Systemic Insulin Resistance and Metabolic Perturbations in Chow Fed Inducible Nitric Oxide Synthase Knockout Male Mice: Partial Reversal by Nitrite Supplementation


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Research Design and Methods
2.1. Animal Studies
2.2. Tolerance Tests
2.3. Body Composition Analysis
2.4. Indirect Calorimetry
2.5. Serum Biochemistry
2.6. Total Nitrite Estimation
2.7. Tissue Biochemistry
2.8. Hematoxylin and Eosin (HE) Staining
2.9. Oil Red O Staining
2.10. Western Blot Analysis and Real Time PCR
2.11. Statistical Analysis
2.12. Data Availability
3. Results
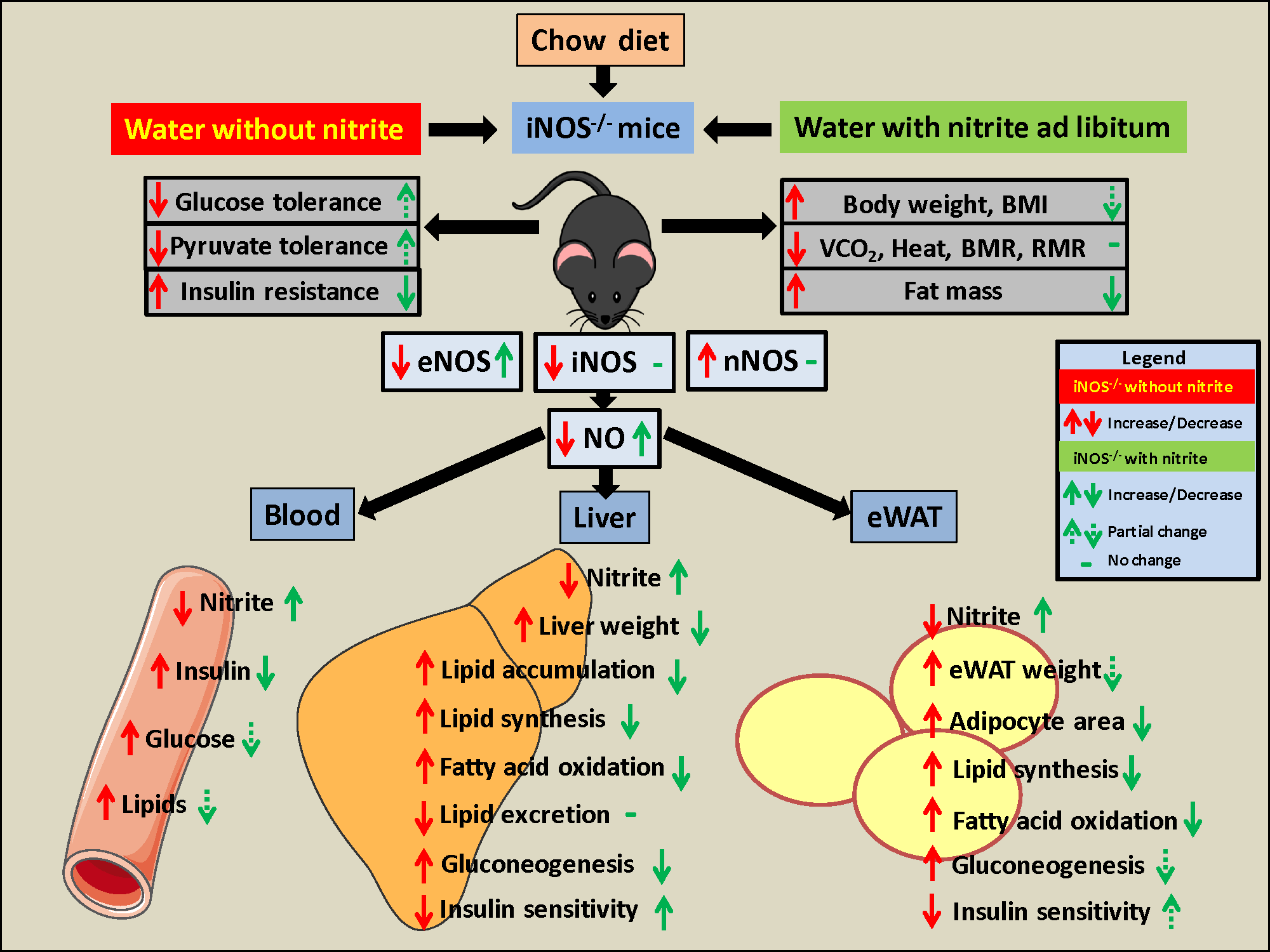
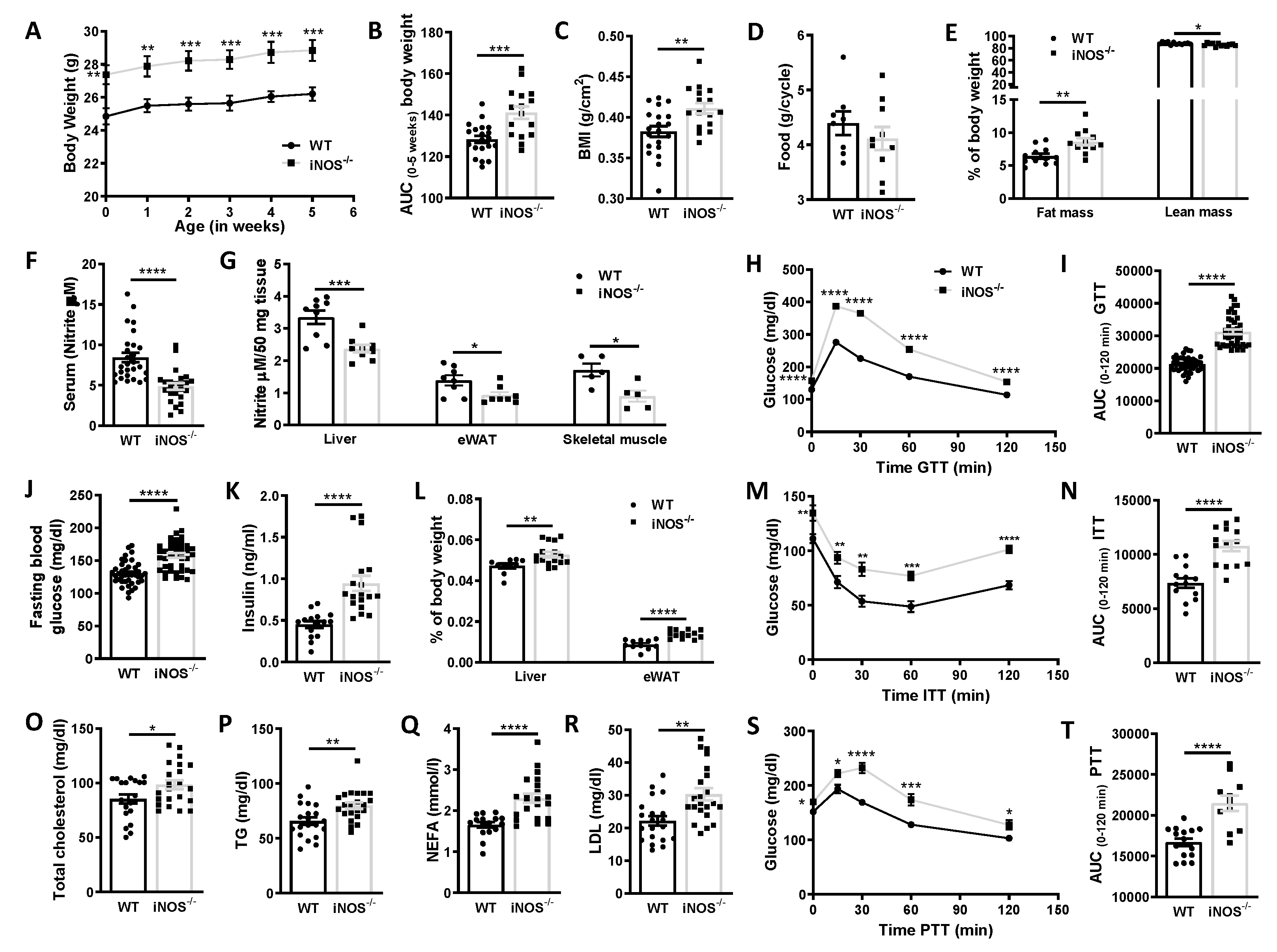
3.1. Gross Parameters, Systemic Insulin, Glucose, Pyruvate Tolerance, and Circulating Lipids
3.2. Status of Metabolic Homeostasis in WT and iNOS−/− Mice
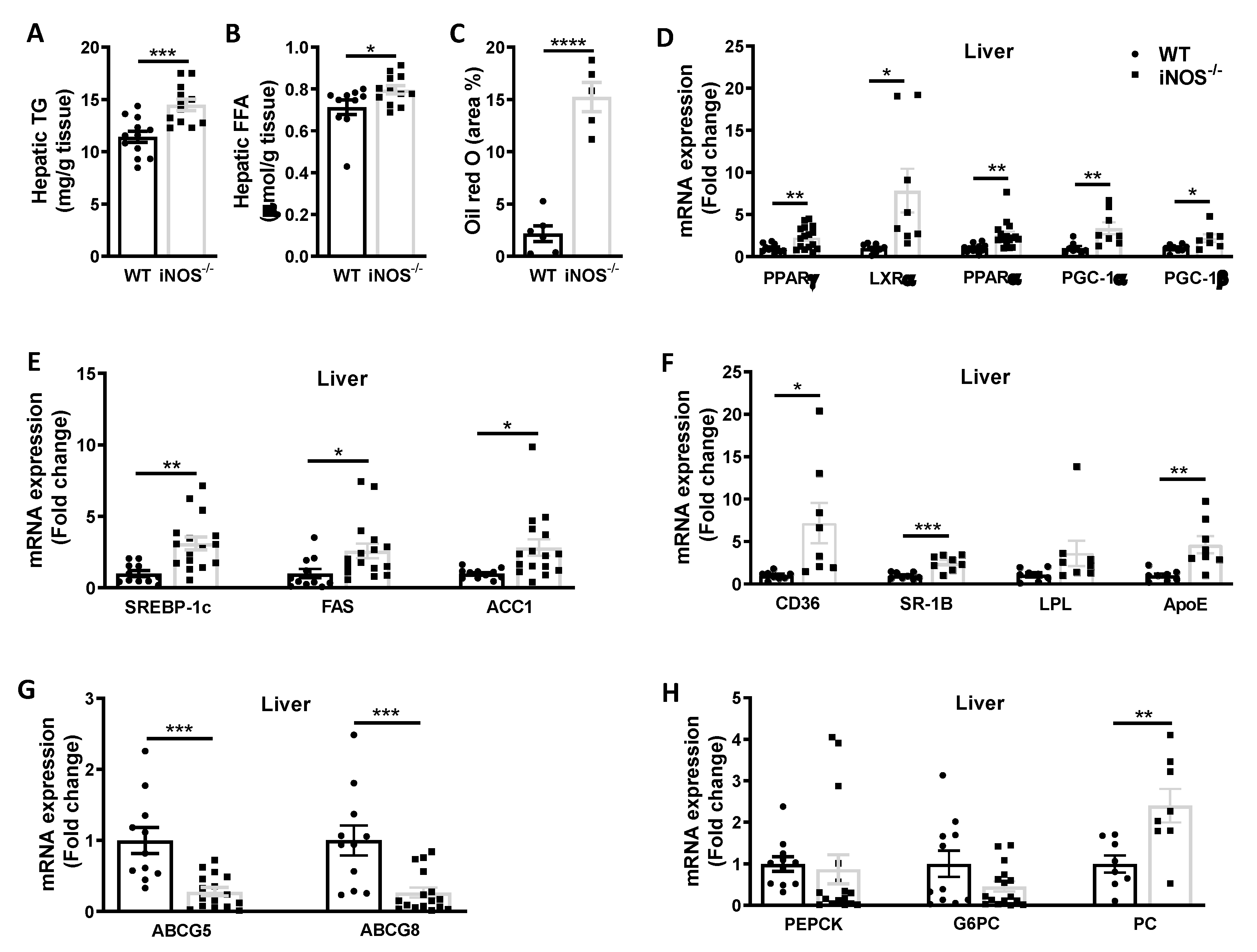
3.2.1. Metabolic Homeostasis in the Liver Tissue
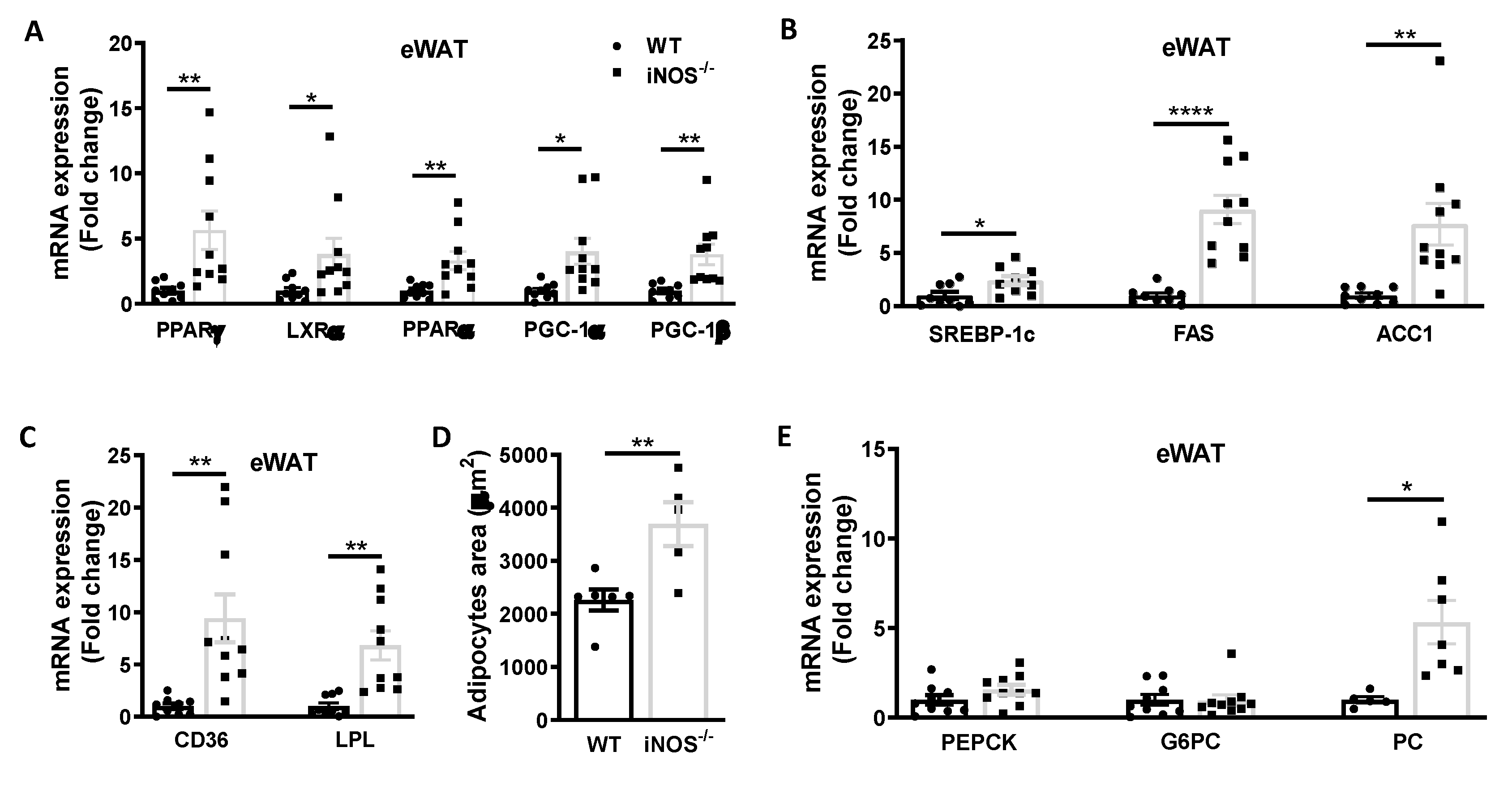
3.2.2. Metabolic Homeostasis in the Adipose Tissue
3.3. Alterations in the Gross Parameters, Glucose Tolerance, Insulin Sensitivity, Gluconeogenesis, and Circulating Lipids after Nitrite Supplementation in Chow Fed iNOS−/− Mice
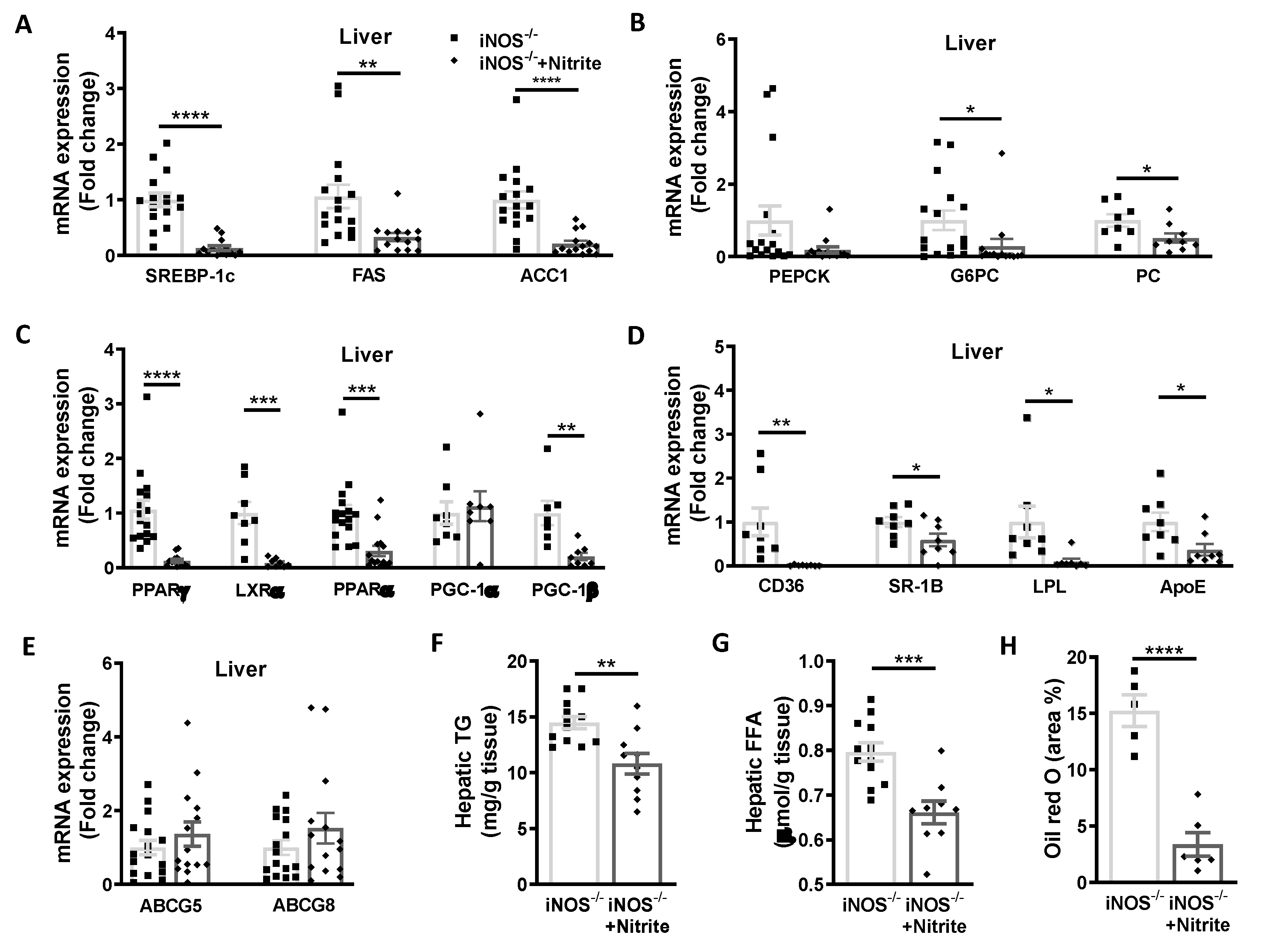
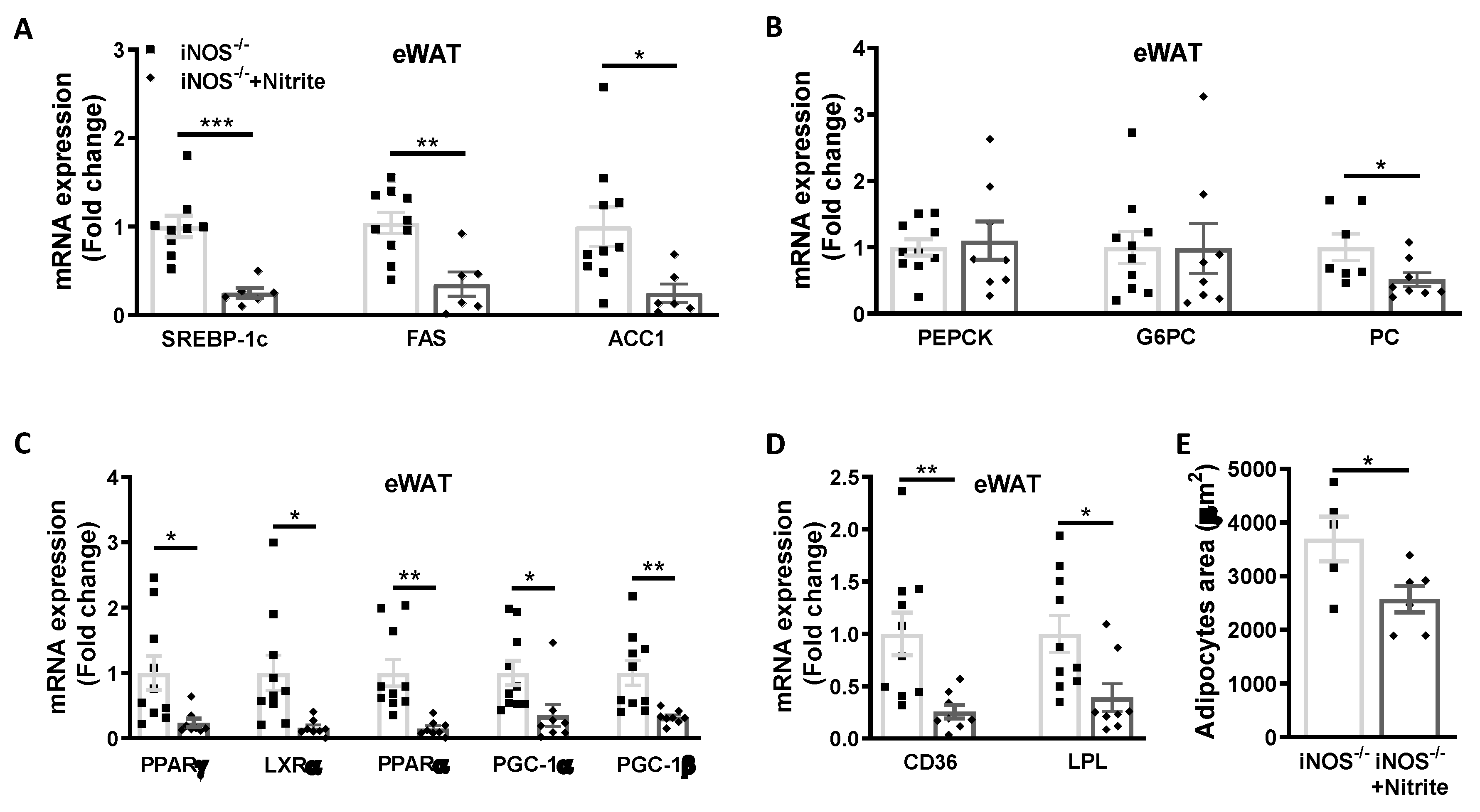
3.4. Metabolic Homeostasis in Liver and Adipose Tissue in Chow Fed iNOS−/− Mice after Nitrite Supplementation
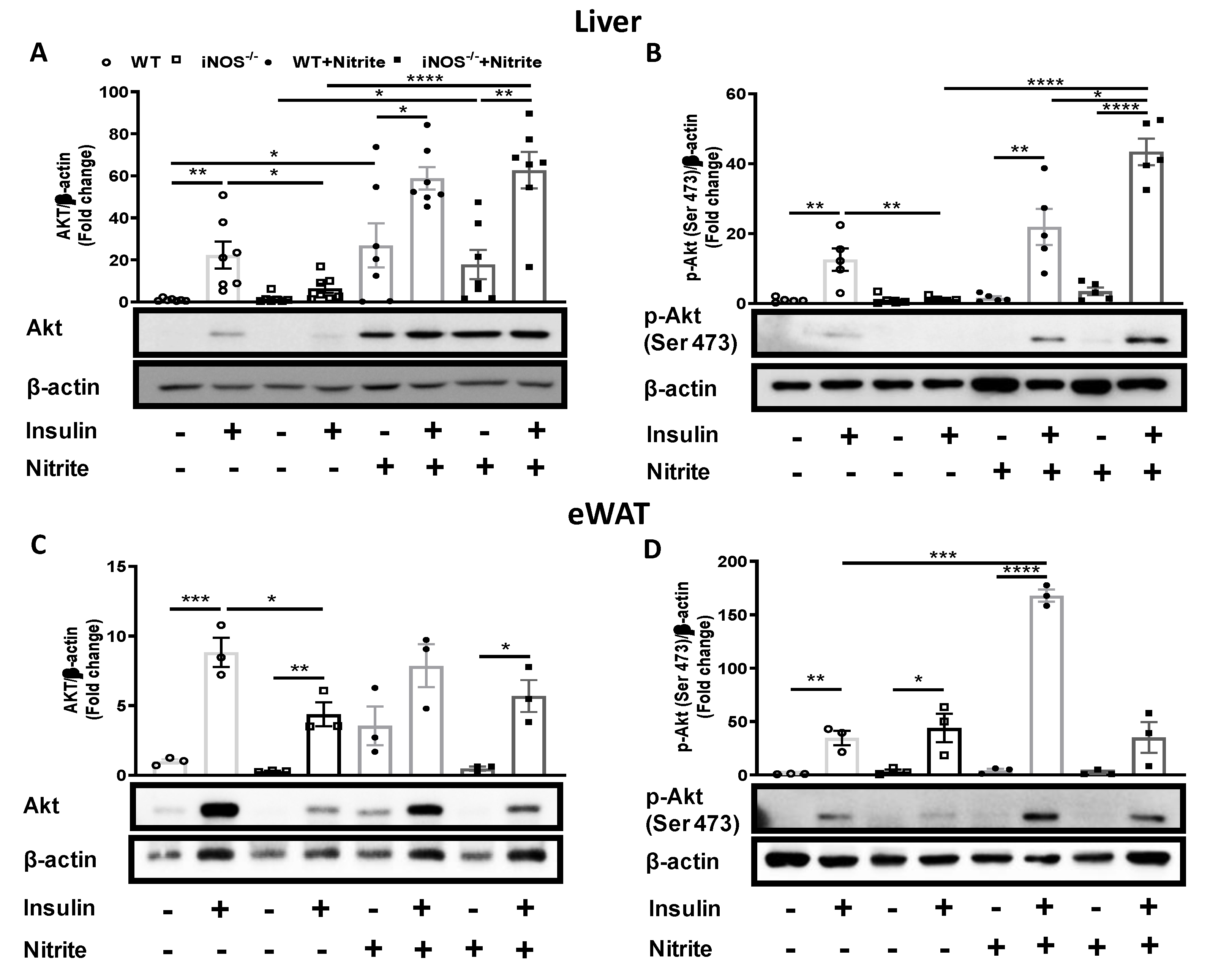
3.5. Insulin Signaling in Nitrite Supplemented Chow Fed WT and iNOS−/− Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABCG5/8 | ATP-binding cassette subfamily G member 5/8; |
| ACC1 | Acetyl-CoA carboxylase 1; |
| ApoE | Apolipoprotein E; |
| BMI | Body mass index; |
| BMR | Basal metabolic rate; |
| CD36 | Cluster of differentiation 36; |
| eNOS | Endothelial-nitric oxide synthase; |
| eWAT | epididymal white adipose tissue; |
| FAS | Fatty acid synthase; |
| FoxO1 | Forkhead box O1 |
| G6PC | Glucose-6-phosphatase; |
| HFD | High fat diet; |
| HMGCoR | 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase; |
| HOMA-IR | Homeostatic Model Assessment of Insulin Resistance; |
| iNOS−/− | Inducible-nitric oxide synthase knockout; |
| IPGTT | Intra-peritoneal glucose tolerance test; |
| IR | Insulin resistance; |
| ITT | Insulin tolerance test; |
| KO | Knock out |
| LDLR | Low-Density Lipoprotein Receptor; |
| LFD | Low fat diet; |
| LPL | Lipoprotein Lipase; |
| LXR | Liver X receptor; |
| nNOS | Neuronal-nitric oxide synthase; |
| NO | Nitric oxide; |
| pAkt | phospho protein kinase B; |
| PC | Pyruvate carboxylase; |
| PCSK9 | Proprotein convertase subtilisin/kexin type 9; |
