The Oxidative Stress Markers in the Erythrocytes and Heart Muscle of Obese Rats: Relate to a High-Fat Diet but Not to DJOS Bariatric Surgery

 , , , ,
, , , ,
Abstract
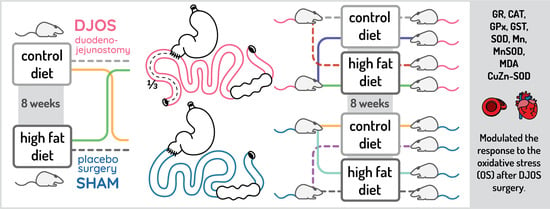
:
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Experimental Design
2.3. Bariatric Procedures
2.4. Blood Collection
2.5. Tissue Collection
2.6. Oxidative Stress Markers’ Analysis
2.6.1. Glutathione Reductase (GR) Activity (EC 1.8.1.7)
2.6.2. Catalase (CAT) Activity (EC 1.11.1.6)
2.6.3. Glutathione Peroxidase (GPx) Activity (EC 1.11.1.9)
2.6.4. Glutathione-S Transferase (GST) Activity (EC 2.5.1.18)
2.6.5. Superoxide Dismutase (SOD) Activity (EC 1.15.1.1)
2.6.6. Malondialdehyde (MDA) Concentration
2.6.7. Protein Concentration
2.7. Statistical Analysis
3. Results
3.1. Erythrocytes
3.1.1. Erythrocytes Glutathione Reductase (GR) Activity
3.1.2. Erythrocytes Catalase (CAT) Activity
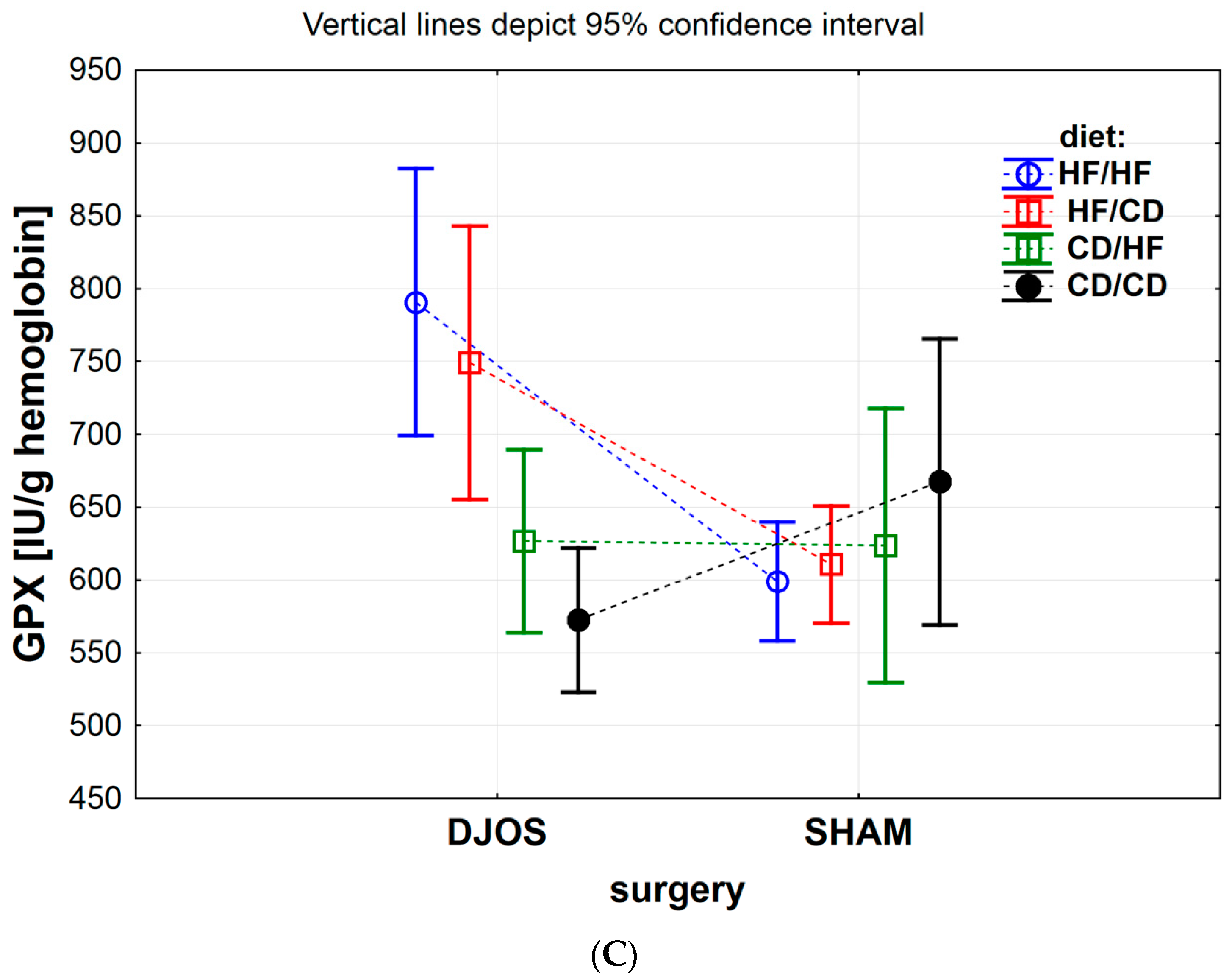
3.1.3. Erythrocytes Glutathione Peroxidase (GPx) Activity
3.1.4. Erythrocytes Glutathione-S-Transferase (GST) Activity
3.1.5. Erythrocytes Total Superoxide Dismutase (SOD) Activity
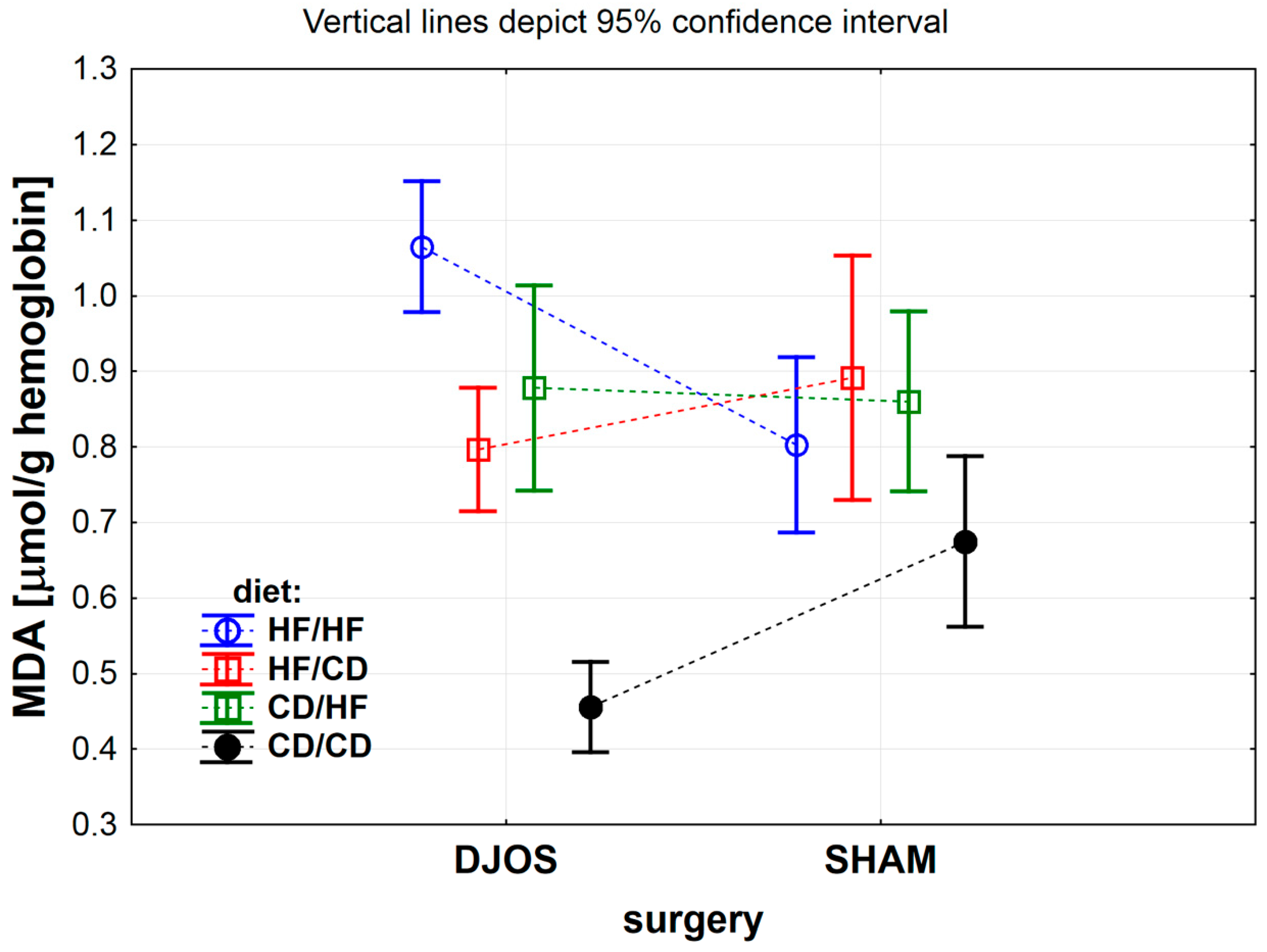
3.1.6. Erythrocytes Malondialdehyde (MDA) Concentration
3.2. Heart Muscle
3.2.1. Heart Muscle Glutathione Reductase (GR) Activity
3.2.2. Heart Muscle Catalase (CAT) Activity
3.2.3. Heart Muscle Glutathione Peroxidase (GPx) Activity
3.2.4. Heart Muscle Glutathione-S-Transferase (GST) Activity
3.2.5. Heart Muscle Total Superoxide Dismutase (SOD) Activity
3.2.6. Heart muscle Mn-Dependent Superoxide Dismutase (MnSOD) Activity
3.2.7. Heart Muscle Copper–Zinc Superoxide Dismutase (CuZnSOD) Activity
3.2.8. Heart Muscle Malondialdehyde (MDA) Concentration
4. Discussion
4.1. Erythrocytes
4.2. Heart Muscle
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Montezano, A.C.; Touyz, R.M. Reactive oxygen species and endothelial function-role of nitric oxide synthase uncoupling and Nox family nicotinamide adenine dinucleotide phosphate oxidases. Basic Clin. Pharmacol. Toxicol. 2012, 110, 87–94. [Google Scholar] [CrossRef]
- Rolo, A.P.; Teodoro, J.S.; Palmeira, C.M. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radic. Biol. Med. 2012, 52, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, A.; Priyadarsini, I.K. Free radicals, oxidative stress and importance of antioxidants in human health. J. Med. Allied Sci. 2011, 1, 53–60. [Google Scholar]
- Keaney, J.F., Jr.; Larson, M.G.; Vasan, R.S.; Wilson, P.W.; Lipinska, I.; Corey, D.; Massaro, J.M.; Sutherland, P.; Vita, J.A.; Benjamin, E.J. Obesity and systemic oxidative stress: Clinical correlates of oxidative stress in the Framingham Study. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 434–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olusi, S.O. Obesity is an independent risk factor for plasma lipid peroxidation and depletion of erythrocyte cytoprotectic enzymes in humans. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 1159–1164. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Eckl, P.; Ortner, A. Possible mutagens derived from lipids and lipid precursors. Mutat. Res. 1990, 238, 223–233. [Google Scholar] [CrossRef]
