
Moving beyond Titers

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
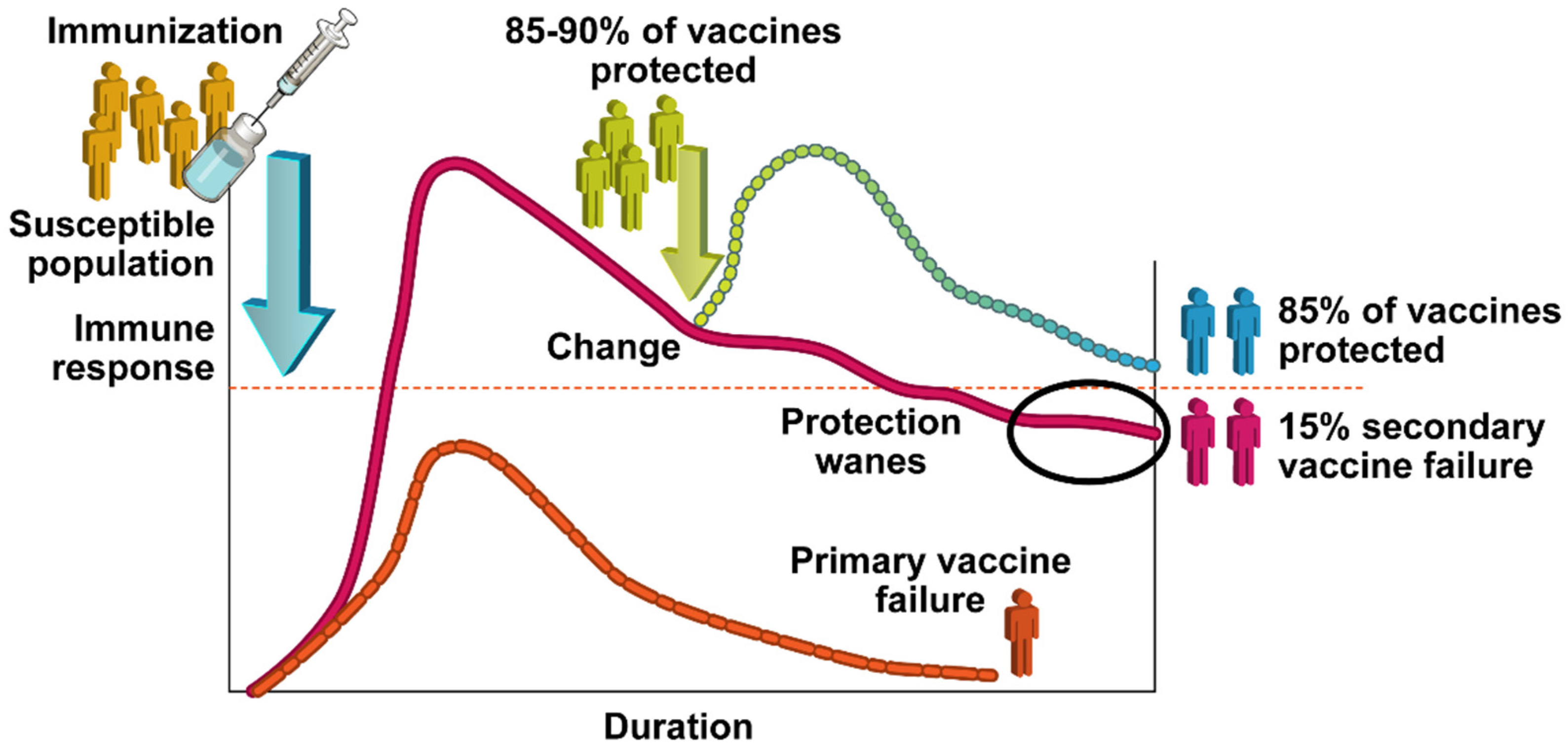
2. Vaccine Effectiveness
3. New and Emerging Technologies
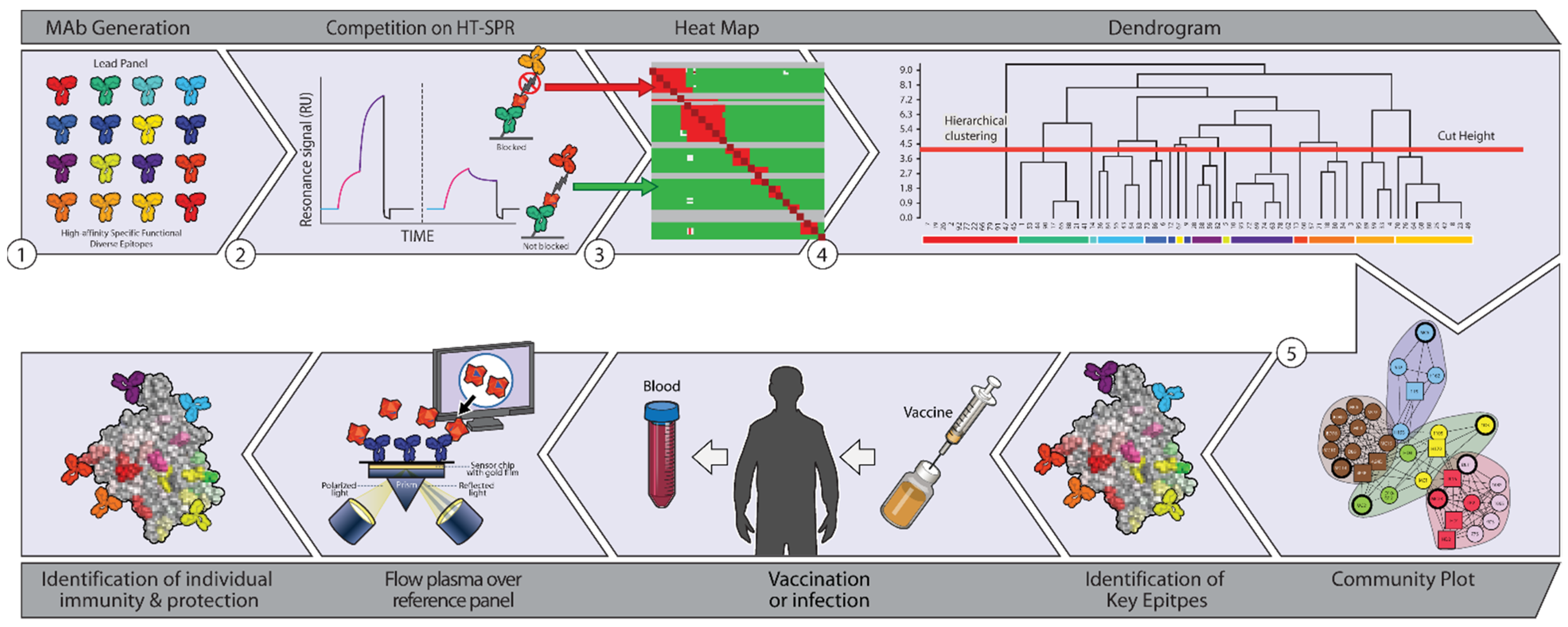
3.1. Epitope Characterization
3.2. Immunogenomics and HLA Typing
3.3. Antibody Repertoire Sequencing
4. Advancement Opportunities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robbins, M.J.; Jacobson, S.H. Analytics for Vaccine Economics and Pricing: Insights and Observations. Expert Rev. Vaccines 2015, 14, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Carias, C.; Reed, C.; Kim, I.K.; Foppa, I.M.; Biggerstaff, M.; Meltzer, M.I.; Finelli, L.; Swerdlow, D.L. Net Costs Due to Seasonal Influenza Vaccination—United States, 2005–2009. PLoS ONE 2015, 10, e0132922. [Google Scholar] [CrossRef] [PubMed]
- Stratton, S.J. COVID-19: Not a Simple Public Health Emergency. Prehospital Disaster Med. 2020, 35, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, G.E.; Elvidge, J.; Davies, L.M. A Systematic Review of Economic Evaluations of Seasonal Influenza Vaccination for the Elderly Population in the European Union. BMJ Open 2017, 7, e014847. [Google Scholar] [CrossRef]
- Wong, C.K.H.; Liao, Q.; Guo, V.Y.W.; Xin, Y.; Lam, C.L.K. Cost-Effectiveness Analysis of Vaccinations and Decision Makings on Vaccination Programmes in Hong Kong: A Systematic Review. Vaccine 2017, 35, 3153–3161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, D.E. Measles Vaccine. Viral Immunol. 2018, 31, 86–95. [Google Scholar] [CrossRef]
- Stanaway, J.D.; Afshin, A.; Gakidou, E.; Lim, S.S.; Abate, D.; Abate, K.H.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; et al. Global, Regional, and National Comparative Risk Assessment of 84 Behavioural, Environmental and Occupational, and Metabolic Risks or Clusters of Risks for 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1923–1994. [Google Scholar] [CrossRef] [Green Version]
- Sheerin, D.; Openshaw, P.J.; Pollard, A.J. Issues in Vaccinology: Present Challenges and Future Directions. Eur. J. Immunol. 2017, 47, 2017–2025. [Google Scholar] [CrossRef] [Green Version]
- Pronker, E.S.; Weenen, T.C.; Commandeur, H.; Claassen, E.H.J.H.M.; Osterhaus, A.D.M.E. Risk in Vaccine Research and Development Quantified. PLoS ONE 2013, 8, e57755. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, S.A. Correlates of Protection. In Vaccines; Elsevier: Amsterdam, The Netherlands, 2018; Volume 11, pp. 35–40.e4. [Google Scholar]
