Changes in Phenolics and Fatty Acids Composition and Related Gene Expression during the Development from Seed to Leaves of Three Cultivated Cardoon Genotypes

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Plant Growth and Sample Collection
2.3. RNA Extraction and qRT-PCR
2.4. Ultrasound-Assisted Extraction of Polyphenolic Compounds
2.5. Antioxidant Activity: ABTS Assay
2.6. Antioxidant Activity: DPPH Assay
2.7. Antioxidant Activity: FRAP Assay
2.8. HRMS Orbitrap Analysis of Bioactive Polyphenols
2.9. Oil Extraction
2.10. Fatty Acids Analysis
2.11. Statistical Analysis
3. Results and Discussion
3.1. Polyphenol Profiling of Cardoon Genotypes
3.2. Qualitative and Quantitative Profile of (Seeds, Hypocotyls, Cotyledons, and Leaves) Oil
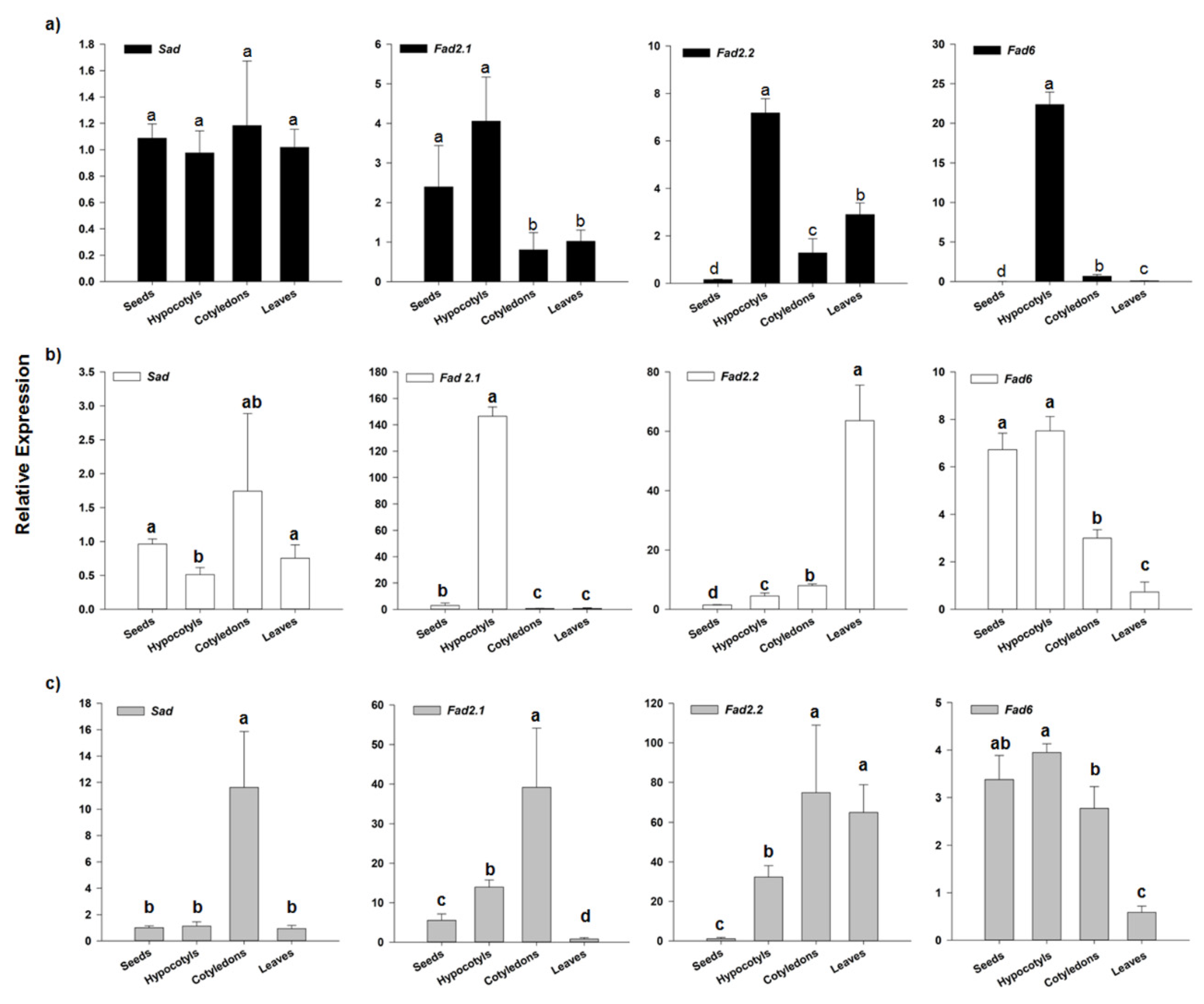
3.3. Transcriptional Analysis of Key Biosynthetic Genes in Chlorogenic Acid and Monounsaturated Fatty Acids
3.3.1. HQT Expression Analysis
3.3.2. SAD and FAD2 Expression Analysis
3.4. Antioxidant Activity of Polyphenolic Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bianco, V.V. Present situation and future potential of artichoke in the mediterranean basin. Acta Hortic. 2005, 681, 39–58. [Google Scholar] [CrossRef]
- Pappalardo, H.D.; Toscano, V.; Puglia, G.D.; Genovese, C.; Raccuia, S.A. Cynara cardunculus L. as a Multipurpose Crop for Plant Secondary Metabolites Production in Marginal Stressed Lands. Front. Plant Sci. 2020, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Ciancolini, A.; Alignan, M.; Pagnotta, M.A.; Vilarem, G.; Crinò, P. Selection of Italian cardoon genotypes as industrial crop for biomass and polyphenol production. Ind. Crop. Prod. 2013, 51, 145–151. [Google Scholar] [CrossRef]
- Foti, S.; Mauromicale, G.; Raccuia, S.A.; Fallico, B.; Fanella, F. Possible alternative utilization of Cynara spp. I. Biomass, grain yield and chemical composition of grain. Ind. Crops Prod. 1999, 10, 219–228. [Google Scholar] [CrossRef]
- Petropoulos, S.; Fernandes, Â.; Pereira, C.; Tzortzakis, N.; Vaz, J.; Soković, M.; Barros, L.; Ferreira, I.C. Bioactivities, chemical composition and nutritional value of Cynara cardunculus L. seeds. Food Chem. 2019, 289, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Ierna, A.; Mauro, R.P.; Mauromicale, G.; Catania, S.; Lancia, S.V.; Industriale, Z.; Palma, B.; Catania, I. Biomass, grain and energy yield in Cynara cardunculus L. as affected by fertilization, genotype and harvest time. Biomass Bioenergy 2011, 36, 404–410. [Google Scholar] [CrossRef]
- Falleh, H.; Ksouri, R.; Chaieb, K.; Karray-Bouraoui, N.; Trabelsi, N.; Boulaaba, M.; Abdelly, C. Phenolic composition of Cynara cardunculus L. organs, and their biological activities. Comptes Rendus Biol. 2008, 331, 372–379. [Google Scholar] [CrossRef]
- Calabria, L.M.; Emerenciano, V.P.; Ferreira, M.J.P.; Scotti, M.T.; Mabry, T.J. A Phylogenetic Analysis of Tribes of the Asteraceae Based on Phytochemical Data. Nat. Prod. Commun. 2007, 2, 277–285. [Google Scholar] [CrossRef]