- Bloomer, R.J.; Kabir, M.M.; Marshall, K.E.; Canale, R.E.; Farney, T.M. Postprandial oxidative stress in response to dextrose and lipid meals of differing size. Lipids Health Dis. 2010, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Manna, P.; Jain, S.K. Obesity, oxidative stress, adipose tissue dysfunction, and the associated health risks: Causes and therapeutic strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [Green Version]
- Ben Saad, H.; Nasri, I.; Elwej, A.; Krayem, N.; Jarraya, R.; Kallel, C.; Zeghal, N.; Amara, I.B. A mineral and antioxidant-rich extract from the red marine algae Alsidium corallinum exhibits cytoprotective effects against potassium bromate-induced erythrocyte oxidative damages in mice. Biol. Trace Elem. Res. 2014, 160, 85–96. [Google Scholar] [CrossRef]
- Cay, M.; Naziroğlu, M. Effects of intraperitoneally-administered vitamin E and seleniumon the blood biochemical and haematological parameters in rats. Cell Biochem. Funct. 1999, 17, 143–148. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species in non-phagocytic cells. J. Leukoc. Biol. 1999, 65, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Ventura-Clapier, R.; Garnier, A.; Veksler, V. Energy metabolism in heart failure. J. Physiol. 2004, 555, 1–13. [Google Scholar] [CrossRef]
- Melanson, K.J.; McInnis, K.J.; Rippe, J.M.; Blackburn, G.; Wilson, P.F. Obesity and cardiovascular disease risk: Research update. Cardiol. Rev. 2001, 9, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Sjöström, L.; Narbro, K.; Sjöström, C.D.; Karason, K.; Larsson, B.; Wedel, H.; Lystig, T.; Sullivan, M.; Bouchard, C.; Carlsson, B.; et al. Effects of bariatric surgery on mortality in Swedish obese subjects. N. Engl. J. Med. 2007, 357, 741–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjöström, L.; Peltonen, M.; Jacobson, P.; Sjöström, C.D.; Karason, K.; Wedel, H.; Ahlin, S.; Anveden, Å.; Bengtsson, C.; Bergmark, G.; et al. Bariatric surgery and long-term cardiovascular events. JAMA 2012, 307, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauck, M.A. Unraveling the science of incretin biology. Am. J. Med. 2009, 122, S3–S10. [Google Scholar] [CrossRef]
- Rubino, F.; Gagner, F. Potential of surgery for curing type 2 diabetes mellitus. Ann. Surg. 2002, 236, 554–559. [Google Scholar] [CrossRef]
- Grueneberger, J.M.; Karcz-Socha, I.; Marjanovic, G.; Kuesters, S.; Zwirska-Korczala, K.; Schmidt, K.; Karcz, W.K. Pylorus preserving loop duodeno-enterostomy with sleeve gastrectomy—Preliminary results. BMC Surg. 2014, 14, 20. [Google Scholar] [CrossRef] [Green Version]
- Stygar, D.; Chełmecka, E.; Sawczyn, T.; Skrzep-Poloczek, B.; Poloczek, J.; Karcz, K.W. Changes of plasma FABP4, CRP, leptin, and chemerin levels in relation to different dietary patterns and duodenal-jejunal omega switch surgery in Sprague–Dawley rats. Oxid. Med. Cell. Longev. 2018, 2018, 2151429. [Google Scholar] [CrossRef] [Green Version]