- Plotkin, S.; Robinson, J.M.; Cunningham, G.; Iqbal, R.; Larsen, S. The Complexity and Cost of Vaccine Manufacturing—An Overview. Vaccine 2017, 35, 4064–4071. [Google Scholar] [CrossRef]
- Offit, P.A. Why Are Pharmaceutical Companies Gradually Abandoning Vaccines? Health Aff. (Millwood) 2005, 24, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Gouglas, D.; Le, T.T.; Henderson, K.; Kaloudis, A.; Danielsen, T.; Hammersland, N.C.; Robinson, J.M.; Heaton, P.M.; Røttingen, J.-A. Estimating the Cost of Vaccine Development against Epidemic Infectious Diseases: A Cost Minimisation Study. Lancet Glob. Health 2018, 6, e1386–e1396. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, S.A. Correlates of Protection Induced by Vaccination. Clin. Vaccine Immunol. CVI 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegrist, C.A. Vaccine Immunology. In Plotkin’s Vaccines; Elsevier: Amsterdam, The Netherlands, 2018; pp. 16–34.e7. [Google Scholar]
- Fenwick, C.; Turelli, P.; Pellaton, C.; Farina, A.; Campos, J.; Raclot, C.; Pojer, F.; Cagno, V.; Nusslé, S.G.; D’Acremont, V.; et al. A High-Throughput Cell- and Virus-Free Assay Shows Reduced Neutralization of SARS-CoV-2 Variants by COVID-19 Convalescent Plasma. Sci. Transl. Med. 2021, 13, eabi8452. [Google Scholar] [CrossRef]
- Plotkin, S.A.; Plotkin, S.A. Correlates of Vaccine-Induced Immunity. Clin. Infect. Dis. 2008, 47, 401–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, T.M.; Ditto, N.T.; Lou, H.; Brooks, B.D.; Atanasiu, D.; Eisenberg, R.J.; Cohen, G.H. Global Sensing of the Antigenic Structure of Herpes Simplex Virus GD Using High-Throughput Array-Based SPR Imaging. PLoS Pathog. 2017, 13, e1006430. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, R.B.; Ovsyannikova, I.G.; Thomas, A.; Larrabee, B.R.; Rubin, S.; Poland, G.A. Differential Durability of Immune Responses to Measles and Mumps Following MMR Vaccination. Vaccine 2019, 37, 1775–1784. [Google Scholar] [CrossRef]
- Bresee, J.S.; Fry, A.M.; Sambhara, S.; Cox, N.J. Inactivated Influenza Vaccines. In Plotkin’s Vaccines; Elsevier: Amsterdam, The Netherlands, 2018; pp. 456–488.e21. [Google Scholar]
- Luke, C.J.; Lakdawala, S.S.; Subbarao, K. Influenza Vaccine. In Plotkin’s Vaccines; Elsevier: Amsterdam, The Netherlands, 2012; pp. 489–510.e7. [Google Scholar]
- Wallis, J.; Shenton, D.P.; Carlisle, R.C. Novel Approaches for the Design, Delivery and Administration of Vaccine Technologies. Clin. Exp. Immunol. 2019, 196, 189–204. [Google Scholar] [CrossRef] [Green Version]
