro, A.; Rocha, A.; Vilanova, M. Protective effect of intranasal immunization with Neospora caninum membrane antigens against murine neosporosis established through the gastrointestinal tract. Immunology 2014, 141, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Ferreirinha, P.; Correia, A.; Teixeira-Coelho, M.; Osório, H.; Teixeira, L.; Rocha, A.; Vilanova, M. Mucosal immunization confers long-term protection against intragastrically established Neospora caninum infection. Vaccine 2016, 34, 6250–6258. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-E.; Lee, S.-Y.; Kim, R.-H.; Ko, M.-K.; Lee, K.-N.; Kim, S.-M.; Kim, B.-K.; Lee, J.-S.; Kim, B.; Park, J.-H. Enhanced immune responses of foot-and-mouth disease vaccine using new oil/gel adjuvant mixtures in pigs and goats. Vaccine 2014, 32, 5221–5227. [Google Scholar] [CrossRef]
- Peters, J.L.; DeMars, P.L.; Collins, L.M.; Stoner, J.A.; Matsumoto, H.; Komori, N.; Singh, A.; Feasley, C.L.; Haddock, J.A.; Levine, M. Effects of immunization with natural and recombinant lysine decarboxylase on canine gingivitis development. Vaccine 2012, 30, 6706–6712. [Google Scholar] [CrossRef]
- Teixeira, L.; Botelho, A.S.; Mesquita, S.D.; Correia, A.; Cerca, F.; Costa, R.; Sampaio, P.; Castro, G.; Vilanova, M. Plasmacytoid and conventional dendritic cells are early producers of IL-12 in Neospora caninum -infected mice. Immunol. Cell Biol. 2009, 88, 79–86. [Google Scholar] [CrossRef]
- Nishikawa, Y.; Inoue, N.; Xuan, X.; Nagasawa, H.; Igarashi, I.; Fujisaki, K.; Otsuka, H.; Mikami, T. Protective efficacy of vaccination by recombinant vaccinia virus against Neospora caninum infection. Vaccine 2001, 19, 1381–1390. [Google Scholar] [CrossRef]
- Williams, M.A.; Bevan, M.J. Effector and Memory CTL Differentiation. Annu. Rev. Immunol. 2007, 25, 171–192. [Google Scholar] [CrossRef]
- Raphael, I.; Nalawade, S.; Eagar, T.N.; Forsthuber, T.G. T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine 2014, 74, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Shirota, H.; Klinman, D.M. Recent progress concerning CpG DNA and its use as a vaccine adjuvant. Expert Rev. Vaccines 2013, 13, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Botelho, A.; Teixeira, L.; Correia-Da-Costa, J.; Faustino, A.; Castro, A.; Vilanova, M. Neospora caninum: High susceptibility to the parasite in C57BL/10ScCr mice. Exp. Parasitol. 2007, 115, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Mineo, T.W.; Benevides, L.; Silva, N.M.; Silva, J.S. Myeloid differentiation factor 88 is required for resistance to Neospora caninum infection. Vet. Res. 2009, 40, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallies, A.; Good-Jacobson, K.L. Transcription Factor T-bet Orchestrates Lineage Development and Function in the Immune System. Trends Immunol. 2017, 38, 287–297. [Google Scholar] [CrossRef]
- Correia, A.; Ferreirinha, P.; Costa, A.A.; Dias, J.; Melo, J.; Costa, R.; Ribeiro, A.; Faustino, A.; Teixeira, L.; Rocha, A.; et al. Mucosal and systemic T cell response in mice intragastrically infected with Neospora caninum tachyzoites. Vet. Res. 2013, 44, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, C.; Oestreich, K.J.; Paley, M.; Crawford, A.; Angelosanto, J.M.; Ali, M.-A.A.; Intlekofer, A.; Boss, J.M.; Reiner, S.L.; Weinmann, A.S.; et al. Transcription factor T-bet represses expression of the inhibitory receptor PD-1 and sustains virus-specific CD8+ T cell responses during chronic infection. Nat. Immunol. 2011, 12, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Valeri, M.; Raffatellu, M. Cytokines IL-17 and IL-22 in the host response to infection. Pathog. Dis. 2016, 74, ftw111. [Google Scholar] [CrossRef] [Green Version]
- Vesely, M.C.A.; Rodríguez, C.; Gruppi, A.; Rodríguez, E.V.A. Interleukin-17 mediated immunity during infections with Trypanosoma cruzi and other protozoans. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165706. [Google Scholar] [CrossRef]
- Kelly, M.N.; Kolls, J.K.; Happel, K.; Schwartzman, J.D.; Schwarzenberger, P.; Combe, C.; Moretto, M.; Khan, I.A. Interleukin-17/Interleukin-17 Receptor-Mediated Signaling Is Important for Generation of an Optimal Polymorphonuclear Response against Toxoplasma gondii Infection. Infect. Immun. 2005, 73, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, P.; Ahmadpour, E.; Daryani, A.; Kazemi, T.; Spotin, A.; Mahami-Oskouei, M.; Flynn, R.J.; Azadi, Y.; Rajabi, S.; Sandoghchian, S. IL-17 and IL-22 elicited by a DNA vaccine encoding ROP13 associated with protection against Toxoplasma gondii in BALB/c mice. J. Cell. Physiol. 2018, 234, 10782–10788. [Google Scholar] [CrossRef]