- Stygar, D.; Sawczyn, T.; Skrzep-Poloczek, B.; Karcz-Socha, I.; Doleżych, B.; Zawisza-Raszka, A.; Augustyniak, M.; Żwirska-Korczala, K.; Karcz, W.K. Ileal transposition in rats influenced glucose metabolism and HSP70 levels. Open Life Sci. 2015, 10, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Stygar, D.; Sawczyn, T.; Skrzep-Poloczek, B.; Poloczek, J.; Klosok, M.; Karcz-Socha, I.; Fink, J.M.; Skonieczna, M.; Dolezych, B.; Zwirska-Korczala, K.; et al. Metabolic surgery in Zucker rats influenced miRNA, caveolin-1 expression and lipid metabolism. Open Life Sci. 2015, 10, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Stygar, D.; Andrare, D.; Bażanów, B.; Chełmecka, E.; Sawczyn, T.; Skrzep-Poloczek, B.; Olszańska, E.; Karcz, K.W.; Jochem, J. The impact of DJOS surgery, a high fat diet and a control diet on the enzymes of glucose metabolism in the liver and muscles of Sprague-Dawley Rats. Front. Physiol. 2019, 10, 571. [Google Scholar] [CrossRef] [PubMed]
- Stygar, D.; Skrzep-Poloczek, B.; Romuk, E.; Chełmecka, E.; Poloczek, J.; Sawczyn, T.; Maciarz, J.; Kukla, M.; Karcz, K.W.; Jochem, J.; et al. The influence of high-fat, high-sugar diet and bariatric surgery on HSP70 and HSP90 plasma and liver concentrations in diet-induced obese rats. Cell Stress Chaperones 2019, 24, 427–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auberval, N.; Dal, S.; Bietiger, W.; Pinget, M.; Jeandidier, N.; Maillard-Pedracini, E.; Schini-Kerth, V.; Sigrist, S. Metabolic and oxidative stress markers in Wistar rats after 2 months on a high-fat diet. Diabetol. Metab. Syndr. 2014, 6, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinho, R.A.; Sepa-Kishi, D.M.; Bikopoulos, G.; Wu, M.V.; Uthayakumar, A.; Mohasses, A.; Hughes, M.C.; Perry, C.G.R.; Ceddia, R.B. High-fat diet induces skeletal muscle oxidative stress in a fiber type-dependent manner in rats. Free Radic. Biol. Med. 2017, 110, 381–389. [Google Scholar] [CrossRef]
- Wietzycoski, C.R.; Marchesini, J.C.D.; Al-Themyat, S.; Meyer, F.S.; Trindade, M.R. Improvement in oxidative stress after duodenojejunostomy in an experimental model of type 2 diabetes mellitus. Arq. Bras. Cir. Dig. 2016, 29 (Suppl. S1), 3–7. [Google Scholar] [CrossRef] [Green Version]
- Skrzep-Poloczek, B.; Stygar, D.; Chelmecka, E.; Nabrdalik, K.; Romuk, E.; Poloczek, J.; Sawczyn, T.; Karcz, K.W.; Gumprecht, J. Antioxidant status in the soleus muscle of Sprague-Dawley rats in relation to duodenal-jejunal omega switch and different dietary patterns. Oxid. Med. Cell. Longev. 2018, 2018, 3795070. [Google Scholar] [CrossRef] [Green Version]
- Skrzep-Poloczek, B.; Stygar, D.; Sawczyn, T.; Romuk, E.; Chełmecka, E.; Poloczek, J.; Dulska, A.; Kazura, W.; Karcz, K.W. Impact of ileal transposition surgical intervention on antioxidant status measured in liver tissue of obese Zucker Rats (Crl:ZUC-Leprfa). Oxid. Med. Cell. Longev. 2018, 2018, 7342451. [Google Scholar] [CrossRef] [Green Version]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Methuen Co., Ltd.: London, UK, 1959. [Google Scholar]