| PEPCK | Phosphoenolpyruvate carboxykinase; |
| PGC | Peroxisome proliferator-activated receptor-γ coactivator; |
| PI3K | Phosphoinositide 3-kinases; |
| PPAR | Peroxisome proliferator-activated receptor; |
| PTT | Pyruvate tolerance test; |
| QUICKI | Quantitative insulin-sensitivity check index; |
| RER | Respiratory Exchange Ratio; |
| RMR | Resting metabolic rate; |
| SR-1B | Scavenger receptor, class B type 1; |
| SREBP-1c | Sterol regulatory element-binding protein 1c; |
| VLDL | Very-low-density lipoprotein; |
| WT | Wild type |
References
- Anavi, S.; Tirosh, O. iNOS as a metabolic enzyme under stress conditions. Free Radic. Biol. Med. 2020, 146, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, C.; Giulivi, C. Subcellular and cellular locations of nitric oxide synthase isoforms as determinants of health and disease. Free Radic. Biol. Med. 2010, 49, 307–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.M.; Billiar, T.R. Regulation and Function of Inducible Nitric Oxide Synthase during Sepsis and Acute Inflammation. Adv. Pharmacol. 1995, 34, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Kröncke, K.D.; Fehsel, K.; Kolb-Bachofen, V. Inducible nitric oxide synthase in human diseases. Clin. Exp. Immunol. 1998, 113, 147–156. [Google Scholar] [CrossRef]
- House, L.M.; Morris, R.T.; Barnes, T.M.; Lantier, L.; Cyphert, T.J.; McGuinness, O.P.; Otero, Y.F. Tissue inflammation and nitric oxide-mediated alterations in cardiovascular function are major determinants of endotoxin-induced insulin resistance. Cardiovasc. Diabetol. 2015, 14, 56. [Google Scholar] [CrossRef] [Green Version]
- McNaughton, L.; Puttagunta, L.; Martinez-Cuesta, M.A.; Kneteman, N.; Mayers, I.; Moqbel, R.; Hamid, Q.; Radomski, M.W. Distribution of nitric oxide synthase in normal and cirrhotic human liver. Proc. Natl. Acad. Sci. USA 2002, 99, 17161–17166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elizalde, M.; Rydén, M.; van Harmelen, V.; Eneroth, P.; Gyllenhammar, H.; Holm, C.; Ramel, S.; Olund, A.; Arner, P.; Andersson, K. Expression of nitric oxide synthases in subcutaneous adipose tissue of nonobese and obese humans. J. Lipid Res. 2000, 41, 1244–1251. [Google Scholar]
- Park, C.-S.; Park, R.; Krishna, G. Constitutive expression and structural diversity of inducible isoform of nitric oxide synthase in human tissues. Life Sci. 1996, 59, 219–225. [Google Scholar] [CrossRef]
- Hoffman, R.A.; Zhang, G.; Nussler, N.C.; Gleixner, S.L.; Ford, H.R.; Simmons, R.L.; Watkins, S.C. Constitutive expression of inducible nitric oxide synthase in the mouse ileal mucosa. Am. J. Physiol. Liver Physiol. 1997, 272, G383–G392. [Google Scholar] [CrossRef]