- Lattanzio, V.; Cardinali, A.; Di Venere, D.; Linsalata, V.; Palmieri, S. Browning phenomena in stored artichoke (Cynara scolymus L.) heads: Enzymic or chemical reactions? Food Chem. 1994, 50, 1–7. [Google Scholar] [CrossRef]
- Williamson, G.; Holst, B. Dietary reference intake (DRI) value for dietary polyphenols: Are we heading in the right direction? Br. J. Nutr. 2008, 99, S55–S58. [Google Scholar] [CrossRef] [Green Version]
- Durazzo, A.; Foddai, M.; Temperini, A.; Azzini, E.; Venneria, E.; Lucarini, M.; Finotti, E.; Maiani, G.; Crinò, P.; Saccardo, F.; et al. Antioxidant Properties of Seeds from Lines of Artichoke, Cultivated Cardoon and Wild Cardoon. Antioxidants 2013, 2, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouveia, S.C.; Castilho, P.C. Phenolic composition and antioxidant capacity of cultivated artichoke, Madeira cardoon and artichoke-based dietary supplements. Food Res. Int. 2012, 48, 712–724. [Google Scholar] [CrossRef]
- Mandim, F.; Petropoulos, S.A.; Dias, M.I.; Pinela, J.; Kostic, M.; Soković, M.; Santos-Buelga, C.; Ferreira, I.C.F.R.; Barros, L. Seasonal variation in bioactive properties and phenolic composition of cardoon (Cynara cardunculus var. altilis) bracts. Food Chem. 2020, 336, 127744. [Google Scholar] [CrossRef]
- Racchi, M.; Daglia, M.; Lanni, C.; Papetti, A.; Govoni, S.; Gazzani, G. Antiradical Activity of Water Soluble Components in Common Diet Vegetables. J. Agric. Food Chem. 2002, 50, 1272–1277. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.A. Spectrophotometric determination of antioxidant activity. Redox Rep. 1996, 2, 161–171. [Google Scholar] [CrossRef]
- Cho, A.S.; Jeon, S.M.; Kim, M.J.; Yeo, J.; Seo, K.I.; Choi, M.S.; Lee, M.K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef]
- Dos Santos, M.D.; Almeida, M.C.; Lopes, N.P.; de Souza, G.E.P. Evaluation of the anti-inflammatory, analgesic and antipyretic activities of the natural polyphenol chlorogenic acid. Biol. Pharm. Bull. 2006, 29, 2236–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D. Chlorogenic Acid: Recent Advances on Its Dual Role as a Food Additive and a Nutraceutical against Metabolic Syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef] [Green Version]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial Activity and Mechanism of Action of Chlorogenic Acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [CrossRef]
- Niggeweg, R.; Michael, A.J.; Martin, C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 2004, 22, 746–754. [Google Scholar] [CrossRef]
- Mahesh, V.; Million-Rousseau, R.; Ullmann, P.; Chabrillange, N.; Bustamante, J.; Mondolot, L.; Morant, M.; Noirot, M.; Hamon, S.; de Kochko, A.; et al. Functional characterization of two p-coumaroyl ester 3′-hydroxylase genes from coffee tree: Evidence of a candidate for chlorogenic acid biosynthesis. Plant Mol. Biol. 2007, 64, 145–159. [Google Scholar] [CrossRef]