- Stygar, D.; Sawczyn, T.; Skrzep-Poloczek, B.; Owczarek, A.J.; Matysiak, N.; Michalski, M.; Mielańczyk, Ł.; Bażanów, B.; Ziora, P.; Choręza, P.; et al. The effects of duodenojejunal omega switch in combination with high-fat diet and control diet on incretins, body weight, and glucose tolerance in Sprague-Dawley rats. Obes. Surg. 2018, 28, 748–759. [Google Scholar] [CrossRef] [Green Version]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Mannervik, B. Glutathione peroxidase. Methods Enzymol. 1985, 113, 490–495. [Google Scholar] [PubMed]
- Habig, W.H.; Jakoby, W.B. Assays for differentiation of glutathione S-transferases. Methods Enzymol. 1981, 77, 398–405. [Google Scholar] [PubMed]
- Oyanagui, Y. Reevaluation of assay methods and establishment of kit for superoxide dismutase activity. Anal. Biochem. 1984, 142, 290–296. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Balasubramaniam, P.; Malathi, A. Comparative study of hemoglobin estimated by Drabkin’s and Sahli’s methods. J. Postgrad. Med. 1992, 38, 8–9. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Ordonez, F.J.; Rosety-Rodriguez, M. Correlation between glutathione peroxidase activity and anthropometrical parameters in adolescents with Down syndrome. Res. Dev. Disabil. 2007, 28, 105–108. [Google Scholar] [CrossRef]
- Pavao, M.; Cordeiro, C.; Costa, A.; Raposo, J.; Santos, M.; Neve, J.; Viegas-Crespo, A.M. Comparison of whole-blood glutathione peroxidase activity, levels of serum selenium, and lipid peroxidation in subjects from the fishing and rural communities of ‘Rabo de Peixe’ village, San Miguel Island, the Azores’ Archipelago, Portugal. Biol. Trace Elem. Res. 2003, 92, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Al-Gadani, Y.; El-Ansary, A.; Attas, O.; Al-Ayadhi, L. Metabolic biomarkers related to oxidative stress and antioxidant status in Saudi autistic children. Clin. Biochem. 2009, 42, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.K.; Sindhu, K.K. Oxidative stress and metabolic syndrome. Life Sci. 2009, 84, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.G.; Gongora, M.C.; Guzik, T.J.; Widder, J. Oxidative stress and hypertension. J. Am. Soc. Hypertens. 2007, 1, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Hermsdorff, H.H.; Puchau, B.; Volp, A.C.; Barbosa, K.B.; Bressan, J.; Zulet, M.A.; Martínez, A.J. Dietary total antioxidant capacity is inversely related to central adiposity as well as to metabolic and oxidative stress markers in healthy young adults. Nutr. Metab. (Lond.) 2011, 8, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skalicky, J.; Muzakova, V.; Kandar, R.; Meloun, M.; Rousar, T.; Palicka, V. Evaluation of oxidative stress and inflammation in obese adults with metabolic syndrome. Clin. Chem. Lab. Med. 2008, 46, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Kempaiah, R.K.; Srinivasan, K. Beneficial influence of dietary curcumin, capsaicin and garlic on erythrocyte integrity in high-fat fed rats. J. Nutr. Biochem. 2006, 17, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Demori, I.; Voci, A.; Fugassa, E.; Burland, B. Combined effects of high-fat diet and ethanol induce oxidative stress in rat liver. Alcohol 2006, 40, 185–191. [Google Scholar] [CrossRef]