- Perner, A.; Andresen, L.; Normark, M.; Rask-Madsen, J. Constitutive Expression of Inducible Nitric Oxide Synthase in the Normal Human Colonic Epithelium. Scand. J. Gastroenterol. 2002, 37, 944–948. [Google Scholar] [CrossRef]
- Saini, R.; Patel, S.; Saluja, R.; Sahasrabuddhe, A.A.; Singh, M.P.; Habib, S.; Bajpai, V.K.; Dikshit, M. Nitric oxide synthase localization in the rat neutrophils: Immunocytochemical, molecular, and biochemical studies. J. Leukoc. Biol. 2006, 79, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlqvist, E.; Storm, P.; Käräjämäki, A.; Martinell, M.; Dorkhan, M.; Carlsson, A.; Vikman, P.; Prasad, R.B.; Aly, D.M.; Almgren, P.; et al. Novel subgroups of adult-onset diabetes and their association with outcomes: A data-driven cluster analysis of six variables. Lancet Diabetes Endocrinol. 2018, 6, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuñiga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Warpeha, K.M.; Xu, W.; Liu, L.; Charles, I.G.; Patterson, C.C.; Ah-Fat, F.; Harding, S.; Hart, P.M.; Chakravarthy, U.; Hughes, A.E. Genotyping and functional analysis of a polymorphic (CCTTT) (n) repeat of NOS2A in diabetic retinopathy. FASEB J. 1999, 13, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Johannesen, J.; Tarnow, L.; Parving, H.H.; Nerup, J.; Pociot, F. CCTTT-repeat polymorphism in the human NOS2-promoter confers low risk of diabetic nephropathy in type 1 diabetic patients. Diabetes Care 2000, 23, 560–562. [Google Scholar] [CrossRef] [Green Version]
- Pathak, P.; Kanshana, J.S.; Kanuri, B.; Rebello, S.C.; Aggarwal, H.; Jagavelu, K.; Dikshit, M. Vasoreactivity of isolated aortic rings from dyslipidemic and insulin resistant inducible nitric oxide synthase knockout mice. Eur. J. Pharmacol. 2019, 855, 90–97. [Google Scholar] [CrossRef]
- Noronha, B.T.; Li, J.-M.; Wheatcroft, S.B.; Shah, A.M.; Kearney, M.T. Inducible nitric oxide synthase has divergent effects on vascular and metabolic function in obesity. Diabetes 2005, 54, 1082–1089. [Google Scholar] [CrossRef] [Green Version]
- Kim, F.; Pham, M.; Maloney, E.; Rizzo, N.O.; Morton, G.J.; Wisse, B.E.; Kirk, E.A.; Chait, A.; Schwartz, M.W. Vascular Inflammation, Insulin Resistance, and Reduced Nitric Oxide Production Precede the Onset of Peripheral Insulin Resistance. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1982–1988. [Google Scholar] [CrossRef]
