
Preparedness for the Dengue Epidemic: Vaccine as a Viable Approach
 , , , , , and
, , , , , and
Abstract
:1. Introduction
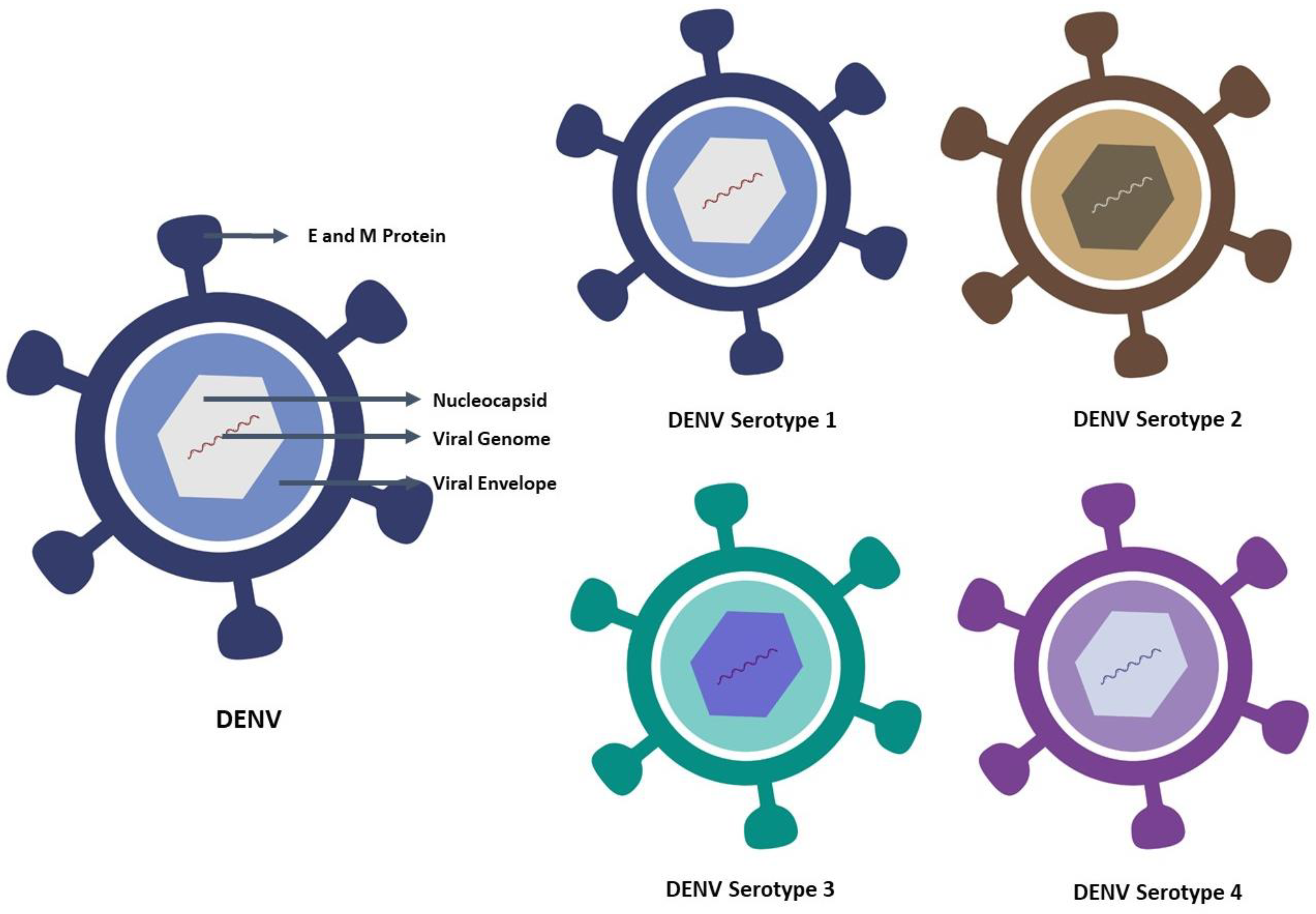
2. Characteristic Features of Dengue Virus and Its Serotype
3. Incidence/Prevalence of Dengue Disease
4. Epidemiology and Global Expansion
5. Transmission of Dengue Virus/Fever
6. Preventive Measures for Dengue Fever
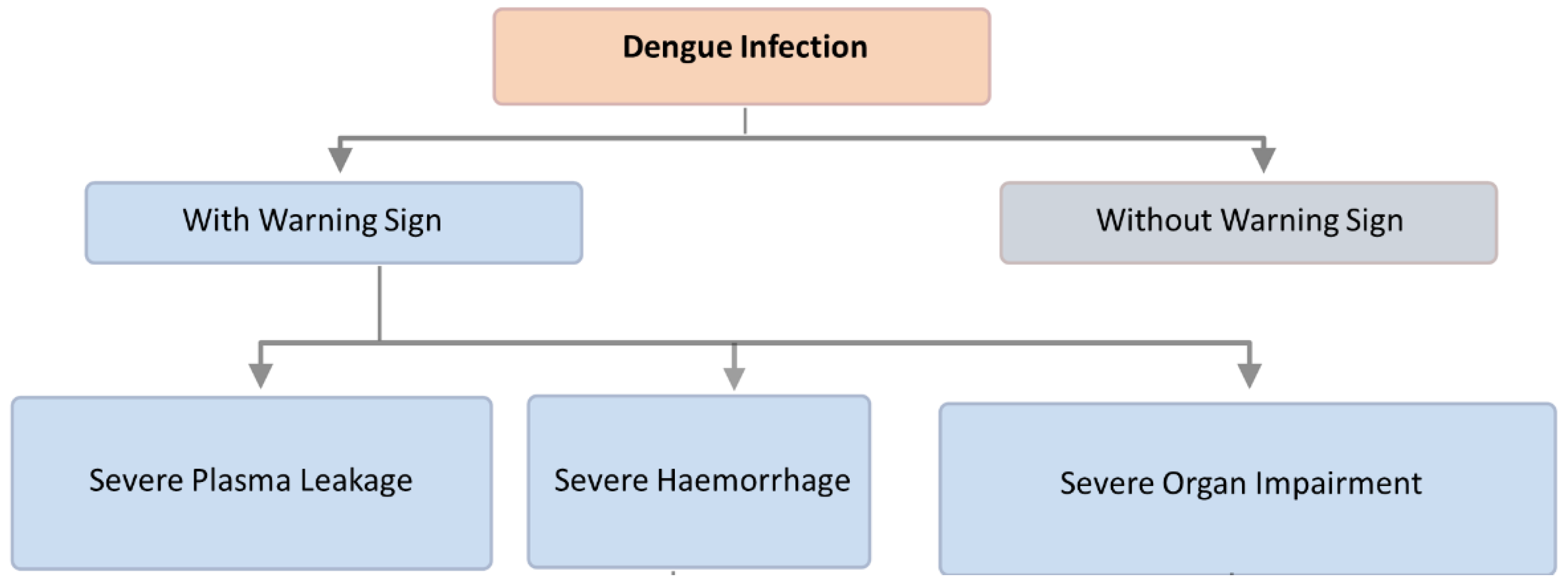
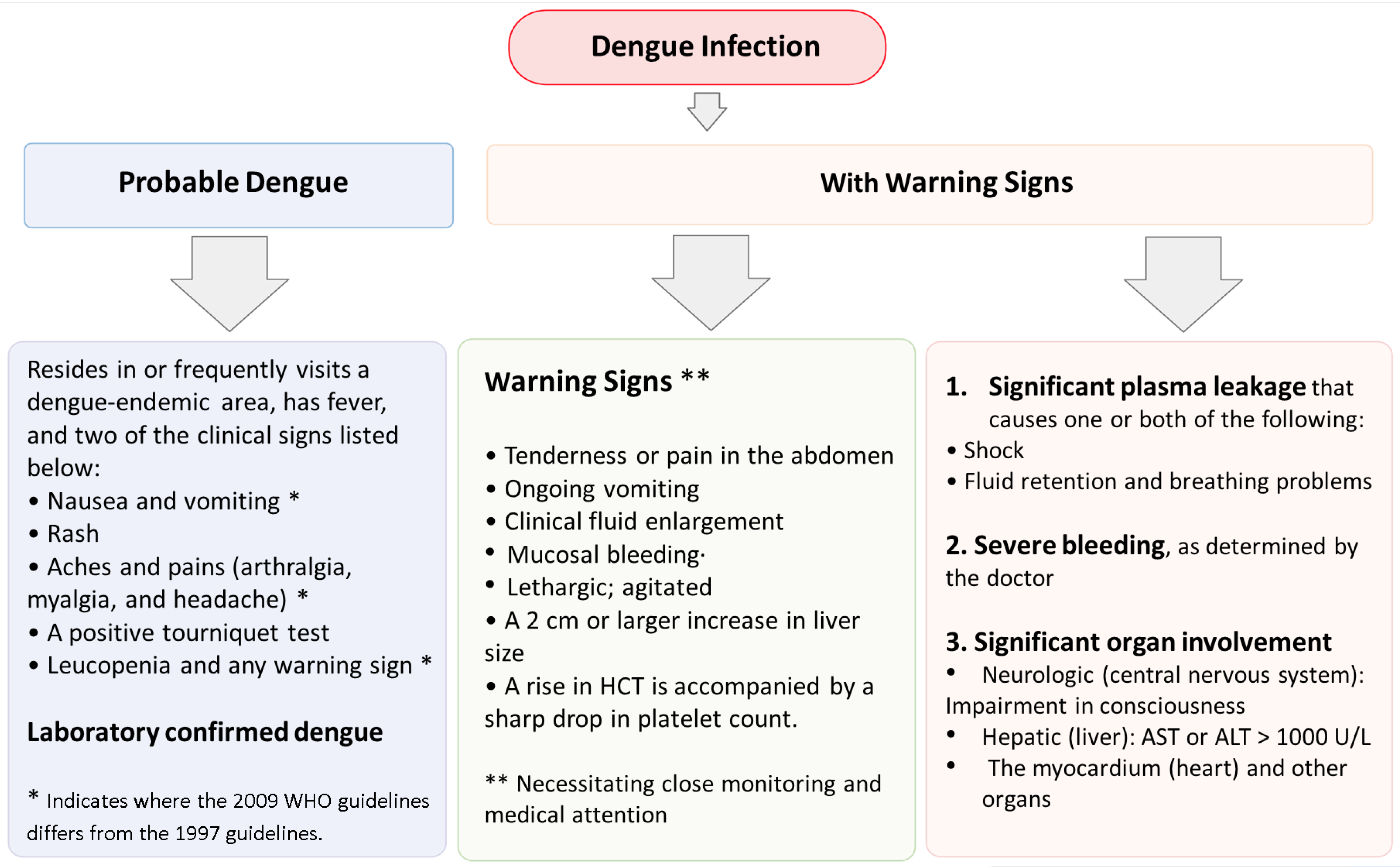
7. Diagnostic Methods for Dengue
8. Diagnostic Assays in Development
9. Treatment of Dengue Fever
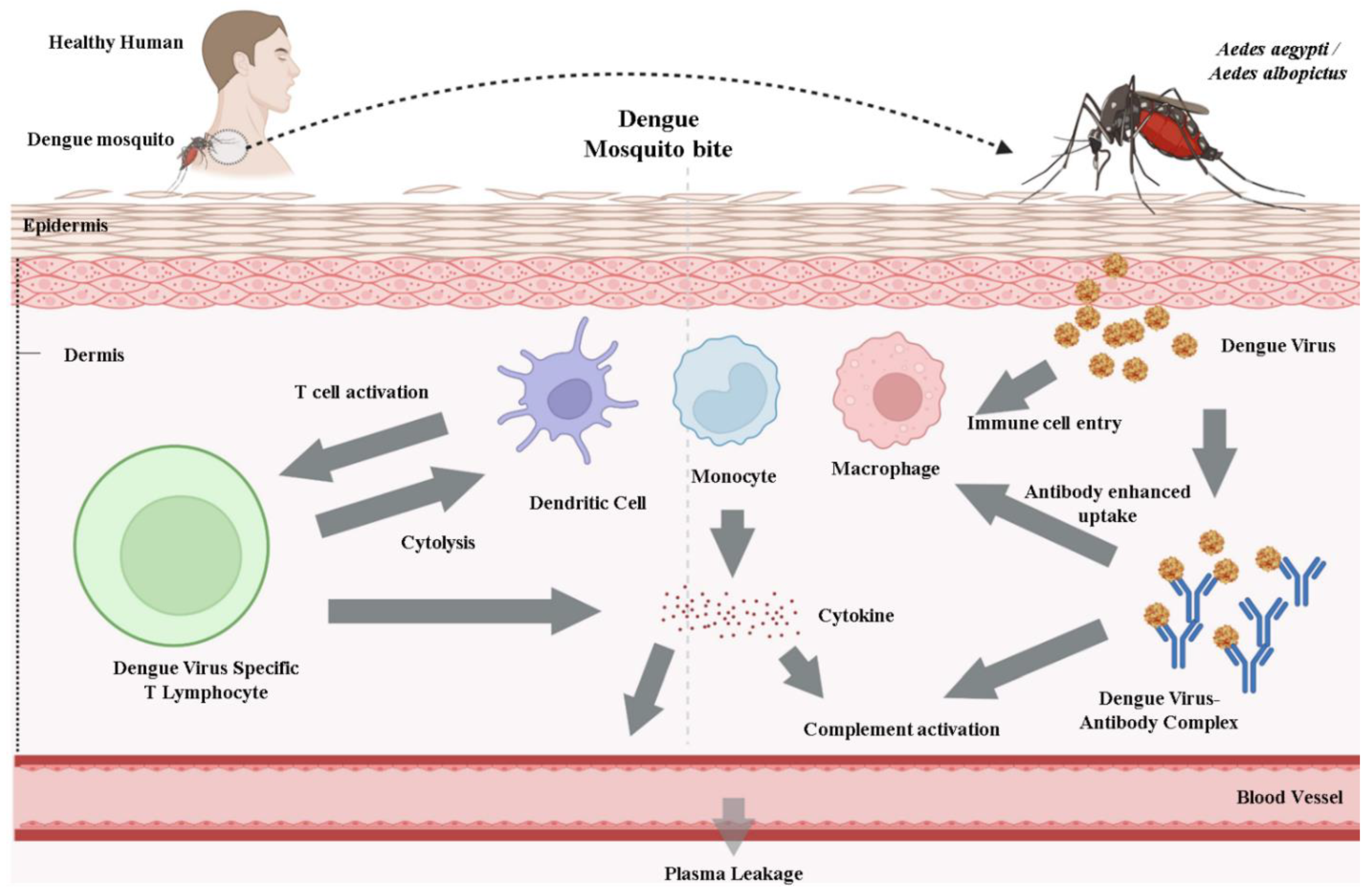
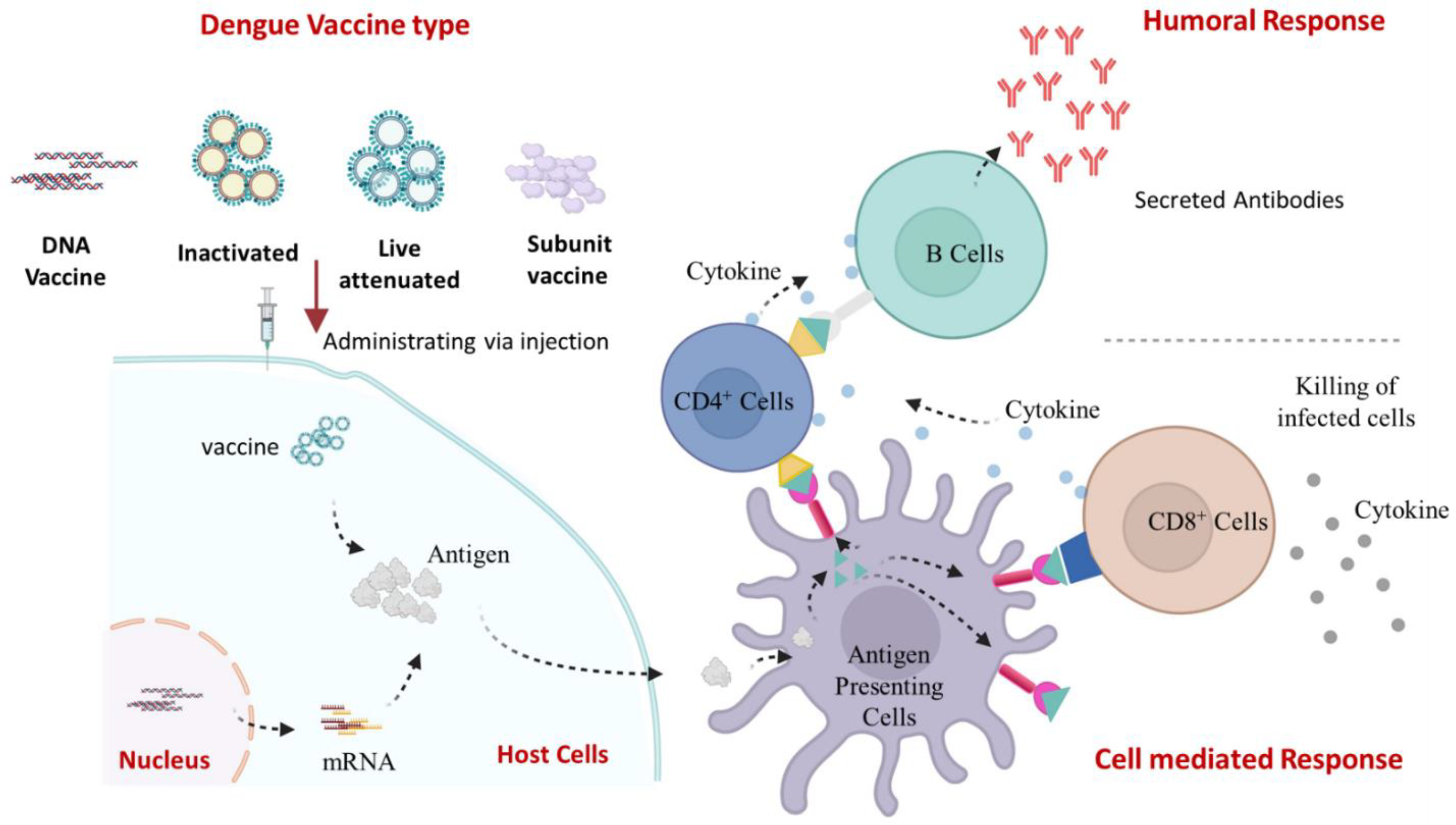
10. Viral Life Cycle and Host Immune Response
11. Dengue Vaccine Development and Progress
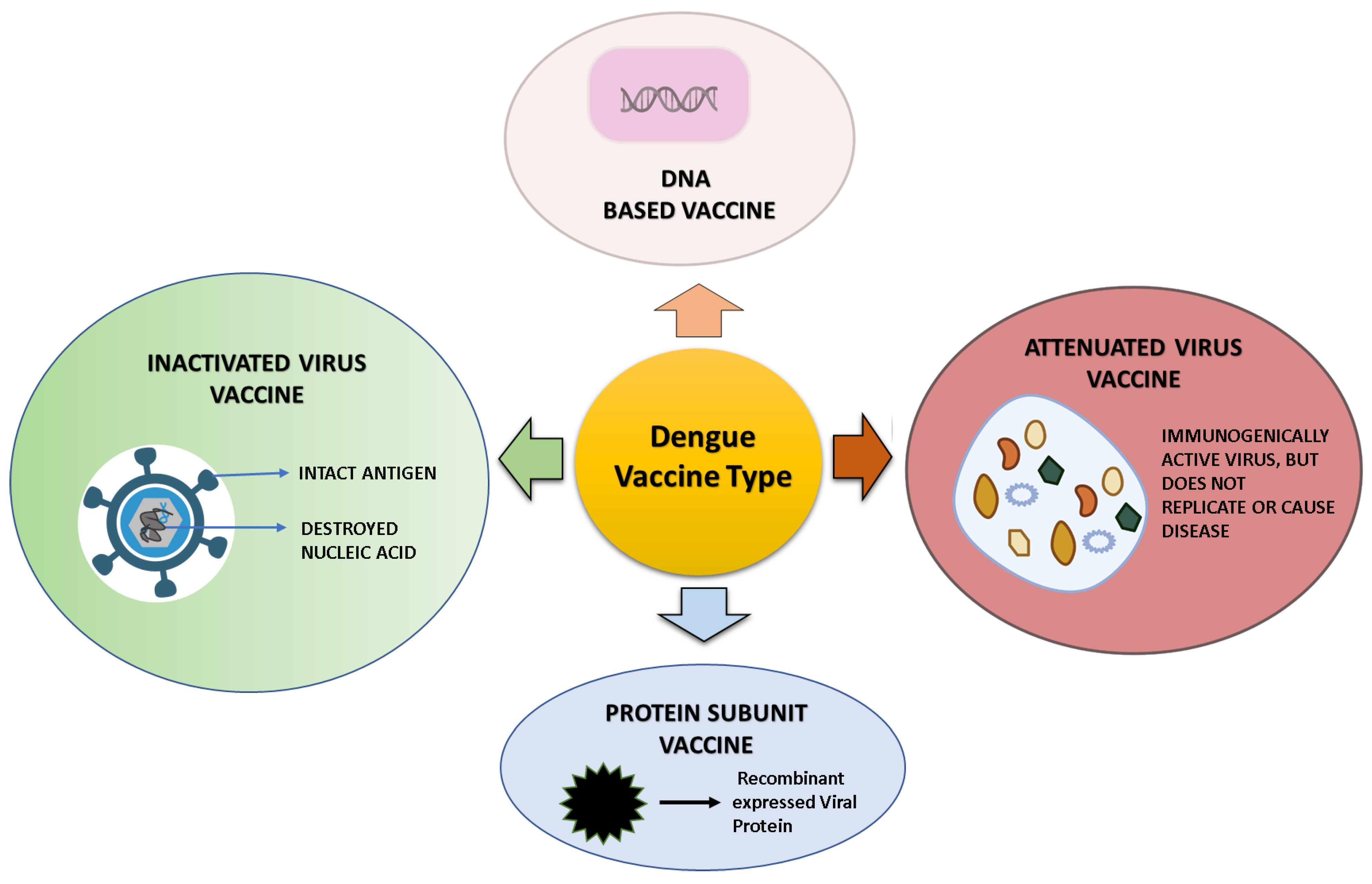
12. Dengue Vaccine Platform and Vaccine in a Developmental Phase
12.1. Live-Attenuated DENV Vaccines
12.2. DENV DNA Vaccine
12.3. Subunit DENV Vaccine
12.4. Inactivated Dengue Vaccine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine/Other Name | Efficacy | Serotype-Specific Efficacy | Immunogenicity | Comments/Side Effect | References |
|---|---|---|---|---|---|
| Dengvaxia® (CYD-TDV) (ChimeriVax; ChimeriVax Tetravalent Dengue Vaccine; ChimeriVax-DEN1-4; ChimeriVax-Dengue; CYD dengue vaccine; CYD dengue vaccine—Sanofi; CYD-4444; CYD-5553; CYD-5555; CYD-TDV; CYD-TDV Dengue Vaccine; Dengue fever vaccine—Sanofi; Dengue Tetravalent Vaccine, Live—Sanofi; Dengue vaccine recombinant tetravalent—Sanofi; Dengue virus vaccine—Sanofi; Dengvaxia; Live-attenuated, dengue serotype 1, 2, 3, 4 virus vaccine; Tetravalent CYD 1, 2, 3, 4 dengue; Tetravalent dengue vaccine—Sanofi; TV-CYD) | 25–59% | DENV-4 > DENV-3 > DENV-1 > DENV-2 | CD8+ respond primarily to NS3, mainly DENV-4-neutralizing antibodies | Age restriction; increased risk of severe dengue in seronegative people; good efficacy and safety in seropositive persons; increases hospitalizations in vaccination recipients | [129,130,131] |
| DENVax/TAK003/TDV (DEN1-DEN2-DEN3-DEN4 vaccine—Takeda Pharmaceuticals USA; Dengue vaccine tetravalent—Takeda Pharmaceuticals USA; DENVax™; Needle-free dengue vaccine; QDENGA; TAK-003; TDV—Takeda Pharmaceuticals USA; Tetravalent dengue vaccine—Takeda Pharmaceuticals USA) | 73.3–85.3% | DENV-2 > DENV-1 > DENV-4 | Antibodies effective against all four serotypes | Children and teenagers tolerate it well; Independent of participant age or serostatus, immunogenic and well tolerated in several phase I and II clinical trials; Safety profile unknown | [89,132,133] |
| TetraVax-DV-V003/TV005 (Tetravalent live-attenuated dengue vaccine admixture TV 005; TetraVax-DV TV005; TetraVax-DV-Vaccine—Admixture TV005; TV-005) | Not yet released | DENV-4 > DENV-3 > DENV-1 > DENV-2 | Rhesus macaque strong neutralizing antibodies | Single dose well tolerated; balanced immunological response; efficient with one dose administration; adverse response (mild rash) | [88,95,131,134] |
| TDEN LAV (Dengue vaccine live—GlaxoSmithKline; T-DEN F-19; T-DEN F17; TDENV-LAV F17; Tetravalent dengue live-attenuated virus formulation 17; WRAIR Live-Attenuated Tetravalent Dengue Vaccine) | Not yet released | Not yet released | Not yet released | Discontinued | [135] |
