
Interactions of Microbiota and Mucosal Immunity in the Ceca of Broiler Chickens Infected with Eimeria tenella


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sampling
2.2. DNA Extraction and Microbiome Analyses in the Cecal Content
2.3. Determination of Secretory Immunoglobulin A (sIgA) and Protein Concentration in the Cecal Contents
2.4. RNA Extraction and Real-Time Reverse Transcription (RT)-PCR Analysis for Analyzing Relative mRNA Abundance of Antimicrobial Peptides, Alkaline Phosphatase, Mucin Gene, Inflammatory Cytokines, and Toll-Like Receptors
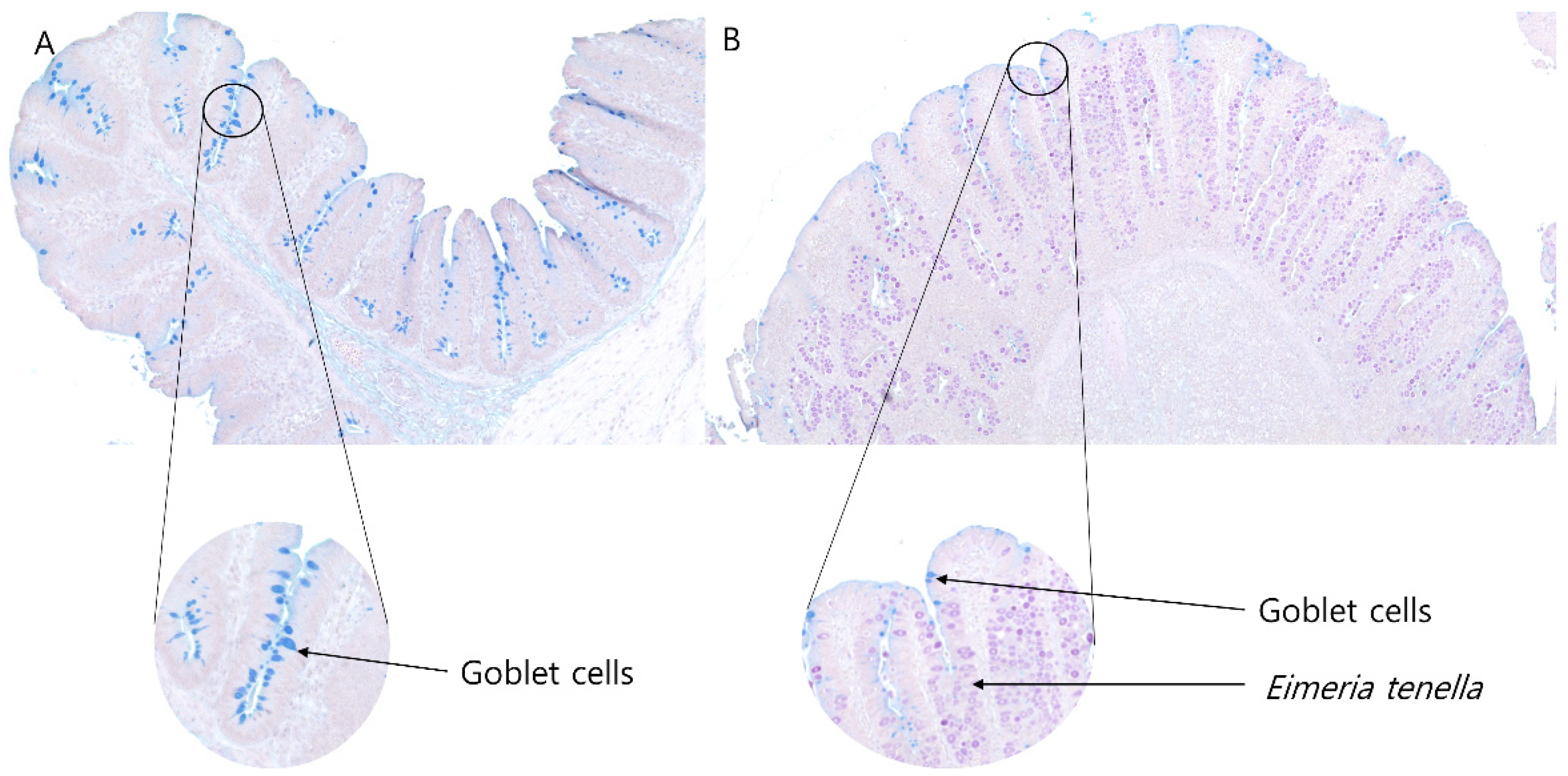
2.5. Quantification of Goblet Cells by Using Alcian Blue/Period Acid-Schiff (AB/PAS) Staining
2.6. Alkaline Phosphatase Activities in the Cecal Tissue and Serum and Serum Endotoxin Levels
2.7. Statistical Analyses
3. Results
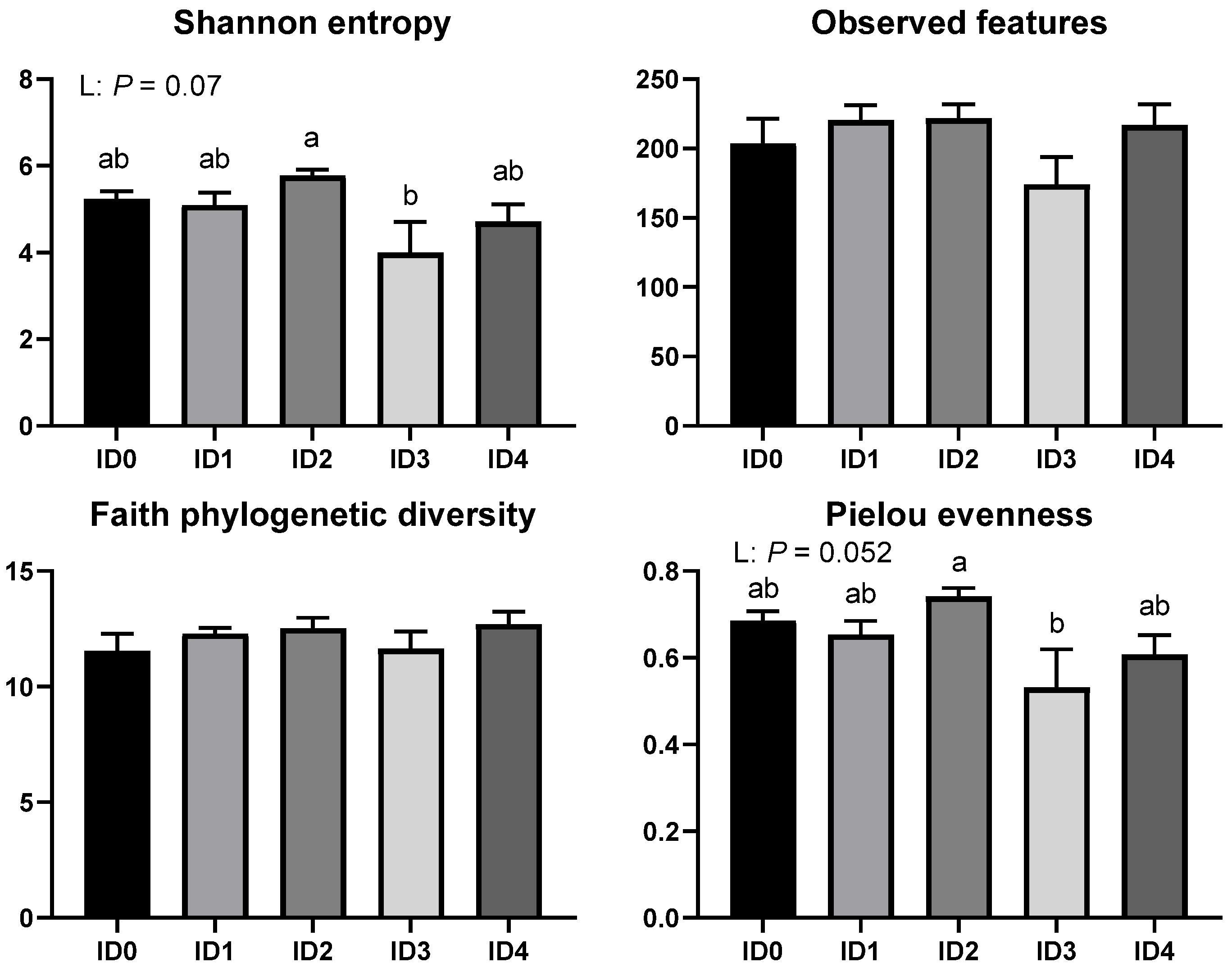
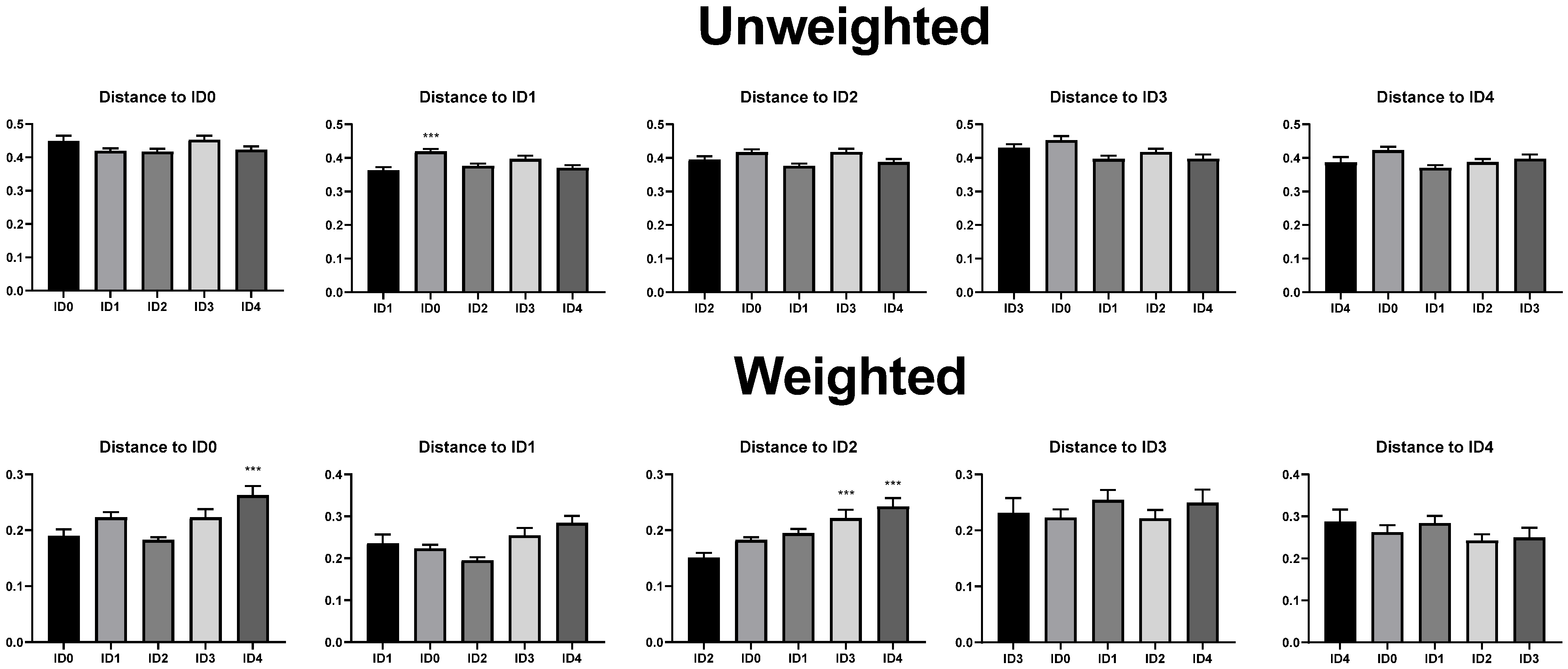
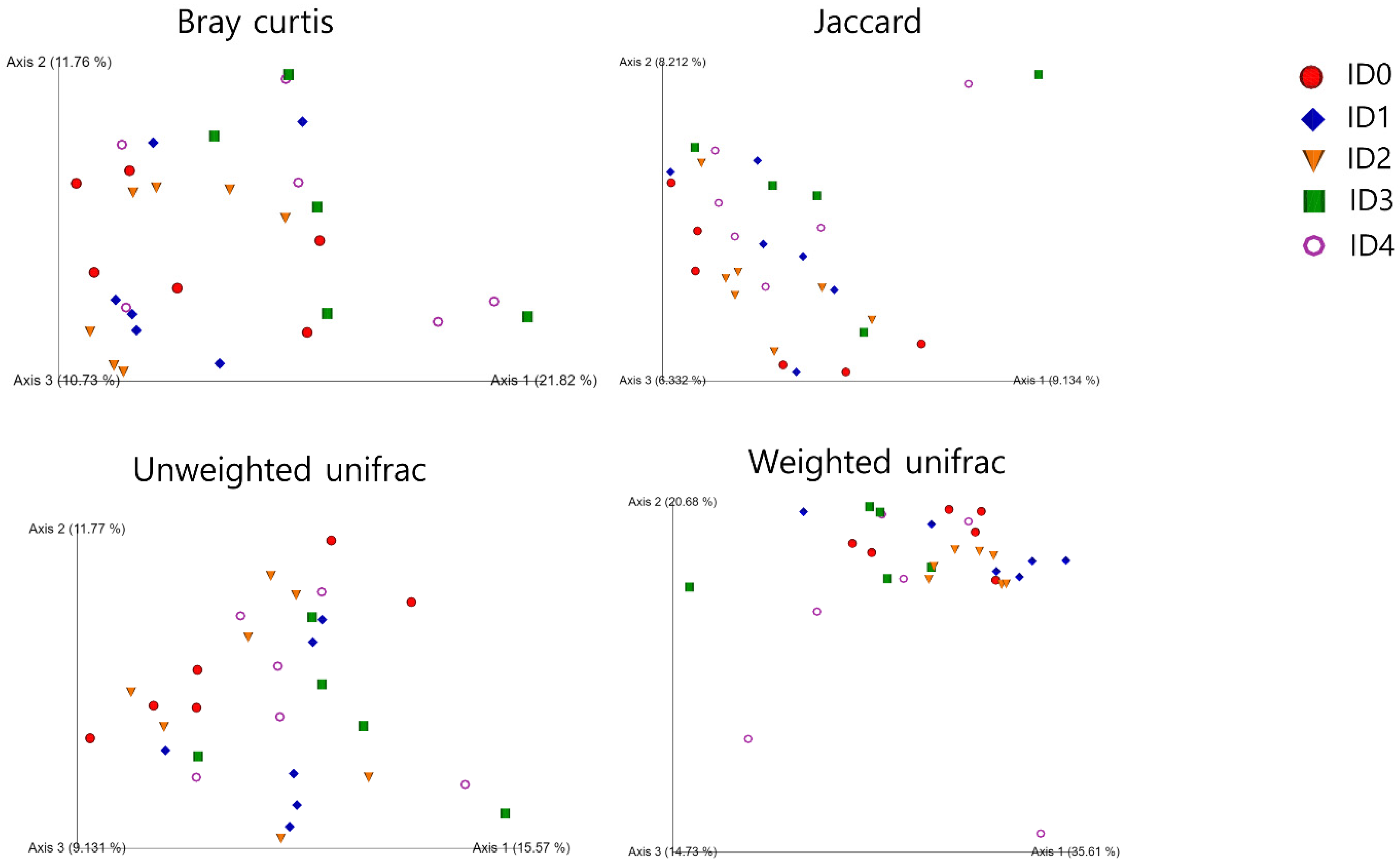
3.1. Alpha and Beta Diversity of the Cecal Microbial Communities
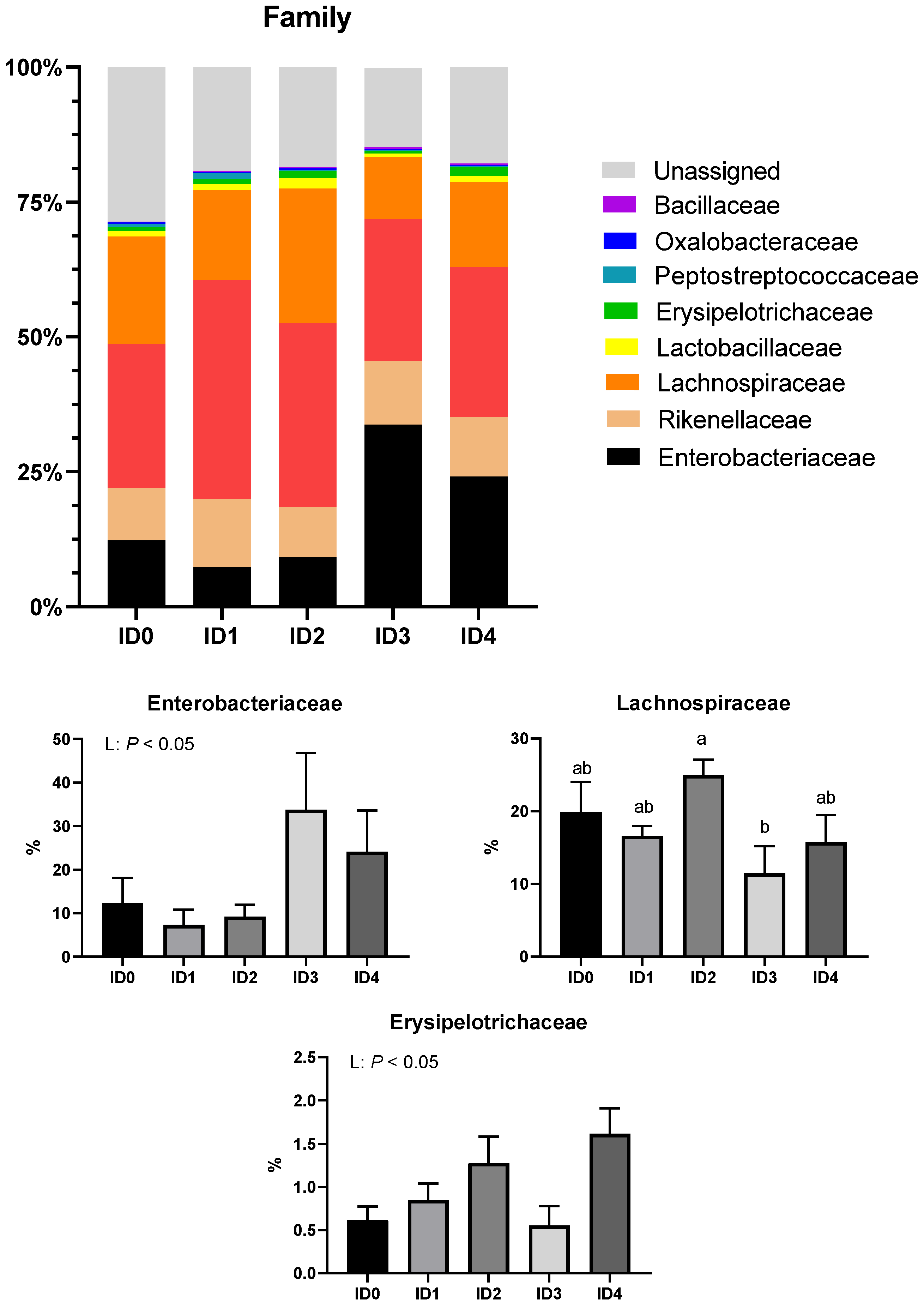
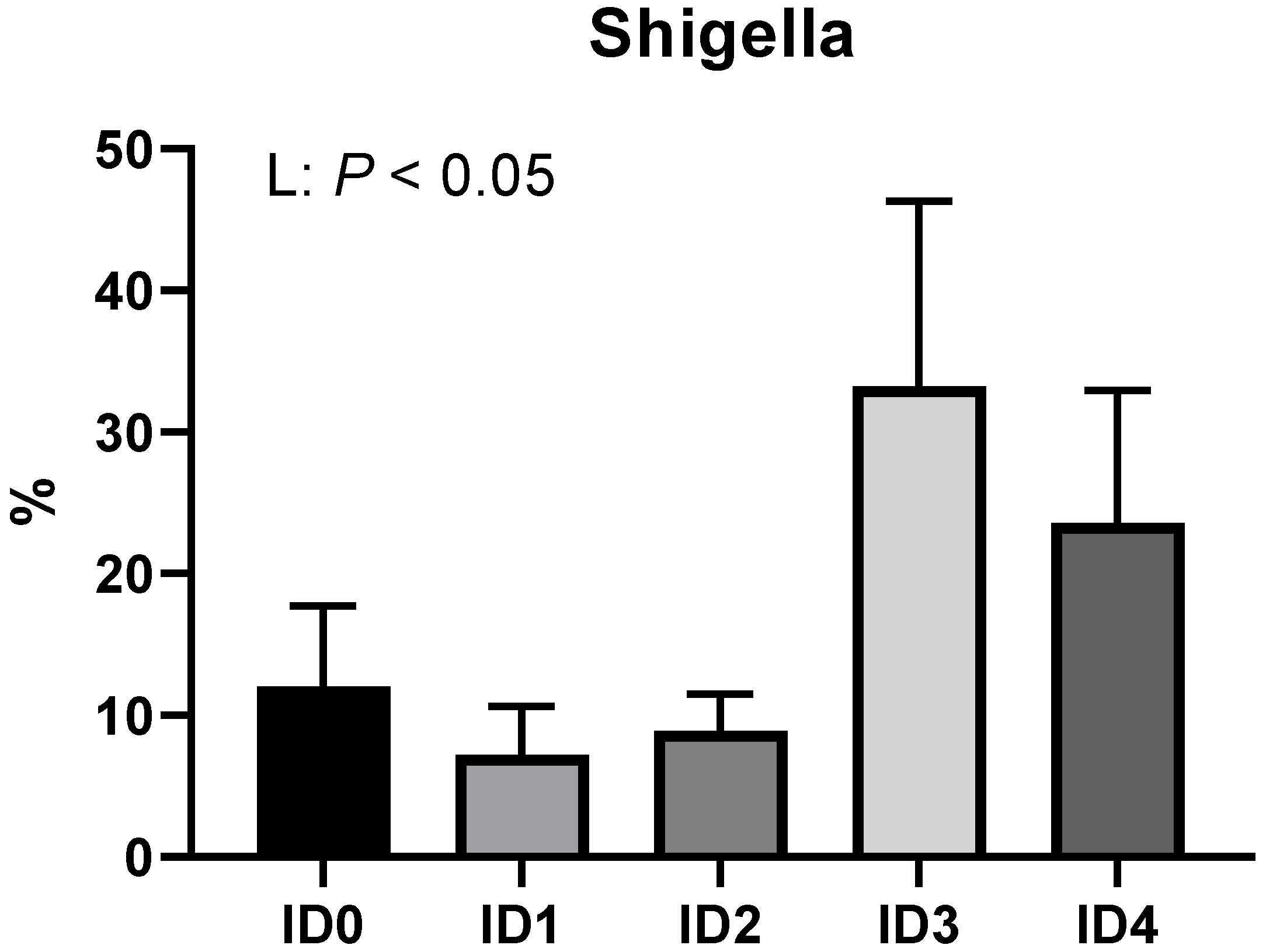
3.2. Phylum- and Family-Level Composition of the Cecal Microbial Communities
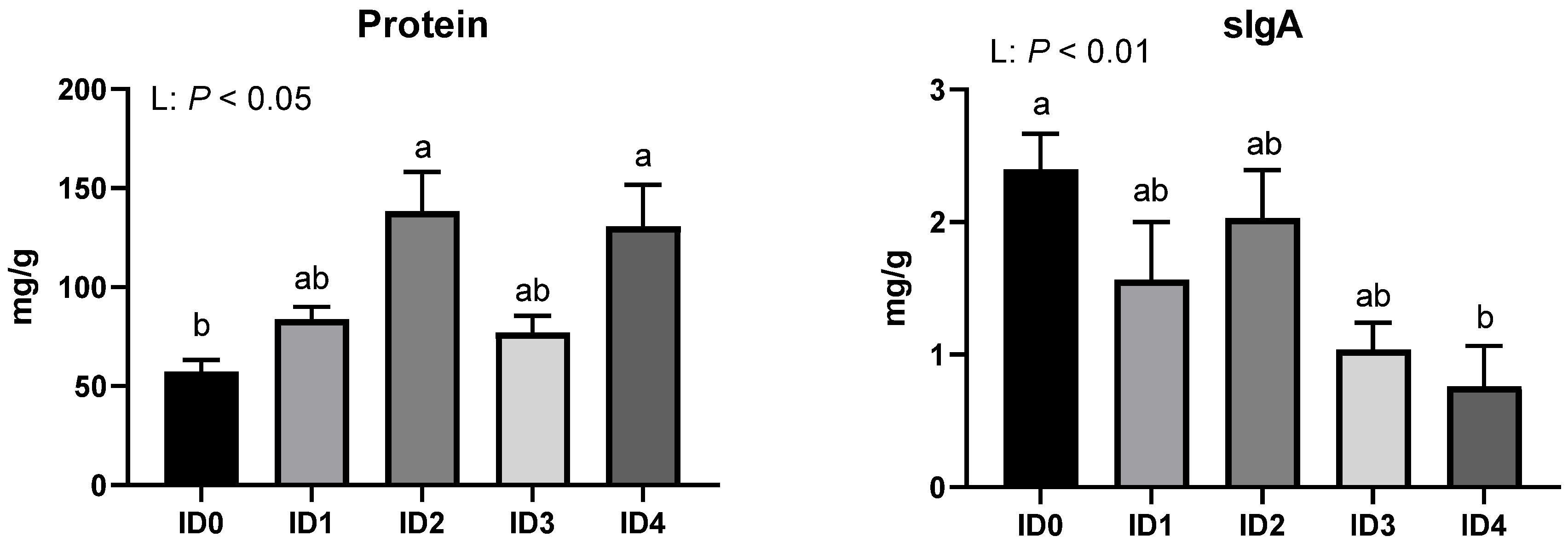
3.3. Concentration of Protein and Secretory Immunoglobulin A (sIgA) in the Cecal Contents
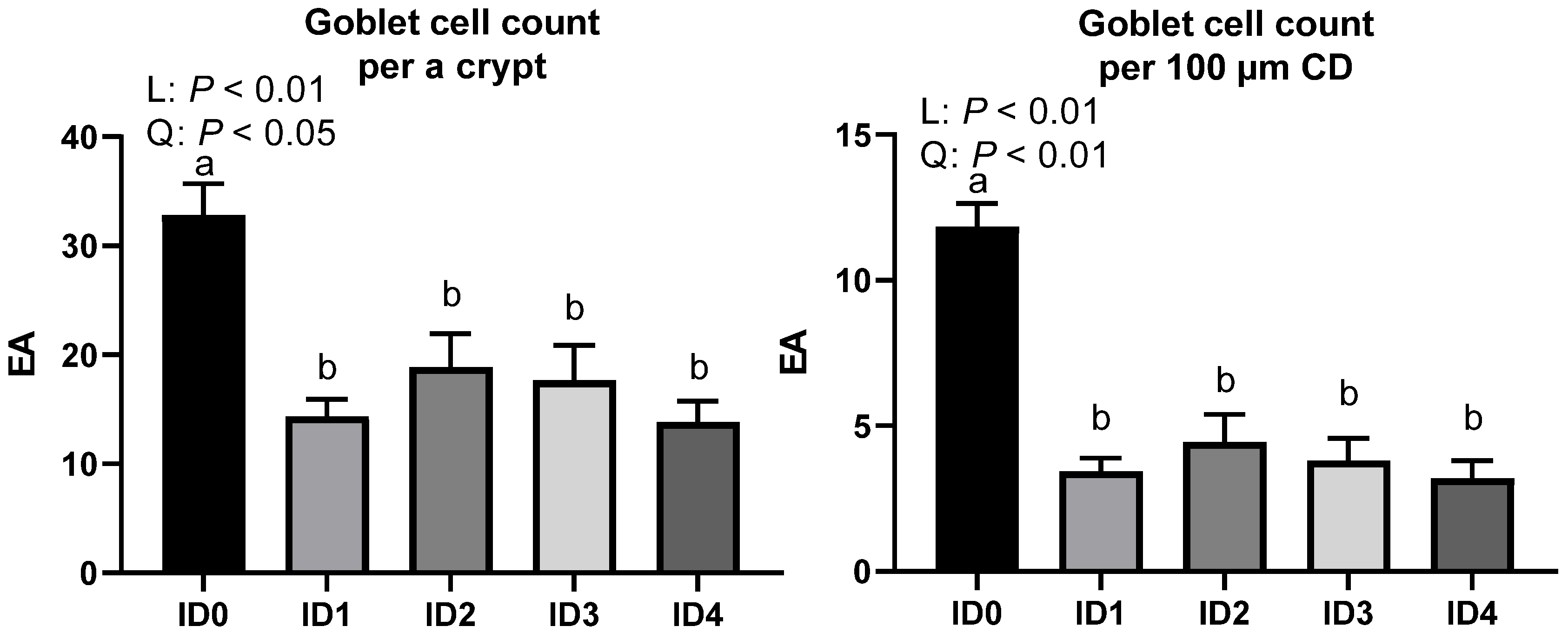
3.4. Goblet Cell Number per a Crypt and Goblet Cell Density in the Ceca
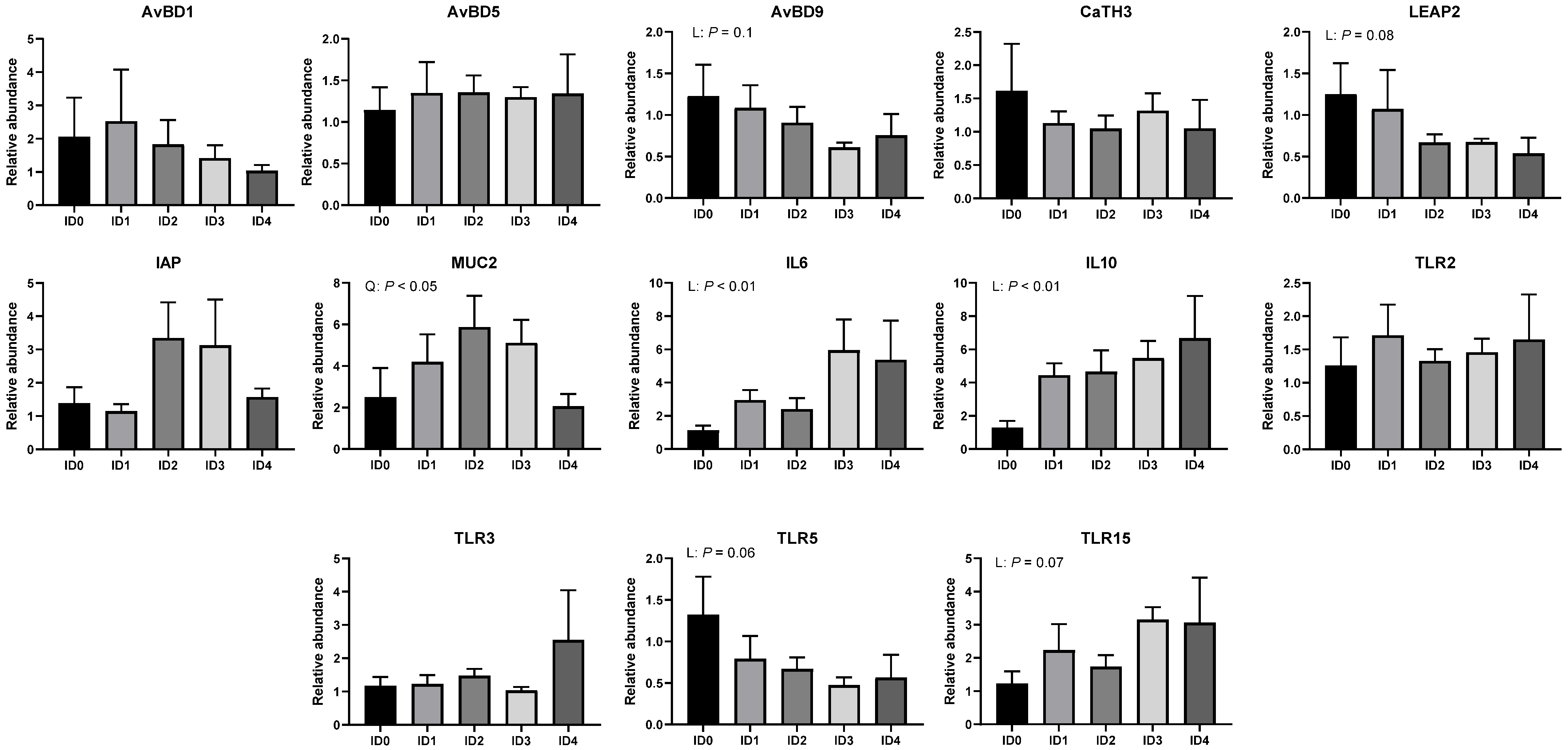
3.5. Relative mRNA Expression
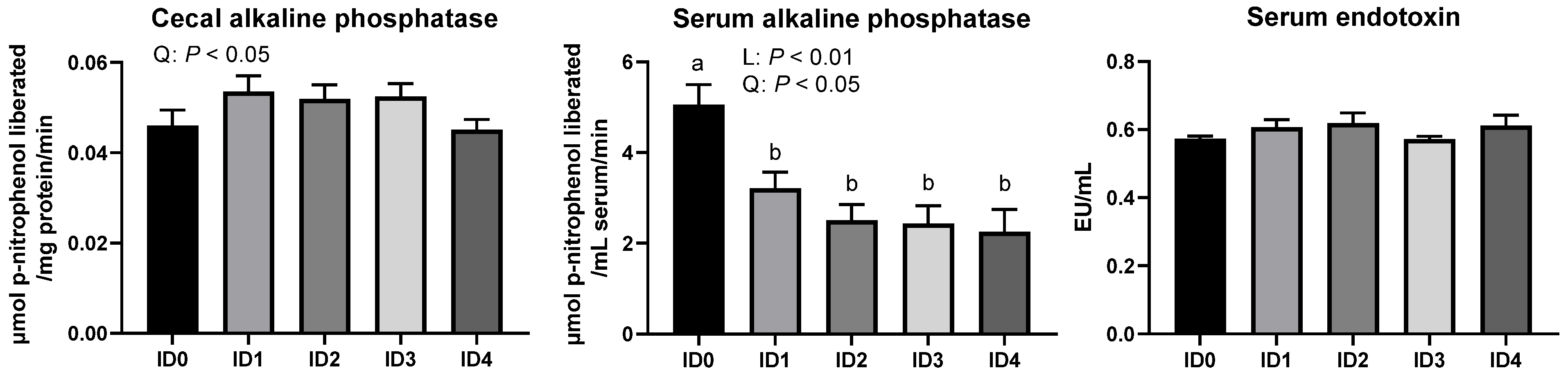
3.6. Alkaline Phosphatase Activities in the Cecal Tissue and Serum and Concentration of Serum Endotoxins in Broilers
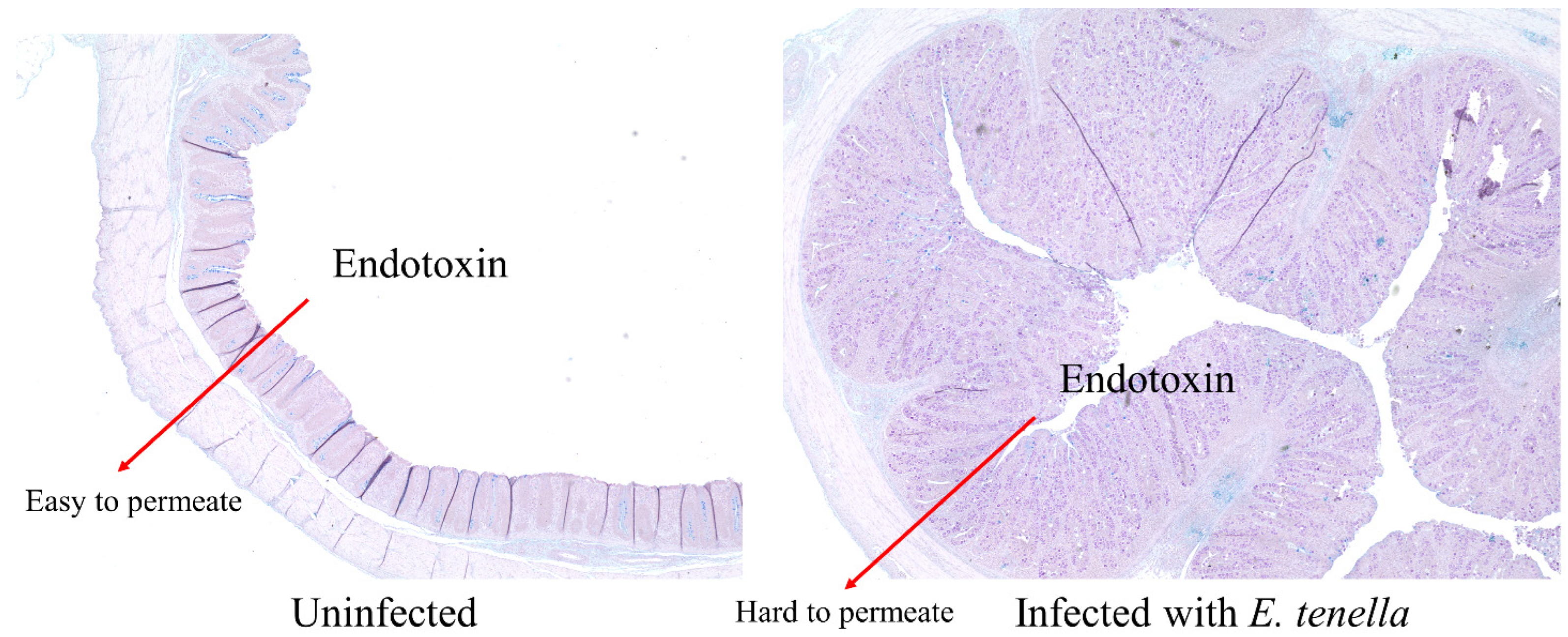
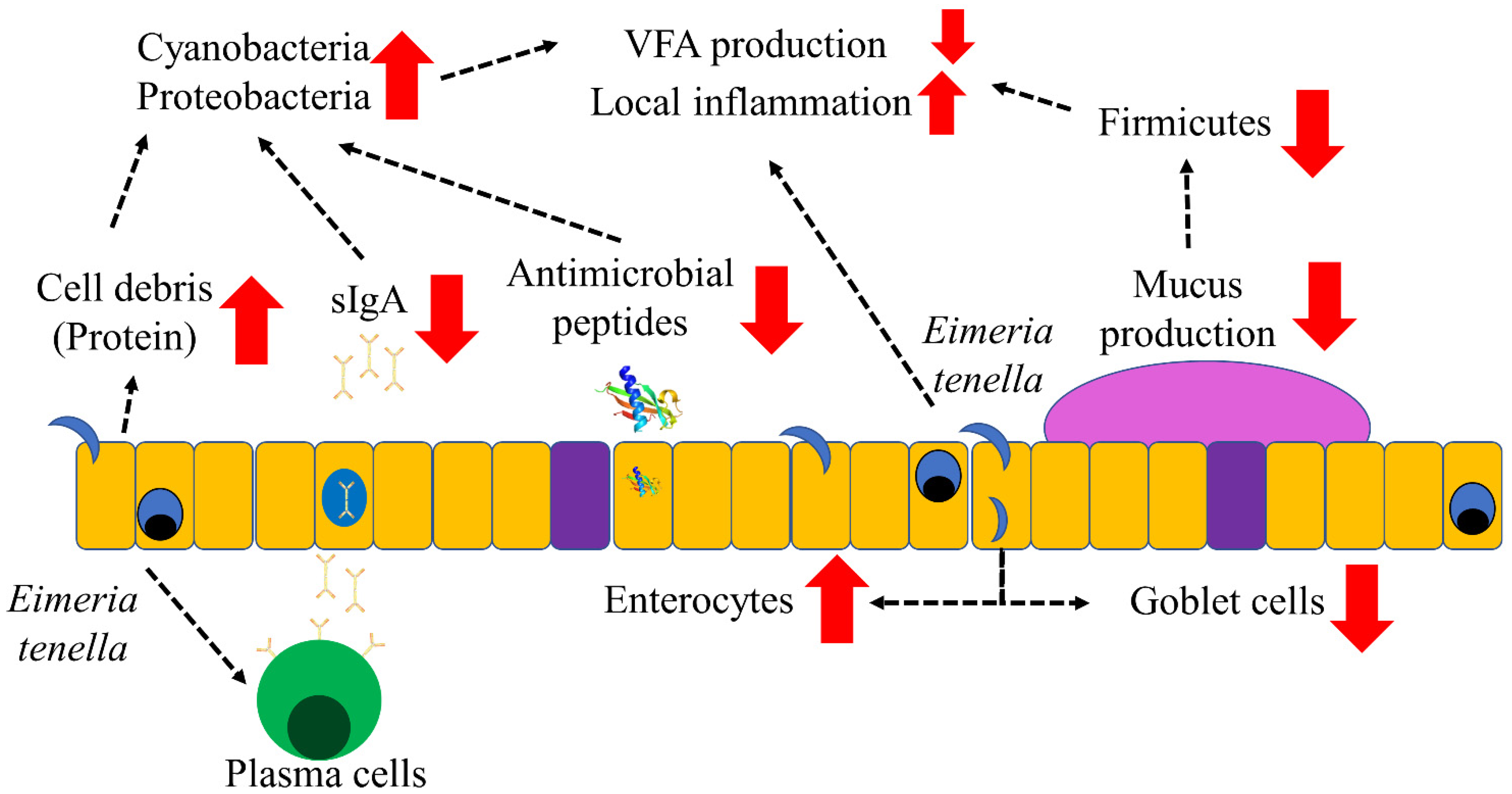
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D. Re-calculating the cost of coccidiosis in chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef] [PubMed]
- Teng, P.-Y.; Choi, J.; Tompkins, Y.; Lillehoj, H.; Kim, W. Impacts of increasing challenge with Eimeria maxima on the growth performance and gene expression of biomarkers associated with intestinal integrity and nutrient transporters. Vet. Res. 2021, 52, 81. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, W.K. Dietary application of tannins as a potential mitigation strategy for current challenges in poultry production: A review. Animals 2020, 10, 2389. [Google Scholar] [CrossRef]
- Williams, R. Epidemiological aspects of the use of live anticoccidial vaccines for chickens. Int. J. Parasitol. 1998, 28, 1089–1098. [Google Scholar] [CrossRef]