- Kina-Tanada, M.; Sakanashi, M.; Tanimoto, A.; Kaname, T.; Matsuzaki, T.; Noguchi, K.; Uchida, T.; Nakasone, J.; Kozuka, C.; Ishida, M.; et al. Long-term dietary nitrite and nitrate deficiency causes the metabolic syndrome, endothelial dysfunction and cardiovascular death in mice. Diabetologia 2017, 60, 1138–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthys, K.E.; Bult, H. Nitric oxide function in atherosclerosis. Mediators Inflamm. 1997, 6, 3. [Google Scholar] [CrossRef]
- Carlström, M.; Larsen, F.J.; Nyström, T.; Hezel, M.; Borniquel, S.; Weitzberg, E.; Lundberg, J.O. Dietary inorganic nitrate reverses features of metabolic syndrome in endothelial nitric oxide synthase-deficient mice. Proc. Natl. Acad. Sci. USA 2010, 107, 17716–17720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, R.R.; Wu, Y.; Shen, H.Q.; Zhu, J.S.; Baron, A.D. Mice with gene disruption of both endothelial and neuronal nitric oxide synthase exhibit insulin resistance. Diabetes 2000, 49, 684–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsutsui, M.; Tanimoto, A.; Tamura, M.; Mukae, H.; Yanagihara, N.; Shimokawa, H.; Otsuji, Y. Significance of nitric oxide synthases: Lessons from triple nitric oxide synthases null mice. J. Pharmacol. Sci. 2015, 127, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Mao, K.; Chen, S.; Chen, M.; Ma, Y.; Wang, Y.; Huang, B.; He, Z.; Zeng, Y.; Hu, Y.; Sun, S.; et al. Nitric oxide suppresses NLRP3 inflammasome activation and protects against LPS-induced septic shock. Cell Res. 2013, 23, 201–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perreault, M.; Marette, A. Targeted disruption of inducible nitric oxide synthase protects against obesity-linked insulin resistance in muscle. Nat. Med. 2001, 7, 1138–1143. [Google Scholar] [CrossRef]
- Spruss, A.; Kanuri, G.; Uebel, K.; Bischoff, S.C.; Bergheim, I. Role of the Inducible Nitric Oxide Synthase in the Onset of Fructose-Induced Steatosis in Mice. Antioxid. Redox Signal. 2011, 14, 2121–2135. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, A.; Marette, A. Inducible Nitric Oxide Synthase Induction Underlies Lipid-Induced Hepatic Insulin Resistance in Mice: Potential Role of Tyrosine Nitration of Insulin Signaling Proteins. Diabetes 2010, 59, 861. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.E.; Ko, M.S.; Yun, J.-Y.; Kim, M.-O.; Kim, J.H.; Park, H.S.; Kim, A.-R.; Kim, H.-J.; Kim, B.J.; Ahn, Y.E.; et al. Nitric Oxide Produced by Macrophages Inhibits Adipocyte Differentiation and Promotes Profibrogenic Responses in Preadipocytes to Induce Adipose Tissue Fibrosis. Diabetes 2016, 65, 2516–2528. [Google Scholar] [CrossRef] [Green Version]