- Abdiche, Y.N.; Yeung, A.Y.; Ni, I.; Stone, D.; Miles, A.; Morishige, W.; Rossi, A.; Strop, P. Antibodies Targeting Closely Adjacent or Minimally Overlapping Epitopes Can Displace One Another. PLoS ONE 2017, 12, e0169535. [Google Scholar] [CrossRef]
- Oscherwitz, J. The Promise and Challenge of Epitope-Focused Vaccines. Hum. Vaccines Immunother. 2016, 12, 2113–2116. [Google Scholar] [CrossRef] [Green Version]
- Abdiche, Y. Accelerating the Discovery of Therapeutic Antibodies Using High Throughput SPR. Available online: https://www.genengnews.com/resources/webinars/accelerating-the-discovery-of-therapeutic-antibodies-using-high-throughput-spr/ (accessed on 4 March 2022).
- Hook, L.M.; Cairns, T.M.; Awasthi, S.; Brooks, B.D.; Ditto, N.T.; Eisenberg, R.J.; Cohen, G.H.; Friedman, H.M. Vaccine-Induced Antibodies to Herpes Simplex Virus Glycoprotein D Epitopes Involved in Virus Entry and Cell-to-Cell Spread Correlate with Protection against Genital Disease in Guinea Pigs. PLoS Pathog. 2018, 14, e1007095. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Hook, L.; Swaminathan, G.; Cairns, T.; Brooks, B.; Smith, J.; Ditto, N.; Gindy, M.; Bett, A.; Espeseth, A.; et al. Antibody Responses to Crucial Functional Epitopes as a Novel Approach to Assess Immunogenicity of Vaccine Adjuvants. Vaccine 2019, 37, 3770–3778. [Google Scholar] [CrossRef] [PubMed]
- Cairns, T.M.; Ditto, N.T.; Atanasiu, D.; Lou, H.; Brooks, B.D.; Saw, W.T.; Eisenberg, R.J.; Cohen, G.H. Surface Plasmon Resonance Reveals Direct Binding of Herpes Simplex Virus Glycoproteins GH/GL to GD and Locates a GH/GL Binding Site on GD. J. Virol. 2019, 93, e00289-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivasubramanian, A.; Estep, P.; Lynaugh, H.; Yu, Y.; Miles, A.; Eckman, J.; Schutz, K.; Piffath, C.; Boland, N.; Niles, R.H.; et al. Broad Epitope Coverage of a Human in Vitro Antibody Library. mAbs 2016, 9, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Abdiche, Y.N.; Miles, A.; Eckman, J.; Foletti, D.; Van Blarcom, T.J.; Yeung, Y.A.; Pons, J.; Rajpal, A. High-Throughput Epitope Binning Assays on Label-Free Array-Based Biosensors Can Yield Exquisite Epitope Discrimination That Facilitates the Selection of Monoclonal Antibodies with Functional Activity. PLoS ONE 2014, 9, e92451. [Google Scholar] [CrossRef]
