Protective Effect of Low Molecular Weight Peptides from Solenocera crassicornis Head against Cyclophosphamide-Induced Nephrotoxicity in Mice via the Keap1/Nrf2 Pathway

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of SCPHs-F1
2.3. Antioxidants Activity of SCHPs-F1
2.3.1. Free Radical Scavenging
- DPPH• Scavenging Activity: SCPHs-F1 samples at indicated concentrations (0.1, 0.25, 0.5, 1, 2, 4, and 8 mg/mL) were prepared in ultrapure water (water only for the control sample). Next, 0.2 mM DPPH (prepared by ethanol) work solution was added to the above solutions (ethanol only for the blank control). All mixtures were reacted at room temperature in the dark for 30 min, and the absorbance (A) of solutions was recorded with UV-vis spectra (SpectraMax M2, Molecular Devices Co., San Jose, CA, USA) at 517 nm. The DPPH• scavenging activity of each sample was computed with the following equation:
- •OH Scavenging Activity: The above samples were mixed with 1, 10-Phenanthroline work solution (1.0 mL, 1.865 mM), FeSO4·7H2O (1.0 mL, 1.865 mM), and H2O2 (1.0 mL, 0.03%, v/v) respectively (control without samples, blank control without H2O2). After reaction at 37 °C for 1 h, the absorbance of solutions was recorded with UV-vis spectra at 536 nm. The •OH scavenging activity of each sample was computed with the following equation:
- O2•− Scavenging Activity: The above samples were mixed with Nitrotetrazolium Blue chloride (1.0 mL, 2.52 mM), Nicotinamide adenine dinucleotide (1.0 mL, 624 mM), and Phenazine methosulphate (1.0 mL, 120 μM), respectively (control without samples). After reaction at room temperature in the dark for 5 min, the absorbance of solutions was recorded with UV-vis spectra at 560 nm. The O2•− scavenging activity of each sample was computed with the following equation:
2.3.2. Reducing Power
2.4. Renoprotective Effects on CTX-Induced Mice
2.4.1. Animals and Experimental Plan
2.4.2. Sample Collection
2.4.3. Biochemistry Assays
2.4.4. Histopathological Examination
2.4.5. Western Blot Analysis
2.4.6. Statistical Analysis
3. Results
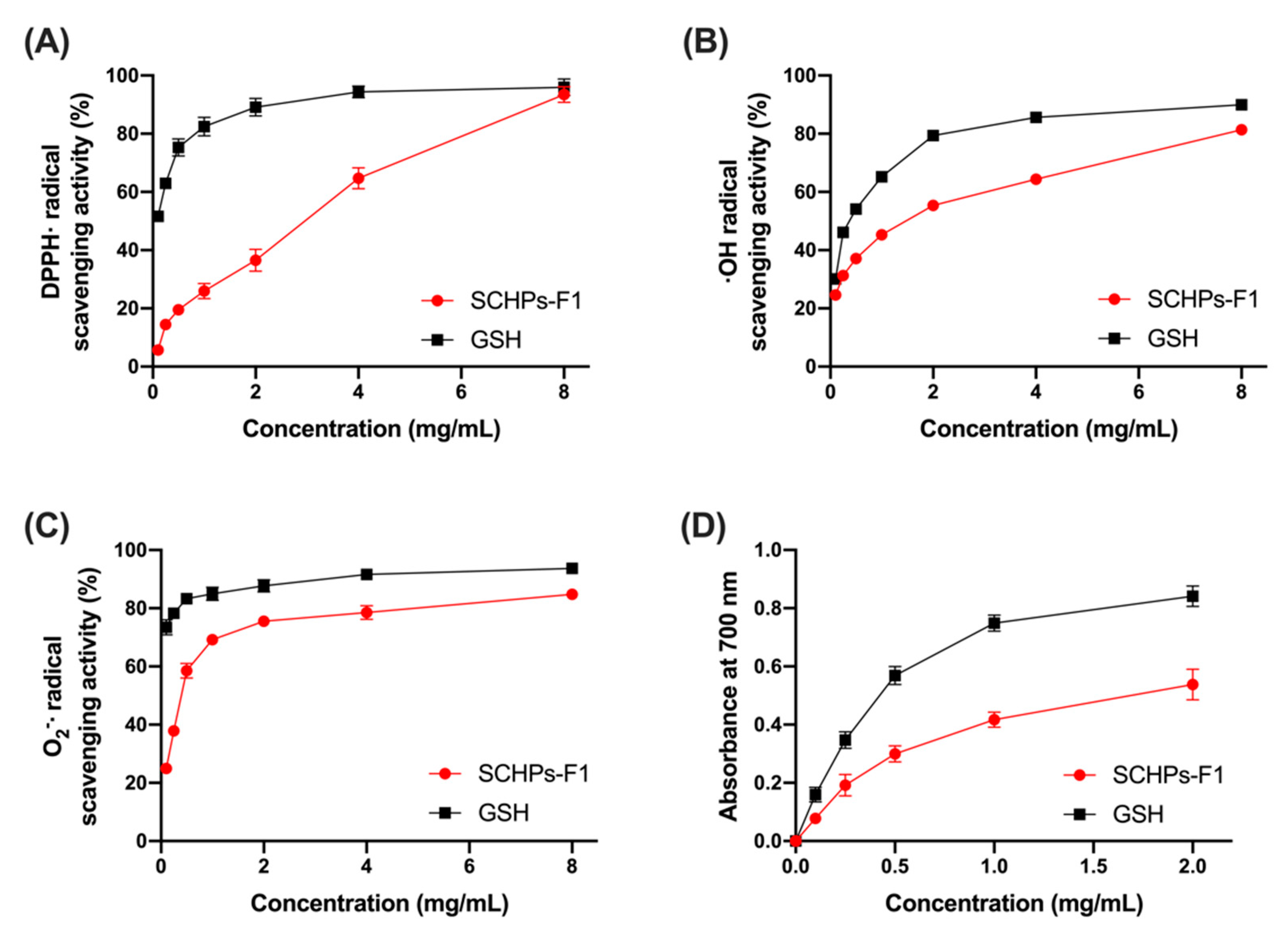
3.1. Radical Scavenging Activity of SCHPs-F1
3.2. SCHPs-F1 Treatment Modulates Renal Function in Mice Exposed to CTX
3.3. SCHPs-F1 Treatment Relieves CTX-Induced Renal Oxidative Stress
3.4. SCHPs-F1 Treatment Mitigates CTX-Induced Renal Inflammation
3.5. SCHPs-F1 Reverses Bax/Bcl-2 Imbalance in CTX-Exposed Kidney
3.6. SCHPs-F1 Treatment Ameliorates Pathomorphology in CTX-Exposed Kidney
3.7. SCHPs-F1 Elevate Nrf2 Related Protein Expression in CTX-Induced Kidney
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Moghe, A.; Ghare, S.; Lamoreau, B.; Mohammad, M.; Barve, S.; McClain, C.; Joshi-Barve, S. Molecular mechanisms of acrolein toxicity: Relevance to human disease. Toxicol. Sci. 2015, 143, 242–255. [Google Scholar] [CrossRef]
- Ponticelli, C.; Escoli, R.; Moroni, G. Does cyclophosphamide still play a role in glomerular diseases? Autoimmun. Rev. 2018, 17, 1022–1027. [Google Scholar] [CrossRef]
