A Reference List of Phenolic Compounds (Including Stilbenes) in Grapevine (Vitis vinifera L.) Roots, Woods, Canes, Stems, and Leaves


Abstract
:1. Introduction
2. The Vegetative Organs of the Vine Plant
2.1. Roots
2.2. Woods
2.3. Canes
2.4. Stems
2.5. Leaves
3. Extraction, Separation, and Identification of Phenolic Compounds in Grapevine
4. Polyphenols (Excluding Stilbenes) Identified in the Vegetative Organs of Grapevine
4.1. Hydroxycinnamic Acids
4.2. Hydroxybenzoic Acids
4.3. Flavan-3-Ols or Flavanols
4.4. Proanthocyanidins
4.5. Anthocyanins
4.6. Flavones
4.7. Flavonols
- -
- Myricetin, myricetin-3-O-galactoside, myricetin-3-O-glucuronide, myricetin-3-O-glucoside, and myricetin-3-O-rhamnoside;
- -
- Quercetin, quercetin-3-O-rutinoside, quercetin-3-O-galactoside, quercetin-3-O-glucoside, quercetin-3-O-glucuronide, quercetin-3-O-rhamnoside, quercetin-3-(6-O-acetyl)glucoside, quercetin-3-(3-O-arabinosyl)glucoside, quercetin-3-(7-O-glucosyl)glucuronide, quercetin-3-O-arabinose (reported as quercetin-O-pentoside by some authors), quercetin-3-(3-O-rhamnosyl)glucoside-7-O-rhamnoside, quercetin-3-(6-O-rhamnosyl)galactoside, and diquercetin-3-(3-O-glucosyl)glucuronide;
- -
- Kaempferol, kaempferol-3-O-galactoside, kaempferol-3-O-rutinoside, kaempferol-3-O-glucuronide, kaempferol-3-O-glucoside, kaempferol-3-O-xyloside (or kaempferol-O-pentoside by some authors), kaempferol-3-O-rhamnoside, dihydrokaempferol-3-O-rhamnoside, kaempferol-3-(6-O-coumaroyl)glucoside, and kaempferol-3-(7-O-glucosyl)galactoside (or kaempferol-3,7-diglucoside by some authors);
- -
- Isorhamnetin-3-O-galactoside, isorhamnetin-3-O-glucoside, isorhamnetin-3-O-arabinose (or isorhamnetin-O-pentoside by some authors), isorhamnetin-3-O-glucuronide, isorhamnetin-3-O-rutinoside, isorhamnetin-3-(6-O-feruloyl)glucoside, and isorhamnetin-3-(4-O-rhamnosyl)rutinoside (or isorhamnetin diglycoside by some authors).
4.8. Flavanones
4.9. Coumarins and Dihydrochalcones
4.10. Non-Phenolic Compounds
5. Stilbenic Compounds Identified in the Vegetative Organs of Grapevine
5.1. Monomeric Stilbenes
5.2. Dimeric Stilbenes
5.3. Trimeric Stilbenes
5.4. Tetrameric Stilbenes
5.5. Pentameric Stilbenes
5.6. Hexameric Stilbenes
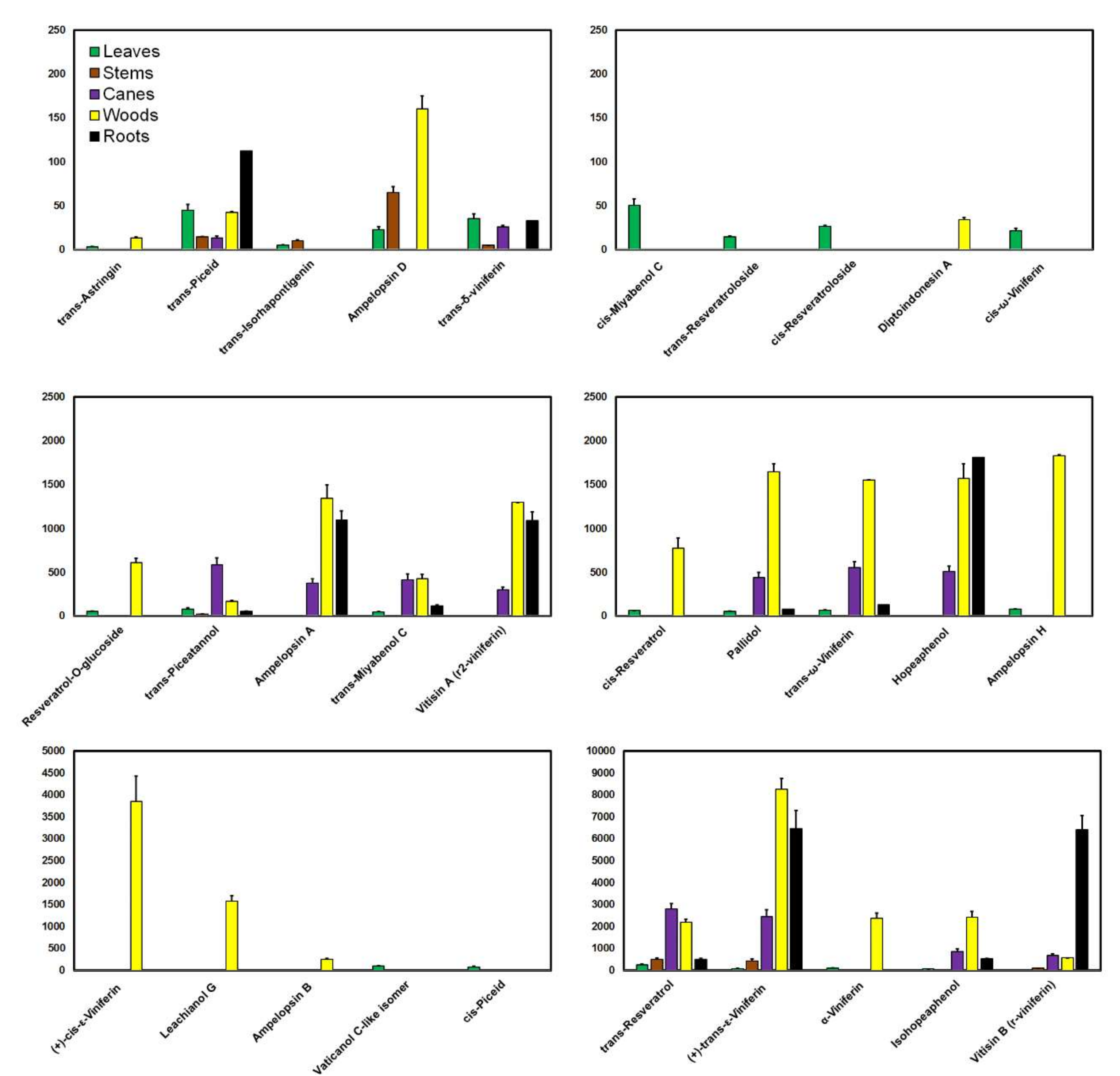
6. Levels of Phenolic Compounds in the Leaves, Stems, Canes, Woods, and Roots of the Vine Plant
6.1. Phenolic Compounds in Grapevine Leaves
6.2. Phenolic Compounds in Grapevine Stems
6.3. Phenolic Compounds in Grapevine Canes
6.4. Phenolic Compounds in Grapevine Woods
6.5. Phenolic Compounds in Grapevine Roots
7. Distribution of Phenolic Compounds in the Vegetative Organs of Grapevine
8. Factors Affecting the Biosynthesis and Levels of Phenolic Compounds in the Vegetative Organs of Grapevine
8.1. Grapevine Cultivars and Rootstocks
8.2. Geographical Location and Climate (Shading, Temperature, Irrigation)
8.3. Vinicultural Practices
8.4. Outside and Biotic Stimuli
9. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hmamouchi, M.; Es-Safi, N.; Lahrichi, M.; Fruchier, A.; Essassi, E.M. Flavones and flavonols in leaves of some Moroccan Vitis vinifera cultivars. Am. J. Enol. Viticult. 1996, 47, 186–192. [Google Scholar]
- Harb, J.; Alseekh, S.; Tohge, T.; Fernie, R.A. Profiling of primary metabolites and flavonols in leaves of two table grape varieties collected from semiarid and temperate regions. Phytochemistry 2015, 117, 444–455. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.M.M.; Felgueiras, M.L.; Cunh, A.; Chicau, G.; Ferreres, F.; Dias, C.P.A. Differential phenolic production in leaves of Vitis vinifera cv. Alvarinho affected with esca disease. Plant Physiol. Biochem. 2017, 112, 45–52. [Google Scholar] [CrossRef]
- Handoussa, H.; Hanafi, R.; El-Khatib, A.; Linscheid, M.; Mahran, L.; Ayoub, N. Computer-assisted HPLC method development using DryLab for determination of major phenolic components in Corchorus olitorius and Vitis vinifera by using HPLC-PDA-ESI-TOF- MSn. Res. Rev. J. Bot. Sci. 2017, 6, 9–16. [Google Scholar]
- Dresch, R.R.; Dresch, K.M.; Guerreiro, F.A.; Biegelmeyer, R.; Holzschuh, M.H.; Rambo, F.D.; Henriques, T.A. Phenolic compounds from the leaves of Vitis labrusca and Vitis vinifera L. as a source of waste byproducts: Development and validation of LC method and antichemotactic activity. Food Anal. Methods 2014, 7, 527–539. [Google Scholar] [CrossRef]
- Leeuwen, V.C.; Roby, J.P.; Villaverde, A.V.; Gindro, K. Impact of clonal variability in Vitis vinifera cabernet franc on grape composition, wine quality, leaf blade stilbene content, and downy mildew resistance. J. Agric. Food Chem. 2013, 61, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Pawlus, D.A.; Sahli, R.; Bisson, J.; Rivière, C.; Delaunay, J.C.; Richard, T.; Gomès, E.; Bordenave, L.; Waffo-Téguo, P.; Mérillon, J.M. Stilbenoid profiles of canes from Vitis and Muscadinia species. J. Agric. Food Chem. 2013, 61, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Pawlus, D.A.; Waffo-Téguo, P.; Shaver, J.; Mérillon, J.M. Stilbenoid chemistry from wine and the genus vitis, a review. J. Int. Sci. Vigne. Vin. 2012, 46, 57–111. [Google Scholar] [CrossRef]
- Wallis, M.C.; Wallingford, K.A.; Chen, J.C. Effects of cultivar, phenology, and Xylella fastidiosa infection on grapevine xylem sap and tissue phenolic content. Physiol. Mol. Plant Pathol. 2013, 84, 28–35. [Google Scholar] [CrossRef]
- Jean-Denis, J.B.; Pezet, R.; Tabacchi, R. Rapid analysis of stilbenes and derivatives from downy mildew-infected grapevine leaves by liquid chromatography–atmospheric pressure photoionisation mass spectrometry. J. Chromatogr. A 2006, 1112, 263–268. [Google Scholar] [CrossRef]
- Schnee, S.; Viret, O.; Gindro, K. Role of stilbenes in the resistance of grapevine to powdery mildew. Physiol. Mol. Plant Pathol. 2008, 72, 128–133. [Google Scholar] [CrossRef]
- Gabaston, J.; Villar, C.E.; Biais, B.; Teguo, W.P.; Renouf, E.; Corio-Costet, M.F.; Richard, T.; Mérillon, J.M. Stilbenes from Vitis vinifera L. waste: A sustainable tool for controlling Plasmopara Viticola. J. Agric. Food Chem. 2017, 65, 2711–2718. [Google Scholar] [CrossRef] [PubMed]
- Amalfitano, C.; Agrelli, D.; Arrigo, A.; Mugnai, L.; Surico, G.; Evidente, A. Stilbene polyphenols in the brown red wood of Vitis vinifera cv. Sangiovese affected by “esca proper”. Phytopathol. Mediterr. 2011, 50, S224–S235. [Google Scholar]
- Bavaresco, L.; Fregoni, C. Physiological role and molecular aspects of grapevine stilbenic compounds. In Molecular Biology & Biotechnology of the Grapevine, Chapter 5, 1st ed.; Roubelakis-Angelakis, K.A., Ed.; Kluwer Academic Publishers: Berlin, Germany, 2001; pp. 153–182. [Google Scholar]
- Goufo, P.; Marques, C.A.; Cortez, I. Exhibition of local but not systemic induced phenolic defenses in Vitis vinifera L. affected by brown wood streaking, grapevine leaf stripe, and apoplexy (Esca complex). Plants 2019, 8, 412. [Google Scholar] [CrossRef] [Green Version]
- Flamini, R.; De Rosso, M. High-resolution mass spectrometry and biological properties of grapevine and wine stilbenoids. In Studies in Natural Products Chemistry; Chapter 5; Rahman, A., Ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2018; Volume 61, pp. 175–210. [Google Scholar]
- Gomès, E.; Coutos-Thévenot, P. Molecular aspects of grapevine-pathogenic fungi interactions. In Grapevine Molecular Physiology & Biotechnology, 2nd ed.; Chapter 15; Roubelakis-Angelakis, K.A., Ed.; Springer Science & Business Media B.V.: Dordrecht, The Netherlands, 2009; pp. 407–428. [Google Scholar]
- Singh, K.R.; Soares, B.; Goufo, P.; Castro, I.; Cosme, F.; Pinto-Sintra, L.A.; Inês, A.; Oliveira, A.A.; Falco, V. Chitosan upregulates the genes of the ROS pathway and enhances the antioxidant potential of grape (Vitis vinifera L. ‘Touriga Franca’ and ’Tinto Cão’) tissues. Antioxidants 2019, 8, 525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aouey, B.; Samet, M.A.; Fetoui, H.; Simmonds, S.J.M.; Bouaziz, M. Anti-oxidant, anti-inflammatory, analgesic and antipyretic activities of grapevine leaf extract (Vitis vinifera) in mice and identification of its active constituents by LC–MS/MS analyses. Biomed. Pharmacother. 2016, 84, 1088–1098. [Google Scholar] [CrossRef]
- Fernandes, F.; Ramalhosa, E.; Verdial, J.; Valentao, P.; Andrade, P.; Bento, A.; Pereira, A.J. Vitis vinifera leaves towards bioactivity. Ind. Crops Prod. 2015, 43, 434–440. [Google Scholar] [CrossRef]
- Apostolou, A.; Stagos, D.; Galitsiou, E.; Spyrou, A.; Haroutounian, S.; Portesis, N.; Trizoglou, I.; Hayes, A.W.; Tsatsakis, M.A.; Kouretas, D. Assessment of polyphenolic content, antioxidant activity, protection against ROS-induced DNA damage and anticancer activity of Vitis vinifera stem extracts. Food Chem. Toxicol. 2013, 61, 60–68. [Google Scholar] [CrossRef]
- Ramalhosa, E.; Valentão, P.; Andrade, P.; Verdial, J.; Bentoa, A.; Pereira, A.J. Phenolic composition and antioxidant potential of Vitis vinifera leaves of six varieties harvested in northeast of Portugal. FaBE Food Biosyst. Eng. 2013, 2, 169–176. [Google Scholar]
- Wei, Y.J.; Zhao, S.R.; Li, J.M.; Xue, B. Stilbene profiles in different tissues of Vitis vinifera L. cv. Cabernet Sauvignon and a comparison of their antioxidant activity. Aust. J. Grape wine Res. 2016, 22, 226–231. [Google Scholar] [CrossRef]
- Biais, B.; Krisa, S.; Cluzet, S.; Da Costa, G.; Waffo-Téguo, P.; Mérillon, J.M.; Richard, T. Antioxidant and cytoprotective activities of grapevine stilbenes. J. Agric. Food Chem. 2017, 65, 4952–4960. [Google Scholar] [CrossRef] [PubMed]
- Macke, S.; Jerz, G.; Empl, T.M.; Steinberg, P.; Winterhalter, P. Activity-guided isolation of resveratrol oligomers from a grapevine-shoot extract using counter current chromatography. J. Agric. Food Chem. 2012, 60, 11919–11927. [Google Scholar] [CrossRef] [PubMed]