- Sonnante, G.; D’Amore, R.; Blanco, E.; Pierri, C.L.; De Palma, M.; Luo, J.; Tucci, M.; Martin, C. Novel hydroxycinnamoyl-coenzyme A quinate transferase genes from artichoke are involved in the synthesis of chlorogenic acid. Plant Physiol. 2010, 153, 1224–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comino, C.; Hehn, A.; Moglia, A.; Menin, B.; Bourgaud, F.; Lanteri, S.; Portis, E. The isolation and mapping of a novel hydroxycinnamoyltransferase in the globe artichoke chlorogenic acid pathway. BMC Plant Biol. 2009, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Cheevarungnapakul, K.; Khaksar, G.; Panpetch, P.; Boonjing, P.; Sirikantaramas, S. Identification and Functional Characterization of Genes Involved in the Biosynthesis of Caffeoylquinic Acids in Sunflower (Helianthus annuus L.). Front. Plant Sci. 2019, 10, 968. [Google Scholar] [CrossRef] [Green Version]
- Raccuia, S.A.; Melilli, M.G. Biomass and grain oil yields in Cynara cardunculus L. genotypes grown in a Mediterranean environment. Field Crop. Res. 2007, 101, 187–197. [Google Scholar] [CrossRef]
- Gominho, J.; Lourenc, A.; Palma, P.; Lourenc, M.E.; Curt, M.D.; Fernández, J.; Pereira, H. Large scale cultivation of Cynara cardunculus L. for biomass production—A case study. Ind. Crops Prod. 2011, 33, 1–6. [Google Scholar] [CrossRef]
- Meï, C.; Michaud, M.; Cussac, M.; Albrieux, C.; Gros, V.; Maréchal, E.; Block, M.A.; Jouhet, J.; Rébeillé, F. Levels of polyunsaturated fatty acids correlate with growth rate in plant cell cultures. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Upchurch, R.G. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Somerville, C.; Browse, J. Plant Lipids: Metabolism, Mutants, and Membranes. Science 1991, 252, 80–87. [Google Scholar] [CrossRef]
- Browse, J.; Somerville, C. Glycerolipid Synthesis: Biochemistry and Regulation. Annu. Rev. Plant Biol. 1991, 42, 467–506. [Google Scholar] [CrossRef]
- Dar, A.A.; Choudhury, A.R.; Kancharla, P.K.; Arumugam, N. The FAD2 Gene in Plants: Occurrence, Regulation, and Role. Front. Plant Sci. 2017, 8, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Sharma, A.; Upadhyaya, K.C. Vegetable Oil: Nutritional and Industrial Perspective. Curr. Genom. 2016, 17, 230–240. [Google Scholar] [CrossRef] [Green Version]
- Orsavova, J.; Misurcova, L.; Ambrozova, J.; Vicha, R.; Mlcek, J. Fatty Acids Composition of Vegetable Oils and Its Contribution to Dietary Energy Intake and Dependence of Cardiovascular Mortality on Dietary Intake of Fatty Acids. Int. J. Mol. Sci. 2015, 16, 12871–12890. [Google Scholar] [CrossRef] [PubMed]
- Clé, C.; Hill, L.M.; Niggeweg, R.; Martin, C.R.; Guisez, Y.; Prinsen, E.; Jansen, M.A.K.K. Modulation of chlorogenic acid biosynthesis in Solanum lycopersicum; consequences for phenolic accumulation and UV-tolerance. Phytochemistry 2008, 69, 2149–2156. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M. Quantification strategies in real-time PCR. AZ Quant. PCR 2004, 1, 87–112. [Google Scholar]