- Huang, G.; Tang, X.; Bi, F.; Hao, Z.; Han, Z.; Suo, J.; Zhang, S.; Wang, S.; Duan, C.; Yu, Z. Eimeria tenella infection perturbs the chicken gut microbiota from the onset of oocyst shedding. Vet. Parasitol. 2018, 258, 30–37. [Google Scholar] [CrossRef]
- Choi, J.; Ko, H.; Tompkins, Y.H.; Teng, P.-Y.; Lourenco, J.M.; Callaway, T.R.; Kim, W.K. Effects of Eimeria tenella Infection on Key Parameters for Feed Efficiency in Broiler Chickens. Animals 2021, 11, 3428. [Google Scholar] [CrossRef]
- Georgieva, N.; Koinarski, V.; Gadjeva, V. Antioxidant status during the course of Eimeria tenella infection in broiler chickens. Vet. J. 2006, 172, 488–492. [Google Scholar] [CrossRef]
- Qasem, M.A.; Dkhil, M.A.; Al-Shaebi, E.M.; Murshed, M.; Mares, M.; Al-Quraishy, S. Rumex nervosus leaf extracts enhance the regulation of goblet cells and the inflammatory response during infection of chickens with Eimeria tenella. J. King Saud. Univ. Sci. 2020, 32, 1818–1823. [Google Scholar] [CrossRef]
- Pham, H.H.S.; Hatabu, T. Eimeria tenella infection modulates the expression levels of intestinal epithelial barrier-related genes in chicken. J. Environ. Sci. Sustain. Soc. 2021, 10, MR04_p13–MR04_p16. [Google Scholar] [CrossRef]
- Györke, A.; Pop, L.; Cozma, V. Prevalence and distribution of Eimeria species in broiler chicken farms of different capacities. Parasite 2013, 20, 50. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Nolan, M.J.; Harman, K.; Boulton, K.; Hume, D.A.; Tomley, F.M.; Stabler, R.A.; Blake, D.P. Effects of Eimeria tenella infection on chicken caecal microbiome diversity, exploring variation associated with severity of pathology. PLoS ONE 2017, 12, e0184890. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.-H.; Jia, L.-S.; Wei, S.-S.; Ding, H.-Y.; Yang, J.-Y.; Wang, H.-W. Effects of Eimeria tenella infection on the barrier damage and microbiota diversity of chicken cecum. Poult. Sci. 2020, 99, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Kley, M.-V.; Oviedo-Rondón, E.; Dowd, S.; Hume, M.; Nalian, A. Effect of Eimeria infection on cecal microbiome of broilers fed essential oils. Int. J. Poult. Sci. 2012, 11, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Stanley, D.; Geier, M.S.; Chen, H.; Hughes, R.J.; Moore, R.J. Comparison of fecal and cecal microbiotas reveals qualitative similarities but quantitative differences. BMC Microbiol. 2015, 15, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clench, M.H.; Mathias, J.R. The avian cecum: A review. Wilson Bull. 1995, 107, 93–121. [Google Scholar]