- Ohmit, S.E.; Petrie, J.G.; Cross, R.T.; Johnson, E.; Monto, A.S. Influenza Hemagglutination-Inhibition Antibody Titer as a Correlate of Vaccine-Induced Protection. J. Infect. Dis. 2011, 204, 1879–1885. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.T.; Markowitz, L.E.; Albrecht, P.; Stewart, J.A.; Mofenson, L.M.; Preblud, S.R.; Orenstein, W.A. Measles Antibody: Reevaluation of Protective Titers. J. Infect. Dis. 1990, 162, 1036–1042. [Google Scholar] [CrossRef]
- To, K.K.W.; Zhang, A.J.X.; Hung, I.F.N.; Xu, T.; Ip, W.C.T.; Wong, R.T.Y.; Ng, J.C.K.; Chan, J.F.W.; Chan, K.-H.; Yuen, K.-Y. High Titer and Avidity of Nonneutralizing Antibodies against Influenza Vaccine Antigen Are Associated with Severe Influenza. Clin. Vaccine Immunol. CVI 2012, 19, 1012–1018. [Google Scholar] [CrossRef] [Green Version]
- Edinger, A.L.; Ahuja, M.; Sung, T.; Baxter, K.C.; Haggarty, B.; Doms, R.W.; Hoxie, J.A. Characterization and Epitope Mapping of Neutralizing Monoclonal Antibodies Produced by Immunization with Oligomeric Simian Immunodeficiency Virus Envelope Protein. J. Virol. 2000, 74, 7922–7935. [Google Scholar] [CrossRef] [Green Version]
- Depla, E.; Van der Aa, A.; Livingston, B.D.; Crimi, C.; Allosery, K.; Brabandere, V.D.; Krakover, J.; Murthy, S.; Huang, M.; Power, S.; et al. Rational Design of a Multiepitope Vaccine Encoding T-Lymphocyte Epitopes for Treatment of Chronic Hepatitis B Virus Infections. J. Virol. 2008, 82, 435–450. [Google Scholar] [CrossRef] [Green Version]
- Poland, G.A.; Ovsyannikova, I.G.; Kennedy, R.B.; Haralambieva, I.H.; Jacobson, R.M. Vaccinomics and a New Paradigm for the Development of Preventive Vaccines against Viral Infections. Omics J. Integr. Biol. 2011, 15, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Ovsyannikova, I.G.; Poland, G.A. Vaccinomics: Current Findings, Challenges and Novel Approaches for Vaccine Development. AAPS J. 2011, 13, 438–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, L.; Calvo-Calle, J. HLA-DR: Molecular Insights and Vaccine Design. Curren Pharm. Des. 2009, 15, 3249–3261. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.; David, J.K.; Maden, S.K.; Wood, M.A.; Weeder, B.R.; Nellore, A.; Thompson, R.F. Human Leukocyte Antigen Susceptibility Map for Severe Acute Respiratory Syndrome Coronavirus 2. J. Virol. 2020, 94, e00510–e00520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, A.; Gaughran, F.; Karasu, A.; Gilbert, A.; Mann, A.; Gelder, C.; Oxford, J.; Stephens, H.; Lambkin-Williams, R. Correlation between Human Leukocyte Antigen Class II Alleles and HAI Titers Detected Post-Influenza Vaccination. Public Libr. Sci. 2013, 8, e71376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyarzún, P.; Ellis, J.; Bodén, M.; Kobe, B. PREDIVAC: CD4+ T-Cell Epitope Prediction for Vaccine Design That Covers 95% of HLA Class II DR Protein Diversity. BioMed Cent. 2013, 14, 52. [Google Scholar] [CrossRef]