| TDENV PIV (Tetravalent purified inactivated dengue vaccine—GlaxoSmithKline/Walter Reed Army Institute) | Not yet released | Not yet released | Not yet released | No recent information, well tolerated and immunogenic in seropositive and naïve people. No reactivation risk and a healthy immunological balance | [128] |
12.5. Commercially Available DENV Vaccine
13. Use of CYD-TDV Vaccine and WHO Position
14. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [Green Version]
- Afreen, N.; Deeba, F.; Khan, W.H.; Haider, S.H.; Kazim, S.N.; Ishrat, R.; Naqvi, I.H.; Shareef, M.Y.; Broor, S.; Ahmed, A.; et al. Molecular characterization of dengue and chikungunya virus strains circulating in New Delhi, India. Microbiol. Immunol. 2014, 58, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Fitzmaurice, C.; Allen, C.; Barber, R.M.; Barregard, L.; Bhutta, Z.A.; Brenner, H.; Dicker, D.J.; Chimed-Orchir, O.; Dandona, R.; Dandona, L. Global, regional, and national cancer incidence, mortality, years of life lost, years lived with disability, and disability-adjusted life-years for 32 cancer groups, 1990 to 2015: A systematic analysis for the global burden of disease study. JAMA Oncol. 2017, 3, 524–548. [Google Scholar] [PubMed]
- Horstick, O.; Tozan, Y.; Wilder-Smith, A. Reviewing dengue: Still a neglected tropical disease? PLoS Negl. Trop. Dis. 2015, 9, e0003632. [Google Scholar] [CrossRef] [Green Version]
- Khan, W.H.; Ahmad, R.; Khan, N.; Ansari, M.A. SARS-CoV-2 Variants Associated Challenges in the Ongoing Vaccination of COVID-19. In Prime Archives in Virology; Raad, H., Ed.; Prime Archives in Virology: Hyderabad, India, 2022; Volume 1, pp. 1–59. [Google Scholar]
- Zeyaullah, M.; AlShahrani, A.M.; Muzammil, K.; Ahmad, I.; Alam, S.; Khan, W.H.; Ahmad, R. COVID-19 and SARS-CoV-2 Variants: Current Challenges and Health Concern. Front. Genet. 2021, 12, 693916. [Google Scholar] [CrossRef] [PubMed]
- Goel, N.; Ahmad, R.; Fatima, H.; Khare, S.K. New threatening of SARS-CoV-2 coinfection and strategies to fight the current pandemic. Med. Drug Discov. 2021, 10, 100089. [Google Scholar] [CrossRef]
- Ramakrishnan, L.; Pillai, M.R.; Nair, R.R. Dengue Vaccine Development: Strategies and Challenges. Viral Immunol. 2015, 28, 76–84. [Google Scholar] [CrossRef]
- Simmons, C.P.; Farrar, J.J.; van Vinh Chau, N.; Wills, B. Dengue. N. Engl. J. Med. 2012, 366, 1423–1432. [Google Scholar] [CrossRef]
- Lindenbach, B.D.; Rice, C.M. Molecular biology of flaviviruses. Adv. Virus Res. 2003, 59, 23–61. [Google Scholar] [CrossRef]
- Perera, R.; Kuhn, R.J. Structural proteomics of dengue virus. Curr. Opin. Microbiol. 2008, 11, 369–377. [Google Scholar] [CrossRef]
- Heinz, F.X.; Allison, S.L. Flavivirus Structure and Membrane Fusion. Adv. Virus Res. 2003, 59, 63–97. [Google Scholar] [CrossRef] [PubMed]
- Wahala, W.M.P.B.; De Silva, A.M. The Human Antibody Response to Dengue Virus Infection. Viruses 2011, 3, 2374–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waggoner, J.J.; Gresh, L.; Vargas, M.J.; Ballesteros, G.; Tellez, Y.; Soda, K.J.; Sahoo, M.K.; Nuñez, A.; Balmaseda, A.; Harris, E.; et al. Viremia and Clinical Presentation in Nicaraguan Patients Infected With Zika Virus, Chikungunya Virus, and Dengue Virus. Clin. Infect. Dis. 2016, 63, 1584–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Genet. 2010, 8, S7–S16. [Google Scholar] [CrossRef] [Green Version]