- Teng, P.-Y.; Kim, W.K. Roles of prebiotics in intestinal ecosystem of broilers. Front. Vet. Sci. 2018, 5, 245. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Xu, S.; Zeng, D.; Ni, X.; Zhou, M.; Zeng, Y.; Wang, H.; Zhou, Y.; Zhu, H.; Pan, K. Disruption in the cecal microbiota of chickens challenged with Clostridium perfringens and other factors was alleviated by Bacillus licheniformis supplementation. PLoS ONE 2017, 12, e0182426. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Liao, R.; Wu, N.; Zhu, G.; Yang, C. Heat stress mediates changes in fecal microbiome and functional pathways of laying hens. Appl. Microbiol. Biotechnol. 2019, 103, 461–472. [Google Scholar] [CrossRef]
- Kau, A.L.; Ahern, P.P.; Griffin, N.W.; Goodman, A.L.; Gordon, J.I. Human nutrition, the gut microbiome and the immune system. Nature 2011, 474, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Yadav, S.; Wang, J.; Lorentz, B.J.; Lourenco, J.M.; Callaway, T.R.; Kim, W.K. Effects of supplemental tannic acid on growth performance, gut health, microbiota, and fat accumulation and optimal doses of tannic acid in broilers. Front. Physiol. 2022, 13, 912797. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Tompkins, Y.H.; Teng, P.-Y.; Gogal, R.M., Jr.; Kim, W.K. Effects of Tannic Acid Supplementation on Growth Performance, Oocyst Shedding, and Gut Health of in Broilers Infected with Eimeria Maxima. Animals 2022, 12, 1378. [Google Scholar] [CrossRef] [PubMed]
- Lackeyram, D.; Yang, C.; Archbold, T.; Swanson, K.C.; Fan, M.Z. Early weaning reduces small intestinal alkaline phosphatase expression in pigs. J. Nutr. 2010, 140, 461–468. [Google Scholar] [CrossRef] [Green Version]
- Ijaz, U.Z.; Sivaloganathan, L.; McKenna, A.; Richmond, A.; Kelly, C.; Linton, M.; Stratakos, A.C.; Lavery, U.; Elmi, A.; Wren, B.W. Comprehensive longitudinal microbiome analysis of the chicken cecum reveals a shift from competitive to environmental drivers and a window of opportunity for Campylobacter. Front. Microbiol. 2018, 9, 2452. [Google Scholar] [CrossRef]
- Abbas, R.; Iqbal, Z.; Mansoor, M. Role of natural antioxidants for the control of coccidiosis in poultry. Pak. Vet. J. 2013, 33, 401. [Google Scholar]