- Gómez-Pérez, Y.; Amengual-Cladera, E.; Català-Niell, A.; Thomàs-Moyà, E.; Gianotti, M.; Proenza, A.M.; Lladó, I. Gender dimorphism in high-fat-diet induced insulin resistance in skeletal muscle of aged rats. Cell Physiol. Biochem. 2008, 22, 539–548. [Google Scholar] [CrossRef]
- Ulla, A.; Alam, M.A.; Sikder, B.; Sumi, F.A.; Rahman, M.M.; Habib, Z.F.; Mohammed, M.K.; Subhan, N.; Hossain, H.; Reza, H.M. Supplementation of Syzygium cumini seed powder prevented obesity, glucose intolerance, hyperlipidemia and oxidative stress in high carbohydrate high fat diet induced obese rats. BMC Complement. Altern. Med. 2017, 17, 289. [Google Scholar] [CrossRef]
- Kindel, T.L.; Stranden, J.L. Bariatric surgery as a treatment for heart failure: Review of the literature and potential mechanisms. Surg. Obes. Relat. Dis. 2018, 14, 117–122. [Google Scholar] [CrossRef]
- Schulze, P.C.; Drosatos, K.; Goldberg, I.J. Lipid use and misuse by the heart. Circ. Res. 2016, 188, 1736–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, H.; John, A.; Howarth, F.C. Alterations in glutathione redox metabolism, oxidative stress, and mitochondrial function in the left ventricle of elderly Zucker diabetic fatty rat heart. Int. J. Mol. Sci. 2012, 13, 16241–16254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frasier, C.R.; Moukdar, F.; Patel, H.D. Redox-dependent increases in glutathione reductase and exercise preconditioning: Role of NADPH oxidase and mitochondria. Cardiovasc. Res. 2013, 98, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Saari, J.T.; Kang, Y.J. Weak antioxidant defenses make the heart a target for damage in copper-deficient rats. Free Radic. Biol. Med. 1994, 17, 529–536. [Google Scholar] [CrossRef]
- Rindler, P.M.; Plafker, S.M.; Szweda, L.I.; Kinter, M. High dietary fat selectively increases catalase expression within cardiac mitochondria. J. Biol. Chem. 2013, 288, 1979–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Littlejohns, B.; Lin, H.; Angelini, G.D.; Halestrap, A.P.; Suleiman, M.S. Switching back to normal diet following high-fat diet feeding reduces cardiac vulnerability to ischaemia and reperfusion injury. Cell Physiol. Biochem. 2014, 34, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.F.; Singal, P.K. Antioxidant and oxidative stress changes during heart failure subsequent to myocardial infarction in rats. Am. J. Pathol. 1996, 148, 291–300. [Google Scholar]
- Heneghan, H.M.; Meron-Eldar, S.; Brethauer, S.A.; Schauer, P.R.; Young, J.B. Effect of bariatric surgery on cardiovascular risk profile. Am. J. Cardiol. 2011, 108, 1499–1507. [Google Scholar] [CrossRef]
- Frezza, E.E.; Wei, C.; Wachtel, M.S. Is surgery the next answer to treat obesity-related hypertension? J. Clin. Hypertens. (Greenwich) 2009, 11, 284–288. [Google Scholar] [CrossRef]