- Alejandria, M.M. Dengue haemorrhagic fever or dengue shock syndrome in children. BMJ Clin. Evid. 2015, 2015, 0917. [Google Scholar]
- Srikiatkhachorn, A. Plasma leakage in dengue haemorrhagic fever. Thromb. Haemost. 2009, 102, 1042–1049. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.; Zhang, R.; Cheng, G. Progress towards understanding the pathogenesis of dengue hemorrhagic fever. Virol. Sin. 2017, 32, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Hermann, L.L.; Gupta, S.B.; Manoff, S.B.; Kalayanarooj, S.; Gibbons, R.V.; Coller, B.-A.G. Advances in the understanding, management, and prevention of dengue. J. Clin. Virol. 2015, 64, 153–159. [Google Scholar] [CrossRef]
- Hladish, T.J.; Pearson, C.A.B.; Chao, D.L.; Rojas, D.P.; Recchia, G.L.; Gómez-Dantés, H.; Halloran, M.E.; Pulliam, J.R.C.; Longini, I.M. Projected impact of dengue vaccination in Yucatán, Mexico. PLoS Negl. Trop. Dis. 2016, 10, e0004661. [Google Scholar] [CrossRef] [Green Version]
- Recker, M.; Vannice, K.; Hombach, J.; Jit, M.; Simmons, C.P. Assessing dengue vaccination impact: Model challenges and future directions. Vaccine 2016, 34, 4461–4465. [Google Scholar] [CrossRef] [Green Version]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the Global Spatial Limits of Dengue Virus Transmission by Evidence-Based Consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue Haemorrhagic Fever: Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 1997. [Google Scholar]
- Kuo, H.-J.; Lee, I.-K.; Liu, J.-W. Analyses of clinical and laboratory characteristics of dengue adults at their hospital presentations based on the World Health Organization clinical-phase framework: Emphasizing risk of severe dengue in the elderly. J. Microbiol. Immunol. Infect. 2018, 51, 740–748. [Google Scholar] [CrossRef] [PubMed]
- WHO. Dengue Guidelines, for Diagnosis, Treatment, Prevention and Control. 2009. Available online: https://www.who.int/publications/i/item/9789241547871 (accessed on 5 October 2022).
- Lee, I.-K.; Hsieh, C.-J.; Lee, C.-T.; Liu, J.-W. Diabetic patients suffering dengue are at risk for development of dengue shock syndrome/severe dengue: Emphasizing the impacts of co-existing comorbidity(ies) and glycemic control on dengue severity. J. Microbiol. Immunol. Infect. 2020, 53, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-H.; Lin, C.-Y.; Chang, K.; Urbina, A.N.; Assavalapsakul, W.; Thitithanyanont, A.; Lu, P.-L.; Chen, Y.-H.; Wang, S.-F. A clinical and epidemiological survey of the largest dengue outbreak in Southern Taiwan in 2015. Int. J. Infect. Dis. 2019, 88, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.-Y.; Lee, I.-K.; Lee, C.-H.; Yang, K.D.; Liu, J.-W. Comparisons of dengue illness classified based on the 1997 and 2009 World Health Organization dengue classification schemes. J. Microbiol. Immunol. Infect. 