- Dallaire, P.; Bellmann, K.; Laplante, M.; Gélinas, S.; Centeno-Baez, C.; Penfornis, P.; Peyot, M.-L.; Latour, M.G.; Lamontagne, J.; Trujillo, M.E.; et al. Obese mice lacking inducible nitric oxide synthase are sensitized to the metabolic actions of peroxisome proliferator-activated receptor-gamma agonism. Diabetes 2008, 57, 1999–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, H.-N.; Song, S.E.; Kim, Y.-W.; Kim, J.-Y.; Won, K.-C.; Park, S.-Y. Lack of inducible nitric oxide synthase prevents lipid-induced skeletal muscle insulin resistance without attenuating cytokine level. J. Pharmacol. Sci. 2011, 117, 77–86. [Google Scholar] [CrossRef]
- Nakata, S.; Tsutsui, M.; Shimokawa, H.; Suda, O.; Morishita, T.; Shibata, K.; Yatera, Y.; Sabanai, K.; Tanimoto, A.; Nagasaki, M.; et al. Spontaneous myocardial infarction in mice lacking all nitric oxide synthase isoforms. Circulation 2008, 117, 2211–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanuri, B.N.; Kanshana, J.S.; Rebello, S.C.; Pathak, P.; Gupta, A.P.; Gayen, J.R.; Jagavelu, K.; Dikshit, M. Altered glucose and lipid homeostasis in liver and adipose tissue pre-dispose inducible NOS knockout mice to insulin resistance. Sci. Rep. 2017, 7, 41009. [Google Scholar] [CrossRef] [PubMed]
- Kakimoto, P.A.; Chausse, B.; Caldeira da Silva, C.C.; Donato Júnior, J.; Kowaltowski, A.J. Resilient hepatic mitochondrial function and lack of iNOS dependence in diet-induced insulin resistance. PLoS ONE 2019, 14, e0211733. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, M.; Shimizu, N.; Kunii, K.; Martyn, J.A.J.; Ueki, K.; Kaneki, M. A Role for iNOS in Fasting Hyperglycemia and Impaired Insulin Signaling in the Liver of Obese Diabetic Mice. Diabetes 2005, 54, 1340–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khedara, A.; Kawai, Y.; Kayashita, J.; Kato, N. Feeding Rats the Nitric Oxide Synthase Inhibitor, L-NωNitroarginine, Elevates Serum Triglyceride and Cholesterol and Lowers Hepatic Fatty Acid Oxidation. J. Nutr. 1996, 126, 2563–2567. [Google Scholar] [CrossRef] [Green Version]
- Goto, T.; Ohnomi, S.; Khedara, A.; Kato, N.; Ogawa, H.; Yanagita, T. Feeding the nitric oxide synthase inhibitor L-N(omega)nitroarginine elevates serum very low density lipoprotein and hepatic triglyceride synthesis in rats. J. Nutr. Biochem. 1999, 10, 274–278. [Google Scholar] [CrossRef]
- Aggarwal, H.; Kanuri, B.N.; Dikshit, M. Role of iNOS in Insulin Resistance and Endothelial Dysfunction. In Oxidative Stress in Heart Diseases; Springer: Singapore, 2019; pp. 461–482. [Google Scholar]
- Milsom, A.B.; Fernandez, B.O.; Garcia-Saura, M.F.; Rodriguez, J.; Feelisch, M. Contributions of Nitric Oxide Synthases, Dietary Nitrite/Nitrate, and Other Sources to the Formation of NO Signaling Products. Antioxid. Redox Signal. 2012, 17, 422–432. [Google Scholar] [CrossRef]