- Cavarretta, E.; Casella, G.; Cali, B. Cardiac remodelling in obese patients after laparoscopic sleeve gastrectomy. World J. Surg. 2013, 37, 565–572. [Google Scholar] [CrossRef]
- Wojciechowska, C.; Romuk, E.; Tomasik, A.; Skrzep-Poloczek, B.; Nowalany-Kozielska, E.; Birkner, E.; Jacheć, W. Oxidative stress markers and C-reactive protein are related to severity of heart failure in patients with dilated cardiomyopathy. Mediat. Inflamm. 2014, 2014, 147040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keith, M.; Geranmayegan, A.; Sole, M.J.; Kurian, R.; Robinson, A.; Omran, A.S.; Jeejeebhoy, K.N. Increased oxidative stress in patients with congestive heart failure. J. Am. Coll. Cardiol. 1998, 31, 1352–1356. [Google Scholar] [CrossRef] [Green Version]
- Simeunovic, D.; Seferovic, P.M.; Ristic, A.D.; Nikolic, D.; Risimic, D.; Seferovic, J.; Maksimovic, R.; Nedeljkovic, I.; Karan, R.; Bajcetic, M. Evaluation of oxidative stress markers and catecholamine changes in patients with dilated cardiomyopathy before and after cardiopulmonary exercise testing. Hell. J. Cardiol. 2015, 56, 394–401. [Google Scholar]
- Nijhawan, S.; Richards, W.; O’Hea, M.F.; Audia, J.P.; Alvarez, D.F. Bariatric surgery rapidly improves mitochondrial respiration in morbidly obese patients. Surg. Endosc. 2013, 27, 4569–4573. [Google Scholar] [CrossRef]
- Hofso, D.; Nordstrand, N.; Johnson, L.K. Obesity-related cardiovascular risk factors after weight loss: A clinical trial comparing gastric bypass surgery and intensive lifestyle intervention. Eur. J. Endocrinol. 2010, 163, 735–745. [Google Scholar] [CrossRef] [Green Version]
- Michiels, C.; Raes, M.; Toussaint, O.; Remacle, J. Importance of Se-glutathione peroxidase, catalase, and Cu/Zn-SOD for cell survival against oxidative stress. Free Radic. Biol. Med. 1994, 17, 235–248. [Google Scholar] [CrossRef]
- Ansari, J.A.; Bhandari, U.; Pillai, K.K.; Haque, S.E. Effect of rosuvastatin on obesity-induced cardiac oxidative stress in Wistar rats-a preliminary study. Indian J. Exp. Biol. 2012, 50, 216–222. [Google Scholar]
- Sawyer, D.B.; Siwik, A.D.; Xiao, L. Role of oxidative stress in myocardial hypertrophy and failure. J. Mol. Cell. Cardiol. 2002, 34, 379–388. [Google Scholar] [CrossRef]
- Li, T.; Danelisen, I.; Bello-Klein, A.K.; Singal, P. Effects of probucol on changes of antioxidant enzymes in adriamycin-induced cardiomyopathy in rats. Cardiovasc. Res. 2000, 46, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Amirkhizi, F.; Siassi, F.; Minaie, S.; Djalali, M.; Rahimi, A.; Chamari, M. Is obesity associated with increased plasma lipid peroxidation and oxidative stress in women. ARYA Atheroscler. J. 2007, 2, 189–192. [Google Scholar]