2013, 46, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Neumayr, A.; Muñoz, J.; Schunk, M.; Bottieau, E.; Cramer, J.; Calleri, G.; López-Vélez, R.; Angheben, A.; Zoller, T.; Visser, L.; et al. Sentinel surveillance of imported dengue via travellers to Europe 2012 to 2014: TropNet data from the DengueTools Research Initiative. Eurosurveillance 2017, 22, 30433. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M. The traveller and emerging infections: Sentinel, courier, transmitter. J. Appl. Microbiol. 2003, 94, 1–11. [Google Scholar] [CrossRef]
- World Health Organization. Dengue and Dengue Haemorrhagic Fever. Factsheet No 117; Revised May 2008. World Health Organization: Geneva, Switzerland. Available online: http://www.who.int/mediacentre/factsheets/fs117/en/ (accessed on 5 October 2022).
- World Health Organization. Dengue Fever and Dengue Haemorrhagic Fever Prevention and Control; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- World Health Organization. Revision of the International Health Regulations. World Health Assembly Resolution WHA58.3, Adopted by the 58th World Health Assembly. 2005. Available online: http://www.who.int/gb/ebwha/pdf_files/WHA58/WHA58_3-en.pdf (accessed on 5 October 2022).
- WHO/EMRO. World Health Organization, Regional Office for the Eastern Mediterranean, Division of Communicable Disease Control, Newsletter, I2005,6:7-8. Available online: http://www.emro.who.int/pdf/dcdnewsletter6.pdf (accessed on 5 October 2022).
- Hyams, K.C.; Oldfield, E.C.; Scott, R.M.; Bourgeois, A.L.; Gardiner, H.; Pazzaglia, G.; Moussa, M.; Saleh, A.S.; Dawi, O.E.; Daniell, F.D. Evaluation of Febrile Patients in Port Sudan, Sudan: Isolation of Dengue Virus. Am. J. Trop. Med. Hyg. 1986, 35, 860–865. [Google Scholar] [CrossRef]
- Kodier, G.R.; Gubler, D.J.; Cope, S.E.; Cropp, C.; Soliman, A.K.; Polycarpe, D.; Abdourhaman, M.A.; Parra, J.-P.; Maslin, J.; Arthur, R.R. Epidemic dengue 2 in the city of Djibouti 1991–1992. Trans. R. Soc. Trop. Med. Hyg. 1996, 90, 237–240. [Google Scholar] [CrossRef]
- Jamil, B.; Hasan, R.; Zafar, A.; Bewley, K.; Chamberlain, J.; Mioulet, V.; Rowlands, M.; Hewson, R. Dengue virus serotype 3, Karachi, Pakistan. Emerg. Infect. Dis. 2007, 13, 182–183. [Google Scholar] [CrossRef]
- Gupta, N.; Srivastava, S.; Jain, A.; Chaturvedi, U.C. Dengue in India. Indian J. Med. Res. 2012, 136, 373–390. [Google Scholar]
- World Health Organization. Denguenet in India. Wkly. Epidemiol. Rec. 2004, 79, 201–203. Available online: https://apps.who.int/iris/bitstream/handle/10665/232483/WER7921_201-203 (accessed on 5 October 2022).
- WHO/SEARO. Concrete Measure Key in Controlling Dengue in South East Asia. Press Release SEA/PR/1479. New Delhi, World Health Organization Regional Office for South-East Asia. Available online: http://www.searo.who.int/EN/Section316/Section503/Section2463_14619.htm (accessed on 5 October 2022).
- World Health Organization. Regional Office for South-East Asia. Comprehensive Guideline for Prevention and Control of Dengue and Dengue Haemorrhagic Fever; Revised and Expanded Edition; WHO Regional Office for South-East Asia. 2011. Available online: https://apps.who.int/iris/handle/10665/204894 (accessed on 5 October 2022).