- Sherif, I.O. Uroprotective mechanism of quercetin against cyclophosphamide-induced urotoxicity: Effect on oxidative stress and inflammatory markers. J. Cell. Biochem. 2018, 119, 7441–7448. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, F.; Nikzad, H.; Taghizadeh, M.; Taherian, A.; Azami-Tameh, A.; Hosseini, S.M.; Moravveji, A. Protective effect of Zingiber officinale extract on rat testis after cyclophosphamide treatment. Andrologia 2014, 46, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Nagi, M.N.; Al-Shabanah, O.A.; Hafez, M.M.; Sayed-Ahmed, M.M. Thymoquinone supplementation attenuates cyclophosphamide-induced cardiotoxicity in rats. J. Biochem. Mol. Toxicol. 2011, 25, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Alhaithloul, H.A.S.; Alotaibi, M.F.; Bin-Jumah, M.; Elgebaly, H.; Mahmoud, A.M. Olea europaea leaf extract up-regulates Nrf2/ARE/HO-1 signaling and attenuates cyclophosphamide-induced oxidative stress, inflammation and apoptosis in rat kidney. Biomed. Pharmacother. 2019, 111, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.N.; Li, Y.Z.; Li, W.; Yan, X.T.; Yang, G.; Zhang, J.; Zhao, L.C.; Yang, L.M. Nephroprotective effects of saponins from leaves of panax quinquefolius against cisplatin-induced acute kidney injury. Int. J. Mol. Sci. 2017, 18, 1407. [Google Scholar] [CrossRef]
- Iqubal, A.; Iqubal, M.K.; Sharma, S.; Ansari, M.A.; Najmi, A.K.; Ali, S.M.; Ali, J.; Haque, S.E. Molecular mechanism involved in cyclophosphamide-induced cardiotoxicity: Old drug with a new vision. Life Sci. 2019, 218, 112–131. [Google Scholar] [CrossRef]
- Singh, R.K.; Kumar, S.; Prasad, D.N.; Bhardwaj, T.R. Therapeutic journery of nitrogen mustard as alkylating anticancer agents: Historic to future perspectives. Eur. J. Med. Chem. 2018, 151, 401–433. [Google Scholar] [CrossRef]
- Jin, L.; Ding, G.; Li, P.; Gu, J.; Zhang, X. Changes in quality attributes of marine-trawling shrimp (Solenocera crassicornis) during storage under different deep-frozen temperatures. J. Food Sci. Technol. 2018, 55, 2890–2898. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; Shahidi, F. Bioactive peptides from shrimp shell processing discards: Antioxidant and biological activities. J. Funct. Foods 2017, 34, 7–17. [Google Scholar] [CrossRef]
- Kim, S.-B.; Yoon, N.Y.; Shim, K.-B.; Lim, C.-W. Antioxidant and angiotensin I-converting enzyme inhibitory activities of northern shrimp (Pandalus borealis) by-products hydrolysate by enzymatic hydrolysis. Fish. Aquat. Sci. 2016, 19, 29. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Guo, N.; Sun, J.; Xue, C. Comprehensive utilization of shrimp waste based on biotechnological methods: A review. J. Clean. Prod. 2017, 143, 814–823. [Google Scholar] [CrossRef]