- Papastamoulis, Y.; Richard, T.; Nassra, M.; Badoc, A.; Krisa, S.; Harakat, D.; Monti, J.P.; Mérillon, J.M.; Téguo, W.P. Viniphenol A, a complex resveratrol hexamer from Vitis vinifera stalks: Structural elucidation and protective effects against amyloid-β-induced toxicity in PC12 cells. J. Nat. Prod. 2014, 77, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Pflieger, A.; Waffo-Téguo, P.; Papastamoulis, Y.; Chaignepain, S.; Subra, F.; Munir, S.; Delelis, O.; Lesbats, P.; Calmels, C.; Andreola, M.L.; et al. Natural stilbenoids isolated from grapevine exhibiting inhibitory effects against HIV-1 integrase and eukaryote MOS1 transposase in vitro activities. PLoS ONE 2013, 8, e81184. [Google Scholar] [CrossRef]
- Choi, Y.H.; Yoo, M.Y.; Choi, C.W.; Cha, M.R.; Yon, G.H.; Kwon, D.Y.; Kim, Y.S.; Park, W.K.; Ryu, S.Y. A new specific BACE-1 inhibitor from the stembark extract of Vitis vinifera. Planta Med. 2009, 75, 537–740. [Google Scholar] [CrossRef]
- Kim, Y.M.; Lee, E.W.; Eom, S.H.; Kim, T.H. Pancreatic lipase inhibitory stilbenoids from the roots of Vitis vinifera. Int. J. Food Sci. Nutr. 2014, 65, 97–100. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Ewald, P.; Yasui, Y.; Yokokawa, H.; Wagner, E.A.; Matsugo, S.; Winterhalter, P.; Rimbach, G. Chemical characterization, free radical scavenging, and cellular antioxidant and anti-inflammatory properties of a stilbenoid-rich root extract of Vitis vinifera. Oxid. Med. Cell. Longev. 2016, 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Waffo-Teguo, P.; Krisa, S.; Richard, T.; Mérillon, J.M. Grapevine stilbenes and their biological effects. In Bioactive Molecules and Medicinal Plants; Chapter 2; Gopal Ramawat, K., Merillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 25–54. [Google Scholar]
- Pantelić, M.M.; Dabić Zagorac, Č.D.; Ćirić, Ž.I.; Pergal, V.M.; Relić, J.D.; Todić, R.S.; Natić, M.M. Phenolic profiles, antioxidant activity and minerals in leaves of different grapevine varieties grown in Serbia. J. Food Compos. Anal. 2017, 62, 76–83. [Google Scholar] [CrossRef]
- Barros, A.; Gironés-Vilaplana, A.; Teixeira, A.; Collado-González, J.; Moreno, A.D.; Gil-Izquierdo, A.; Rosa, E.; Domínguez-Perles, R. Evaluation of grape (Vitis vinifera L.) stems from Portuguese varieties as a resource of (poly)phenolic compounds: A comparative study. Food Res. Int. 2014, 65, 375–384. [Google Scholar] [CrossRef]
- Makris, P.D.; Boskou, G.; Andrikopoulos, K.N.; Kefalas, P. Characterisation of certain major polyphenolic antioxidants in grape (Vitis vinifera cv. Roditis) stems by liquid chromatography-mass spectrometry. Eur. Food Res. Technol. 2008, 226, 1075–1079. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F. Bioactive chemicals and health benefits of grapevine products. In Bioactive Foods in Promoting Health: Fruits and Vegetables; Chapter 38; Watson, R., Preedy, V., Eds.; Academic Press: Cambridge, MA, USA, 2010; pp. 581–620. [Google Scholar]
- Gindro, K.; Alonso-Villaverde, V.; Viret, O.; Spring, J.L.; Marti, G.; Wolfender, J.L.; Pezet, R. Stilbenes: Biomarkers of grapevine resistance to disease of high relevance for agronomy, oenology and human health. In Plant Defence: Biological Control. Progress in Biological Control; Chapter 2; Merillon, J.-M., Ramawat, K.G., Eds.; Springer Science & Business Media B.V.: Berlin, Germany, 2012; Volume 2, pp. 25–54. [Google Scholar]
- Spatafora, C.; Barbagallo, E.; Amico, V.; Tringali, C. Grape stems from Sicilian Vitis vinifera cultivars as a source of polyphenol-enriched fractions with enhanced antioxidant activity. LWT Food Sci. Technol. 2013, 54, 542–548. [Google Scholar] [CrossRef]
- Jeandet, P.; Jacquot, C.; Parmentier, M.; Conreux, A.; Sbaghi, M.; Adrian, M.; Waffo-Téguo, P. Role of stilbene dehydrodimers (δ- and ε-viniferins) in the Botrytis cinerea/grapevine interaction. In Macromolecules and Secondary Metabolites of Grapevine and Wines; Jeandet, P., Clément, C., Conreux, A., Eds.; Intercept Lavoisier: New York, NY, USA, 2007; pp. 105–110. [Google Scholar]
- Waffo-Teguo, P.; Krisa, S.; Pawlus, D.A.; Richard, T.; Monti, J.P.; Me’rillon, J.M. Grapevine stilbenoids: Bioavailability and neuroprotection. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Chapter 73; Gopal Ramawat, K., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2275–2309. [Google Scholar]
- Gorena, T.; Saez, V.; Mardones, C.; Vergara, C.; Winterhalter, P.; Von Baer, D. Influence of post-pruning storage on stilbenoid levels in Vitis vinifera. L. canes. Food Chem. 2014, 155, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Fujii, F.; He, Y.H.; Terashima, K.; Takaya, Y.; Niwa, M. Three new stilbeneoligomers from the roots of Vitis vinifera ‘Kyohou’. Heterocycles 2005, 65, 2461–2469. [Google Scholar]
- Reniero, F.; Rudolph, M.; Angioni, A.; Bernreuther, A.; Cabras, P.; Mattivi, F. Identification of two stilbenoids from Vitis roots. VITIS 1996, 35, 125–127. [Google Scholar]
- Martin, N.; Vesentini, D.; Rego, C.; Monteiro, S.; Oliveira, H.; Ferreira, B.R. Phaeomoniella chlamydospora infection induces changes in phenolic compounds content in Vitis vinifera. Phytopathol. Mediterr. 2009, 48, 101–116. [Google Scholar]
- Rusjan, D.; Persic, M.; Likar, M.; Biniari, K.; Mikulic-Petkovsek, M. Phenolic responses to esca-associated fungi in differently decayed grapevine woods from different trunk parts of ‘Cabernet Sauvignon’. J. Agric. Food Chem. 2017, 65, 6615–6624. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Bisson, J.; Waffo-Téguo, P.; Papastamoulis, Y.; Richard, T.; Corio-Costet, M.F.; Mérillon, J.M.; Cluzet, S. Phenolics and their antifungal role in grapevine wood decay: Focus on the Botryosphaeriaceae family. J. Agric. Food Chem. 2012, 60, 11859–11860. [Google Scholar] [CrossRef]
- Delgado-Torre, M.P.; Ferreiro-Vera, C.; Priego-Capote, F.; Pérez-Juan, M.P.; Luque de Castro, D.M. Comparison of accelerated methods for the extraction of phenolic compounds from different vine-shoot cultivars. J. Agric. Food Chem. 2012, 60, 3051–3060. [Google Scholar] [CrossRef]
- Max, B.; Salgado, J.M.; Cortés, S.; Domínguez, J.M. Extraction of phenolic acids by alkaline hydrolysis from the solid residue obtained after prehydrolysis of trimming vine shoots. J. Agric. Food Chem. 2010, 58, 1909–1917. [Google Scholar] [CrossRef]
- Sánchez-Gómez, R.; Zalacain, A.; Alonso, G.L.; Salinas, M.R. Effect of toasting on non-volatile and volatile vine-shoots low molecular weight phenolic compounds. Food Chem. 2016, 204, 499–505. [Google Scholar] [CrossRef]
- Sánchez-Gómez, R.; Zalacain, A.; Alonso, G.L.; Salinas, M.R. Vine-shoot waste aqueous extracts for re-use in agriculture obtained by different extraction techniques: Phenolic, volatile, and mineral compounds. J. Agric. Food Chem. 2014, 62, 10861–10872. [Google Scholar] [CrossRef] [PubMed]
- Sáez, V.; Gayoso, C.; Riquelme, S.; Pérez, J.; Vergara, C.; Mardones, C.; Von Baer, D. C18 core-shell column with in-series absorbance and fluorescence detection for simultaneous monitoring of changes in stilbenoid and proanthocyanidin concentrations during grape cane storage. J. Chromatogr. B 2018, 1074, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Richard, T.; Renouf, E.; Bisson, J.; Waffo-Téguo, P.; Bordenave, I.; Ollat, N.; Mérillon, J.M.; Cluzet, S. Comparative analyses of stilbenoids in canes of major Vitis vinifera L. cultivars. J. Agric. Food Chem. 2013, 61, 11392–11399. [Google Scholar] [CrossRef] [PubMed]
- Houillé, B.; Besseau, S.; Delanoue, G.; Oudin, A.; Papon, N.; Clastre, M.; Simkin, J.A.; Guérin, L.; Courdavault, V.; Giglioli-Guivarch, N.; et al. Composition and tissue-specific distribution of stilbenoids in grape canes are affected by downy mildew pressure in the vineyard. J. Agric. Food Chem. 2015, 63, 8472–8477. [Google Scholar] [CrossRef]
- Püssa, T.; Floren, J.; Kuldkepp, P.; Raal, A. Survey of grapevine Vitis vinifera stem polyphenols by Liquid Chromatography-Diode Array Detection-Tandem Mass Spectrometry. J. Agric. Food Chem. 2006, 54, 7488–7494. [Google Scholar]
- Dias, C.; Domínguez-Perles, R.; Aires, A.; Teixeira, A.; Rosa, E.; Barros, A.; Saavedra, J.M. Phytochemistry and activity against digestive pathogens of grape (Vitis vinifera L.) stem’s (poly)phenolic extracts. LWT Food Sci. Technol. 2015, 61, 25–32. [Google Scholar] [CrossRef]
- Wallis, M.C.; Chen, J.C. Grapevine phenolic compounds in xylem sap and tissues are significantly altered during infection by Xylella fastidiosa. Phytopathology 2012, 102, 816–826. [Google Scholar] [CrossRef] [Green Version]
- Yan, K.X.; Terashima, K.; Takaya, Y.; Niwa, M. A novel oligostilbene named (+)-viniferol A from the stem of Vitis vinifera ‘Kyohou’. Tetrahedron 2001, 57, 2711–2715. [Google Scholar] [CrossRef]
- Yan, K.X.; Terashima, K.; Takaya, Y.; Niwa, M. Two new stilbenetetramers from the stem of Vitis vinifera ‘Kyohou’. Tetrahedron 2002, 58, 6931–6935. [Google Scholar] [CrossRef]
- Ewald, P.; Delker, U.; Winterhalter, P. Quantification of stilbenoids in grapevine canes and grape cluster stems with a focus on long-term storage effects on stilbenoid concentration in grapevine canes. Food Res. Int. 2017, 100, 326–331. [Google Scholar] [CrossRef]
- Schoedl, K.; Forneck, A.; Sulyok, M.; Schuhmacher, R. Optimization, in-house validation, and application of a Liquid Chromatography_Tandem Mass Spectrometry (LC_MS/MS)-based method for the quantification of selected polyphenolic compounds in leaves of grapevine (Vitis vinifera L.). J. Agric. Food Chem. 2011, 59, 10787–10794. [Google Scholar] [CrossRef] [PubMed]