- Docimo, T.; De Stefano, R.; Cappetta, E.; Lisa Piccinelli, A.; Celano, R.; De Palma, M.; Tucci, M. Physiological, biochemical, and metabolic responses to short and prolonged saline stress in two cultivated cardoon genotypes. Plants 2020, 9, 554. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Garros, L.; Drouet, S.; Renouard, S.; Lainé, E.; Hano, C. Green Ultrasound Assisted Extraction of trans Rosmarinic Acid from Plectranthus scutellarioides (L.) R.Br. Leaves. Plants 2019, 8, 50. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma as a measure of antioxodant. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Graziani, G.; Ritieni, A.; Cirillo, A.; Cice, D.; Di Vaio, C. Effects of Biostimulants on Annurca Fruit Quality and Potential Nutraceutical Compounds at Harvest and during Storage. Plants 2020, 9, 775. [Google Scholar] [CrossRef]
- Kostić, M.D.; Joković, N.M.; Stamenković, O.S.; Rajković, K.M.; Milić, P.S.; Veljković, V.B. Optimization of hempseed oil extraction by n-hexane. Ind. Crop. Prod. 2013, 48, 133–143. [Google Scholar] [CrossRef]
- Pandino, G.; Lombardo, S.; Mauromicale, G.; Williamson, G. Phenolic acids and flavonoids in leaf and floral stem of cultivated and wild Cynara cardunculus L. genotypes. Food Chem. 2011, 126, 417–422. [Google Scholar] [CrossRef]
- Dias, M.I.; Barros, L.; Barreira, J.C.M.; Alves, M.J.; Barracosa, P.; Ferreira, I.C.F.R. Phenolic profile and bioactivity of cardoon (Cynara cardunculus L.) inflorescence parts: Selecting the best genotype for food applications. Food Chem. 2018, 268, 196–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, P.A.B.; Santos, S.A.O.; Guerra, Â.R.; Guerreiro, O.; Freire, C.S.R.; Rocha, S.M.; Duarte, M.F.; Silvestre, A.J.D. Phenolic composition and antioxidant activity of different morphological parts of Cynara cardunculus L. var. altilis (DC). Ind. Crop. Prod. 2014, 61, 460–471. [Google Scholar] [CrossRef]
- Lattanzio, V.; Kroon, P.A.; Linsalata, V.; Cardinali, A. Globe artichoke: A functional food and source of nutraceutical ingredients. J. Funct. Foods 2009, 1, 131–144. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus × acerifolia plants during Mediterranean summers. New Phytol. 2015, 207, 613–626. [Google Scholar] [CrossRef]
- Penfield, S.; Graham, S.; Graham, I.A. Storage reserve mobilization in germinating oilseeds: Arabidopsis as a model system. Biochem. Soc. Trans. 2005, 33, 380–383. [Google Scholar] [CrossRef]
- Gominho, J.; Curt, M.D.; Lourenço, A.; Fernández, J.; Pereira, H. Cynara cardunculus L. as a biomass and multi-purpose crop: A review of 30 years of research. Biomass Bioenergy 2018, 109, 257–275. [Google Scholar] [CrossRef]
- Raccuia, S.A.; Piscioneri, I.; Sharma, N.; Melilli, M.G. Genetic variability in Cynara cardunculus L. domestic and wild types for grain oil production and fatty acids composition. Biomass Bioenergy 2011, 35, 3167–3173. [Google Scholar] [CrossRef]