- Biasato, I.; Ferrocino, I.; Dabbou, S.; Evangelista, R.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Black soldier fly and gut health in broiler chickens: Insights into the relationship between cecal microbiota and intestinal mucin composition. J. Anim. Sci. Biotechnol. 2020, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Xiang, Y.; Zhou, W.; Chen, J.; Li, K.; Yang, H. Microbial community mapping in intestinal tract of broiler chicken. Poult. Sci. 2017, 96, 1387–1393. [Google Scholar] [CrossRef]
- Prasai, T.P.; Walsh, K.B.; Bhattarai, S.P.; Midmore, D.J.; Van, T.T.; Moore, R.J.; Stanley, D. Zeolite food supplementation reduces abundance of enterobacteria. Microbiol. Res. 2017, 195, 24–30. [Google Scholar] [CrossRef] [PubMed]
- McDermott, C.; Feola, R.; Plude, J. Detection of cyanobacterial toxins (microcystins) in waters of northeastern Wisconsin by a new immunoassay technique. Toxicon 1995, 33, 1433–1442. [Google Scholar] [CrossRef]
- Zhang, B.; Lv, Z.; Li, Z.; Wang, W.; Li, G.; Guo, Y. Dietary L-arginine supplementation alleviates the intestinal injury and modulates the gut microbiota in broiler chickens challenged by Clostridium perfringens. Front. Microbiol. 2018, 9, 1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, S.; Sar, A.; Misra, A.; Pal, S.; Chakraborty, A.; Dam, B. Increased productivity in poultry birds by sub-lethal dose of antibiotics is arbitrated by selective enrichment of gut microbiota, particularly short-chain fatty acid producers. Microbiology 2018, 164, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Hui, S.T.; Parks, B.W.; Org, E.; Norheim, F.; Che, N.; Pan, C.; Castellani, L.W.; Charugundla, S.; Dirks, D.L.; Psychogios, N. The genetic architecture of NAFLD among inbred strains of mice. Elife 2015, 4, e05607. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, J.; Zhong, H.; Li, N.; Xu, H.; Zhu, Q.; Liu, Y. Effect of probiotics on the meat flavour and gut microbiota of chicken. Sci. Rep. 2017, 7, 6400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diether, N.E.; Willing, B.P. Microbial fermentation of dietary protein: An important factor in diet–microbe–host interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Witlock, D.; Danforth, H.; Ruff, M. Scanning electron microscopy of Eimeria tenella infection and subsequent repair in chicken caeca. J. Comp. Pathol. 1975, 85, 571–581. [Google Scholar] [CrossRef]
- Yan, Z.; Ming-xue, Z.; Zhi-yong, X.; Huan-cheng, X.; Xiao-zhen, C.; Sha-sha, Y.; Wen-long, Z.; Shan, L.; Qiang-hua, L.; Rui, B. Relationship between Eimeria tenella development and host cell apoptosis in chickens. Poult. Sci. 2015, 94, 2970–2979. [Google Scholar] [CrossRef]