- Griesser, M.; Weingart, G.; Schoedl-Hummel, K.; Neumann, N.; Becker, M.; Varmuza, K.; Liebner, F.; Schuhmacher, R.; Forneck, A. Severe drought stress is affecting selected primary metabolites, polyphenols, and volatile metabolites in grapevine leaves (Vitis vinifera cv. Pinot noir). Plant Physiol. Biochem. 2015, 88, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Mattivi, F.; Vrhovsek, U.; Malacarne, G.; Masuero, D.; Zulini, L.; Stefanini, M.; Moser, C.; Velasco, R.; Guella, G. Profiling of resveratrol oligomers, important stress metabolites, accumulating in the leaves of hybrid Vitis vinifera (Merzling × Teroldego) genotypes infected with Plasmopara viticola. J. Agric. Food Chem. 2011, 59, 5364–5375. [Google Scholar] [CrossRef] [PubMed]
- Pezet, R.; Perret, C.; Jean-Denis, J.B.; Tabacchi, R.; Gindro, K.; Viret, O. δ-Viniferin, a resveratrol dehydrodimer: One of the major stilbenes synthesized by stressed grapevine leaves. J. Agric. Food Chem. 2003, 51, 5488–5492. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J. The Identification of stilbene compounds and the change of their contents in UV-irradiated grapevine leaves. Korean J. Hortic Sci. 2011, 29, 374–381. [Google Scholar]
- Tříska, J.; Vrchotová, N.; Olejníčková, J.; Jílek, R.; Sotolář, R. Separation and identification of highly fluorescent compounds derived from trans-resveratrol in the leaves of Vitis vinifera infected by Plasmopara viticola. Molecules 2012, 17, 2773–2783. [Google Scholar] [CrossRef] [Green Version]
- Braidot, E.; Zancani, M.; Petrussa, E.; Peresson, C.; Bertolini, A.; Patui, S.; Macrì, F.; Vianello, A. Transport and accumulation of flavonoids in grapevine (Vitis vinifera L.). Plant Signal. Behav. 2008, 3, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Tartaglione, L.; Gambuti, A.; De Cicco, P.; Ercolano, G.; Ianaro, A.; Taglialatela-Scafati, O.; Moio, L.; Forino, M. NMR-based phytochemical analysis of Vitis vinifera cv Falanghina leaves. Characterization of a previously undescribed biflavonoid with antiproliferative activity. Fitoterapia 2018, 125, 13–17. [Google Scholar] [CrossRef]
- Takaya, Y.; Terashima, K.; Yan, K.X.; Niwa, M. (+)-Viniferol D, a new stilbenetrimer from the stem of Vitis vinifera ‘Kyohou’. Heterocycles 2003, 60, 1433–1439. [Google Scholar] [CrossRef]
- Becker, L.; Carré, V.; Poutaraud, A.; Merdinoglu, D.; Chaimbault, P. MALDI mass spectrometry imaging for the simultaneous location of resveratrol, pterostilbene and viniferins on grapevine leaves. Molecules 2014, 19, 10587–10600. [Google Scholar] [CrossRef] [Green Version]
- Jordão, A.M.; Ricardo-da-Silva, J.M.; Laureano, O. Evolution of proanthocyanidins in bunch stems during berry development (Vitis vinifera L.). Vitis 2001, 40, 17–22. [Google Scholar]
- Bruisson, S.; Maillot, P.; Schellenbaum, P.; Walter, B.; Gindro, K.; Deglène-Benbrahim, L. Arbuscular mycorrhizal symbiosis stimulates key genes of the phenylpropanoid biosynthesis and stilbenoid production in grapevine leaves in response to downy mildew and grey mould infection. Phytochemistry 2016, 131, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Bavaresco, L.; Fregoni, C.; Gonçalves, M.I.; Vezzulli, S. Physiology & molecular biology of grapevine stilbenes: An update. In Grapevine Molecular Physiology & Biotechnology, 2nd ed.; Roubelakis-Angelakis, K.A., Ed.; Springer Science & Business Media B.V.: Dordrecht, The Netherlands, 2009; pp. 341–364. [Google Scholar]
- Montero, L.; Sáez, V.; Von Baer, D.; Cifuentes, A.; Herrero, M. Profiling of Vitis vinifera L. canes (poly)phenolic compounds using comprehensive two-dimensional liquid chromatography. J. Chromatogr. A 2018, 1536, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anđelković, M.; Radovanović, B.; Anđelković, M.A.; Radovanović, V. Phenolic compounds and bioactivity of healthy and infected grapevine leaf extracts from red varieties Merlot and Vranac (Vitis vinifera L.). Plant Foods Hum. Nutr. 2015, 70, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Karvela, E.; Makris, P.D.; Kalogeropoulos, N.; Karathanos, T.V. Deployment of response surface methodology to optimise recovery of grape (Vitis vinifera) stem polyphenols. Talanta 2009, 79, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Topalović, A.; Mikulič-Petkovšek, M.; Perović, N.; Trifunović, S.; Knežević, M. Phenolic composition of the leaf of grapevine cv. cardinal. Agr. Forest. 2012, 52, 5–15. [Google Scholar]
- Huang, Y.F.; Cheynier, V.; Terrier, N. Shedding light on the black boxes of the proanthocyanidin pathway with grapevine. In Recent Advances in Polyphenol Research, 1st ed.; Chapter 7; Cheynier, V., Sarni-Manchado, P., Quideau, S., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; Volume 3, pp. 161–190. [Google Scholar]
- Cui, Z.H.; Bi, W.L.; Hao, X.Y.; Li, P.M.; Duan, Y.; Walker, M.A.; Xu, Y.; Wang, Q.C. Drought stress enhances up-regulation of anthocyanin biosynthesis in grapevine leafroll-associated virus 3-infected in vitro grapevine (Vitis vinifera) leaves. Plant Dis. 2017, 101, 1606–1615. [Google Scholar] [CrossRef] [Green Version]
- Caramanico, L.; Rustioni, L.; De Lorenzis, G. Iron deficiency stimulates anthocyanin accumulation in grapevine apical leaves. Plant Physiol. Biochem. 2017, 119, 286–293. [Google Scholar] [CrossRef]
- Schneider, E.; Heydt, V.H.; Esperester, A. Evaluation of polyphenol composition in red leaves from different varieties of Vitis vinifera. Planta Med. 2008, 74, 565–572. [Google Scholar] [CrossRef]
- Park, H.J.; Lim, C.E.; Park, C.W.; Cha, H.C. Analysis of flavonols extracted from leaves of various grapevine cultivars by HPLC. Hortic. Environ. Biot. 2004, 45, 138–142. [Google Scholar]
- Chitarrini, G.; Zulini, L.; Masuero, D.; Vrhovsek, U. Lipid, phenol and carotenoid changes in ‘Bianca’ grapevine leaves after mechanical wounding: A case study. Protoplasma 2017, 254, 2095–2106. [Google Scholar] [CrossRef] [PubMed]
- Vezzulli, S.; Malacarne, G.; Masuero, D.; Vecchione, A.; Dolzani, C.; Goremykin, V.; Mehari, H.Z.; Banchi, E.; Velasco, R.; Stefanini, M.; et al. The Rpv3-3 haplotype and stilbenoid induction mediate downy mildew resistance in a grapevine interspecific population. Front. Plant Sci. 2019, 10, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luque-Rodríguez, J.M.; Pérez-Juan, P.; Luque de Castro, M.D. Extraction of polyphenols from vine shoots of Vitis vinifera by superheated ethanol-water mixtures. J. Agric. Food Chem. 2006, 54, 8775–8781. [Google Scholar] [CrossRef] [PubMed]
- Schoedl, K.; Schuhmacher, R.; Forneck, A. Studying the polyphenols of grapevine leaves according to age and insertion level under controlled conditions. Sci. Hortic. 2012, 141, 37–41. [Google Scholar] [CrossRef]
- Kocsis, M.; Abrankó, L.; Ayaydin, F.; Csepregi, K.; Papp, N.; Teszlák, P.; Jakab, G. Main leaf polyphenolic components of berry color variant grapevines and their acclimative responses to sunlight exposure. Appl. Sci. 2015, 5, 1955–1969. [Google Scholar] [CrossRef] [Green Version]
- Martín, L.; Fontaine, F.; Castañoa, J.F.; Songy, A.; Roda, R.; Vallet, J.; Ferrer-Gallego, R. Specific profile of Tempranillo grapevines related to Esca-leaf symptoms and climate conditions. Plant Physiol. Biochem. 2019, 135, 575–587. [Google Scholar] [CrossRef]
- Farhadi, K.; Esmaeilzadeh, F.; Hatami, M.; Forough, M.; Molaie, R. Determination of phenolic compounds content and antioxidant activity in skin, pulp, seed, cane and leaf of five native grape cultivars in West Azerbaijan province, Iran. Food Chem. 2016, 199, 847–855. [Google Scholar] [CrossRef]
- Taware, B.P.; Dhumal, N.K.; Oulkar, P.D.; Patil, H.S.; Banerjee, K. Phenolic alterations in grape leaves, berries and wines due to foliar and cluster powdery mildew infections. Int. J. Pharma Bio. Sci. 2010, 1, 1–14. [Google Scholar]
- Vrhovsek, U.; Malacarne, G.; Masuero, D.; Zulini, I.; Guella, G.; Stefanini, M.; Velasco, R.; Mattivi, F. Profiling and accurate quantification of trans-resveratrol, trans-piceid, trans-pterostilbene and 11 viniferins induced by Plasmopara viticola in partially resistant grapevine leaves. Aust. J. Grape wine Res. 2012, 18, 11–19. [Google Scholar] [CrossRef]
- Mikeš, O.; Vrchotová, N.; Tříska, J.; Kyseláková, M.; Šmidrkal, J. Distribution of major polyphenolic compounds in vine grapes of different cultivars growing in south Moravian vineyards. Czech J. Food Sci. 2008, 26, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Piñeiro, Z.; Guerrero, R.F.; Fernández-Marin, M.I.; Cantos-Villar, E.; Palma, M. Ultrasound-assisted extraction of stilbenoids from grape stems. J. Agric. Food Chem. 2013, 61, 12549–12556. [Google Scholar] [CrossRef] [PubMed]
- Bavaresco, L.; Mattivi, F.; De Rosso, M.; Flamini, R. Effects of elicitors, viticultural factors, and enological practices on resveratrol and stilbenes in grapevine and wine. Mini-Rev. Med. Chem. 2012, 12, 1366–1381. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| ID 1 | Compound Group | Compound Name 1 | Chemical Formula 2 | MW (g/mol) | [M–H]– | Main MS/MS Fragments (m/z) 3,4 | λmax (CH3OH) (nm) | Detection Mode 5 | Tissue Distribution 6 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LEAVES | STEMS | CANES | WOODS | ROOTS | |||||||||
| 1 | Hydroxybenzoic acid | Quinic acid | C7H12O6 | 192.167 | 191 | 111, 173, 85, 127 | 308, 280 | MS | ✓ | ||||
| 2 | Hydroxybenzoic acid | Gallic acid | C7H6O5 | 170.120 | 169 | 125, 124, 79, 51 | 278, 214 | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 4 | Hydroxybenzoic acid | Protocatechuic acid | C7H6O4 | 154.121 | 153 | 109, 108 | 297, 258 | MS | ✓ | ✓ | |||
| 7 | Hydroxybenzoic acid | p-Hydroxybenzoic acid | C7H6O3 | 138.122 | 137 | 93, 60, 79, 108, 118, 137 | 272, 310sh | MS | ✓ | ✓ | |||
| 8 | Hydroxybenzoic acid | Gentisic acid | C7H6O4 | 154.121 | 153 | 109, 81, 42, 108 | 281, 228, 330sh | MS | ✓ | ||||
| 9 | Hydroxybenzoic acid | γ-Resorcylic acid | C7H6O4 | 154.121 | 153 | 109, 136, 154, 110, 52, 80, 137, 39, 155 | 313, 245 | MS | ✓ | ||||