- Francisco, M.; Elena Cartea, M.; Maria Butron, A.; Sotelo, T.; Velasco, P. Environmental and Genetic Effects on Yield and Secondary Metabolite Production in Brassica rapa Crops. J. Agric. Food Chem. 2012, 60, 5507–5514. [Google Scholar] [CrossRef]
- Maccarone, E.; Fallico, B.; Fanella, F.; Mauromicale, G.; Raccuia, S.A. Possible alternative utilization of Cynara spp. II. Chemical characterization of their grain oil. Ind. Crops Prod. 1999, 10, 229–237. [Google Scholar] [CrossRef]
- Curt, M.D.; Sánchez, G.; Fernández, J. The potential of Cynara cardunculus L. for seed oil production in a perennial cultivation system. Biomass Bioenergy 2002, 23, 33–46. [Google Scholar] [CrossRef]
- Angelova, V.; Nemska, M.P.; Krustev, L. Chemical composition of cardoon (Cynara cardunculus L.) Grown in south bulgaria. AGROFOR Int. J. 2019, 4, 100–110. [Google Scholar]
- Volpi E Silva, N.; Mazzafera, P.; Cesarino, I. Should I stay or should I go: Are chlorogenic acids mobilized towards lignin biosynthesis? Phytochemistry 2019, 166, 112063. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Navarre, D.; Kuhl, J.C.; Pantoja, A.; Pillai, S.S. Differential effects of environment on potato phenylpropanoid and carotenoid expression. BMC Plant Biol. 2012, 12, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payyavula, R.S.; Shakya, R.; Sengoda, V.G.; Munyaneza, J.E.; Swamy, P.; Navarre, D.A. Synthesis and regulation of chlorogenic acid in potato: Rerouting phenylpropanoid flux in HQT-silenced lines. Plant Biotechnol. J. 2015, 13, 551–564. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Bai, M.; He, H.; Wang, H.; Wu, H. Correlation of the temporal and spatial expression patterns of HQT with the biosynthesis and accumulation of chlorogenic acid in Lonicera japonica flowers. Hortic. Res. 2019, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Vijaya Kumar Reddy, C.; Sreeramulu, D.; Raghunath, M. Antioxidant activity of fresh and dry fruits commonly consumed in India. Food Res. Int. 2010, 43, 285–288. [Google Scholar] [CrossRef]
- Kollia, E.; Markaki, P.; Zoumpoulakis, P.; Proestos, C. Antioxidant activity of Cynara scolymus L. and Cynara cardunculus L. extracts obtained by different extraction techniques. Nat. Prod. Res. 2017, 31, 1163–1167. [Google Scholar] [CrossRef]
- Arnao, M.B. Some methodological problems in the determination of antioxidant activity using chromogen radicals: A practical case. Trends Food Sci. Technol. 2000, 11, 419–421. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Svecova, E.; Rea, E.; Lucini, L. Effects of saline stress on mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon genotypes grown in floating system. J. Sci. Food Agric. 2013, 93, 1119–1127. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Peak Number | Polyphenol | Retention Time (min) | Chemical Formula | Theoretical Mass (m/z) | Measured Mass (m/z) | Accuracy (ppm) |