- Gilbert, M.S.; Ijssennagger, N.; Kies, A.K.; van Mil, S.W. Protein fermentation in the gut; implications for intestinal dysfunction in humans, pigs, and poultry. Am. J. Physiol. Gastrointest. 2018, 315, G159–G170. [Google Scholar] [CrossRef]
- Brisbin, J.T.; Gong, J.; Sharif, S. Interactions between commensal bacteria and the gut-associated immune system of the chicken. Anim. Health Res. Rev. 2008, 9, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macpherson, A.; McCoy, K.; Johansen, F.; Brandtzaeg, P. The immune geography of IgA induction and function. Mucosal Immunol. 2008, 1, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Mantis, N.J.; Forbes, S.J. Secretory IgA: Arresting microbial pathogens at epithelial borders. Immunol. Investig. 2010, 39, 383–406. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Palm, N.W. Immunoglobulin A and the microbiome. Curr. Opin. Microbiol. 2020, 56, 89–96. [Google Scholar] [CrossRef]
- Okahashi, N.; Yamamoto, M.; Vancott, J.L.; Chatfield, S.N.; Roberts, M.; Bluethmann, H.; Hiroi, T.; Kiyono, H.; McGhee, J.R. Oral immunization of interleukin-4 (IL-4) knockout mice with a recombinant Salmonella strain or cholera toxin reveals that CD4+ Th2 cells producing IL-6 and IL-10 are associated with mucosal immunoglobulin A responses. Infect. Immun. 1996, 64, 1516–1525. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Kwakkel, R.; Williams, B.; Parmentier, H.; Li, W.; Yang, Z.; Verstegen, M. Effects of mushroom and herb polysaccharides on cellular and humoral immune responses of Eimeria tenella-infected chickens. Poult. Sci. 2004, 83, 1124–1132. [Google Scholar] [CrossRef]
- Tian, E.-j.; Zhou, B.-h.; Wang, X.-y.; Zhao, J.; Deng, W.; Wang, H.-w. Effect of diclazuril on intestinal morphology and SIgA expression in chicken infected with Eimeria tenella. Parasitol. Res. 2014, 113, 4057–4064. [Google Scholar] [CrossRef]
- Bun, S.; Guo, Y.; Guo, F.; Ji, F.; Cao, H. Influence of organic zinc supplementation on the antioxidant status and immune responses of broilers challenged with Eimeria tenella. Poult. Sci. 2011, 90, 1220–1226. [Google Scholar] [CrossRef]
- Gubatan, J.; Holman, D.R.; Puntasecca, C.J.; Polevoi, D.; Rubin, S.J.; Rogalla, S. Antimicrobial peptides and the gut microbiome in inflammatory bowel disease. World J. Gastroenterol. 2021, 27, 7402. [Google Scholar] [CrossRef]