- Cha, H.-N.; Kim, Y.-W.; Kim, J.-Y.; Kim, Y.-D.; Song, I.H.; Min, K.-N.; Park, S.-Y. Lack of inducible nitric oxide synthase does not prevent aging-associated insulin resistance. Exp. Gerontol. 2010, 45, 711–718. [Google Scholar] [CrossRef]
- Ghasemi, A.; Jeddi, S. Anti-obesity and anti-diabetic effects of nitrate and nitrite. Nitric Oxide 2017, 70, 9–24. [Google Scholar] [CrossRef]
- Kleemann, R.; van Erk, M.; Verschuren, L.; van den Hoek, A.M.; Koek, M.; Wielinga, P.Y.; Jie, A.; Pellis, L.; Bobeldijk-Pastorova, I.; Kelder, T.; et al. Time-Resolved and Tissue-Specific Systems Analysis of the Pathogenesis of Insulin Resistance. PLoS ONE 2010, 5, e8817. [Google Scholar] [CrossRef]
- Jiang, H.; Torregrossa, A.C.; Potts, A.; Pierini, D.; Aranke, M.; Garg, H.K.; Bryan, N.S. Dietary nitrite improves insulin signaling through GLUT4 translocation. Free Radic. Biol. Med. 2014, 67, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Ohtake, K.; Nakano, G.; Ehara, N.; Sonoda, K.; Ito, J.; Uchida, H.; Kobayashi, J. Dietary nitrite supplementation improves insulin resistance in type 2 diabetic KKAy mice. Nitric Oxide 2015, 44, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ables, G.P.; Perrone, C.E.; Orentreich, D.; Orentreich, N. Methionine-Restricted C57BL/6J Mice Are Resistant to Diet-Induced Obesity and Insulin Resistance but Have Low Bone Density. PLoS ONE 2012, 7, e51357. [Google Scholar] [CrossRef] [Green Version]
- Kanuri, B.N.; Rebello, S.C.; Pathak, P.; Agarwal, H.; Kanshana, J.S.; Awasthi, D.; Gupta, A.P.; Gayen, J.R.; Jagavelu, K.; Dikshit, M. Glucose and lipid metabolism alterations in liver and adipose tissue pre-dispose p47phox knockout mice to systemic insulin resistance. Free Radic. Res. 2018, 52, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Sadaf, S.; Singh, A.K.; Awasthi, D.; Nagarkoti, S.; Agrahari, A.K.; Srivastava, R.N.; Jagavelu, K.; Kumar, S.; Barthwal, M.K.; Dikshit, M. Augmentation of iNOS expression in myeloid progenitor cells expedites neutrophil differentiation. J. Leukoc. Biol. 2019, 106, 397–412. [Google Scholar] [CrossRef]
- Kanshana, J.S.; Rebello, S.C.; Pathak, P.; Kanuri, B.N.; Aggarwal, H.; Srivastava, V.; Khanna, V.; Singh, V.; Jagavelu, K.; Barthwal, M.K.; et al. Standardized fraction of Xylocarpus moluccensis fruits improve vascular relaxation and plaque stability in dyslipidemic models of atherosclerosis. J. Ethnopharmacol. 2018, 213, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Brial, F.; Le Lay, A.; Hedjazi, L.; Tsang, T.; Fearnside, J.F.; Otto, G.W.; Alzaid, F.; Wilder, S.P.; Venteclef, N.; Cazier, J.-B.; et al. Systems Genetics of Hepatic Metabolome Reveals Octopamine as a Target for Non-Alcoholic Fatty Liver Disease Treatment. Sci. Rep. 2019, 9, 3656. [Google Scholar] [CrossRef] [Green Version]