- Bashford-Rogers, R.J.M.; Palser, A.L.; Idris, S.F.; Carter, L.; Epstein, M.; Callard, R.E.; Douek, D.C.; Vassiliou, G.S.; Follows, G.A.; Hubank, M.; et al. Capturing Needles in Haystacks: A Comparison of B-Cell Receptor Sequencing Methods. BMC Immunol. 2014, 15, 29. [Google Scholar] [CrossRef] [Green Version]
- Robinson, W.H. Sequencing the Functional Antibody Repertoire--Diagnostic and Therapeutic Discovery. Nat. Rev. Rheumatol. 2015, 11, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Slatko, B.E.; Gardner, A.F.; Ausubel, F.M. Overview of Next Generation Sequencing Technologies. Curr. Protoc. Mol. Biol. 2018, 122, e59. [Google Scholar] [CrossRef]
- Rees, A.R. Understanding the Human Antibody Repertoire. mAbs 2020, 12, 1729683. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, K.; Raybould, M.I.J.; Kovaltsuk, A.; Deane, C.M. Looking for Therapeutic Antibodies in Next-Generation Sequencing Repositories. mAbs 2019, 11, 1197–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finlay, W.J.J.; Lugovskoy, A.A. De novo discovery of antibody drugs–great promise demands scrutiny. mAbs 2019, 11, 809–811. [Google Scholar] [CrossRef] [PubMed]
- Yaari, G.; Kleinstein, S.H. Practical Guidelines for B-Cell Receptor Repertoire Sequencing Analysis. Genome Med. 2015, 7, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Sok, D.; Azadnia, P.; Hsueh, J.; Landais, E.; Simek, M.; Koff, W.C.; Poignard, P.; Burton, D.R.; Zhu, J. Toward a More Accurate View of Human B-Cell Repertoire by next-Generation Sequencing, Unbiased Repertoire Capture and Single-Molecule Barcoding. Sci. Rep. 2014, 4, 6778. [Google Scholar] [CrossRef]
- Bregenholt, S.; Jensen, A.; Lantto, J.; Hyldig, S.; Haurum, J.S. Recombinant Human Polyclonal Antibodies: A New Class of Therapeutic Antibodies against Viral Infections. Curr. Pharm. Des. 2006, 12, 2007–2015. [Google Scholar] [CrossRef]
- Zhu, J.; Wu, X.; Zhang, B.; McKee, K.; O’Dell, S.; Soto, C.; Zhou, T.; Casazza, J.P.; NISC Comparative Sequencing Program; Mullikin, J.C.; et al. De Novo Identification of VRC01 Class HIV-1-Neutralizing Antibodies by next-Generation Sequencing of B-Cell Transcripts. Proc. Natl. Acad. Sci. USA 2013, 110, E4088–E4097. [Google Scholar] [CrossRef] [Green Version]