- Nazeer, N.; Uribe-Diaz, S.; Rodriguez-Lecompte, J.C.; Ahmed, M. Antimicrobial peptides as an alternative to relieve antimicrobial growth promoters in poultry. Br. Poult. Sci. 2021, 62, 672–685. [Google Scholar] [CrossRef]
- Hong, Y.; Song, W.; Lee, S.; Lillehoj, H. Differential gene expression profiles of β-defensins in the crop, intestine, and spleen using a necrotic enteritis model in 2 commercial broiler chicken lines. Poult. Sci. 2012, 91, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Yacoub, H.A.; El-Hamidy, S.M.; Mahmoud, M.M.; Baeshen, M.N.; Almehdar, H.A.; Uversky, V.N.; Redwan, E.M.; Al-Maghrabi, O.A.; Elazzazy, A.M. Biocidal activity of chicken defensin-9 against microbial pathogens. Biochem. Cell Biol. 2016, 94, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Dwyer, D.; Miska, K.B.; Fetterer, R.H.; Jenkins, M.C.; Wong, E.A. Expression of host defense peptides in the intestine of Eimeria-challenged chickens. Poult. Sci. 2017, 96, 2421–2427. [Google Scholar] [CrossRef]
- Arike, L.; Hansson, G.C. The densely O-glycosylated MUC2 mucin protects the intestine and provides food for the commensal bacteria. J. Mol. Biol. 2016, 428, 3221–3229. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.E.; Gustafsson, J.K.; Holmén-Larsson, J.; Jabbar, K.S.; Xia, L.; Xu, H.; Ghishan, F.K.; Carvalho, F.A.; Gewirtz, A.T.; Sjövall, H. Bacteria penetrate the normally impenetrable inner colon mucus layer in both murine colitis models and patients with ulcerative colitis. Gut 2014, 63, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.X.; Wu, C.M.; Ribbeck, K. Home, sweet home: How mucus accommodates our microbiota. FEBS J. 2021, 288, 1789–1799. [Google Scholar] [CrossRef] [PubMed]
- Koropatkin, N.M.; Cameron, E.A.; Martens, E.C. How glycan metabolism shapes the human gut microbiota. Nat. Rev. Microbiol. 2012, 10, 323–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Eicher, S.D.; Applegate, T.J. Development of intestinal mucin 2, IgA, and polymeric Ig receptor expressions in broiler chickens and Pekin ducks. Poult. Sci. 2015, 94, 172–180. [Google Scholar] [CrossRef]
- Gao, K.; Jin, J.; Huang, C.; Li, J.; Luo, H.; Li, L.; Huang, Y.; Jiang, Y. Exosomes derived from septic mouse serum modulate immune responses via exosome-associated cytokines. Front. Immunol. 2019, 10, 1560. [Google Scholar] [CrossRef] [Green Version]
- Mutlu, E.A.; Gillevet, P.M.; Rangwala, H.; Sikaroodi, M.; Naqvi, A.; Engen, P.A.; Kwasny, M.; Lau, C.K.; Keshavarzian, A. Colonic microbiome is altered in alcoholism. Am. J. Physiol. Gastrointest. 2012, 302, G966–G978. [Google Scholar] [CrossRef] [Green Version]