- Becares, N.; Gage, M.C.; Voisin, M.; Shrestha, E.; Martin-Gutierrez, L.; Liang, N.; Louie, R.; Pourcet, B.; Pello, O.M.; Luong, T.V.; et al. Impaired LXRα Phosphorylation Attenuates Progression of Fatty Liver Disease. Cell Rep. 2019, 26, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, Y.; Fujita, K.; Wada, K.; Yoneda, M.; Kessoku, T.; Shinohara, Y.; Imajo, K.; Ogawa, Y.; Nakamuta, M.; Saito, S.; et al. Deficiency of iNOS-derived NO accelerates lipid accumulation-independent liver fibrosis in non-alcoholic steatohepatitis mouse model. BMC Gastroenterol. 2015, 15, 42. [Google Scholar] [CrossRef] [Green Version]
- Kubant, R.; Poon, A.N.; Sánchez-Hernández, D.; Domenichiello, A.F.; Huot, P.S.P.; Pannia, E.; Cho, C.E.; Hunschede, S.; Bazinet, R.P.; Anderson, G.H. A comparison of effects of lard and hydrogenated vegetable shortening on the development of high-fat diet-induced obesity in rats. Nutr. Diabetes 2015, 5, e188. [Google Scholar] [CrossRef]
- Khedara, A.; Goto, T.; Morishima, M.; Kayashita, J.; Kato, N. Elevated Body Fat in Rats by the Dietary Nitric Oxide Synthase Inhibitor, L-Nω Nitroarginine. Biosci. Biotechnol. Biochem. 1999, 63, 698–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadowska, J.; Gębczyński, A.K.; Konarzewski, M. Metabolic risk factors in mice divergently selected for BMR fed high fat and high carb diets. PLoS ONE 2017, 12, e0172892. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Beg, M.; Kumar, D.; Shankar, K.; Varshney, S.; Rajan, S.; Srivastava, A.; Singh, K.; Sonkar, S.; Mahdi, A.A.; et al. Chronic hyper-leptinemia induces insulin signaling disruption in adipocytes: Implications of NOS2. Free Radic. Biol. Med. 2017, 112, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Li, P.; Pferdekamper, J.; Fan, W.; Saberi, M.; Schenk, S.; Olefsky, J.M. Inducible nitric oxide synthase deficiency in myeloid cells does not prevent diet-induced insulin resistance. Mol. Endocrinol. 2010, 24, 1413–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenopoulou, M.; Doulias, P.-T.; Nakamoto, K.; Berrios, K.; Zura, G.; Li, C.; Faust, M.; Yakovishina, V.; Evans, P.; Tan, L.; et al. Oral nitrite restores age-dependent phenotypes in eNOS-null mice. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Gheibi, S.; Jeddi, S.; Carlström, M.; Gholami, H.; Ghasemi, A. Effects of long-term nitrate supplementation on carbohydrate metabolism, lipid profiles, oxidative stress, and inflammation in male obese type 2 diabetic rats. Nitric Oxide 2018, 75, 27–41. [Google Scholar] [CrossRef]