- Kepler, T.B.; Liao, H.-X.; Alam, S.M.; Bhaskarabhatla, R.; Zhang, R.; Yandava, C.; Stewart, S.; Anasti, K.; Kelsoe, G.; Parks, R.; et al. Immunoglobulin Gene Insertions and Deletions in the Affinity Maturation of HIV-1 Broadly Reactive Neutralizing Antibodies. Cell Host Microbe 2014, 16, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Sok, D.; Laserson, U.; Laserson, J.; Liu, Y.; Vigneault, F.; Julien, J.-P.; Briney, B.; Ramos, A.; Saye, K.F.; Le, K.; et al. The Effects of Somatic Hypermutation on Neutralization and Binding in the PGT121 Family of Broadly Neutralizing HIV Antibodies. PLoS Pathog. 2013, 9, e1003754. [Google Scholar] [CrossRef]
- Lavinder, J.J.; Wine, Y.; Giesecke, C.; Ippolito, G.C.; Horton, A.P.; Lungu, O.I.; Hoi, K.H.; DeKosky, B.J.; Murrin, E.M.; Wirth, M.M.; et al. Identification and Characterization of the Constituent Human Serum Antibodies Elicited by Vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 2259–2264. [Google Scholar] [CrossRef] [Green Version]
- Lindesmith, L.C.; McDaniel, J.R.; Changela, A.; Verardi, R.; Kerr, S.A.; Costantini, V.; Brewer-Jensen, P.D.; Mallory, M.L.; Voss, W.N.; Boutz, D.R.; et al. Sera Antibody Repertoire Analyses Reveal Mechanisms of Broad and Pandemic Strain Neutralizing Responses after Human Norovirus Vaccination. Immunity 2019, 50, 1530–1541.e8. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Paparoditis, P.; Horton, A.P.; Frühwirth, A.; McDaniel, J.R.; Jung, J.; Boutz, D.R.; Hussein, D.A.; Tanno, Y.; Pappas, L.; et al. Persistent Antibody Clonotypes Dominate the Serum Response to Influenza over Multiple Years and Repeated Vaccinations. Cell Host Microbe 2019, 25, 367–376.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belongia, E.A.; Skowronski, D.M.; McLean, H.Q.; Chambers, C.; Sundaram, M.E.; De Serres, G. Repeated Annual Influenza Vaccination and Vaccine Effectiveness: Review of Evidence. Expert Rev. Vaccines 2017, 16, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, K. Can We Improve Vaccine Efficacy by Targeting T and B Cell Repertoire Convergence? Front. Immunol. 2019, 10, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, W.; Rosenfeld, A.M.; Luning Prak, E.T. Mining the Antibody Repertoire for Solutions to SARS-CoV-2. Cell Host Microbe 2020, 28, 499–501. [Google Scholar] [CrossRef]