- Witlock, D.; Ruff, M.; Chute, M. Physiological basis of Eimeria tenella-induced mortality in individual chickens. J. Parasitol. 1981, 67, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Bilski, J.; Mazur-Bialy, A.; Wojcik, D.; Zahradnik-Bilska, J.; Brzozowski, B.; Magierowski, M.; Mach, T.; Magierowska, K.; Brzozowski, T. The role of intestinal alkaline phosphatase in inflammatory disorders of gastrointestinal tract. Mediat. Inflamm. 2017, 2017, 9074601. [Google Scholar] [CrossRef] [PubMed]
- Malo, M.S.; Moaven, O.; Muhammad, N.; Biswas, B.; Alam, S.N.; Economopoulos, K.P.; Gul, S.S.; Hamarneh, S.R.; Malo, N.S.; Teshager, A. Intestinal alkaline phosphatase promotes gut bacterial growth by reducing the concentration of luminal nucleotide triphosphates. Am. J. Physiol. Gastrointest. 2014, 306, G826–G838. [Google Scholar] [CrossRef]
- Chen, K.T.; Malo, M.S.; Moss, A.K.; Zeller, S.; Johnson, P.; Ebrahimi, F.; Mostafa, G.; Alam, S.N.; Ramasamy, S.; Warren, H.S. Identification of specific targets for the gut mucosal defense factor intestinal alkaline phosphatase. Am. J. Physiol. Gastrointest. 2010, 299, G467–G475. [Google Scholar] [CrossRef]
- Koyama, H.; Ono, T. Induction by short-chain fatty acids of alkaline phosphatase activity in cultured mammalian cells. J. Cell. Physiol. 1976, 88, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Keestra, A.M.; de Zoete, M.R.; Bouwman, L.I.; Vaezirad, M.M.; van Putten, J.P. Unique features of chicken Toll-like receptors. Dev. Comp. Immunol. 2013, 41, 316–323. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, R.; Ma, L.; Wang, Y.; Pan, B.; Cai, J.; Wang, M. Eimeria tenella: Expression profiling of toll-like receptors and associated cytokines in the cecum of infected day-old and three-week old SPF chickens. Exp. Parasitol. 2012, 130, 442–448. [Google Scholar] [CrossRef]
- Weis, S.; Rubio, I.; Ludwig, K.; Weigel, C.; Jentho, E. Hormesis and defense of infectious disease. Int. J. Mol. Sci. 2017, 18, 1273. [Google Scholar] [CrossRef] [Green Version]
- Williams, R. Quantification of the crowding effect during infections with the seven Eimeria species of the domesticated fowl: Its importance for experimental designs and the production of oocyst stocks. Int. J. Parasitol. 2001, 31, 1056–1069. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | D 14 to 20 |
|---|---|
| Corn | 700.8 |
| Soybean meal (480 g crude protein/kg) | 241.73 |
| Soybean oil | 15.84 |