- Romuk, E.; Wojciechowska, C.; Jacheć, W.; Nowak, J.; Niedziela, J.; Malinowska-Borowska, J.; Głogowska-Gruszka, A.; Birkner, E.; Rozentryt, P. Comparison of oxidative stress parameters in heart failure patients depending on ischaemic or nonischaemic aetiology. Oxid. Med. Cell. Longev. 2019. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, D.B. Oxidative stress in heart failure: What are we missing? Am. J. Med. Sci. 2011, 342, 120–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.; Marwick, T.H. Obesity cardiomyopathy: Diagnosis and therapeutic implications. Nat. Clin. Pract. Cardiovasc. Med. 2007, 4, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.L.; Lang, C.H. Etiology of alcoholic cardiomyopathy: Mitochondria, oxidative stress and apoptosis. Int. J. Biochem. Cell Biol. 2017, 89, 125–135. [Google Scholar] [CrossRef]
- Pei, J.; Fu, W.; Yang, L.; Zhang, Z.; Liu, Y. Oxidative stress is involved in the pathogenesis of Keshan disease (an endemic dilated cardiomyopathy) in China. Oxid. Med. Cell Longev. 2013, 2013, 474203. [Google Scholar] [CrossRef]
- Machado, F.S.; Tanowitz, H.B.; Ribeiro, A.L. Pathogenesis of Chagas cardiomyopathy: Role of inflammation and oxidative stress. J. Am. Heart. Assoc. 2013, 2, e000539. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxidative Stress Marker | DJOS-Operated | SHAM-Operated | p ANOVA | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| HF/HF | HF/CD | CD/HF | CD/CD | HF/HF | HF/CD | CD/HF | CD/CD | Group | Op. | Int. | |
| GR (IU/g) | 27.56 ± 1.39 | 15.41 ± 1.95 | 14.19 ± 2.35 | 16.45 ± 2.22 | 66.19 ± 17.50 | 30.30 ± 7.77 | 30.53 ± 5.01 | 14.09 ± 2.29 | <0.001 | <0.001 | <0.001 |
| CAT (IU/g) | 59.91 ± 3.55 | 104.42 ± 12.84 | 84.68 ± 42.72 | 92.33 ± 9.48 | 306.63 ± 62.55 | 93.14 ± 7.01 | 78.80 ± 14.33 | 100.82 ± 27.05 | <0.001 | <0.001 | <0.001 |
| GPx (IU/g) | 15.81 ± 1.23 | 19.42 ± 1.70 | 18.76 ± 5.28 | 18.28 ± 1.82 | 14.58 ± 4.72 | 14.76 ± 2.44 | 15.49 ± 2.44 | 16.35 ± 1.38 | 0.253 | <0.01 | 0.506 |
| GST (IU/g) | 3.44 ± 0.61 | 3.74 ± 0.30 | 2.89 ± 1.08 | 3.50 ± 0.57 | 2.66 ± 0.73 | 3.58 ± 0.82 | 3.34 ± 0.25 | 3.43 ± 0.48 | 0.121 | 0.632 | 0.074 |
| Total SOD (NU/mg) | 79.75 ± 4.71 | 129.37 ± 27.69 | 105.22 ± 23.89 | 93.70 ± 9.57 | 146.11 ± 6.99 | 118.33 ± 17.91 | 77.54 ± 11.64 | 123.13 ± 26.37 | <0.001 | <0.01 | <0.001 |
| MnSOD (NU/mg) | 67.40 ± 4.63 | 89.05 ± 2.89 | 82.62 ± 20.55 | 80.50 ± 7.10 | 45.38 ± 22.15 | 81.13 ± 16.28 | 66.17 ± 8.26 | 69.25 ± 23.10 | <0.001 | <0.01 | 0.700 |
| CuZnSOD (NU/mg) | 11.96 (11.54–12.74) | 42.38 (27.63–49.71) | 19.98 (14.80–30.62) | 16.47 (12.94–18.51) | 16.85 (14.83–18.81) | 26.67 (15.80–33.67) | 10.34 (8.75–11.82) | 59.60 (43.52–93.75) | <0.001 | 0.333 | <0.001 |
| MDA (μmol/g) | 3.48 ± 1.33 | 2.47 ± 0.24 | 6.10 ± 2.58 | 2.94 ± 0.79 | 8.66 ± 1.65 | 10.53 ± 1.91 | 2.08 ± 0.78 | 2.07 ± 0.58 | <0.001 | <0.001 | <0.001 |
| Oxidative Stress Markers Post hoc | DJOS vs. SHAM | DJOS | SHAM | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1: HF/HF | 2: HF/CD | 3: CD/HF | 4: CD/CD | 1 vs. 2 | 1 vs. 3 | 1 vs. 4 | 2 vs. 3 | 2 vs. 4 | 3 vs. 4 | 1 vs. 2 | 1 vs. 3 | 1 vs. 4 | 2 vs. 3 | 2 vs. 4 | 3 vs. 4 | |
| Erythrocytes | ||||||||||||||||
| GR (IU/g) | 0.189 | <0.001 | <0.001 | 0.557 | <0.001 | <0.05 | 0.207 | <0.001 | <0.001 | 0.226 | 0.114 | <0.001 | 0.594 | <0.001 | <0.05 | <0.001 |