| 30 | Hydroxybenzoic acid | Vanillic acid | C8H8O4 | 168.149 | 167 | 123, 152, 108, 91 | 292, 260 | MS, NMR, DAD | ✓ | ✓ | |||
| 36 | Hydroxybenzoic acid | Syringic acid | C9H10O5 | 198.174 | 197 | 182, 153, 167, 138 | 276 | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 50 | Hydroxybenzoic acid | Ellagic acid | C14H6O8 | 302.194 | 301 | 284, 300, 257, 229, 184 | 367, 256, 301sh | MS | ✓ | ✓ | |||
| 14 | Hydroxycinnamic acid | 1-O-Sinapoyl-β-D-glucose | C17H22O10 | 386.353 | 385 | 223, 205, 341, 265, 190, 179, 119, 247 | 282 | MS | ✓ | ||||
| 16 | Hydroxycinnamic acid | 1-O-(4-Coumaroyl)-glucose | C15H18O8 | 326.301 | 325 | 163, 145, 119, 187, 265, 205 | 322 | MS | ✓ | ✓ | |||
| 20 | Hydroxycinnamic acid | 1-Caffeoyl-β-D-glucose | C15H18O9 | 342.300 | 341 | 179, 161, 143, 149, 131, 135 | 290, 304sh, 328 | MS | ✓ | ✓ | |||
| 21 | Hydroxycinnamic acid | Ferulic acid pentose | NA | NA | 325 | 149, 178, 193 | 326, 275 | MS | ✓ | ✓ | |||
| 22 | Hydroxycinnamic acid | Caftaric acid isomer | C13H11O9 | 311.224 | 311 | 179, 135, 149 | 325, 286 | MS | ✓ | ||||
| 24 | Hydroxycinnamic acid | Caftaric acid | C13H12O9 | 312.230 | 311 | 179, 135, 149, 267, 161, 237 | 326, 298sh, 243 | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 27 | Hydroxycinnamic acid | Coutaric acid | C13H12O8 | 296.231 | 295 | 163, 149, 119 | 316, 234, 300sh | MS, DAD | ✓ | ✓ | |||
| 28 | Hydroxycinnamic acid | Chlorogenic acid | C16H18O9 | 354.311 | 353 | 191, 179, 135, 161, 335, 172, 284 | 328, 244, 303sh | MS, DAD | ✓ | ||||
| 31 | Hydroxycinnamic acid | Chicoric acid | C22H18O12 | 474.374 | 473 | 311, 293, 179, 149, 135, 219 | 328, 305sh, 279 | MS | ✓ | ||||
| 32 | Hydroxycinnamic acid | Fertaric acid | C14H14O9 | 326.259 | 325 | 193, 175, 149, 281, 134 | 314, 279 | MS | ✓ | ||||
| 35 | Hydroxycinnamic acid | Caffeic acid | C9H8O4 | 180.159 | 179 | 135, 134, 96 | 324, 299sh, 240 | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 43 | Hydroxycinnamic acid | p-Coumaric acid | C9H8O3 | 164.160 | 163 | 119, 104, 93 | 310, 225, 211, 310sh | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 46 | Hydroxycinnamic acid | Ferulic acid | C10H10O4 | 194.186 | 193 | 134, 149, 178, 116 | 323, 289, 238sh | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 48 | Hydroxycinnamic acid | Sinapic acid | C11H12O5 | 224.212 | 223 | 164, 149, 208, 164, 193, 179 | 318, 238 | MS | ✓ | ✓ | |||
| 83 | Hydroxycinnamic acid | Cinnamic acid | C9H8O2 | 148.161 | 147 | 103, 77, 87, 129 | 276, 215, 203 | DAD | ✓ | ||||
| 3 | Flavan-3-ol | Gallocatechin | C15H14O7 | 306.270 | 305 | 179, 221, 219, 165, 261, 125, 137 | 274, 370 | MS | ✓ | ||||
| 6 | Flavan-3-ol | Procyanidin B1 | C30H26O12 | 578.526 | 577 | 425, 407, 289, 451, 287, 245, 451 | 275, 322 | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 10 | Flavan-3-ol | Procyanidin A1 | C30H24O12 | 576.501 | 575 | 449, 289, 423, 539, 477, 407, 441 | 280 | MS | ✓ | ✓ | ✓ | ||
| 15 | Flavan-3-ol | Epigallocatechin | C15H14O7 | 306.270 | 305 | 179, 165, 219, 221, 261, 125, 261, 125 | 274, 212, 235 | MS | ✓ | ||||
| 18 | Flavan-3-ol | Procyanidin C1 | C45H38O18 | 866.778 | 865 | 695, 407, 577, 287, 713, 739, 575, 425, 289 | 279 | MS, DAD | ✓ | ✓ | |||
| 19 | Flavan-3-ol | Procyanidin T2 | C45H38O18 | 866.778 | 865 | 577, 713, 289, 287, 425, 575, 695, 407, 739 | 280 | MS, DAD | ✓ | ✓ | |||
| 23 | Flavan-3-ol | Catechin | C15H14O6 | 290.271 | 289 | 245, 203, 179, 205, 227, 109, 123, 165, 125, 151 | 275, 222 | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 25 | Flavan-3-ol | Procyanidin B3 | C30H26O12 | 578.526 | 577 | 425, 407, 289, 151, 559, 445 | 270, 330 | MS, NMR, DAD | ✓ | ✓ | |||
| 26 | Flavan-3-ol | Procyanidin B4 | C30H26O12 | 578.526 | 577 | 425, 407, 289 | 280, 240 | MS, DAD | ✓ | ✓ | |||
| 29 | Flavan-3-ol | Procyanidin B2 | C30H26O12 | 578.526 | 577 | 425, 407, 289, 451, 287, 245, 125 | 280, 240, 370 | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 37 | Flavan-3-ol | Epigallocatechin gallate | C22H18O11 | 458.375 | 457 | 305, 219, 261, 221, 359, 169, 305, 289, 271, 125, 331 | 274, 238 | MS, DAD | ✓ | ||||
| 38 | Flavan-3-ol | Prodelphinidin A-type | C30H26O13 | 594.527 | 593 | 425, 441, 573, 423, 407, 289, 531, 273, 339, 245, 177, 161 | 276, 228, 320 | MS | ✓ | ✓ | |||
| 39 | Flavan-3-ol | Procyanidin dimer gallate | NA | NA | 729 | 577, 407, 559, 451, 711, 289, 593, 437, 425 | 280 | MS, DAD | ✓ | ✓ | |||
| 40 | Flavan-3-ol | Epicatechin | C15H14O6 | 290.271 | 289 | 245, 203, 109, 179, 205, 123, 125, 151 | 277, 226 | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 42 | Flavan-3-ol | Gallocatechin gallate | C22H18O11 | 458.375 | 457 | 169, 161, 359, 331, 169, 305, 193 | 276, 240 | MS | ✓ | ||||
| 47 | Flavan-3-ol | Epicatechin gallate | C22H18O10 | 442.376 | 441 | 289, 245, 205, 169, 125, 331, 271, 179 | 278, 240 | MS, NMR, DAD | ✓ | ✓ | |||
| 61 | Flavan-3-ol | Catechin gallate | C22H18O10 | 442.376 | 441 | 289, 245, 205, 331, 169, 125, 425, 271, 395, 169, 193, 405 | 278 | MS | ✓ | ✓ | |||
| 33 | Anthocyanin | Delphinidin-3-O-glucoside | C21H21O12+ | 465.387 | 463 | 301, 300, 271, 125 | 526, 361, 277, 402sh | DAD | ✓ | ||||
| 34 | Anthocyanin | Cyanidin-3-O-glucoside | C21H21O11+ | 449.388 | 447 | 284, 211, 285, 255, 147, 227 | 516, 262, 301sh | MS, DAD | ✓ | ||||
| 44 | Anthocyanin | Petunidin-3-O-glucoside | C22H23O12+ | 479.414 | 477 | 314, 315, 299, 300 | 526, 344sh, 277 | DAD | ✓ | ||||
| 45 | Anthocyanin | Peonidin-3-O-glucoside | C22H23O11+ | 463.415 | 461 | 299, 298, 284, 255, 227, 211 | 517, 280, 330sh, 421sh | MS, DAD | ✓ | ||||
| 52 | Anthocyanin | Malvidin-3-O-glucoside | C23H25O12+ | 493.441 | 491 | 329, 314, 299 | 528, 348sh, 288 | MS, DAD | ✓ | ✓ | |||
| 59 | Anthocyanin | Petunidin-3-(6-O-acetyl)glucoside | C24H25O13+ | 521.451 | 519 | 315, 302, 274, 149 | 528, 270, 350sh | DAD | ✓ | ||||
| 62 | Anthocyanin | Peonidin-3-(6-O-acetyl)glucoside | C24H25O12+ | 505.452 | 504 | 301, 286, 230, 258, 268 | 522, 280 | DAD | ✓ | ||||
| 67 | Anthocyanin | Malvidin-3-(6-O-acetyl)glucoside | C25H27O13+ | 535.478 | 533 | 329, 315 | 522, 344, 278 | DAD | ✓ | ||||
| 72 | Anthocyanin | Cyanidin-3-(6-O-coumaroyl)glucoside | C30H27O13+ | 595.533 | 593 | 287, 259, 231, 213, 259 | 524, 314, 284, 449sh | DAD | ✓ | ||||
| 80 | Anthocyanin | Petunidin-3-(6-O-coumaroyl)glucoside | C31H29O14+ | 625.553 | 624 | 317, 302, 274, 218, 228, 246 | 534, 282, 313 | DAD | ✓ | ||||
| 81 | Anthocyanin | Peonidin-3-(6-O-coumaroyl)glucoside | C31H29O13+ | 609.554 | 608 | 301, 286, 230, 258, 268 | 522, 312 | MS, DAD | ✓ | ||||
| 85 | Anthocyanin | Malvidin-3-(6-O-coumaroyl)glucoside | C32H31O14+ | 639.586 | 637 | 329, 299, 281 | 534, 318 | DAD | ✓ | ||||
| 86 | Anthocyanin | Malvidin-3-(6-O-caffeoyl)glucoside | C32H31O15+ | 655.581 | 655 | 331, 299, 637, 315, 475 | 532, 324, 284 | MS | ✓ | ||||
| 87 | Anthocyanin | Malvidin-3-O-rutinoside | C29H35O16+ | 639.583 | 637 | 331 | 526, 288 | MS | ✓ | ||||
| 41 | Flavanone | Taxifolin | C15H12O7 | 304.254 | 303 | 285, 125, 177, 275, 151, 259, 217 | 290, 326sh | MS | ✓ | ||||
| 55 | Flavanone | Taxifolin-O-pentoside | C20H20O11 | 436.371 | 435 | 303, 285, 399, 151, 241, 217, 175 | 274, 317 | MS | ✓ | ||||
| 56 | Flavanone | Taxifolin-3-O-glucoside | C21H22O12 | 466.395 | 465 | 285, 303, 151, 339, 177, 259, 447 | 290 | MS | ✓ | ||||
| 57 | Flavanone | Taxifolin-3-O-rhamnoside | C21H22O11 | 450.396 | 449 | 303, 285, 151, 323, 431 | 292, 235 | MS | ✓ | ||||
| 88 | Flavanone | Hesperetin | C16H14O6 | 302.282 | 301 | 258, 143, 157, 137, 286 | 284, 324sh, 221 | MS | ✓ | ||||
| 95 | Flavanone | Eriodictyol-7-O-glucoside | C21H22O11 | 450.396 | 449 | 287, 269, 151, 135, 259, 225, 209, 431 | 281, 327 | MS | ✓ | ||||
| 103 | Flavanone | Naringenin | C15H12O5 | 272.256 | 271 | 151, 177, 119, 165, 125, 107, 227, 191 | 289, 228, 336sh | MS | ✓ | ||||
| 104 | Flavanone | Naringenin-7-O-glucoside | C21H22O10 | 434.397 | 433 | 271, 269, 313, 177, 151, 119, 107 | 282, 222 | MS | ✓ | ||||
| 49 | Flavonol | Myricetin-3-O-galactoside | C21H20O13 | 480.378 | 479 | 317, 316, 178, 271 | 360, 265 | MS, DAD | ✓ | ||||
| 51 | Flavonol | Myricetin-3-O-glucuronide | C21H18O14 | 494.361 | 493 | 317 | 353, 300sh, 261 | MS, DAD | ✓ | ||||
| 53 | Flavonol | Myricetin-3-O-glucoside | C21H20O13 | 480.378 | 479 | 317, 316, 169, 271, 303, 227, 179, 151 | 362, 298sh, 260 | MS, DAD | ✓ | ||||
| 54 | Flavonol | Quercetin-3-O-rutinoside | C27H30O16 | 610.521 | 609 | 301, 300, 271, 255, 179, 343, 151 | 353, 256, 294sh | MS, NMR, DAD | ✓ | ✓ | |||
| 58 | Flavonol | Quercetin-3-O-galactoside | C21H20O12 | 464.379 | 463 | 301, 300, 179, 273, 257, 151 | 362, 256, 301sh | MS, NMR, DAD | ✓ | ✓ | ✓ | ||
| 60 | Flavonol | Quercetin-3-O-glucoside | C21H20O12 | 464.379 | 463 | 301, 300, 271, 161, 179, 255, 151 | 358, 256, 300sh | MS, NMR, DAD | ✓ | ✓ | |||
| 63 | Flavonol | Quercetin-3-O-glucuronide | C21H18O13 | 478.362 | 477 | 301, 179, 151, 283, 459, 431, 501 | 356, 254, 300sh | MS, NMR, DAD | ✓ | ✓ | |||
| 65 | Flavonol | Myricetin-3-O-rhamnoside | C21H20O12 | 464.379 | 463 | 317, 316, 271, 300, 179, 287, 151 | 372, 302sh, 248 | MS | ✓ | ||||
| 68 | Flavonol | Myricetin | C15H10O8 | 318.237 | 317 | 151, 179, 137, 287, 271, 109, 192 | 372, 253, 303sh, 207 | MS, NMR, DAD | ✓ | ||||
| 69 | Flavonol | Quercetin-3-O-rhamnoside | C21H20O11 | 448.380 | 447 | 301, 300, 179, 151, 271, 257 | 354, 258, 307sh | MS, NMR, DAD | ✓ | ✓ | |||
| 70 | Flavonol | Kaempferol-3-O-galactoside | C21H20O11 | 448.380 | 447 | 285, 255, 227, 327 | 361, 260 | MS | ✓ | ||||
| 71 | Flavonol | Kaempferol-3-O-rutinoside | C27H30O15 | 594.522 | 593 | 285, 257, 151, 447, 199, 241, 93 | 354, 274 | MS, DAD | ✓ | ✓ | |||