- Muller, D.A.; Depelsenaire, A.; Young, P.R. Clinical and Laboratory Diagnosis of Dengue Virus Infection. J. Infect. Dis. 2017, 215, S89–S95. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, E.; Mileguir, F.; Grossman, Z.; Mendelson, E. Evaluation of ELISA-based sero-diagnosis of dengue fever in travelers. J. Clin. Virol. 2000, 19, 169–173. [Google Scholar] [CrossRef]
- Guzmán, M.G.; Kourı, G. Dengue diagnosis, advances and challenges. Int. J. Infect. Dis. 2004, 8, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Hunsperger, E.A.; Yoksan, S.; Buchy, P.; Nguyen, V.C.; Sekaran, S.D.; Enria, D.A.; Pelegrino, J.L.; Vázquez, S.; Artsob, H.; Drebot, M.; et al. Evaluation of Commercially Available Anti–Dengue Virus Immunoglobulin M Tests. Emerg. Infect. Dis. 2009, 15, 436–440. [Google Scholar] [CrossRef]
- Callahan, J.D.; Wu, S.-J.L.; Dion-Schultz, A.; Mangold, B.E.; Peruski, L.F.; Watts, D.M.; Porter, K.R.; Murphy, G.R.; Suharyono, W.; King, C.-C.; et al. Development and Evaluation of Serotype- and Group-Specific Fluorogenic Reverse Transcriptase PCR (TaqMan) Assays for Dengue Virus. J. Clin. Microbiol. 2001, 39, 4119–4124. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-J.L.; Lee, E.M.; Putvatana, R.; Shurtliff, R.N.; Porter, K.R.; Suharyono, W.; Watts, D.M.; King, C.-C.; Murphy, G.S.; Hayes, C.G.; et al. Detection of Dengue Viral RNA Using a Nucleic Acid Sequence-Based Amplification Assay. J. Clin. Microbiol. 2001, 39, 2794–2798. [Google Scholar] [CrossRef] [Green Version]
- Fry, S.R.; Meyer, M.; Semple, M.G.; Simmons, C.P.; Sekaran, S.D.; Huang, J.X.; McElnea, C.; Huang, C.-Y.; Valks, A.; Young, P.R.; et al. The Diagnostic Sensitivity of Dengue Rapid Test Assays Is Significantly Enhanced by Using a Combined Antigen and Antibody Testing Approach. PLoS Negl. Trop. Dis. 2011, 5, e1199. [Google Scholar] [CrossRef] [Green Version]
- Hang, V.T.; Nguyet, N.M.; Trung, D.T.; Tricou, V.; Yoksan, S.; Dung, N.M.; Van Ngoc, T.; Hien, T.T.; Farrar, J.; Wills, B.; et al. Diagnostic Accuracy of NS1 ELISA and Lateral Flow Rapid Tests for Dengue Sensitivity, Specificity and Relationship to Viraemia and Antibody Responses. PLoS Negl. Trop. Dis. 2009, 3, e360. [Google Scholar] [CrossRef]
- Guzman, M.G.; Jaenisch, T.; Gaczkowski, R.; Hang, V.T.T.; Sekaran, S.D.; Kroeger, A.; Vazquez, S.; Ruiz, D.; Martinez, E.; Mercado, J.C.; et al. Multi-Country Evaluation of the Sensitivity and Specificity of Two Commercially-Available NS1 ELISA Assays for Dengue Diagnosis. PLoS Negl. Trop. Dis. 2010, 4, e811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaterji, S.; Allen, J.C., Jr.; Chow, A.; Leo, Y.-S.; Ooi, E.-E. Evaluation of the NS1 rapid test and the WHO dengue classification schemes for use as bedside diagnosis of acute dengue fever in adults. Am. J. Trop. Med. Hyg. 2011, 84, 224. [Google Scholar] [CrossRef] [PubMed]
- Duong, V.; Ly, S.; Try, P.L.; Tuiskunen, A.; Ong, S.; Chroeung, N.; Lundkvist, A.; Leparc-Goffart, I.; Deubel, V.; Vong, S.; et al. Clinical and Virological Factors Influencing the Performance of a NS1 Antigen-Capture Assay and Potential Use as a Marker of Dengue Disease Severity. PLoS Negl. Trop. Dis. 2011, 5, e1244. [Google Scholar] [CrossRef] [PubMed]
- Tricou, V.; Minh, N.N.; Farrar, J.; Tran, H.T.; Simmons, C.P. Kinetics of Viremia and NS1 Antigenemia Are Shaped by Immune Status and Virus Serotype in Adults with Dengue. PLoS Negl. Trop. Dis. 2011, 5, e1309. [Google Scholar] [CrossRef] [Green Version]
- Huhtamo, E.; Hasu, E.; Uzcátegui, N.Y.; Erra, E.; Nikkari, S.; Kantele, A.; Vapalahti, O.; Piiparinen, H. Early diagnosis of dengue in travelers: Comparison of a novel real-time RT-PCR, NS1 antigen detection and serology. J. Clin. Virol. 2010, 47, 49–53. [Google Scholar] [CrossRef]
- Watthanaworawit, W.; Turner, P.; Turner, C.L.; Tanganuchitcharnchai, A.; Jarman, R.G.; Blacksell, S.D.; Nosten, F.H. A prospective evaluation of diagnostic methodologies for the acute diagnosis of dengue virus infection on the Thailand-Myanmar border. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Fox, A.; Hoa, L.N.M.; Simmons, C.P.; Wolbers, M.; Wertheim, H.; Khuong, P.T.; Ninh, T.T.H.; Lien, T.T.M.; Lien, N.T.; Trung, N.V.; et al. Immunological and Viral Determinants of Dengue Severity in Hospitalized Adults in Ha Noi, Viet Nam. PLoS Negl. Trop. Dis. 2011, 5, e967. [Google Scholar] [CrossRef]
- Domingo, C.; de Ory, F.; Sanz, J.C.; Reyes, N.; Gascón, J.; Wichmann, O.; Puente, S.; Schunk, M.; López-Vélez, R.; Ruiz, J.; et al. Molecular and serologic markers of acute dengue infection in naive and flavivirus-vaccinated travelers. Diagn. Microbiol. Infect. Dis. 2009, 65, 42–48. [Google Scholar] [CrossRef]
- Matheus, S.; Deparis, X.; Labeau, B.; Lelarge, J.; Morvan, J.; Dussart, P. Discrimination between Primary and Secondary Dengue Virus Infection by an Immunoglobulin G Avidity Test Using a Single Acute-Phase Serum Sample. J. Clin. Microbiol. 2005, 43, 2793–2797. [Google Scholar] [CrossRef] [Green Version]
- Teoh, B.-T.; Sam, S.-S.; Tan, K.-K.; Johari, J.; Danlami, M.B.; Hooi, P.-S.; Md-Esa, R.; AbuBakar, S. Detection of dengue viruses using reverse transcription-loop-mediated isothermal amplification. BMC Infect. Dis. 2013, 13, 387. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-R.; Lai, Y.-C.; Yeh, T.-M. Dengue virus non-structural protein 1: A pathogenic factor, therapeutic target, and vaccine candidate. J. Biomed. Sci. 2018, 25, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, E.V.R.; De Oliveira, J.L.; Abrantes, D.C.; Rogério, C.B.; Bueno, C.; Miranda, V.R.; Monteiro, R.A.; Fraceto, L.F. Recent Developments in Nanotechnology for Detection and Control of Aedes aegypti-Borne Diseases. Front. Bioeng. Biotechnol. 2020, 8, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batool, K.; Alam, I.; Wu, S.; Liu, W.; Zhao, G.; Chen, M.; Wang, J.; Xu, J.; Huang, T.; Pan, X.; et al. Transcriptomic Analysis of Aedes aegypti in Response to Mosquitocidal Bacillus thuringiensis LLP29 Toxin. Sci. Rep. 2018, 8, 12650. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Khatoon, N.; Sardar, M. Antibacterial Effect of Green Synthesized TiO2 Nanoparticles. Adv. Sci. Lett. 2014, 20, 1616–1620. [Google Scholar] [CrossRef]
- Ahmad, R.; Mohsin, M.; Ahmad, T.; Sardar, M. Alpha amylase assisted synthesis of TiO2 nanoparticles: Structural characterization and application as antibacterial agents. J. Hazard. Mater. 2015, 283, 171–177. [Google Scholar] [CrossRef]
- Ahmad, R.; Srivastava, S.; Ghosh, S.; Khare, S.K. Phytochemical delivery through nanocarriers: A review. Colloids Surf. B Biointerfaces 2020, 197, 111389. [Google Scholar] [CrossRef]
- Sadaf, A.; Ahmad, R.; Ghorbal, A.; Elfalleh, W.; Khare, S.K. Synthesis of cost-effective magnetic nano-biocomposites mimicking peroxidase activity for remediation of dyes. Environ. Sci. Pollut. Res. 2019, 27, 27211–27220. [Google Scholar] [CrossRef]
- Suganya, P.; Vaseeharan, B.; Vijayakumar, S.; Balan, B.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Benelli, G. Biopolymer zein-coated gold nanoparticles: Synthesis, antibacterial potential, toxicity and histopathological effects against the Zika virus vector Aedes aegypti. J. Photochem. Photobiol. B Biol. 2017, 173, 404–411. [Google Scholar] [CrossRef]
- Noori, R.; Ahmad, R.; Sardar, M. Nanobiosensor in Health Sector: The Milestones Achieved and Future Prospects. In Nanobiosensors for Agricultural, Medical and Environmental Applications; Springer: Berlin/Heidelberg, Germany, 2020; pp. 63–90. [Google Scholar]
- Durán, N.; Islan, G.A.; Durán, M.; Castro, G.R. Nanobiotechnology Solutions against Aedes aegypti. J. Braz. Chem. Soc. 2016, 27, 1139–1149. [Google Scholar] [CrossRef]