- Rathore, J.S.; Wang, Y. Protective role of Th17 cells in pulmonary infection. Vaccine 2016, 34, 1504–1514. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Bhattacharya, P.; Dagur, P.K.; Karmakar, S.; Ismail, N.; Joshi, A.B.; Akue, A.D.; KuKuruga, M.; McCoy, J.P.; Dey, R.; et al. Live Attenuated Leishmania donovani Centrin Gene–Deleted Parasites Induce IL-23–Dependent IL-17–Protective Immune Response against Visceral Leishmaniasis in a Murine Model. J. Immunol. 2017, 200, 163–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, Y.; Hamano, S.; Wang, S.; Shimanoe, Y.; Iwakura, Y.; Yoshida, H. IL-17 Is Necessary for Host Protection against Acute-Phase Trypanosoma cruzi Infection. J. Immunol. 2010, 185, 1150–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, A.C.; Wilk, M.; Misiak, A.; Borkner, L.; Murphy, D.; Mills, K.H.G. Sustained protective immunity against Bordetella pertussis nasal colonization by intranasal immunization with a vaccine-adjuvant combination that induces IL-17-secreting TRM cells. Mucosal Immunol. 2018, 11, 1763–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen-Kilic, E.; Blackwood, C.; Boehm, D.T.; Witt, W.T.; Malkowski, A.C.; Bevere, J.R.; Wong, T.Y.; Hall, J.M.; Bradford, S.; Varney, M.; et al. Intranasal Peptide-Based FpvA-KLH Conjugate Vaccine Protects Mice From Pseudomonas aeruginosa Acute Murine Pneumonia. Front. Immunol. 2019, 10, 2497. [Google Scholar] [CrossRef] [PubMed]
- Cabral, M.P.; Correia, A.; Vilanova, M.; Gärtner, M.D.F.; Moscoso, M.; García, P.; Vallejo, J.A.; Pérez, A.; Francisco-Tomé, M.; Fuentes-Valverde, V.; et al. A live auxotrophic vaccine confers mucosal immunity and protection against lethal pneumonia caused by Pseudomonas aeruginosa. PLoS Pathog. 2020, 16, e1008311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashtekar, A.R.; Katz, J.; Xu, Q.; Michalek, S.M. A Mucosal Subunit Vaccine Protects against Lethal Respiratory Infection with Francisella tularensis LVS. PLoS ONE 2012, 7, e50460. [Google Scholar] [CrossRef] [PubMed]
- Moroda, M.; Takamoto, M.; Iwakura, Y.; Nakayama, J.; Aosai, F. Interleukin-17A-Deficient Mice Are Highly Susceptible to Toxoplasma gondii Infection Due to Excessively Induced T. gondii HSP70 and Interferon Gamma Production. Infect. Immun. 2017, 85, e00399-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreirinha, P.; Fróis-Martins, R.; Teixeira, L.; Rocha, A.; Vilanova, M.; Correia, A. Interferon-γ-dependent protection against Neospora caninum infection conferred by mucosal immunization in IL-12/IL-23 p40-deficient mice. Vaccine 2018, 36, 4890–4896. [Google Scholar] [CrossRef]
- Pereyra, R.; Mansilla, F.C.; Petersen, M.I.; Suarez, V.; Capozzo, A.V. Evidence of reduced vertical transmission of Neospora caninum associated with higher IgG1 than IgG2 serum levels and presence of IFN-γ in non-aborting chronically infected cattle under natural condition. Vet. Immunol. Immunopathol. 2019, 208, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Aguado-Martínez, A.; Basto, A.; Leitao, A.; Hemphill, A. Neospora caninum in non-pregnant and pregnant mouse models: Cross-talk between infection and immunity. Int. J. Parasitol. 2017, 47, 723–735. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, A.; Alves, P.; Fróis-Martins, R.; Teixeira, L.; Vilanova, M. Protective Effect against Neosporosis Induced by Intranasal Immunization with Neospora caninum Membrane Antigens Plus Carbomer-Based Adjuvant. Vaccines 2022, 10, 925. https://doi.org/10.3390/vaccines10060925
Correia A, Alves P, Fróis-Martins R, Teixeira L, Vilanova M. Protective Effect against Neosporosis Induced by Intranasal Immunization with Neospora caninum Membrane Antigens Plus Carbomer-Based Adjuvant. Vaccines. 2022; 10(6):925. https://doi.org/10.3390/vaccines10060925
Chicago/Turabian StyleCorreia, Alexandra, Pedro Alves, Ricardo Fróis-Martins, Luzia Teixeira, and Manuel Vilanova. 2022. "Protective Effect against Neosporosis Induced by Intranasal Immunization with Neospora caninum Membrane Antigens Plus Carbomer-Based Adjuvant" Vaccines 10, no. 6: 925. https://doi.org/10.3390/vaccines10060925