| 73 | Flavonol | Kaempferol-3-O-glucuronide | C21H18O12 | 462.363 | 461 | 285, 267, 443, 417, 257, 229 | 348, 265 | MS, DAD | ✓ | ||||
| 74 | Flavonol | Quercetin-3-(6-O-acetyl)glucoside | C23H22O13 | 506.416 | 505 | 301, 300, 463, 271, 255 | 354, 256, 267sh, 298sh | MS | ✓ | ||||
| 75 | Flavonol | Quercetin-3-(3-O-arabinosyl)glucoside | C26H28O16 | 596.493 | 595 | 301, 517, 151, 300, 463, 179, 445, 271, 255 | 354, 260, 231 | MS | ✓ | ||||
| 76 | Flavonol | Quercetin-3-(7-O-glucosyl)glucuronide | C27H28O18 | 640.503 | 639 | 477, 301, 179, 151 | 361, 300, 268, 256 | DAD | ✓ | ||||
| 77 | Flavonol | Quercetin-3-O-arabinose | C20H18O11 | 434.350 | 433 | 301, 179, 151, 300, 283 | 358, 311 | MS | ✓ | ✓ | |||
| 78 | Flavonol | C33H40O21 | 772.662 | 771 | 609, 301 | 355, 259, 299sh, 204 | NMR, DAD | ✓ | |||||
| 79 | Flavonol | Kaempferol-3-O-glucoside | C21H20O11 | 448.380 | 447 | 285, 255, 151, 227, 327, 243 | 348, 263, 297sh | MS, DAD | ✓ | ✓ | |||
| 82 | Flavonol | Quercetin | C15H10O7 | 302.239 | 301 | 151, 179, 273, 193, 257, 229 | 372, 255, 202sh, 300sh | MS, NMR, DAD | ✓ | ✓ | |||
| 84 | Flavonol | Kaempferol | C15H10O6 | 286.239 | 285 | 187, 117, 211, 127, 257, 151, 169, 241 | 369, 258, 390 | MS, NMR, DAD | ✓ | ✓ | |||
| 89 | Flavonol | Kaempferol-3-O-xyloside | C20H18O10 | 418.354 | 417 | 285, 255, 227 | 350 | MS | ✓ | ||||
| 90 | Flavonol | Kaempferol-3-O-rhamnoside | C21H20O10 | 432.381 | 431 | 285 | 351, 264, 202, 294sh | MS | ✓ | ||||
| 91 | Flavonol | Dihydrokaempferol-3-O-rhamnoside | C21H22O10 | 434.397 | 433 | 269, 287, 259, 180, 151, 368 | 286, 230 | MS | ✓ | ||||
| 92 | Flavonol | Isorhamnetin-3-O-galactoside | C22H22O12 | 478.406 | 477 | 315,.314 271, 300, 357 | 366, 289, 259 | MS | ✓ | ||||
| 93 | Flavonol | Isorhamnetin-3-O-glucoside | C22H22O12 | 478.406 | 477 | 315, 314, 285, 357, 271 | 354, 265sh | MS, DAD | ✓ | ||||
| 94 | Flavonol | Quercetin-3-(6-O-rhamnosyl)galactoside | C27H30O16 | 610.517 | 609 | 301, 541, 463, 300, 271, 255, 179, 447, 151 | 356, 256, 300 | NMR, DAD | ✓ | ||||
| 96 | Flavonol | Isorhamnetin-3-O-arabinose | C21H20O11 | 448.381 | 447 | 315, 314, 271, 243 | 345, 258 | MS | ✓ | ||||
| 97 | Flavonol | Isorhamnetin-3-O-glucuronide | C22H20O13 | 492.389 | 491 | 315, 255, 151 | 355, 265sh | MS | ✓ | ||||
| 98 | Flavonol | Isorhamnetin-3-O-rutinoside | C28H32O16 | 624.548 | 623 | 315, 300, 468 | 354, 256 | MS | ✓ | ||||
| 99 | Flavonol | Isorhamnetin-3-(6-O-feruloyl)glucoside | C32H30O15 | 654.577 | 653 | 315 | 320, 274 | MS | ✓ | ||||
| 100 | Flavonol | Isorhamnetin-3-(4-O-rhamnosyl)rutinoside | C34H42O20 | 770.685 | 769 | 461, 623, 163 | 354, 256 | MS, DAD | ✓ | ||||
| 101 | Flavonol | Kaempferol-3-(6-O-coumaroyl)glucoside | C30H26O13 | 594.525 | 593 | 285, 227, 255 | 317, 265, 356sh, 310sh | MS | ✓ | ||||
| 102 | Flavonol | Kaempferol-3 (7-O-glucosyl)galactoside | C27H30O16 | 610.521 | 609 | 447, 489, 285 | 343, 300sh, 265 | DAD | ✓ | ||||
| 105 | Flavonol | Diquercetin-3-(3-O-glucosyl)glucuronide | C42H36O24 | 924.722 | 923 | 765, 755, 837, 903, 935, 808 | 374 | MS | ✓ | ||||
| 64 | Flavone | Apigenin-7-O-glucoside | C21H20O10 | 432.381 | 431 | 269, 283, 311, 413, 231, 225, 201, 197, 149 | 335, 269, 253 | MS, NMR, DAD | ✓ | ||||
| 66 | Flavone | Luteolin-7-O-glucoside | C21H20O11 | 448.380 | 447 | 285, 226, 257, 217, 241, 198 | 349, 254sh, 205 | MS, NMR, DAD | ✓ | ||||
| 5 | Coumarin | Aesculin | C15H16O9 | 340.282 | 339 | 133, 177, 150 | 346, 289 | MS | ✓ | ||||
| 11 | Dihydrochalcone | Phlorizin | C21H24O10 | 436.413 | 435 | 273, 167, 229, 297 | 285, 230sh | MS | ✓ | ||||
| 12 | Coumarin | Fraxin | C16H18O10 | 370.310 | 369 | 207, 192, 354, 149, 123 | 332, 308sh | MS | ✓ | ||||
| 13 | Coumarin | Aesculetin | C9H6O4 | 178.143 | 177 | 149, 133, 105, 91, 115, 89, 65 | 334, 288sh | MS | ✓ | ||||
| 17 | Coumarin | Umbelliferone | C9H6O3 | 162.144 | 161 | 133, 117, 105, 51, 78 | 323, 236 | MS | ✓ | ||||
| ID 1 | Compound Group | Compound Name 1,2 | Chemical Formula 3 | MW (g/mol) | [M–H]– Precursor Ion | Main MS/MS Fragments (m/z) 4 | λmax (CH3OH) (nm) | Detection Mode 5 | Tissue Distribution 6 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LEAVES | STEMS | CANES | WOODS | ROOTS | |||||||||
| 106 | Monomers | trans-Astringin | C20H22O9 | 406.383 | 405 | 243, 225, 201, 322, 159, 199, 173 | 331, 305 | MS | ✓ | ✓ | ✓ | ||
| 107 | Monomers | trans-Resveratroloside | C20H22O8 | 390.388 | 389 | 227, 185, 179, 269, 143, 305, 371, 209 | 311 | MS | ✓ | ✓ | |||
| 108 | Monomers | trans-Resveratrol-2-C-glucoside | NA | NA | 389 | 269, 241, 299, 175, 163 | 326 | MS, NMR | ✓ | ✓ | |||
| 109 | Monomers | trans-Resveratrol-10-C-glucoside | NA | NA | 435 | 389, 227 | 315 | MS, NMR | ✓ | ||||
| 110 | Monomers | trans-Resveratrol-O-glucoside | C20H22O8 | 390.388 | 389 | 227, 305, 175, 185 | 321 | MS, NMR | ✓ | ✓ | ✓ | ||
| 183 | Monomers | cis-Resveratrol-O-glucoside | C20H23O8 | 391.391 | 389 | 227 | 319, 306 | MS | ✓ | ||||
| 113 | Monomers | trans-Piceid | C20H22O8 | 390.383 | 389 | 227, 185, 251, 269, 209, 371, 143 | 318, 306, 229 | MS, NMR, DAD | ✓ | ✓ | ✓ | ✓ | ✓ |
| 117 | Monomers | cis-Astringin | C20H22O9 | 406.383 | 405 | 243, 225, 201, 322, 405, 159, 228, 157 | 324, 260 | MS | ✓ | ||||
| 119 | Monomers | trans-Piceatannol | C14H12O4 | 244.246 | 243 | 175, 225, 149, 215, 201, 159, 181, 132, 199, 143 | 325, 290, 306 | MS, NMR | ✓ | ✓ | ✓ | ✓ | ✓ |
| 120 | Monomers | cis-Resveratroloside | C20H22O8 | 390.388 | 389 | 227, 371, 209 | 280 | MS | ✓ | ||||
| 121 | Monomers | cis-Piceid | C20H22O8 | 390.383 | 389 | 227, 269, 241, 185, 209, 371, 143 | 284, 230 | MS, NMR | ✓ | ✓ | ✓ | ||
| 122 | Monomers | trans-Isorhapontin | C21H24O9 | 420.411 | 419 | 257, 241, 225, 175, 242, 201, 159, 281, 132 | 326, 303, 290 | MS | ✓ | ||||
| 123 | Monomers | trans-Resveratrol | C14H12O3 | 228.247 | 227 | 185, 143, 183, 159, 157, 212, 205 | 306, 319, 228 | MS, NMR, DAD | ✓ | ✓ | ✓ | ✓ | ✓ |
| 124 | Monomers | 2,4,6-Trihydroxyphenanthrene-2-O-glucoside | C20H20O8 | 388.372 | 389 | 371, 353, 335, 227, 209, 199 | 261, 222 | MS | ✓ | ||||
| 126 | Monomers | trans-Isorhapontigenin | C15H14O4 | 258.270 | 257 | 242, 241, 224, 172, 213, 185 | 325, 303, 290 | MS | ✓ | ✓ | ✓ | ||
| 127 | Monomers | trans-Pinostilbene-4′-O-glucoside | C21H24O8 | 404.410 | 403 | 241, 226, 225 | NA | MS | ✓ | ||||
| 128 | Monomers | cis-Resveratrol | C14H12O3 | 228.247 | 227 | 185, 159, 143, 157, 212, 143 | 285, 232 | MS, NMR | ✓ | ✓ | |||
| 145 | Monomers | trans-Pterostilbene | C16H16O3 | 256.296 | 255 | 239, 197, 209, 226, 165 | 298, 305, 275 | MS, NMR, DAD | ✓ | ✓ | |||
| 146 | Monomers | cis-Pterostilbene | C16H16O3 | 256.296 | 255 | 197, 239, 209, 226, 165 | 279 | MS | ✓ | ||||
| 153 | Monomers | cis-Isorhapontigenin | C15H14O4 | 258.270 | 257 | 241, 213, 185, 224 | 318, 220 | MS | ✓ | ||||
| 154 | Monomers | trans-Rhaponticin | C21H24O9 | 420.414 | 419 | 257, 241, 281, 299, 323, 405, 389, 243, 169, 395 | 324, 220 | MS | ✓ | ||||
| 155 | Monomers | trans-Pinostilbene | C15H14O3 | 242.270 | 241 | 181, 225, 197, 169 | NA | MS | ✓ | ||||
| 156 | Monomers | cis-Pinostilbene | C15H14O3 | 242.270 | 241 | 181, 225, 197, 169 | NA | MS | ✓ | ||||
| 111 | Dimers | Leachianol G | C28H24O7 | 472.496 | 471 | 387, 377, 349, 255, 121 | 280, 218 | MS, NMR | ✓ | ✓ | ✓ | ||
| 112 | Dimers | Leachianol F | C28H24O7 | 472.496 | 471 | 349, 453, 255, 287, 153, 241, 121 | 280, 218 | MS, NMR | ✓ | ✓ | ✓ | ||
| 114 | Dimers | Restrytisol A | C28H24O7 | 472.486 | 471 | 377, 255, 349, 121, 471 | 280, 221 | MS | ✓ | ✓ | ✓ | ||
| 115 | Dimers | Ampelopsin A | C28H22O7 | 470.479 | 469 | 345, 451, 375, 363, 257, 357, 423, 317, 241 | 283 | MS, NMR | ✓ | ✓ | ✓ | ✓ | |
| 116 | Dimers | Pallidol | C28H22O6 | 454.478 | 453 | 359, 265, 435, 406, 391, 346, 273 | 284 | MS, NMR | ✓ | ✓ | ✓ | ✓ | ✓ |
| 118 | Dimers | Caraphenol B | C28H22O7 | 470.473 | 469 | 451, 281, 363, 375, 227, 423 | 326, 291 | MS | ✓ | ✓ | |||
| 130 | Dimers | Ampelopsin D | C28H22O6 | 454.478 | 453 | 359, 361, 437, 215, 343, 199, 255, 289 | 314, 280 | MS, NMR | ✓ | ✓ | ✓ | ✓ | |
| 131 | Dimers | Quadrangularin A | C28H22O6 | 454.471 | 453 | 359, 289, 411, 435, 347, 253 | 314 | MS, NMR | ✓ | ✓ | |||
| 132 | Dimers | (+)-cis-ε-Viniferin | C28H22O6 | 454.471 | 453 | 435, 347, 411, 333, 359, 369, 253 | 286, 201, 230 | MS, NMR | ✓ | ✓ | |||
| 134 | Dimers | (+)-trans-ε-Viniferin | C28H22O6 | 454.471 | 453 | 359, 435, 347, 369, 411, 333, 253, 225 | 327, 285, 308 | MS, NMR, DAD | ✓ | ✓ | ✓ | ✓ | ✓ |
| 135 | Dimers | Viniferifuran | C28H20O6 | 452.455 | 451 | NA | 317, 289, 209 | MS, NMR | ✓ | ||||
| 136 | Dimers | Diptoindonesin A | C34H32O11 | 616.610 | 615 | 453, 359, 411, 347, 585 | 326, 226 | MS, NMR | ✓ | ✓ | |||
| 141 | Dimers | trans-ω-Viniferin | C28H22O6 | 454.478 | 453 | 435, 359, 347, 411, 395, 333, 285 | 324, 280 | MS, NMR | ✓ | ✓ | ✓ | ✓ | |
| 144 | Dimers | cis-ω-Viniferin | C28H22O6 | 454.478 | 453 | 435, 411, 395, 333, 285, 359, 225 | 294 | MS. NMR | ✓ | ||||
| 149 | Dimers | trans-δ-Viniferin | C28H22O6 | 454.478 | 453 | 435, 369, 411, 347, 333, 359, 225, 409 | 312, 225 | MS, DAD | ✓ | ✓ | ✓ | ✓ | |
| 152 | Dimers | cis-δ-Viniferin | C28H22O6 | 454.478 | 453 | 435, 411, 369, 359, 333, 347, 317, 307, 251, 267 | 285, 232 | MS | ✓ | ||||
| 157 | Dimers | trans-ε-Viniferin derivative (dimethylated) | C30H26O6 | 482.523 | 481 | 387, 375, 226, 197, 466 | 325 | MS | ✓ | ||||
| 158 | Dimers | trans-δ-Viniferin derivative (dimethylated) | C30H26O6 | 482.523 | 481 | 397, 361, 439, 387, 463 | 313 | MS | ✓ | ||||
| 159 | Dimers | trans-Scirpusin A | C28H22O7 | 470.470 | 469 | 375, 451, 385, 359, 241, 427, 728, 445, 287, 514, 955 | 320, 286, 204 | MS, NMR | ✓ | ||||
| 162 | Dimers | Maackin A | C28H22O8 | 486.470 | 485 | 244, 226, 137 | 327, 288, 204 | MS, NMR | ✓ | ||||