- Jiang, S.; Zhang, Z.; Yu, F.; Zhang, Z.; Yang, Z.; Tang, Y.; Ding, G. Ameliorative effect of low molecular weight peptides from the head of red shrimp (Solenocera crassicornis) against cyclophosphamide-induced hepatotoxicity in mice. J. Funct. Foods 2020, 72, 104085. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, G.-X.; Zhao, Y.-Q.; Qiu, Y.-T.; Chi, C.-F.; Wang, B. Identification and active evaluation of antioxidant peptides from protein hydrolysates of skipjack tuna (Katsuwonus pelamis) head. Antioxidants 2019, 8, 318. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.-Y.; Wang, Y.-M.; Li, L.; Chi, C.-F.; Wang, B. Four antioxidant peptides from protein hydrolysate of red stingray (Dasyatis akajei) cartilages: Isolation, identification, and in vitro activity evaluation. Mar. Drugs 2019, 17, 263. [Google Scholar] [CrossRef] [Green Version]
- AAT Bioquest, Inc. Quest Graph™ EC50 Calculator. Available online: https://www.aatbio.com/tools/ec50-calculator (accessed on 1 July 2020).
- He, Y.; Pan, X.; Chi, C.-F.; Sun, K.-L.; Wang, B. Ten new pentapeptides from protein hydrolysate of miiuy croaker (Miichthys miiuy) muscle: Preparation, identification, and antioxidant activity evaluation. LWT-Food Sci. Technol. 2019, 105, 1–8. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, Z.; Ye, S.; Hong, X.; Jin, H.; Huang, F.; Yang, Z.; Tang, Y.; Chen, Y.; Ding, G. Immunoenhancement effects of pentadecapeptide derived from Cyclina sinensis on immune-deficient mice induced by Cyclophosphamide. J. Funct. Foods 2019, 60, 103408. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, X.; Lin, L.; Ding, G.; Yu, F. Immunomodulatory activity of low molecular-weight peptides from nibea japonica in RAW264.7 cells via NF-κB pathway. Mar. Dugs 2019, 17, 404. [Google Scholar] [CrossRef] [Green Version]
- Kang, X.; Jing, M.; Zhang, G.; He, L.; Hong, P.; Deng, C. The ameliorating effect of plasma protein from tachypleus tridentatus on cyclophosphamide-induced acute kidney injury in mice. Mar. Drugs 2019, 17, 227. [Google Scholar] [CrossRef] [Green Version]
- Philips, F.S.; Sternberg, S.S.; Cronin, A.P.; Vidal, P.M. Cyclophosphamide and urinary bladder toxicity. Cancer Res. 1961, 21, 1577–1589. [Google Scholar] [PubMed]
- El-Naggar, S.A.; Alm-Eldeen, A.A.; Germoush, M.O.; El-Boray, K.F.; Elgebaly, H.A. Ameliorative effect of propolis against cyclophosphamide-induced toxicity in mice. Pharm. Biol. 2015, 53, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Haque, R.; Bin-Hafeez, B.; Parvez, S.; Pandey, S.; Sayeed, I.; Ali, M.; Raisuddin, S. Aqueous extract of walnut (Juglans regia L.) protects mice against cyclophosphamideinduced biochemical toxicity. Hum. Exp. Toxicol. 2016, 22, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Qu, T.; Wang, E.; Li, A.; Du, G.; Li, Z.; Qin, X. NMR based metabolomic approach revealed cyclophosphamide-induced systematic alterations in a rat model. RSC Adv. 2016, 6, 111020–111030. [Google Scholar] [CrossRef]
- Dhanisha, S.S.; Drishya, S.; Guruvayoorappan, C. Pithecellobium dulce fruit extract mitigates cyclophosphamide-mediated toxicity by regulating proinflammatory cytokines. J. Food Biochem. 2020, 44, e13083. [Google Scholar] [CrossRef]