- Benelli, G.; Caselli, A.; Canale, A. Nanoparticles for mosquito control: Challenges and constraints. J. King Saud Univ. Sci. 2017, 29, 424–435. [Google Scholar] [CrossRef]
- Nicolini, A.M.; McCracken, K.E.; Yoon, J.-Y. Future developments in biosensors for field-ready Zika virus diagnostics. J. Biol. Eng. 2017, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Khare, S.K. Immobilization of Aspergillus niger cellulase on multiwall carbon nanotubes for cellulose hydrolysis. Bioresour. Technol. 2018, 252, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Sardar, M. Enzyme immobilization: An overview on nanoparticles as immobilization matrix. Biochem. Anal. Biochem. 2015, 4, 1. [Google Scholar]
- Ghosh, S.; Ahmad, R.; Gautam, V.K.; Khare, S.K. Cholesterol-oxidase-magnetic nanobioconjugates for the production of 4-cholesten-3-one and 4-cholesten-3, 7-dione. Bioresour. Technol. 2018, 254, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Sousa, J.o.; Pais, A.; Vitorino, C. Nanomedicine: Principles, properties, and regulatory issues. Front. Chem. 2018, 6, 360. [Google Scholar] [CrossRef]
- Abdulla, N.K.; Siddiqui, S.I.; Fatima, B.; Sultana, R.; Tara, N.; Hashmi, A.A.; Ahmad, R.; Mohsin, M.; Nirala, R.K.; Linh, N.T.; et al. Silver based hybrid nanocomposite: A novel antibacterial material for water cleansing. J. Clean. Prod. 2020, 284, 124746. [Google Scholar] [CrossRef]
- Ellah, N.H.A.; Gad, S.F.; Muhammad, K.; E Batiha, G.; Hetta, H.F. Nanomedicine as a promising approach for diagnosis, treatment and prophylaxis against COVID-19. Nanomedicine 2020, 15, 2085–2102. [Google Scholar] [CrossRef]
- Ahmad, R.; Mishra, A.; Sardar, M. Simultaneous Immobilization and Refolding of Heat Treated Enzymes on TiO2 Nanoparticles. Adv. Sci. Eng. Med. 2014, 6, 1264–1268. [Google Scholar] [CrossRef]
- Ghosh, S.; Ahmad, R.; Banerjee, K.; AlAjmi, M.F.; Rahman, S. Mechanistic Aspects of Microbe-Mediated Nanoparticle Synthesis. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Ghosh, S.; Ahmad, R.; Khare, S. Refolding of thermally denatured cholesterol oxidases by magnetic nanoparticles. Int. J. Biol. Macromol. 2019, 138, 958–965. [Google Scholar] [CrossRef]
- Ghosh, S.; Ahmad, R.; Zeyaullah, M.; Khare, S.K. Microbial Nano-Factories: Synthesis and Biomedical Applications. Front. Chem. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.; Tripathi, N.K.; Dash, P.K.; Parida, M. Working towards dengue as a vaccine-preventable disease: Challenges and opportunities. Expert Opin. Biol. Ther. 2017, 17, 1193–1199. [Google Scholar] [CrossRef]
- Garcia, M.; Wehbe, M.; Lévêque, N.; Bodet, C. Skin innate immune response to flaviviral infection. Eur. Cytokine Netw. 2017, 28, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Welsch, S.; Miller, S.; Romero-Brey, I.; Merz, A.; Bleck, C.K.E.; Walther, P.; Fuller, S.D.; Antony, C.; Krijnse-Locker, J.; Bartenschlager, R. Composition and Three-Dimensional Architecture of the Dengue Virus Replication and Assembly Sites. Cell Host Microbe 2009, 5, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.-W.; Wu-Hsieh, B.A.; Lin, Y.-S.; Chen, W.-Y.; Huang, Y.; Anderson, R. The monocyte-macrophage-mast cell axis in dengue pathogenesis. J. Biomed. Sci. 2018, 25, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, C.A.; Wegman, A.D.; Endy, T.P. Mobilization and Activation of the Innate Immune Response to Dengue Virus. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Whitehead, S.S.; Durbin, A.P.; Pierce, K.K.; Elwood, D.; McElvany, B.D.; Fraser, E.A.; Carmolli, M.P.; Tibery, C.M.; Hynes, N.A.; Jo, M.; et al. In a randomized trial, the live attenuated tetravalent dengue vaccine TV003 is well-tolerated and highly immunogenic in subjects with flavivirus exposure prior to vaccination. PLoS Negl. Trop. Dis. 2017, 11, e0005584. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, S.S. Development of TV003/TV005, a single dose, highly immunogenic live attenuated dengue vaccine; what makes this vaccine different from the Sanofi-Pasteur CYD™ vaccine? Expert Rev. Vaccines 2016, 15, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Sáez-Llorens, X.; Tricou, V.; Yu, D.; Rivera, L.; Tuboi, S.; Garbes, P.; Borkowski, A.; Wallace, D. Safety and immunogenicity of one versus two doses of Takeda's tetravalent dengue vaccine in children in Asia and Latin America: Interim results from a phase 2, randomised, placebo-controlled study. Lancet Infect. Dis. 2017, 17, 615–625. [Google Scholar] [CrossRef]
- Bauer, K.; Esquilin, I.O.; Cornier, A.S.; Thomas, S.J.; del Rio, A.I.Q.; Bertran-Pasarell, J.; Ramirez, J.O.M.; Diaz, C.; Carlo, S.; Eckels, K.H.; et al. A Phase II, Randomized, Safety and Immunogenicity Trial of a Re-Derived, Live-Attenuated Dengue Virus Vaccine in Healthy Children and Adults Living in Puerto Rico. Am. J. Trop. Med. Hyg. 2015, 93, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.C.; Lin, L.; Martinez, L.J.; Ruck, R.C.; Eckels, K.H.; Collard, A.; De La Barrera, R.; Paolino, K.M.; Toussaint, J.-F.; Lepine, E.; et al. Phase 1 Randomized Study of a Tetravalent Dengue Purified Inactivated Vaccine in Healthy Adults in the United States. Am. J. Trop. Med. Hyg. 2017, 96, 1325–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannice, K.S.; Durbin, A.; Hombach, J. Status of vaccine research and development of vaccines for dengue. Vaccine 2016, 34, 2934–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadinegoro, S.R.; Arredondo-García, J.L.; Capeding, M.R.; Deseda, C.; Chotpitayasunondh, T.; Dietze, R.; Ismail, H.H.M.; Reynales, H.; Limkittikul, K.; Rivera-Medina, D.M.; et al. Efficacy and Long-Term Safety of a Dengue Vaccine in Regions of Endemic Disease. N. Engl. J. Med. 2015, 373, 1195–1206. [Google Scholar] [CrossRef] [Green Version]
- Villar, L.; Dayan, G.H.; Arredondo-García, J.L.; Rivera, D.M.; Cunha, R.; Deseda, C.; Reynales, H.; Costa, M.S.; Morales-Ramírez, J.O.; Carrasquilla, G.; et al. Efficacy of a Tetravalent Dengue Vaccine in Children in Latin America. N. Engl. J. Med. 2015, 372, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, B.D.; Durbin, A.P.; Pierce, K.K.; Carmolli, M.P.; Tibery, C.M.; Grier, P.L.; Hynes, N.; Diehl, S.A.; Elwood, D.; Jarvis, A.P.; et al. Robust and Balanced Immune Responses to All 4 Dengue Virus Serotypes Following Administration of a Single Dose of a Live Attenuated Tetravalent Dengue Vaccine to Healthy, Flavivirus-Naive Adults. J. Infect. Dis. 2015, 212, 702–710. [Google Scholar] [CrossRef]
- Sirivichayakul, C.; Barranco-Santana, E.A.; Esquilin-Rivera, I.; Oh, H.M.L.; Raanan, M.; Sariol, C.A.; Shek, L.; Simasathien, S.; Smith, M.K.; Velez, I.D.; et al. Safety and Immunogenicity of a Tetravalent Dengue Vaccine Candidate in Healthy Children and Adults in Dengue-Endemic Regions: A Randomized, Placebo-Controlled Phase 2 Study. J. Infect. Dis. 2015, 213, 1562–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.J.; Eckels, K.H.; Carletti, I.; De La Barrera, R.; Dessy, F.; Fernandez, S.; Putnak, R.; Toussaint, J.-F.; Sun, W.; Bauer, K.; et al. A Phase II, Randomized, Safety and Immunogenicity Study of a Re-Derived, Live-Attenuated Dengue Virus Vaccine in Healthy Adults. Am. J. Trop. Med. Hyg. 2013, 88, 73–88. [Google Scholar] [CrossRef]
- Diaz, C.; Koren, M.; Lin, L.; Martinez, L.J.; Eckels, K.H.; Campos, M.; Jarman, R.G.; De La Barrera, R.; Lepine, E.; Febo, I.; et al. Safety and Immunogenicity of Different Formulations of a Tetravalent Dengue Purified Inactivated Vaccine in Healthy Adults from Puerto Rico: Final Results after 3 Years of Follow-Up from a Randomized, Placebo-Controlled Phase I Study. Am. J. Trop. Med. Hyg. 2020, 102, 951–954. [Google Scholar] [CrossRef] [Green Version]