| Defluorinated phosphate | 13.99 |
| Sand | 7.00 |
| Limestone | 6.11 |
| Titanium dioxide | 3.00 |
| DL-Methionine 99% | 2.86 |
| L-Lysine HCl 78% | 2.80 |
| Vitamin Premix 1 | 2.50 |
| Sodium chloride | 1.79 |
| L-threonine | 0.77 |
| Mineral Premix 2 | 0.80 |
| Total | 1000 |
| Calculated energy and nutrient value, % | |
| Metabolizable energy, kcal/kg | 3100 |
| Crude protein | 18.375 |
| SID 3 Methionine | 0.552 |
| SID Total sulfur amino acids | 0.8 |
| SID Lysine | 1.02 |
| SID Threonine | 0.66 |
| Total calcium | 0.76 |
| Available phosphate | 0.38 |
| Target | Primers | Size of PCR Product |
|---|---|---|
| GAPDH | 5′-GCTAAGGCTGTGGGGAAAGT-3′ | 161 |
| 5′-TCAGCAGCAGCCTTCACTAC-3′ | ||
| Beta actin | 5′-CAACACAGTGCTGTCTGGTGGTA-3′ | 205 |
| 5′-ATCGTACTCCTGCTTGCTGATCC-3′ | ||
| AvBD1 | 5′-GGATCGTGTACCTGCTCCTC-3′ | 113 |
| 5′-TGCACAGAAGCCACTCTTTC-3′ | ||
| AvBD5 | 5′-CTCTTTGCTGTCCTCCTCCT-3′ | 118 |
| 5′-CTGGAGGACATGACTTGTGG-3′ | ||
| AvBD9 | 5′-GCTGACACCTTAGCATGCAG-3′ | 113 |
| 5′-CATTTGCAGCATTTCAGCTT-3′ | ||
| CaTH3 | 5′-GCTGTGGACTCCTACAACCA-3′ | 124 |
| 5′-CCATGATGGTGAAGTTGAGG-3′ | ||
| LEAP2 | 5′-TATTCTTCTCGCTGCTGCTC-3′ | 123 |
| 5′-AGGCTCCAACAGGTCTCAGT-3′ | ||
| Alkaline phosphatase | 5′-CTTCCTCGGAGATGGATTTG-3′ | 123 |
| 5′-AGAGCCACATAGGGGAAAGA-3′ | ||
| MUC2 | 5′-ATGCGATGTTAACACAGGACTC-3′ | 110 |
| 5′-GTGGAGCACAGCAGACTTTG-3′ | ||
| IL6 | 5′-ATAAATCCCGATGAAGTGG-3′ | 146 |
| 5′-CTCACGGTCTTCTCCATAAA-3′ | ||
| IL10 | 5′-CTGTCACCGCTTCTTCACC-3′ | 85 |
| 5′-CCCGTTCTCATCCATCTTCT-3′ | ||
| TLR2 | 5′-CGGTGGAAAGGGAGAAAG-3′ | 103 |
| 5′-CTTGCCACATCAGCTTCATT-3′ | ||
| TLR3 | 5′-GGCTAAACGACACTCAAGCA-3′ | 113 |
| 5′-CTTGCAGGCTGAGGTATCAA-3′ | ||
| TLR5 | 5′-CGTTAGTGAGAATGGCTGGA-3′ | 106 |
| 5′-TGAGCCCATTGTATGAGAGC-3′ | ||
| TLR15 | 5′-ATTGAACCTGGTGCATTTGA-3′ | 102 |
| 5′-TTTCAGGTGAGGTGCAAGAC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.; Kim, W. Interactions of Microbiota and Mucosal Immunity in the Ceca of Broiler Chickens Infected with Eimeria tenella. Vaccines 2022, 10, 1941. https://doi.org/10.3390/vaccines10111941
Choi J, Kim W. Interactions of Microbiota and Mucosal Immunity in the Ceca of Broiler Chickens Infected with Eimeria tenella. Vaccines. 2022; 10(11):1941. https://doi.org/10.3390/vaccines10111941
Chicago/Turabian StyleChoi, Janghan, and Wookyun Kim. 2022. "Interactions of Microbiota and Mucosal Immunity in the Ceca of Broiler Chickens Infected with Eimeria tenella" Vaccines 10, no. 11: 1941. https://doi.org/10.3390/vaccines10111941