- Schultheiß, C.; Paschold, L.; Simnica, D.; Mohme, M.; Willscher, E.; von Wenserski, L.; Scholz, R.; Wieters, I.; Dahlke, C.; Tolosa, E.; et al. Next-Generation Sequencing of T and B Cell Receptor Repertoires from COVID-19 Patients Showed Signatures Associated with Severity of Disease. Immunity 2020, 53, 442–455.e4. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, S.; Lee, Y.; Grubbs, G.; Coyle, E.M.; Klenow, L.; Akasaka, O.; Koga, M.; Adachi, E.; Saito, M.; Nakachi, I.; et al. Longitudinal Antibody Repertoire in “Mild” versus “Severe” COVID-19 Patients Reveals Immune Markers Associated with Disease Severity and Resolution. Sci. Adv. 2021, 7, eabf2467. [Google Scholar] [CrossRef] [PubMed]
- Hook, L.M.; Awasthi, S.; Dubin, J.; Flechtner, J.; Long, D.; Friedman, H.M. A Trivalent GC2/GD2/GE2 Vaccine for Herpes Simplex Virus Generates Antibody Responses That Block Immune Evasion Domains on GC2 Better than Natural Infection. Vaccine 2019, 37, 664–669. [Google Scholar] [CrossRef]
- Lurie, N.; Saville, M.; Hatchett, R.; Halton, J. Developing COVID-19 Vaccines at Pandemic Speed. N. Engl. J. Med. 2020, 21, 1969–1973. [Google Scholar] [CrossRef]
- Yamey, G.; Schäferhoff, M.; Hatchett, R.; Pate, M.; Zhao, F.; McDade, K.K. Ensuring Global Access to COVID-19 Vaccines. Lancet 2020, 50, 273–275. [Google Scholar] [CrossRef]
- Tripp, R.A.; Ross, T.M. Development of a Zika Vaccine. Expert Rev. Vaccines 2016, 15, 1083–1085. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; He, Y.; Liu, S. SARS Vaccine Development. Emerg. Infect. Dis. 2005, 11, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.L.; Bavari, S. A Race for an Ebola Vaccine: Promises and Obstacles. Trends Microbiol. 2015, 23, 65–66. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.D. The Importance of Epitope Binning for Biological Drug Discovery. Curr. Drug Discov. Technol. 2014, 11, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Noy-Porat, T.; Makdasi, E.; Alcalay, R.; Mechaly, A.; Levy, Y.; Bercovich-Kinori, A.; Zauberman, A.; Tamir, H.; Yahalom-Ronen, Y.; Israeli, M.; et al. A Panel of Human Neutralizing MAbs Targeting SARS-CoV-2 Spike at Multiple Epitopes. Nat. Commun. 2020, 11, 4303. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients with COVID-19 in Wuhan, China; Social Science Research Network: Rochester, NY, USA, 2020. [Google Scholar]