- Coller, B.-A.G.; Clements, D.E.; Bett, A.J.; Sagar, S.L.; Ter Meulen, J.H. The development of recombinant subunit envelope-based vaccines to protect against dengue virus induced disease. Vaccine 2011, 29, 7267–7275. [Google Scholar] [CrossRef] [Green Version]
- Manoff, S.B.; Sausser, M.; Russell, A.F.; Martin, J.; Radley, D.; Hyatt, D.; Roberts, C.; Lickliter, J.; Krishnarajah, J.; Bett, A.; et al. Immunogenicity and safety of an investigational tetravalent recombinant subunit vaccine for dengue: Results of a Phase I randomized clinical trial in flavivirus-naïve adults. Hum. Vaccines Immunother. 2019, 15, 2195–2204. [Google Scholar] [CrossRef]
- Beckett, C.G.; Tjaden, J.; Burgess, T.; Danko, J.R.; Tamminga, C.; Simmons, M.; Wu, S.-J.; Sun, P.; Kochel, T.; Raviprakash, K.; et al. Evaluation of a prototype dengue-1 DNA vaccine in a Phase 1 clinical trial. Vaccine 2011, 29, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Danko, J.R.; Kochel, T.; Teneza-Mora, N.; Luke, T.C.; Raviprakash, K.; Sun, P.; Simmons, M.; Moon, J.E.; De La Barrera, R.; Martinez, L.J.; et al. Safety and Immunogenicity of a Tetravalent Dengue DNA Vaccine Administered with a Cationic Lipid-Based Adjuvant in a Phase 1 Clinical Trial. Am. J. Trop. Med. Hyg. 2018, 98, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Kochel, T.; Raviprakash, K.; Hayes, C.; Watts, D.; Russell, K.; Gozalo, A.; Phillips, I.; Ewing, D.; Murphy, G.; Porter, K. A dengue virus serotype-1 DNA vaccine induces virus neutralizing antibodies and provides protection from viral challenge in Aotus monkeys. Vaccine 2000, 18, 3166–3173. [Google Scholar] [CrossRef]
- Nivarthi, U.K.; Swanstrom, J.; Delacruz, M.J.; Patel, B.; Durbin, A.P.; Whitehead, S.S.; Kirkpatrick, B.D.; Pierce, K.K.; Diehl, S.A.; Katzelnick, L.; et al. A tetravalent live attenuated dengue virus vaccine stimulates balanced immunity to multiple serotypes in humans. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Capeding, M.R.; Tran, N.H.; Hadinegoro, S.R.S.; Ismail, H.I.H.M.; Chotpitayasunondh, T.; Chua, M.N.; Luong, C.Q.; Rusmil, K.; Wirawan, D.N.; Nallusamy, R.; et al. Clinical efficacy and safety of a novel tetravalent dengue vaccine in healthy children in Asia: A phase 3, randomised, observer-masked, placebo-controlled trial. Lancet 2014, 384, 1358–1365. [Google Scholar] [CrossRef]
- Sabchareon, A.; Wallace, D.; Sirivichayakul, C.; Limkittikul, K.; Chanthavanich, P.; Suvannadabba, S.; Jiwariyavej, V.; Dulyachai, W.; Pengsaa, K.; Wartel, T.A.; et al. Protective efficacy of the recombinant, live-attenuated, CYD tetravalent dengue vaccine in Thai schoolchildren: A randomised, controlled phase 2b trial. Lancet 2012, 380, 1559–1567. [Google Scholar] [CrossRef]
- Tully, D.; Griffiths, C.L. Dengvaxia: The world’s first vaccine for prevention of secondary dengue. Ther. Adv. Vaccines Immunother. 2021, 9. [Google Scholar] [CrossRef]
- FDA. Dengvaxia FDA 2019. Available online: https://www.fda.gov/vaccines-blood-biologics/dengvaxia (accessed on 26 October 2021).
- Guy, B.; Jackson, N. Dengue vaccine: Hypotheses to understand CYD-TDV-induced protection. Nat. Rev. Genet. 2016, 14, 45–54. [Google Scholar] [CrossRef]
- Shukla, R.; Ramasamy, V.; Shanmugam, R.K.; Ahuja, R.; Khanna, N. Antibody-Dependent Enhancement: A Challenge for Developing a Safe Dengue Vaccine. Front. Cell. Infect. Microbiol. 2020, 10, 572681. [Google Scholar] [CrossRef]
- Halstead, S.B.; Katzelnick, L.C.; Russell, P.K.; Markoff, L.; Aguiar, M.; Dans, L.R.; Dans, A.L. Ethics of a partially effective dengue vaccine: Lessons from the Philippines. Vaccine 2020, 38, 5572–5576. [Google Scholar] [CrossRef]
- Swaminathan, S.; Khanna, N. Dengue vaccine development: Global and Indian scenarios. Int. J. Infect. Dis. 2019, 84, S80–S86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, L.J.; Young, E.F.; Stoops, M.J.; Henein, S.R.; Adams, E.C.; Baric, R.S.; de Silva, A.M. Defining levels of dengue virus serotype-specific neutralizing antibodies induced by a live attenuated tetravalent dengue vaccine (TAK-003). PLoS Negl. Trop. Dis. 2021, 15, e0009258. [Google Scholar] [CrossRef] [PubMed]
- Rivino, L.; Lim, M.Q. CD4+and CD8+T-cell immunity to Dengue—Lessons for the study of Zika virus. Immunology 2017, 150, 146–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norshidah, H.; Vignesh, R.; Lai, N.S. Updates on Dengue Vaccine and Antiviral: Where Are We Heading? Molecules 2021, 26, 6768. [Google Scholar] [CrossRef] [PubMed]
- Watanaveeradej, V.; Tournay, E.; Kerdpanich, A.; Eckels, K.H.; Dessy, F.; Innis, B.; De La Barrerra, R.; Simasathien, S.; Toussaint, J.-F.; Jarman, R.G.; et al. Safety and Immunogenicity of a Rederived, Live-Attenuated Dengue Virus Vaccine in Healthy Adults Living in Thailand: A Randomized Trial. Am. J. Trop. Med. Hyg. 2014, 91, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Torresi, J.; Ebert, G.; Pellegrini, M. Vaccines licensed and in clinical trials for the prevention of dengue. Hum. Vaccines Immunother. 2017, 13, 1059–1072. [Google Scholar] [CrossRef]
- Raviprakash, K.; Apt, D.; Brinkman, A.; Skinner, C.; Yang, S.; Dawes, G.; Ewing, D.; Wu, S.-J.; Bass, S.; Punnonen, J.; et al. A chimeric tetravalent dengue DNA vaccine elicits neutralizing antibody to all four virus serotypes in rhesus macaques. Virology 2006, 353, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Raviprakash, K. Immunogenicity of dengue virus type 1 DNA vaccines expressing truncated and full length envelope protein. Vaccine 2000, 18, 2426–2434. [Google Scholar] [CrossRef]
- Kochel, T.; Wu, S.-J.; Raviprakash, K.; Hobart, P.; Hoffman, S.; Porter, K.; Hayes, C. Inoculation of plasmids expressing the dengue-2 envelope gene elicit neutralizing antibodies in mice. Vaccine 1997, 15, 547–552. [Google Scholar] [CrossRef]
- Porter, K.R.; Ewing, D.; Chen, L.; Wu, S.-J.; Hayes, C.G.; Ferrari, M.; Teneza-Mora, N.; Raviprakash, K. Immunogenicity and protective efficacy of a vaxfectin-adjuvanted tetravalent dengue DNA vaccine. Vaccine 2012, 30, 336–341. [Google Scholar] [CrossRef]
- Blair, P.J.; Kochel, T.J.; Raviprakash, K.; Guevara, C.; Salazar, M.; Wu, S.-J.; Olson, J.G.; Porter, K.R. Evaluation of immunity and protective efficacy of a dengue-3 premembrane and envelope DNA vaccine in Aotus nancymae monkeys. Vaccine 2006, 24, 1427–1432. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y. Dengue 2 PreM-E/LAMP chimera targeted to the MHC class II compartment elicits long-lasting neutralizing antibodies. Vaccine 2003, 21, 2178–2189. [Google Scholar] [CrossRef]
- Raviprakash, K.; Marques, E.; Ewing, D.; Lu, Y.; Phillips, I.; Porter, K.R.; Kochel, T.J.; August, T.J.; Hayes, C.G.; Murphy, G.S. Synergistic Neutralizing Antibody Response to a Dengue Virus Type 2 DNA Vaccine by Incorporation of Lysosome-Associated Membrane Protein Sequences and Use of Plasmid Expressing GM-CSF. Virology 2001, 290, 74–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durbin, A.P.; Pierce, K.K.; Kirkpatrick, B.D.; Grier, P.; Sabundayo, B.P.; He, H.; Sausser, M.; Russell, A.F.; Martin, J.; Hyatt, D.; et al. Immunogenicity and Safety of a Tetravalent Recombinant Subunit Dengue Vaccine in Adults Previously Vaccinated with a Live Attenuated Tetravalent Dengue Vaccine: Results of a Phase-I Randomized Clinical Trial. Am. J. Trop. Med. Hyg. 2020, 103, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, D.; Meschino, S.; Guan, L.; Clements, D.E.; ter Meulen, J.H.; Casimiro, D.R.; Coller, B.-A.G.; Bett, A.J. Preclinical development of a dengue tetravalent recombinant subunit vaccine: Immunogenicity and protective efficacy in nonhuman primates. Vaccine 2015, 33, 4105–4116. [Google Scholar] [CrossRef]