- Larsen, F.J.; Schiffer, T.A.; Ekblom, B.; Mattsson, M.P.; Checa, A.; Wheelock, C.E.; Nyström, T.; Lundberg, J.O.; Weitzberg, E. Dietary nitrate reduces resting metabolic rate: A randomized, crossover study in humans. Am. J. Clin. Nutr. 2014, 99, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Hezel, M.P.; Liu, M.; Schiffer, T.A.; Larsen, F.J.; Checa, A.; Wheelock, C.E.; Carlström, M.; Lundberg, J.O.; Weitzberg, E. Effects of long-term dietary nitrate supplementation in mice. Redox Biol. 2015, 5, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konrad, D.; Wueest, S. The Gut-Adipose-Liver Axis in the Metabolic Syndrome. Physiology 2014, 29, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Penfornis, P.; Marette, A. Inducible nitric oxide synthase modulates lipolysis in adipocytes. J. Lipid Res. 2005, 46, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puigserver, P.; Rhee, J.; Donovan, J.; Walkey, C.J.; Yoon, J.C.; Oriente, F.; Kitamura, Y.; Altomonte, J.; Dong, H.; Accili, D.; et al. Insulin-regulated hepatic gluconeogenesis through FOXO1–PGC-1α interaction. Nature 2003, 423, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Camporez, J.-P.G.; Kursawe, R.; Titchenell, P.M.; Zhang, D.; Perry, C.J.; Jurczak, M.J.; Abudukadier, A.; Han, M.S.; Zhang, X.-M.; et al. Hepatic acetyl CoA links adipose tissue inflammation to hepatic insulin resistance and type 2 diabetes. Cell 2015, 160, 745–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aggarwal, H.; Pathak, P.; Singh, P.; Gayen, J.R.; Jagavelu, K.; Dikshit, M. Systemic Insulin Resistance and Metabolic Perturbations in Chow Fed Inducible Nitric Oxide Synthase Knockout Male Mice: Partial Reversal by Nitrite Supplementation. Antioxidants 2020, 9, 736. https://doi.org/10.3390/antiox9080736
Aggarwal H, Pathak P, Singh P, Gayen JR, Jagavelu K, Dikshit M. Systemic Insulin Resistance and Metabolic Perturbations in Chow Fed Inducible Nitric Oxide Synthase Knockout Male Mice: Partial Reversal by Nitrite Supplementation. Antioxidants. 2020; 9(8):736. https://doi.org/10.3390/antiox9080736
Chicago/Turabian StyleAggarwal, Hobby, Priya Pathak, Pragati Singh, Jiaur R. Gayen, Kumaravelu Jagavelu, and Madhu Dikshit. 2020. "Systemic Insulin Resistance and Metabolic Perturbations in Chow Fed Inducible Nitric Oxide Synthase Knockout Male Mice: Partial Reversal by Nitrite Supplementation" Antioxidants 9, no. 8: 736. https://doi.org/10.3390/antiox9080736