| CAT (IU/g) | <0.001 | <0.05 | <0.001 | <0.05 | <0.001 | <0.001 | <0.001 | 0.056 | 0.375 | 0.259 | <0.01 | <0.001 | <0.01 | <0.001 | 0.490 | <0.001 |
| GPx (IU/g) | <0.001 | <0.01 | 0.939 | <0.05 | 0.324 | <0.001 | <0.001 | <0.01 | <0.001 | 0.180 | 0.771 | 0.542 | 0.094 | 0.758 | 0.178 | 0.296 |
| GST (IU/g) | <0.001 | 0.303 | <0.001 | 0.178 | <0.001 | <0.001 | <0.001 | 0.382 | <0.001 | <0.001 | 0.058 | <0.001 | 0.127 | <0.001 | <0.01 | <0.001 |
| Total SOD (NU/mg) | 0.876 | 0.763 | 0.857 | 0.542 | <0.01 | 0.181 | <0.001 | 0.115 | 0.293 | <0.01 | <0.01 | 0.156 | <0.01 | 0.091 | 0.898 | 0.070 |
| MDA (μmol/g) | <0.001 | 0.142 | 0.774 | <0.001 | <0.001 | <0.01 | <0.001 | 0.205 | <0.001 | <0.001 | 0.154 | 0.356 | <0.05 | 0.621 | <0.01 | <0.01 |
| Heart Muscle | ||||||||||||||||
| GR (IU/g) | <0.001 | <0.001 | <0.001 | 0.572 | <0.01 | <0.01 | <0.05 | 0.771 | 0.803 | 0.589 | <0.001 | <0.001 | <0.001 | 0.956 | <0.001 | <0.001 |
| CAT (IU/g) | <0.001 | 0.529 | 0.742 | 0.635 | <0.05 | 0.171 | 0.075 | 0.273 | 0.500 | 0.669 | <0.001 | <0.001 | <0.001 | 0.425 | 0.668 | 0.222 |
| GPx (IU/g) | 0.461 | <0.01 | 0.064 | 0.270 | <0.05 | 0.095 | 0.160 | 0.705 | 0.513 | 0.782 | 0.911 | 0.586 | 0.291 | 0.676 | 0.361 | 0.618 |
| Total SOD (NU/mg) | <0.001 | 0.300 | <0.05 | <0.01 | <0.001 | <0.05 | 0.192 | <0.05 | <0.01 | 0.280 | <0.05 | <0.001 | <0.05 | <0.001 | 0.650 | <0.001 |
| MnSOD (NU/mg) | <0.05 | 0.380 | 0.113 | 0.215 | <0.05 | 0.096 | 0.150 | 0.475 | 0.343 | 0.813 | <0.001 | <0.05 | <0.01 | 0.154 | 0.190 | 0.904 |
| CuZnSOD (NU/mg) | 0.222 | <0.05 | <0.01 | <0.001 | <0.001 | <0.05 | 0.277 | <0.05 | <0.001 | 0.188 | 0.116 | <0.05 | <0.001 | <0.001 | <0.001 | <0.001 |
| MDA (μmol/g) | <0.001 | <0.001 | <0.001 | 0.303 | 0.230 | <0.01 | 0.514 | <0.001 | 0.578 | <0.001 | <0.05 | <0.001 | <0.001 | <0.001 | <0.001 | 0.990 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skrzep-Poloczek, B.; Poloczek, J.; Chełmecka, E.; Dulska, A.; Romuk, E.; Idzik, M.; Kazura, W.; Nabrdalik, K.; Gumprecht, J.; Jochem, J.; et al. The Oxidative Stress Markers in the Erythrocytes and Heart Muscle of Obese Rats: Relate to a High-Fat Diet but Not to DJOS Bariatric Surgery. Antioxidants 2020, 9, 183. https://doi.org/10.3390/antiox9020183
Skrzep-Poloczek B, Poloczek J, Chełmecka E, Dulska A, Romuk E, Idzik M, Kazura W, Nabrdalik K, Gumprecht J, Jochem J, et al. The Oxidative Stress Markers in the Erythrocytes and Heart Muscle of Obese Rats: Relate to a High-Fat Diet but Not to DJOS Bariatric Surgery. Antioxidants. 2020; 9(2):183. https://doi.org/10.3390/antiox9020183
Chicago/Turabian StyleSkrzep-Poloczek, Bronisława, Jakub Poloczek, Elżbieta Chełmecka, Agnieszka Dulska, Ewa Romuk, Maciej Idzik, Wojciech Kazura, Katarzyna Nabrdalik, Janusz Gumprecht, Jerzy Jochem, and et al. 2020. "The Oxidative Stress Markers in the Erythrocytes and Heart Muscle of Obese Rats: Relate to a High-Fat Diet but Not to DJOS Bariatric Surgery" Antioxidants 9, no. 2: 183. https://doi.org/10.3390/antiox9020183