| 164 | Dimers | trans-ε-Viniferin derivative (γ-lactam ring) | C32H26O7N | 936.550 | 536 | NA | NA | MS, NMR | ✓ | ||||
| 165 | Dimers | trans-Resveratrol derivative (γ-lactam ring) | C18H16O4N | 310.324 | 310 | NA | NA | MS, NMR | ✓ | ||||
| 171 | Dimers | Malibatol A | C28H20O7 | 468.454 | 467 | NA | NA | MS, NMR | ✓ | ||||
| 172 | Dimers | Ampelopsin F | C28H22O6 | 454.471 | 453 | NA | 282, 220 | MS, NMR | ✓ | ||||
| 176 | Dimers | Viniferal | C35H26O8 | 574.579 | 573 | NA | NA | MS, NMR | ✓ | ||||
| 177 | Dimers | Vitisinol C | C27H24O5 | 428.482 | 427 | NA | 358, 279 | MS | ✓ | ||||
| 178 | Dimers | Vitisinol E | C27H24O6 | 444.475 | 444 | NA | 281, 230, 204 | MS, NMR | ✓ | ||||
| 179 | Dimers | Vitisinol B | C35H26O8 | 574.579 | 573 | NA | 282, 228, 204 | MS, NMR | ✓ | ||||
| 181 | Dimers | Viniferether A | C29H26O7 | 486.509 | 485 | NA | 280, 229 | MS, NMR | ✓ | ||||
| 182 | Dimers | Viniferether B | C29H26O7 | 486.513 | 485 | NA | 280, 231 | MS, NMR | ✓ | ||||
| 125 | Trimers | Ampelopsin B | C28H22O6 | 454.478 | 453 | 359, 243, 211, 183, 265 | 281, 328 | MS, NMR | ✓ | ✓ | ✓ | ||
| 139 | Trimers | trans-Miyabenol C | C42H32O9 | 680.698 | 679 | 661, 573, 479, 451, 637, 585, 447 | 322, 279 | MS, NMR | ✓ | ✓ | ✓ | ✓ | ✓ |
| 140 | Trimers | cis-Miyabenol C | C42H32O9 | 680.699 | 679 | 661, 573, 479, 451, 637, 585, 447 | 285 | MS, NMR | ✓ | ✓ | |||
| 142 | Trimers | Davidiol A | C42H32O9 | 680.704 | 679 | 585, 447, 491, 385, 479, 465, 567 | 284, 219 | MS | ✓ | ✓ | |||
| 143 | Trimers | α-Viniferin | C42H30O9 | 678.682 | 677 | 571, 583, 437, 449, 463, 501, 331 | 284, 309 | MS, NMR | ✓ | ✓ | ✓ | ||
| 161 | Trimers | Ampelopsin C | C42H32O9 | 680.709 | 679 | NA | 283 | MS, NMR | ✓ | ✓ | |||
| 169 | Trimers | Viniferol D | C42H32O9 | 680.702 | 679 | NA | NA | MS, NMR | ✓ | ✓ | |||
| 173 | Trimers | Ampelopsin E | C42H32O9 | 680.701 | 679 | NA | 325, 285 | MS, NMR | ✓ | ✓ | |||
| 129 | Tetramers | Hopeaphenol | C56H42O12 | 906.925 | 905 | 811, 717, 451, 611, 359, 299 | 283, 226 | MS, NMR | ✓ | ✓ | ✓ | ✓ | |
| 133 | Tetramers | Isohopeaphenol | C56H42O12 | 906.925 | 905 | 451, 675, 811, 717, 358, 265 | 284 | MS, NMR | ✓ | ✓ | ✓ | ✓ | ✓ |
| 137 | Tetramers | Ampelopsin H | C56H42O12 | 906.925 | 905 | 811, 717, 705, 793 | 281 | MS, NMR | ✓ | ✓ | ✓ | ||
| 138 | Tetramers | Vaticanol C-like isomer | C56H42O12 | 906.929 | 905 | 811, 717, 793, 705, 611 | 281 | MS, NMR | ✓ | ||||
| 147 | Tetramers | Vitisin A (r2-viniferin) | C56H42O12 | 906.920 | 905 | 811, 887, 717, 693, 545, 451, 359, 265 | 328, 285 | MS, NMR | ✓ | ✓ | ✓ | ✓ | |
| 148 | Tetramers | Vitisifuran A | C56H40O12 | 904.907 | 903 | NA | 322, 232 | MS, NMR | ✓ | ||||
| 150 | Tetramers | Vitisin B (r-viniferin) | C56H42O12 | 906.920 | 905 | 799, 887, 811, 717, 545, 451, 359, 317 | 321, 286 | MS, NMR | ✓ | ✓ | ✓ | ✓ | |
| 151 | Tetramers | Vitisifuran B | C56H40O12 | 904.907 | 903 | NA | 324, 228 | MS, NMR | ✓ | ||||
| 160 | Tetramers | Vitisin C | C56H42O12 | 906.926 | 905 | NA | NA | MS, NMR | ✓ | ||||
| 166 | Tetramers | Viniferol A | C56H42O12 | 906.925 | 905 | 559, 813, 361, 453, 651, 541, 801, 783 | 284, 227 | MS, NMR | ✓ | ||||
| 167 | Tetramers | Viniferol B | C56H42O12 | 906.929 | 905 | 559, 813, 361, 453, 651, 541, 801, 783 | 283, 225 | MS, NMR | ✓ | ||||
| 168 | Tetramers | Viniferol C | C56H42O12 | 906.929 | 905 | NA | 284, 228 | MS, NMR | ✓ | ||||
| 170 | Tetramers | Viniferol E | C56H44O13 | 924.940 | 923 | NA | 284, 231 | MS, NMR | ✓ | ||||
| 174 | Tetramers | Wilsonol C | C56H42O12 | 906.929 | 905 | NA | 231 | NMR | ✓ | ||||
| 175 | Tetramers | Heyneanol A | C56H42O12 | 906.929 | 905 | 320, 284 | 322, 237 | NMR | ✓ | ||||
| 180 | Tetramers | Stenophyllol C | C56H42O12 | 906.923 | 905 | NA | 285, 330, 223 | MS, NMR | ✓ | ||||
| 163 | Hexamers | Viniphenol A | C84H64O18 | 1361.391 | 1360 | NA | NA | MS, NMR | ✓ | ||||
| Id | Compound Name 1 | Minimum Value 2,3 | Maximum Value | Mean Value 4 | Standard Deviation | N5 | References |
|---|---|---|---|---|---|---|---|
| 63 | Quercetin-3-O-glucuronide | 868.63 | 46,528.55 | 10,305.10 | 13,363.51 | 10 | [1,2,4,5,19,59,60,66,75,79,81,82,84,85,86] |
| 58 | Quercetin-3-O-galactoside | 21.72 | 28,831.11 | 7436.94 | 9880.90 | 14 | [2,3,5,20,22,73,75,79,81,85] |
| 60 | Quercetin-3-O-glucoside | 27.65 | 22,610.13 | 7256.42 | 7628.71 | 24 | [1,2,3,5,19,20,22,59,60,66,73,75,79,80,81,82,84,85,86] |
| 24 | Caftaric acid | 12.46 | 14,052.62 | 4151.97 | 3984.79 | 18 | [3,4,5,19,20,22,59,60,66,73,75,79,81,82,84,85,86] |
| 69 | Quercetin-3-O-rhamnoside | 1210.53 | 4206.67 | 2708.60 | 1498.07 | 2 | [1,2] |
| 79 | Kaempferol-3-O-glucoside | 2.56 | 6203.85 | 1730.09 | 1812.15 | 20 | [2,3,19,20,22,59,60,66,73,75,79,80,81,84,85,86] |
| 73 | Kaempferol-3-O-glucuronide | 47.92 | 1698.41 | 662.34 | 736.83 | 3 | [2,19,66,79,81,82,85,86] |
| 27 | Coutaric acid | 4.54 | 1491.02 | 635.81 | 432.70 | 10 | [3,19,20,22,73,75,79,81,82,85] |
| 53 | Myricetin-3-O-glucoside | ND | 850.12 | 291.23 | 254.65 | 8 | [2,3,20,22,73,85] |
| 54 | Quercetin-3-O-rutinoside | 1.30 | 1650.01 | 257.51 | 473.26 | 12 | [2,4,5,32,73,75,79,81,82,85,87] |
| 71 | Kaempferol-3-O-rutinoside | 0.12 | 730.01 | 200.34 | 307.13 | 4 | [2,19,75,79,81,85,86] |
| 32 | Fertaric acid | 85.48 | 85.48 | 85.48 | 0.00 | 1 | [81,82] |
| 7 | p-Hydroxybenzoic acid | 15.80 | 151.00 | 83.40 | 67.60 | 2 | [19,32] |
| 42 | Gallocatechin gallate | 20.10 | 78.70 | 49.40 | 29.30 | 2 | [32] |
| 68 | Myricetin | 1.00 | 193.28 | 44.75 | 74.37 | 5 | [1,3,22,81,88] |
| 3 | Gallocatechin | 4.84 | 102.00 | 40.20 | 43.85 | 3 | [32,81,82] |
| 5 | Aesculin | 1.60 | 50.70 | 25.63 | 20.06 | 3 | [32,81] |
| 15 | Epigallocatechin | 1.67 | 66.30 | 23.77 | 30.08 | 3 | [32,81,82] |
| 37 | Epigallocatechin gallate | 0.04 | 43.81 | 10.69 | 16.71 | 6 | [32,73,81,82] |
| 23 | Catechin | 0.02 | 76.58 | 10.62 | 21.44 | 18 | [19,32,59,60,73,75,79,81,82,84,87,88] |
| 46 | Ferulic acid | 0.008 | 89.80 | 9.07 | 25.61 | 11 | [32,59,60,73,79,81,84] |
| 82 | Quercetin | 0.13 | 52.17 | 8.84 | 16.55 | 16 | [1,4,19,32,59,60,73,84,87,88] |
| 6 | Procyanidin B1 | 0.39 | 25.56 | 6.80 | 10.83 | 4 | [73,75,79,81,82] |
| 4 | Protocatechuic acid | 1.25 | 10.50 | 5.88 | 4.63 | 2 | [32] |
| 8 | Gentisic acid | 0.59 | 8.85 | 4.72 | 4.13 | 2 | [32] |
| 11 | Phlorizin | 2.95 | 2.95 | 2.95 | 0.00 | 1 | [81,82] |
| 2 | Gallic acid | 0.01 | 7.80 | 2.77 | 2.95 | 9 | [32,79,81,82,87,88] |
| 78 | Quercetin-3-(3-O-rhamnosyl)glucoside-7-O-rhamnoside | 1.32 | 4.21 | 2.77 | 1.44 | 2 | [1] |
| 40 | Epicatechin | 0.01 | 15.02 | 2.46 | 4.69 | 18 | [32,59,60,73,75,79,81,82,84,87,88] |
| 94 | Quercetin-3-(6-O-rhamnosyl)galactoside | 0.02 | 4.02 | 2.02 | 2.00 | 2 | [1] |
| 29 | Procyanidin B2 | 0.35 | 5.69 | 1.91 | 2.21 | 4 | [75,79,81,82] |
| 47 | Epicatechin gallate | 0.01 | 8.45 | 1.74 | 2.57 | 9 | [59,60,81,82,84,88] |
| 28 | Chlorogenic acid | 0.01 | 11.50 | 1.74 | 3.70 | 8 | [32,73,79,88] |
| 35 | Caffeic acid | 0.003 | 19.60 | 1.68 | 4.84 | 15 | [1,32,59,73,79,81,82,84,87,88] |
| 25 | Procyanidin B3 | 0.74 | 2.41 | 1.57 | 0.84 | 2 | [75,81,82] |
| 26 | Procyanidin B4 | 0.61 | 2.38 | 1.49 | 0.89 | 2 | [75,81,86] |
| 93 | Isorhamnetin-3-O-glucoside | 1.48 | 1.48 | 1.48 | 0.00 | 1 | [2,80,81,85] |
| 84 | Kaempferol | 0.01 | 6.77 | 1.28 | 2.19 | 8 | [1,32,59,60,84,86,88] |
| 34 | Cyanidin-3-O-glucoside | 0.01 | 6.40 | 1.17 | 2.04 | 8 | [4,5,59,60,77,78,79,84,88] |
| 98 | Isorhamnetin-3-O-rutinoside | 1.12 | 1.12 | 1.12 | 0.00 | 1 | [2,81] |
| 43 | p-Coumaric acid | 0.01 | 8.17 | 0.92 | 2.42 | 10 | [32,59,60,79,84,88] |
| 64 | Apigenin-7-O-glucoside | 0.09 | 1.60 | 0.85 | 0.75 | 2 | [1] |
| 10 | Procyanidin A1 | 0.72 | 0.72 | 0.72 | 0.00 | 1 | [75] |
| 66 | Luteolin-7-O-glucoside | 0.02 | 1.91 | 0.60 | 0.69 | 5 | [1,4,73,81,82] |
| 48 | Sinapic acid | 0.55 | 0.55 | 0.55 | 0.00 | 1 | [81] |
| 50 | Ellagic acid | 0.06 | 0.77 | 0.41 | 0.36 | 2 | [32] |
| 41 | Taxifolin | 0.37 | 0.37 | 0.37 | 0.00 | 1 | [81] |
| 83 | Cinnamic acid | 0.17 | 0.51 | 0.34 | 0.17 | 2 | [79] |
| 75 | Quercetin-3-(3-O-arabinosyl)glucoside | 0.31 | 0.31 | 0.31 | 0.00 | 1 | [2,81] |
| 89 | Kaempferol-3-O-xyloside | 0.23 | 0.23 | 0.23 | 0.00 | 1 | [2,75] |
| 45 | Peonidin-3-O-glucoside | 0.01 | 0.60 | 0.20 | 0.24 | 4 | [5,77,78,79,88] |
| 30 | Vanillic acid | 0.01 | 0.54 | 0.19 | 0.19 | 5 | [77,78,79,81,88] |
| 14 | 1-O-Sinapoyl-β-D-glucose | 0.15 | 0.15 | 0.15 | 0.00 | 1 | [75] |
| 70 | Kaempferol-3-O-galactoside | 0.06 | 0.06 | 0.06 | 0.00 | 1 | [2,75,85,86] |
| 36 | Syringic acid | 0.01 | 0.07 | 0.04 | 0.03 | 2 | [88] |
| 52 | Malvidin-3-O-glucoside | 0.01 | 0.06 | 0.04 | 0.03 | 2 | [1,2,3,4,5,19,32,59,66,75,77,78,79,80,81,82,84,85,86,88] |
| 16 | 1-O-(4-Coumaroyl)-glucose | 0.03 | 0.03 | 0.03 | 0.00 | 1 | [75] |
| 1 | Quinic acid | NQ | NQ | NQ | NQ | 0 | [4,19] |
| 9 | γ-Resorcylic acid | NQ | NQ | NQ | NQ | 0 | [82] |
| 12 | Fraxin | NQ | NQ | NQ | NQ | 0 | [82] |
| 13 | Aesculetin | NQ | NQ | NQ | NQ | 0 | [19] |
| 17 | Umbelliferone | NQ | NQ | NQ | NQ | 0 | [19] |
| 20 | 1-Caffeoyl-β-D-glucose | NQ | NQ | NQ | NQ | 0 | [81] |
| 21 | Ferulic acid pentose | NQ | NQ | NQ | NQ | 0 | [19] |
| 22 | Caftaric acid isomer | NQ | NQ | NQ | NQ | 0 | [85] |
| 33 | Delphinidin-3-O-glucoside | NQ | NQ | NQ | NQ | 0 | [77,78,79] |
| 44 | Petunidin-3-O-glucoside | NQ | NQ | NQ | NQ | 0 | [77,78,79] |
| 49 | Myricetin-3-O-galactoside | NQ | NQ | NQ | NQ | 0 | [2,79,85] |
| 51 | Myricetin-3-O-glucuronide | NQ | NQ | NQ | NQ | 0 | [2,79] |
| 59 | Petunidin-3-(6-O-acetyl)glucoside | NQ | NQ | NQ | NQ | 0 | [77] |
| 61 | Catechin gallate | NQ | NQ | NQ | NQ | 0 | [86] |
| 62 | Peonidin-3-(6-O-acetyl)glucoside | NQ | NQ | NQ | NQ | 0 | [79] |
| 65 | Myricetin-3-O-rhamnoside | NQ | NQ | NQ | NQ | 0 | [2] |
| 67 | Malvidin-3-(6-O-acetyl)glucoside | NQ | NQ | NQ | NQ | 0 | [77,79] |