- Basu, A.; Singha Roy, S.; Bhattacharjee, A.; Bhuniya, A.; Baral, R.; Biswas, J.; Bhattacharya, S. Vanadium(III)-L-cysteine protects cisplatin-induced nephropathy through activation of Nrf2/HO-1 pathway. Free Radic. Res. 2016, 50, 39–55. [Google Scholar] [CrossRef]
- Domitrović, R.; Cvijanović, O.; Šušnić, V.; Katalinić, N. Renoprotective mechanisms of chlorogenic acid in cisplatin-induced kidney injury. Toxicology 2014, 324, 98–107. [Google Scholar] [CrossRef]
- Boddy, A.V.; Yule, S.M. Metabolism and pharmacokinetics of oxazaphosphorines. Clin. Pharmacokinet. 2000, 38, 291–304. [Google Scholar] [CrossRef]
- Wagner, T. Ifosfamide clinical pharmacokinetics. Clin. Pharmacokinet. 1994, 26, 439–456. [Google Scholar] [CrossRef]
- Goudarzi, M.; Khodayar, M.J.; Hosseini Tabatabaei, S.M.T.; Ghaznavi, H.; Fatemi, I.; Mehrzadi, S. Pretreatment with melatonin protects against cyclophosphamide-induced oxidative stress and renal damage in mice. Fundam. Clin. Pharmacol. 2017, 31, 625–635. [Google Scholar] [CrossRef]
- Yaman, T.; Yener, Z.; Celik, I. Histopathological and biochemical investigations of protective role of honey in rats with experimental aflatoxicosis. BMC Complement. Altern. Med. 2016, 16, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharjee, A.; Basu, A.; Biswas, J.; Bhattacharya, S.J.M.; Biochemistry, C. Nano-Se attenuates cyclophosphamide-induced pulmonary injury through modulation of oxidative stress and DNA damage in Swiss albino mice. Mol. Cell. Biochem. 2015, 405, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Zhang, J.; Cao, Z.; Ji, Q.; Han, Y.; Song, M.; Shao, B.; Li, Y. Lycopene attenuates AFB1-induced renal injury with the activation of the Nrf2 antioxidant signaling pathway in mice. Food Funct. 2018, 9, 6427–6434. [Google Scholar] [CrossRef] [PubMed]
- Abraham, P.; Rabi, S. Protective effect of aminoguanidine against cyclophosphamide-induced oxidative stress and renal damage in rats. Redox Rep. 2011, 16, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Lin, X.; Li, H.; Yuan, J.; Peng, Y.; Dong, L.; Dai, S. Paeoniflorin ameliorates renal function in cyclophosphamide-induced mice via AMPK suppressed inflammation and apoptosis. Biomed. Pharmacother. 2016, 84, 1899–1905. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, P.; Kulurkar, P.; Singh, D.; Kumar, D.; Patial, V. Iridoid glycosides fraction from Picrorhiza kurroa attenuates cyclophosphamide-induced renal toxicity and peripheral neuropathy via PPAR-γ mediated inhibition of inflammation and apoptosis. Phytomedicine 2017, 36, 108–117. [Google Scholar] [CrossRef]
- Caglayan, C.; Temel, Y.; Kandemir, F.M.; Yildirim, S.; Kucukler, S.J.E.S.; Research, P. Naringin protects against cyclophosphamide-induced hepatotoxicity and nephrotoxicity through modulation of oxidative stress, inflammation, apoptosis, autophagy, and DNA damage. Environ. Sci. Pollut. Res. 2018, 25, 20968–20984. [Google Scholar] [CrossRef]
- Cakir, M.; Duzova, H.; Baysal, I.; Gul, C.C.; Kuscu, G.; Kutluk, F.; Cakin, H.; Seker, S.; Ilbegi, E.; Uslu, S.; et al. The effect of hypericum perforatum on kidney ischemia/reperfusion damage. Ren. Fail. 2017, 39, 385–391. [Google Scholar] [CrossRef]