- McArthur, M.A.; Sztein, M.B.; Edelman, R. Dengue vaccines: Recent developments, ongoing challenges and current candidates. Expert Rev. Vaccines 2013, 12, 933–953. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, S.; Thomas, S.J.; De La Barrera, R.; Im-Erbsin, R.; Jarman, R.G.; Baras, B.; Toussaint, J.-F.; Mossman, S.; Innis, B.; Schmidt, A.; et al. An Adjuvanted, Tetravalent Dengue Virus Purified Inactivated Vaccine Candidate Induces Long-Lasting and Protective Antibody Responses Against Dengue Challenge in Rhesus Macaques. Am. J. Trop. Med. Hyg. 2015, 92, 698–708. [Google Scholar] [CrossRef]
- Da Silveira, L.T.C.; Tura, B.; Santos, M. Systematic review of dengue vaccine efficacy. BMC Infect. Dis. 2019, 19, 1–8. [Google Scholar] [CrossRef]
- Henein, S.; Swanstrom, J.; Byers, A.M.; Moser, J.M.; Shaik, S.F.; Bonaparte, M.; Jackson, N.; Guy, B.; Baric, R.; de Silva, A.M. Dissecting Antibodies Induced by a Chimeric Yellow Feverâ Dengue, Live-Attenuated, Tetravalent Dengue Vaccine (CYD-TDV) in Naive and Dengue-Exposed Individuals. J. Infect. Dis. 2017, 215, 351–358. [Google Scholar]
- Turner, M.; Papadimitriou, A.; Winkle, P.; Segall, N.; Levin, M.; Doust, M.; Johnson, C.; Lucksinger, G.; Fierro, C.; Pickrell, P.; et al. Immunogenicity and safety of lyophilized and liquid dengue tetravalent vaccine candidate formulations in healthy adults: A randomized, phase 2 clinical trial. Hum. Vaccines Immunother. 2020, 16, 2456–2464. [Google Scholar] [CrossRef] [Green Version]
- Biswal, S.; Reynales, H.; Saez-Llorens, X.; Lopez, P.; Borja-Tabora, C.; Kosalaraksa, P.; Sirivichayakul, C.; Watanaveeradej, V.; Rivera, L.; Espinoza, F.; et al. Efficacy of a Tetravalent Dengue Vaccine in Healthy Children and Adolescents. N. Engl. J. Med. 2019, 381, 2009–2019. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.E.; Wallace, D.; Stinchcomb, D.T. A recombinant, chimeric tetravalent dengue vaccine candidate based on a dengue virus serotype 2 backbone. Expert Rev. Vaccines 2016, 15, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, S.S.; Falgout, B.; Hanley, K.A.; Blaney, J.E., Jr.; Markoff, L.; Murphy, B.R. A live, attenuated dengue virus type 1 vaccine candidate with a 30-nucleotide deletion in the 3′ untranslated region is highly attenuated and immunogenic in monkeys. J. Virol. 2003, 77, 1653–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; A Koren, M.; Paolino, K.M.; Eckels, K.H.; De La Barrera, R.; Friberg, H.; Currier, J.R.; Gromowski, G.D.; E Aronson, N.; Keiser, P.B.; et al. Immunogenicity of a Live-Attenuated Dengue Vaccine Using a Heterologous Prime-Boost Strategy in a Phase 1 Randomized Clinical Trial. J. Infect. Dis. 2020, 223, 1707–1716. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro-Michelsen, J.R.; Souza, R.D.S.O.; Santana, I.V.R.; Da Silva, P.D.S.; Mendez, E.C.; Luiz, W.B.; Amorim, J.H. Anti-dengue vaccines: From development to clinical trials. Front. Immunol. 2020, 11, 1252. [Google Scholar] [CrossRef]
- Guy, B.; Charnay, C.; Burdin, N.; Lang, J.; Aguirre, M.; Mantel, N.; Ramirez, L.; Dumas, R.; Pontvianne, J.; Gulia, S.; et al. Evaluation of Interferences between Dengue Vaccine Serotypes in a Monkey Model. Am. J. Trop. Med. Hyg. 2009, 80, 302–311. [Google Scholar] [CrossRef] [Green Version]
- Wilder-Smith, A. The first licensed dengue vaccine: Can it be used in travelers? Curr. Opin. Infect. Dis. 2019, 32, 394–400. [Google Scholar] [CrossRef]
- World Health Organization. Dengue Vaccine: WHO Position Paper, September 2018—Recommendations. Vaccine 2019, 37, 4848–4849. [Google Scholar] [CrossRef]
- World Health Organization. Position Paper on Dengue Vaccines. Wkly. Epidemiol. Rec. 2018, 93, 457–476. Available online: https://reliefweb.int/report/world/weekly-epidemiological-record-wer-7-september-2018-vol-93-no-36-pp-457-476-enfr (accessed on 5 October 2022).






| Clinical Sample | Approach | Technique | Time Required | |
|---|---|---|---|---|
| Detection of virus and viral product | Acute serum (1–5 days after onset of fever) and necropsy tissue |
| Mosquito or mosquito cell culture | 1 week or more |
| RT-PCR and real-time RT-PCR | 1–2 days | |||
| NS1 antigen rapid test | Minutes | |||
| NS1 antigen-capture ELISA | 1 day | |||
| Immunohistochemistry | 2–5 days | |||
| Serological response | Paired sera
|
| ELISA, HI | 1–2 days |
| Plaque reduction neutralization test | >7 days | |||
| MAC-ELISA | 1–2 days | |||
| Immunoglobulin M rapid tests (LFA) | Minutes | |||
| Immunoglobulin G ELISA, HI | 1–2 days | |||
| Immunoglobulin G rapid tests (LFA) | Minutes |
| Vaccine Candidate, Developer, and Status | Approach and Type of Vaccine | Status | References |
|---|---|---|---|
| Dengvaxia ® (CYD-TDV); Sanofi Pasteur; Licensed | YF-17D backbone with prM and E genes of dengue virus 1–4 (Live-attenuated vaccine) | Age limit; increased risk of severe dengue in seronegative subjects but high effectiveness and safe in seropositive individuals | [87,94] |
| TetraVax-DV-V003/TV005; NIAID/NIH; Phase III | Full-length dengue virus 1/2/3/4 lacking 30 nucleotides in 3′ UTR (Live-attenuated vaccine) | Well tolerated; balanced immune response in subjects, effective with administration of a single dose; adverse reaction (mild rash) | [88,89,95] |
| DENVax/TAK003/TDV; Takeda/Inviragen; Phase III | Dengue virus 2 PDK53 backbone with dengue virus 1,3, and 4 prM and E gene chimera (Live-attenuated vaccine) | Immunogenic and well tolerated in multiple phase I and II clinical studies, independent of the participant’s age or serostatus, safety profile not entirely known | [90,96] |
| TDEN LAV; WRAIR/GSK; Phase II (Discontinued) | Dengue virus 1/2/3/4 PDK virus (Live-attenuated vaccine) | Proven to be a safe, well-tolerated, and immunogenic DENV vaccine candidate in phase II trial | [91,97,98] |
| TDEN PIV; WRAIR/GSK/Fiocruz; Phase I | Formalin inactivated dengue virus 1/2/3/4 (Purified inactivated vaccine) | Well tolerated, immunogenic in naïve and seropositive individuals. No risk of re-activation and good immunological balance | [92,98] |
| V180; WRAIR/GSK/Fiocruz; Phase I/II | DEN-80E-containing recombinant truncated protein (Recombinant subunit) | Induced steady immune response against all DENV serotypes, decreasing the likelihood of the ADE effect | [99,100] |
| TVDV; US Naval Medical Research Centre; Phase I | prM and E proteins Nucleic acid (DNA) | Stable but lack of immunogenicity, Plasmid modification required | [101,102] |
| D1ME100; US Naval Medical Research Centre; Phase I | Recombinant plasmid vector encoding prM/E Nucleic acid (DNA) | No neutralizing antibody, Response detected in individuals with low-dose immunization | [103] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeyaullah, M.; Muzammil, K.; AlShahrani, A.M.; Khan, N.; Ahmad, I.; Alam, M.S.; Ahmad, R.; Khan, W.H. Preparedness for the Dengue Epidemic: Vaccine as a Viable Approach. Vaccines 2022, 10, 1940. https://doi.org/10.3390/vaccines10111940
Zeyaullah M, Muzammil K, AlShahrani AM, Khan N, Ahmad I, Alam MS, Ahmad R, Khan WH. Preparedness for the Dengue Epidemic: Vaccine as a Viable Approach. Vaccines. 2022; 10(11):1940. https://doi.org/10.3390/vaccines10111940
Chicago/Turabian StyleZeyaullah, Md., Khursheed Muzammil, Abdullah M. AlShahrani, Nida Khan, Irfan Ahmad, Md. Shane Alam, Razi Ahmad, and Wajihul H. Khan. 2022. "Preparedness for the Dengue Epidemic: Vaccine as a Viable Approach" Vaccines 10, no. 11: 1940. https://doi.org/10.3390/vaccines10111940