| 72 | Cyanidin-3-(6-O-coumaroyl)glucoside | NQ | NQ | NQ | NQ | 0 | [77] |
| 74 | Quercetin-3-(6-O-acetyl)glucoside | NQ | NQ | NQ | NQ | 0 | [19] |
| 76 | Quercetin-3-(7-O-glucosyl)glucuronide | NQ | NQ | NQ | NQ | 0 | [80] |
| 80 | Petunidin-3-(6-O-coumaroyl)glucoside | NQ | NQ | NQ | NQ | 0 | [77] |
| 81 | Peonidin-3-(6-O-coumaroyl)glucoside | NQ | NQ | NQ | NQ | 0 | [2,77,79] |
| 85 | Malvidin-3-(6-O-coumaroyl)glucoside | NQ | NQ | NQ | NQ | 0 | [77,79] |
| 88 | Hesperetin | NQ | NQ | NQ | NQ | 0 | [4] |
| 90 | Kaempferol-3-O-rhamnoside | NQ | NQ | NQ | NQ | 0 | [2] |
| 92 | Isorhamnetin-3-O-galactoside | NQ | NQ | NQ | NQ | 0 | [2,85] |
| 95 | Eriodictyol-7-O-glucoside | NQ | NQ | NQ | NQ | 0 | [19] |
| 96 | Isorhamnetin-3-O-arabinose | NQ | NQ | NQ | NQ | 0 | [2] |
| 97 | Isorhamnetin-3-O-glucuronide | NQ | NQ | NQ | NQ | 0 | [2] |
| 100 | Isorhamnetin-3(4-O-rhamnosyl)rutinoside | NQ | NQ | NQ | NQ | 0 | [2,80,85] |
| 101 | Kaempferol-3-(6-O-coumaroyl)glucoside | NQ | NQ | NQ | NQ | 0 | [4] |
| 102 | Kaempferol-3 (7-O-glucosyl)galactoside | NQ | NQ | NQ | NQ | 0 | [80] |
| 103 | Naringenin | NQ | NQ | NQ | NQ | 0 | [82] |
| 104 | Naringenin-7-O-glucoside | NQ | NQ | NQ | NQ | 0 | [82] |
| 105 | Diquercetin-3-(3-O-glucosyl)glucuronide | NQ | NQ | NQ | NQ | 0 | [66] |
| Id | Compound Name | Minimum Value 1,2 | Maximum Value | Mean Value 3 | Standard Deviation | N4 | References |
|---|---|---|---|---|---|---|---|
| 123 | trans-Resveratrol | ND | 1886.80 | 137.88 | 444.15 | 24 | [6,10,11,19,23,32,59,60,61,62,63,64,68,70,73,81,82,84,87,88,89] |
| 138 | Vaticanol C-like isomer | ND | 226.80 | 96.85 | 102.35 | 6 | [61,81,82,89] |
| 121 | cis-Piceid | ND | 368.40 | 78.38 | 132.57 | 6 | [10,62,63,64,68,81,82] |
| 119 | trans-Piceatannol | ND | 232.10 | 78.04 | 108.94 | 5 | [23,63,82,88] |
| 137 | Ampelopsin H | ND | 226.80 | 76.08 | 106.58 | 6 | [61,81,82,89] |
| 143 | α-Viniferin | ND | 189.06 | 71.61 | 75.19 | 6 | [10,61,81,89] |
| 140 | cis-Miyabenol C | ND | 148.60 | 50.67 | 69.29 | 6 | [61,81,82,89] |
| 110 | cis-Resveratrol-3-O-glucoside | ND | 232.63 | 47.41 | 83.30 | 6 | [59,60,84] |
| 113 | trans-Piceid | ND | 170.23 | 44.71 | 64.69 | 17 | [6,10,11,23,61,62,63,64,68,70,73,81,82,89] |
| 139 | trans-Miyabenol C | ND | 121.30 | 41.57 | 56.43 | 6 | [61,81,82,89] |
| 149 | trans-δ-Viniferin | 1.09 | 165.71 | 35.55 | 53.31 | 8 | [6,10,11,62,68,70,82] |
| 120 | cis-Resveratroloside | 15.20 | 37.50 | 26.35 | 11.15 | 2 | [63] |
| 134 | (+)-trans-ε-Viniferin | ND | 98.20 | 25.11 | 35.79 | 15 | [6,10,11,23,61,62,68,70,73,81,82,89] |
| 130 | Ampelopsin D | ND | 67.60 | 22.78 | 31.70 | 6 | [10,61,81,82,89] |
| 141 | trans-ω-Viniferin | ND | 63.55 | 21.35 | 29.84 | 6 | [10,61,81,82,89] |
| 144 | cis-ω-Viniferin | ND | 63.55 | 21.24 | 29.92 | 6 | [10,61,81,82,89] |
| 128 | cis-Resveratrol | ND | 53.10 | 19.46 | 20.50 | 4 | [62,63,64,73,82] |
| 107 | trans-Resveratroloside | 7.50 | 21.80 | 14.65 | 7.15 | 2 | [63] |
| 116 | Pallidol | ND | 26.71 | 11.52 | 12.09 | 6 | [61,81,82,89] |
| 131 | Quadrangularin A | ND | 33.80 | 11.29 | 15.92 | 6 | [10,61,81,82,89] |
| 133 | Isohopeaphenol | ND | 131.17 | 7.12 | 12.33 | 6 | [61,81,82,89] |
| 153 | cis-Isorhapontigenin | 0.10 | 13.00 | 6.55 | 6.45 | 2 | [63] |
| 122 | trans-Isorhapontin | 0.07 | 21.30 | 6.44 | 8.69 | 4 | [63,81,82] |
| 126 | trans-Isorhapontigenin | 0.10 | 9.60 | 4.85 | 4.75 | 2 | [63] |
| 145 | trans-Pterostilbene | ND | 10.83 | 3.92 | 4.24 | 10 | [6,10,61,62,64,68,70,82,89] |
| 132 | (+)-cis-ε-Viniferin | ND | 7.31 | 1.83 | 3.17 | 4 | [10,62,68,82,89] |
| 106 | trans-Astringin | 0.04 | 7.60 | 3.02 | 3.09 | 4 | [63,81,82] |
| 152 | cis-δ-Viniferin | ND | 3.42 | 1.71 | 1.71 | 2 | [62,68] |
| 127 | trans-Pinostilbene-4′-O-glucoside | 0.10 | 3.30 | 1.70 | 1.60 | 2 | [63] |
| 117 | cis-Astringin | 0.20 | 2.10 | 1.15 | 0.95 | 2 | [63] |
| 155 | trans-Pinostilbene | 0.10 | 2.00 | 1.05 | 0.95 | 2 | [63] |
| 154 | trans-Rhaponticin | 0.10 | 1.80 | 0.95 | 0.85 | 2 | [63] |
| 156 | cis-Pinostilbene | 0.10 | 0.30 | 0.20 | 0.10 | 2 | [63] |
| 114 | Restrytisol A | NQ | NQ | NQ | NQ | 0 | [10] |
| 124 | 2,4,6-Trihydroxyphenanthrene-2-O-glucoside | NQ | NQ | NQ | NQ | 0 | [64] |
| 125 | Ampelopsin B | NQ | NQ | NQ | NQ | 0 | [10] |
| 142 | Davidiol A | NQ | NQ | NQ | NQ | 0 | [10] |
| 146 | cis-Pterostilbene | NQ | NQ | NQ | NQ | 0 | [10] |
| 157 | trans-ε-Viniferin derivative (dimethylated) | NQ | NQ | NQ | NQ | 0 | [10] |
| 158 | trans-δ-Viniferin derivative (dimethylated) | NQ | NQ | NQ | NQ | 0 | [10] |
| Id | Compound Name 1 | Minimum Value 2 | Maximum Value | Mean Value 3 | Standard Deviation | N4 | References |
|---|---|---|---|---|---|---|---|
| 58 | Quercetin-3-O-galactoside | 1920.34 | 41,831.70 | 17,403.61 | 15,457.56 | 4 | [21,50] |
| 23 | Catechin | 283.72 | 98,290.95 | 14,900.45 | 27,191.10 | 12 | [9,21,37,50,53,55,69,74,90] |
| 2 | Gallic acid | 386.54 | 32,960.41 | 10,307.36 | 13,374.20 | 4 | [9,21,34,37,55] |
| 40 | Epicatechin | 193.61 | 33,154.03 | 9251.64 | 12,435.12 | 14 | [9,21,33,50,53,54,55,69,90] |
| 6 | Procyanidin B1 | 215.36 | 50,709.00 | 9216.18 | 14,385.61 | 10 | [9,33,37,50,53,54,55,69] |
| 19 | Procyanidin T2 | 1388.90 | 35,015.04 | 9100.99 | 8406.54 | 2 | [9,34,50,55,69] |
| 25 | Procyanidin B3 | 186.04 | 23,108.65 | 8724.23 | 5791.29 | 4 | [9,21,55,69] |
| 47 | Epicatechin gallate | 2371.55 | 9862.08 | 6362.96 | 2950.30 | 6 | [9,21,33,54,55] |
| 18 | Procyanidin C1 | 305.51 | 9710.00 | 5007.76 | 4702.25 | 2 | [9,50,55,69] |
| 82 | Quercetin | 321.88 | 8210.20 | 4266.04 | 3944.16 | 2 | [21] |
| 24 | Caftaric acid | 110.35 | 16,110.62 | 3373.18 | 5723.49 | 6 | [33,54,74,87] |
| 26 | Procyanidin B4 | 131.00 | 4355.20 | 2243.10 | 2112.10 | 2 | [69] |
| 39 | Procyanidin dimer gallate | 110.04 | 4358.12 | 2234.08 | 2124.04 | 2 | [9,33,34,50,54,69] |
| 29 | Procyanidin B2 | 10.49 | 6670.76 | 2056.93 | 2735.52 | 4 | [9,21,50,53,55,69,74] |
| 60 | Quercetin-3-O-glucoside | 29.88 | 7270.12 | 1785.38 | 2544.06 | 6 | [9,21,37,53,55,90] |
| 69 | Quercetin-3-O-rhamnoside | 320.20 | 2820.00 | 1570.10 | 1249.90 | 2 | [21] |
| 10 | Procyanidin A1 | 674.91 | 1833.85 | 1254.38 | 579.47 | 4 | [33,50,54] |
| 84 | Kaempferol | 70.12 | 1830.57 | 950.35 | 880.23 | 2 | [21] |
| 63 | Quercetin-3-O-glucuronide | 391.52 | 1424.35 | 908.82 | 469.54 | 6 | [33,34,37,54,74] |
| 87 | Malvidin-3-O-rutinoside | 451.00 | 628.77 | 539.89 | 88.88 | 4 | [33,54] |
| 52 | Malvidin-3-O-glucoside | 224.88 | 801.37 | 513.13 | 288.25 | 4 | [33,54] |
| 43 | p-Coumaric acid | 12.00 | 934.08 | 473.04 | 461.04 | 2 | [9,21,55] |
| 35 | Caffeic acid | 10.18 | 647.32 | 328.75 | 318.57 | 2 | [9,21,55] |
| 38 | Prodelphinidin A-type | 27.46 | 292.88 | 160.17 | 132.71 | 2 | [33,50,54] |
| 99 | Isorhamnetin-3-(6-O-feruloyl)glucoside | 81.10 | 115.07 | 98.09 | 16.99 | 4 | [33,54] |
| 86 | Malvidin-3-(6-O-caffeoyl)glucoside | 47.33 | 119.20 | 83.27 | 35.94 | 4 | [33,54] |
| 71 | Kaempferol-3-O-rutinoside | 21.99 | 127.39 | 74.69 | 52.70 | 4 | [33,54] |
| 54 | Quercetin-3-O-rutinoside | 10.55 | 126.73 | 65.84 | 46.33 | 6 | [21,33,34,54,74,90] |
| 79 | Kaempferol-3-O-glucoside | 20.14 | 79.08 | 49.61 | 29.47 | 4 | [33,54] |
| 36 | Syringic acid | 6.48 | 32.23 | 19.36 | 12.88 | 2 | [21] |
| 46 | Ferulic acid | 8.01 | 25.55 | 16.78 | 8.77 | 2 | [9,21,55] |
| 16 | 1-O-(4-Coumaroyl)-glucose | NQ | NQ | NQ | NQ | 0 | [53] |
| 20 | 1-Caffeoyl-β-D-glucose | NQ | NQ | NQ | NQ | 0 | [53] |
| 21 | Ferulic acid pentose | NQ | NQ | NQ | NQ | 0 | [9,55] |
| 31 | Chicoric acid | NQ | NQ | NQ | NQ | 0 | [9,55] |
| 55 | Taxifolin-O-pentoside | NQ | NQ | NQ | NQ | 0 | [9,55] |
| 56 | Taxifolin-3-O-glucoside | NQ | NQ | NQ | NQ | 0 | [53] |
| 57 | Taxifolin-3-O-rhamnoside | NQ | NQ | NQ | NQ | 0 | [9,34,55,74] |
| 61 | Catechin gallate | NQ | NQ | NQ | NQ | 0 | [53] |
| 77 | Quercetin-3-O-arabinose | NQ | NQ | NQ | NQ | 0 | [50] |
| 91 | Dihydrokaempferol-3-O-rhamnoside | NQ | NQ | NQ | NQ | 0 | [9,55] |
| Id | Compound Name | Minimum Value 1,2 | Maximum Value | Mean Value 3 | Standard Deviation | N4 | References |
|---|---|---|---|---|---|---|---|
| 123 | trans-Resveratrol | ND | 2130.00 | 506.41 | 570.04 | 6 | [9,23,26,27,53,55,58,91] |
| 134 | (+)-trans-ε-Viniferin | 14.30 | 1400.67 | 433.49 | 765.23 | 6 | [9,23,26,27,28,53,55,56,58,67,91] |
| 130 | Ampelopsin D | ND | 130.00 | 65.00 | 65.00 | 2 | [53] |
| 150 | Vitisin B (r-Viniferin) | 6.80 | 61.10 | 33.95 | 27.15 | 2 | [26,27,28,53,58,67,91] |
| 113 | trans-Piceid | 14.52 | 14.52 | 14.52 | 0.00 | 1 | [9,23,26,27,53,55] |
| 126 | trans-Isorhapontigenin | ND | 19.80 | 9.90 | 9.90 | 2 | [91] |
| 119 | trans-Piceatannol | ND | 21.10 | 7.42 | 9.68 | 3 | [9,23,26,27,53,55,58,91] |
| 149 | trans-δ-Viniferin | 4.86 | 4.86 | 4.86 | 0.00 | 1 | [23] |
| 106 | trans-Astringin | NQ | NQ | NQ | NQ | 0 | [9,53,55] |
| 107 | trans-Resveratroloside | NQ | NQ | NQ | NQ | 0 | [53] |
| 108 | trans-Resveratrol-2-C-glucoside | NQ | NQ | NQ | NQ | 0 | [53] |
| 109 | trans-Resveratrol-10-C-glucoside | NQ | NQ | NQ | NQ | 0 | [56] |
| 110 | trans-Resveratrol-O-glucoside | NQ | NQ | NQ | NQ | 0 | [56] |
| 111 | Leachianol G | NQ | NQ | NQ | NQ | 0 | [26,27] |
| 112 | Leachianol F | NQ | NQ | NQ | NQ | 0 | [26,27] |
| 114 | Restrytisol A | NQ | NQ | NQ | NQ | 0 | [58] |
| 115 | Ampelopsin A | NQ | NQ | NQ | NQ | 0 | [26,27,28,53,56,58] |
| 116 | Pallidol | NQ | NQ | NQ | NQ | 0 | [9,26,27,55] |
| 118 | Caraphenol B | NQ | NQ | NQ | NQ | 0 | [53] |
| 129 | Hopeaphenol | NQ | NQ | NQ | NQ | 0 | [9,26,53,55,56,58] |
| 131 | Quadrangularin A | NQ | NQ | NQ | NQ | 0 | [26,27,53] |
| 133 | Isohopeaphenol | NQ | NQ | NQ | NQ | 0 | [26,27,53,56,67] |
| 135 | Viniferifuran | NQ | NQ | NQ | NQ | 0 | [67] |