- Blanco, A.; Blanco, G. Chapter 32—Apoptosis. In Medical Biochemistry; Blanco, A., Blanco, G., Eds.; Academic Press: New York, NY, USA, 2017; pp. 791–796. [Google Scholar] [CrossRef]
- Fouad, A.A.; Abdel-Gaber, S.A.; Abdelghany, M.I. Hesperidin opposes the negative impact of cyclophosphamide on mice kidneys. Drug Chem. Toxicol. 2019, 1–6. [Google Scholar] [CrossRef]
- Bhakkiyalakshmi, E.; Sireesh, D.; Ramkumar, K.M. Chapter 12—Redox sensitive transcription via Nrf2-keap1 in suppression of inflammation. In Immunity and Inflammation in Health and Disease; Chatterjee, S., Jungraithmayr, W., Bagchi, D., Eds.; Academic Press: New York, NY, USA, 2018; pp. 149–161. [Google Scholar] [CrossRef]
- Liang, Y.; Li, J.; Lin, Q.; Huang, P.; Zhang, L.; Wu, W.; Ma, Y. Research progress on signaling pathway-associated oxidative stress in endothelial cells. Oxid. Med. Cell. Longev. 2017, 7156941. [Google Scholar] [CrossRef]
- Zhu, Z.; Shi, Z.; Xie, C.; Gong, W.; Hu, Z.; Peng, Y. A novel mechanism of Gamma-aminobutyric acid (GABA) protecting human umbilical vein endothelial cells (HUVECs) against H2O2-induced oxidative injury. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2018, 217, 68–75. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MDA (nmol/mg Prot) | CAT (U/mg Prot) | SOD (U/mg Prot) | GSH-Px (U/mg Prot) | T-AOC (U/mg Prot) | |
|---|---|---|---|---|---|
| Control | 18.67 ± 1.31 | 39.28 ± 1.18 | 11.31 ± 0.25 | 11.15 ± 0.33 | 2.08 ± 0.09 |
| Model | 34.01 ± 2.23 ** | 30.65 ± 1.29 ** | 7.62 ± 0.63 ** | 8.44 ± 0.43 ** | 1.16 ± 0.05 ** |
| SCHPs-F1 100 | 28.92 ± 1.36 **,++ | 32.52 ± 2.20 ** | 8.37 ± 0.19 ** | 8.66 ± 0.42 ** | 1.35 ± 0.09 **,+ |
| SCHPs-F1 200 | 26.64 ± 1.29 **,++ | 35.25 ± 2.45 *,++ | 8.83 ± 0.64 **,+ | 8.79 ± 0.11 ** | 1.54 ± 0.04 **,++ |
| SCHPs-F1 400 | 20.88 ± 0.85 ++ | 36.63 ± 1.23 **,++ | 10.60 ± 0.81 ++ | 10.07 ± 0.24 **,++ | 1.83 ± 0.11 **,++ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Zhang, Z.; Huang, F.; Yang, Z.; Yu, F.; Tang, Y.; Ding, G. Protective Effect of Low Molecular Weight Peptides from Solenocera crassicornis Head against Cyclophosphamide-Induced Nephrotoxicity in Mice via the Keap1/Nrf2 Pathway. Antioxidants 2020, 9, 745. https://doi.org/10.3390/antiox9080745
Jiang S, Zhang Z, Huang F, Yang Z, Yu F, Tang Y, Ding G. Protective Effect of Low Molecular Weight Peptides from Solenocera crassicornis Head against Cyclophosphamide-Induced Nephrotoxicity in Mice via the Keap1/Nrf2 Pathway. Antioxidants. 2020; 9(8):745. https://doi.org/10.3390/antiox9080745
Chicago/Turabian StyleJiang, Shuoqi, Zhuangwei Zhang, FangFang Huang, Zuisu Yang, Fangmiao Yu, Yunping Tang, and Guofang Ding. 2020. "Protective Effect of Low Molecular Weight Peptides from Solenocera crassicornis Head against Cyclophosphamide-Induced Nephrotoxicity in Mice via the Keap1/Nrf2 Pathway" Antioxidants 9, no. 8: 745. https://doi.org/10.3390/antiox9080745