- Andrew, M.K.; Bowles, S.K.; Pawelec, G.; Haynes, L.; Kuchel, G.A.; McNeil, S.A.; McElhaney, J.E. Influenza Vaccination in Older Adults: Recent Innovations and Practical Applications. Drugs Aging 2019, 36, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Stamatatos, L.; Czartoski, J.; Wan, Y.-H.; Homad, L.J.; Rubin, V.; Glantz, H.; Neradilek, M.; Seydoux, E.; Jennewein, M.F.; MacCamy, A.J.; et al. MRNA Vaccination Boosts Cross-Variant Neutralizing Antibodies Elicited by SARS-CoV-2 Infection. Science 2021, 372, 1413–1418. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Quadeer, A.A.; McKay, M.R. Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies. Viruses 2020, 12, 254. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.A.; Burkley, B.M.; Langaee, T.Y.; Clare-Salzler, M.J.; Klein, T.E.; Altman, R.B. Implementing Personalized Medicine: Development of a Cost-Effective Customized Pharmacogenetics Genotyping Array. Clin. Pharmacol. Ther. 2012, 92, 437–439. [Google Scholar] [CrossRef] [Green Version]
- Poland, G.A.; Ovsyannikova, I.G.; Jacobson, R.M. Personalized Vaccines: The Emerging Field of Vaccinomics. Expert Opin. Biol. Ther. 2008, 8, 1659–1667. [Google Scholar] [CrossRef]
- Loomis, R.J.; Johnson, P.R. Emerging Vaccine Technologies. Vaccines 2015, 3, 429–447. [Google Scholar] [CrossRef] [Green Version]
- Koff, W.C.; Burton, D.R.; R. Johnson, P.; Walker, B.D.; King, C.R.; Nabel, G.J.; Ahmed, R.; Bhan, M.K.; Plotkin, S.A. Accelerating Next Generation Vaccine Development for Global Disease Prevention. Science 2013, 340, 1232910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet Lond. Engl. 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Pronker, E.S.; Weenen, T.C.; Commandeur, H.R.; Osterhaus, A.D.M.E.; Claassen, H.J.H.M. The Gold Industry Standard for Risk and Cost of Drug and Vaccine Development Revisited. Vaccine 2011, 29, 5846–5849. [Google Scholar] [CrossRef] [PubMed]
- Poland, G.A.; Ovsyannikova, I.G.; Kennedy, R.B. Personalized Vaccinology: A Review. Vaccine 2018, 36, 5350–5357. [Google Scholar] [CrossRef]
- Ohashi, W.; Mizushima, H.; Tanaka, H. Economic Advantage of Pharmacogenomics - Clinical Trials with Genetic Information. Stud. Health Technol. Inform. 2008, 136, 585–590. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brooks, B.D.; Beland, A.; Aguero, G.; Taylor, N.; Towne, F.D. Moving beyond Titers. Vaccines 2022, 10, 683. https://doi.org/10.3390/vaccines10050683
Brooks BD, Beland A, Aguero G, Taylor N, Towne FD. Moving beyond Titers. Vaccines. 2022; 10(5):683. https://doi.org/10.3390/vaccines10050683
Chicago/Turabian StyleBrooks, Benjamin D., Alexander Beland, Gabriel Aguero, Nicholas Taylor, and Francina D. Towne. 2022. "Moving beyond Titers" Vaccines 10, no. 5: 683. https://doi.org/10.3390/vaccines10050683