| 136 | Diptoindonesin A | NQ | NQ | NQ | NQ | 0 | [26,27,53] |
| 137 | Ampelopsin H | NQ | NQ | NQ | NQ | 0 | [26,27] |
| 139 | trans-Miyabenol C | NQ | NQ | NQ | NQ | 0 | [26,27,53,58] |
| 140 | cis-Miyabenol C | NQ | NQ | NQ | NQ | 0 | [53] |
| 142 | Davidiol A | NQ | NQ | NQ | NQ | 0 | [26,27,53] |
| 143 | α-Viniferin | NQ | NQ | NQ | NQ | 0 | [9,55] |
| 145 | trans-Pterostilbene | NQ | NQ | NQ | NQ | 0 | [9,26,27,55] |
| 147 | Vitisin A (r2-Viniferin) | NQ | NQ | NQ | NQ | 0 | [28,56,58,67] |
| 148 | Vitisifuran A | NQ | NQ | NQ | NQ | 0 | [67] |
| 151 | Vitisifuran B | NQ | NQ | NQ | NQ | 0 | [67] |
| 159 | trans-Scirpusin A | NQ | NQ | NQ | NQ | 0 | [26,27,53] |
| 160 | Vitisin C | NQ | NQ | NQ | NQ | 0 | [26,27,67] |
| 161 | Ampelopsin C | NQ | NQ | NQ | NQ | 0 | [26,27] |
| 162 | Maackin A | NQ | NQ | NQ | NQ | 0 | [26,27] |
| 163 | Viniphenol A | NQ | NQ | NQ | NQ | 0 | [26,27] |
| 166 | Viniferol A | NQ | NQ | NQ | NQ | 0 | [56] |
| 167 | Viniferol B | NQ | NQ | NQ | NQ | 0 | [56] |
| 168 | Viniferol C | NQ | NQ | NQ | NQ | 0 | [56] |
| 169 | Viniferol D | NQ | NQ | NQ | NQ | 0 | [67] |
| 171 | Malibatol A | NQ | NQ | NQ | NQ | 0 | [56] |
| 172 | Ampelopsin F | NQ | NQ | NQ | NQ | 0 | [56] |
| 173 | Ampelopsin E | NQ | NQ | NQ | NQ | 0 | [56] |
| 176 | Viniferal | NQ | NQ | NQ | NQ | 0 | [67] |
| 178 | Vitisinol E | NQ | NQ | NQ | NQ | 0 | [28] |
| Id | Compound Name | Minimum Value 1,2 | Maximum Value | Mean Value 3 | Standard Deviation | N4 | References |
|---|---|---|---|---|---|---|---|
| 123 | trans-Resveratrol | ND | 6526.29 | 2797.17 | 2559.72 | 17 | [7,12,24,25,40,48,49,50,51,52,58,72] |
| 134 | (+)-trans-ε-Viniferin | 21.00 | 12,612.22 | 2449.25 | 3197.26 | 13 | [7,12,24,25,40,50,51,52,58] |
| 133 | Isohopeaphenol | ND | 3521.52 | 851.43 | 1133.46 | 7 | [12,24,51,52] |
| 150 | Vitisin B (r-Viniferin) | 0.01 | 2159.00 | 668.08 | 818.46 | 10 | [7,12,24,40,50,51,52,58] |
| 119 | trans-Piceatannol | 0.50 | 1710.24 | 583.88 | 799.71 | 11 | [7,12,24,25,40,50,51,52,58] |
| 141 | trans-ω-Viniferin | ND | 1714.63 | 556.41 | 628.60 | 3 | [7,12,24,25,50] |
| 129 | Hopeaphenol | ND | 1439.21 | 511.39 | 585.40 | 9 | [12,24,25,40,50,51,52,58] |
| 116 | Pallidol | 4.00 | 1276.43 | 440.81 | 591.08 | 3 | [12,24,40,50] |
| 139 | trans-Miyabenol C | 0.01 | 2108.47 | 412.53 | 702.07 | 7 | [7,12,24,25,40,51,52,58] |
| 115 | Ampelopsin A | 0.01 | 1684.16 | 370.88 | 534.32 | 8 | [12,24,25,40,50,51,52,58] |
| 147 | Vitisin A (r2-Viniferin) | 43.00 | 717.55 | 293.18 | 301.67 | 3 | [12,24,25,50,51,58] |
| 149 | trans-δ-Viniferin | 9.00 | 43.00 | 26.00 | 17.00 | 2 | [24,50] |
| 113 | trans-Piceid | 0.50 | 36.21 | 13.50 | 16.11 | 5 | [7,40,48,49,50] |
| 177 | Vitisinol C | 1.00 | 29.00 | 15.00 | 14.00 | 2 | [24] |
| 108 | trans-Resveratrol-2-C-glucoside | NQ | NQ | NQ | NQ | 0 | [7] |
| 110 | trans-Resveratrol-O-glucoside | NQ | NQ | NQ | NQ | 0 | [72] |
| 111 | Leachianol G | NQ | NQ | NQ | NQ | 0 | [50] |
| 112 | Leachianol F | NQ | NQ | NQ | NQ | 0 | [50] |
| 114 | Restrytisol A | NQ | NQ | NQ | NQ | 0 | [50,58] |
| 118 | Caraphenol B | NQ | NQ | NQ | NQ | 0 | [72] |
| 121 | cis-Piceid | NQ | NQ | NQ | NQ | 0 | [50] |
| 125 | Ampelopsin B | NQ | NQ | NQ | NQ | 0 | [40] |
| 126 | trans-Isorhapontigenin | NQ | NQ | NQ | NQ | 0 | [50] |
| 130 | Ampelopsin D | NQ | NQ | NQ | NQ | 0 | [50] |
| 164 | trans-ε-Viniferin derivative (γ-lactam ring) | NQ | NQ | NQ | NQ | 0 | [25] |
| 165 | trans-Resveratrol derivative (γ-lactam ring) | NQ | NQ | NQ | NQ | 0 | [25] |
| Id | Compound Name | Minimum Value 1 | Maximum Value | Mean Value 2 | Standard Deviation | N3 | References |
|---|---|---|---|---|---|---|---|
| 23 | Catechin | 65.16 | 6735.24 | 1747.01 | 2525.88 | 10 | [46,48,49,50,83] |
| 48 | Sinapic acid | 26.41 | 2283.20 | 1154.81 | 1128.40 | 4 | [46,47,83] |
| 6 | Procyanidin B1 | 215.36 | 806.87 | 511.12 | 295.76 | 2 | [50] |
| 40 | Epicatechin | 45.53 | 896.17 | 269.40 | 289.87 | 6 | [48,49,50] |
| 46 | Ferulic acid | 0.92 | 650.13 | 165.63 | 279.74 | 8 | [46,47,48,49,83] |
| 2 | Gallic acid | 7.21 | 570.13 | 165.06 | 234.37 | 8 | [46,47,48,49,83] |
| 38 | Prodelphinidin A-type | 27.46 | 292.88 | 160.17 | 132.71 | 2 | [50] |
| 4 | Protocatechuic acid | 3.25 | 379.85 | 103.31 | 159.84 | 8 | [46,47,48,49,83] |
| 24 | Caftaric acid | 18.64 | 77.60 | 48.12 | 29.48 | 4 | [48,49] |
| 7 | p-Hydroxybenzoic acid | 0.01 | 95.22 | 47.62 | 47.61 | 4 | [46,47,83] |
| 30 | Vanillic acid | 0.01 | 152.10 | 40.13 | 64.74 | 8 | [46,47,48,49,83] |
| 36 | Syringic acid | 0.01 | 113.09 | 31.05 | 47.43 | 8 | [46,47,48,49,83] |
| 50 | Ellagic acid | 0.01 | 53.25 | 18.78 | 20.99 | 8 | [46,48,49,83] |
| 27 | Coutaric acid | 5.20 | 19.39 | 12.30 | 7.10 | 4 | [48,49] |
| 43 | p-Coumaric acid | 0.01 | 31.20 | 11.13 | 11.97 | 8 | [46,47,48,49,83] |
| 35 | Caffeic acid | 1.15 | 3.43 | 2.29 | 1.14 | 4 | [47,48,49] |
| 10 | Procyanidin A1 | NQ | NQ | NQ | NQ | 0 | [50] |
| 18 | Procyanidin C1 | NQ | NQ | NQ | NQ | 0 | [50] |
| 19 | Procyanidin T2 | NQ | NQ | NQ | NQ | 0 | [50] |
| 29 | Procyanidin B2 | NQ | NQ | NQ | NQ | 0 | [50] |
| 39 | Procyanidin dimer gallate | NQ | NQ | NQ | NQ | 0 | [50] |
| 58 | Quercetin-3-O-galactoside | NQ | NQ | NQ | NQ | 0 | [50] |
| 77 | Quercetin-3-O-arabinose | NQ | NQ | NQ | NQ | 0 | [50] |
| Id | Compound Name | Minimum Value 1,2 | Maximum Value | Mean Value 3 | Standard Deviation | N4 | References |
|---|---|---|---|---|---|---|---|
| 134 | (+)-trans-ε-Viniferin | 122.57 | 14,080.88 | 8263.87 | 4866.89 | 7 | [12,13,43,45] |
| 132 | (+)-cis-ε-Viniferin | 2504.08 | 14,023.65 | 3851.97 | 5759.79 | 2 | [13] |
| 133 | Isohopeaphenol | 11.68 | 7913.00 | 2429.69 | 2544.59 | 7 | [12,13,43,45] |
| 143 | α-Viniferin | ND | 4732.06 | 2366.03 | 2366.03 | 2 | [13] |
| 123 | trans-Resveratrol | 15.11 | 3604.04 | 2195.12 | 1414.54 | 7 | [12,13,43,45] |
| 137 | Ampelopsin H | 1144.77 | 2518.08 | 1831.43 | 686.66 | 2 | [13] |
| 116 | Pallidol | 410.60 | 2602.15 | 1647.36 | 916.64 | 3 | [12,13] |
| 111 | Leachianol G | 350.32 | 2800.34 | 1575.33 | 1225.01 | 2 | [13] |
| 129 | Hopeaphenol | 20.09 | 5006.77 | 1570.13 | 1683.09 | 7 | [12,13,43,45] |
| 141 | trans-ω-Viniferin | 1554.16 | 1554.16 | 1554.16 | 0.00 | 1 | [12] |
| 112 | Leachianol F | 35.57 | 2805.13 | 1420.35 | 1384.78 | 2 | [13] |
| 115 | Ampelopsin A | 151.00 | 3684.01 | 1345.17 | 1541.97 | 5 | [12,13,45] |
| 147 | Vitisin A (r2-Viniferin) | 1298.67 | 1298.67 | 1298.67 | 0.00 | 1 | [12] |
| 128 | cis-Resveratrol | 780.58 | 3609.66 | 774.64 | 1192.86 | 2 | [13] |
| 110 | trans-Resveratrol-O-glucoside | 131.00 | 1090.00 | 610.50 | 479.50 | 2 | [44] |
| 150 | Vitisin B (r-Viniferin) | 569.18 | 569.18 | 569.18 | 0.00 | 1 | [12] |
| 139 | trans-Miyabenol C | ND | 1339.51 | 430.02 | 482.17 | 5 | [12,13,45] |
| 125 | Ampelopsin B | ND | 493.44 | 246.72 | 246.72 | 2 | [13] |
| 119 | trans-Piceatannol | 38.00 | 378.07 | 160.69 | 154.14 | 3 | [12,45] |
| 130 | Ampelopsin D | 10.51 | 310.22 | 160.37 | 149.86 | 2 | [43] |
| 113 | trans-Piceid | 35.00 | 50.00 | 42.50 | 7.50 | 2 | [45] |
| 136 | Diptoindonesin A | 9.78 | 57.70 | 33.74 | 23.96 | 2 | [44] |
| 106 | trans-Astringin | 2.56 | 24.00 | 13.28 | 10.72 | 2 | [44] |
| Id | Compound Name | Minimum Value 1 | Maximum Value | Mean Value 2 | Standard Deviation | N3 | References |
|---|---|---|---|---|---|---|---|
| 134 | (+)-trans-ε-Viniferin | 125.10 | 18,000.98 | 6475.95 | 8163.57 | 3 | [12,23,30,43] |
| 150 | Vitisin B (r-Viniferin) | 11.10 | 12,829.85 | 6420.48 | 6409.38 | 2 | [12,30,41,42] |
| 129 | Hopeaphenol | 1814.90 | 1814.90 | 1814.90 | 0.00 | 1 | [12,41,42] |
| 115 | Ampelopsin A | 15.60 | 2178.23 | 1096.92 | 1081.32 | 2 | [12,29,30,41,42] |
| 147 | Vitisin A (r2-viniferin) | 87.10 | 2093.13 | 1090.12 | 1003.02 | 2 | [12,30] |
| 133 | Isohopeaphenol | 529.54 | 529.54 | 529.54 | 0.00 | 1 | [12] |
| 123 | trans-Resveratrol | 46.30 | 1095.24 | 503.25 | 438.74 | 3 | [12,23,29,30] |
| 141 | trans-ω-Viniferin | 127.70 | 127.70 | 127.70 | 0.00 | 1 | [12] |
| 139 | trans-Miyabenol C | 12.70 | 212.34 | 112.52 | 99.82 | 2 | [12,30] |
| 113 | trans-Piceid | 112.07 | 112.07 | 112.07 | 0.00 | 1 | [23,29] |
| 116 | Pallidol | 73.06 | 73.06 | 73.06 | 0.00 | 1 | [12,29] |
| 119 | trans-Piceatannol | 4.20 | 121.33 | 47.18 | 52.66 | 3 | [12,23,30] |
| 149 | trans-δ-viniferin | 32.77 | 32.77 | 32.77 | 0.00 | 1 | [23] |
| 121 | cis-Piceid | NQ | NQ | NQ | NQ | 0 | [29] |
| 161 | Ampelopsin C | NQ | NQ | NQ | NQ | 0 | [41,42] |
| 169 | Viniferol D | NQ | NQ | NQ | NQ | 0 | [41,42] |
| 170 | Viniferol E | NQ | NQ | NQ | NQ | 0 | [41,42] |
| 173 | Ampelopsin E | NQ | NQ | NQ | NQ | 0 | [41,42] |
| 174 | Wilsonol C | NQ | NQ | NQ | NQ | 0 | [29] |
| 175 | Heyneanol A | NQ | NQ | NQ | NQ | 0 | [29] |
| 179 | Vitisinol B | NQ | NQ | NQ | NQ | 0 | [41,42] |
| 180 | Stenophyllol C | NQ | NQ | NQ | NQ | 0 | [41,42] |
| 181 | Viniferether A | NQ | NQ | NQ | NQ | 0 | [41,42] |
| 182 | Viniferether B | NQ | NQ | NQ | NQ | 0 | [41,42] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goufo, P.; Singh, R.K.; Cortez, I. A Reference List of Phenolic Compounds (Including Stilbenes) in Grapevine (Vitis vinifera L.) Roots, Woods, Canes, Stems, and Leaves. Antioxidants 2020, 9, 398. https://doi.org/10.3390/antiox9050398
Goufo P, Singh RK, Cortez I. A Reference List of Phenolic Compounds (Including Stilbenes) in Grapevine (Vitis vinifera L.) Roots, Woods, Canes, Stems, and Leaves. Antioxidants. 2020; 9(5):398. https://doi.org/10.3390/antiox9050398
Chicago/Turabian StyleGoufo, Piebiep, Rupesh Kumar Singh, and Isabel Cortez. 2020. "A Reference List of Phenolic Compounds (Including Stilbenes) in Grapevine (Vitis vinifera L.) Roots, Woods, Canes, Stems, and Leaves" Antioxidants 9, no. 5: 398. https://doi.org/10.3390/antiox9050398