|---|---|---|---|---|---|---|
| 13 | Luteolin | 10.99 | C15H10O6 | 285.04046 | 285.04071 | 0.88 |
| 15 | Apigenin | 11.45 | C15H10O5 | 269.04555 | 269.04605 | 1.86 |
| 6 | Diosmin | 9.96 | C28H32O15 | 607.16684 | 607.16821 | 2.26 |
| 5 | Apigenin-8-C-glucoside | 9.89 | C21H20O10 | 431.09837 | 431.09973 | 3.15 |
| 11 | (+/−) Naringenin | 10.58 | C15H12O5 | 271.06120 | 271.06165 | 1.66 |
| 4 | Quercitin-3-O-glucoside | 9.71 | C21H19O12 | 463.08820 | 463.08926 | 2.29 |
| 12 | Quercetin | 10.71 | C15H10O7 | 301.03538 | 301.03540 | 0.07 |
| 14 | Kaempferol | 11.31 | C15H10O6 | 285.04046 | 285.04077 | 1.09 |
| 9 | Myricetin | 10.06 | C15H10O8 | 317.03029 | 317.03119 | 2.84 |
| 7 | Naringin | 10.01 | C27H32O14 | 579.54123 | 579.54162 | 0.67 |
| 10 | Kaempferol-3-O-glucoside | 10.11 | C21H20O11 | 447.09328 | 447.09396 | 1.52 |
| 8 | Luteolin-7-O-glucoside | 10.05 | C21H20O12 | 447.09328 | 447.09311 | −0.38 |
| 3 | p-Coumaric acid | 8.66 | C9H8O3 | 163.04007 | 163.04056 | 3.00 |
| 2 | Chlorogenic acid | 7.85 | C16H18O9 | 353.08781 | 353.08856 | 2.12 |
| 1 | 4-Hydroxybenzoic acid | 6.76 | C7H6O3 | 137.02442 | 137.02489 | 3.43 |
| Phenolic Compounds | SEEDS | HYPOCOTYLS | COTYLEDONS | LEAVES | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gigante | Spagnolo | B. Avorio | Gigante | Spagnolo | B. Avorio | Gigante | Spagnolo | B. Avorio | Gigante | Spagnolo | B. Avorio | |
| 4-hydroxy benzoic acid | 2.750a | 2.510b | 2.040c | 5.620a | 3.760c | 3.910b | 12.592b | 8.795c | 16.771a | 1.221a | 0.661b | 0.329c |
| Vitexin | 107.300a | 91.400b | 83.700c | 121.700b | 111.710c | 250.110a | 350.432c | 876.529b | 1182.892a | 528.396c | 2100.344b | 2107.906a |
| luteolin-7-O-glucoside | 1.470b | 2.760a | 0.910c | 1.610c | 8.610b | 12.700a | 18.092c | 34.299a | 24.086b | 81.922c | 122.687a | 88.773b |
| naringin | 10.900c | 11.700b | 14.400a | 5.710c | 51.700b | 67.100a | 14.599b | 438.433a | 436.022a | 0.251b | 0.357a | 0.038c |
| chlorogenic acid | 1036.310c | 1201.990b | 1430.460a | 3461.130b | 1006.780c | 3735.790a | 786.031a | 769.301a | 463.390b | 1468.968b | 2467.679a | 2637.733a |
| coumaric acid | 1.790a | 1.840a | 1.820a | 3.620a | 3.410a | 3.720a | 0.180a | 0.080b | 0.080b | 0.080c | 0.560a | 0.200b |
| quercetin-3-glucoside | 0.090c | 0.270b | 1.160a | 0.700b | 0.180c | 1.180a | 1.587b | 2.670a | 1.970b | 3.386a | 3.618a | 2.050b |
| diosmin | 0.050a | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| kaempferol-3-O-glucoside | 2.180a | 2.140a | 2.200a | 4.910ab | 4.460b | 5.000a | 0.420b | 0.780a | 0.860a | 9.960a | 9.960a | 6.720b |
| myricetin | 0.440a | 0.440a | 0.450a | 0.880a | 0.890a | 0.880a | 2.470c | 3.110b | 8.470a | 6.884a | 6.063b | 5.493c |
| naringenin | 1.730a | 1.610b | 1.680ab | 3.130a | 3.110a | 3.110a | 0.120a | nd | 0.040b | 0.160c | 0.320b | 0.500a |
| luteolin | 0.320a | 0.020b | 0.020b | 0.040b | 0.200a | nd | 11.700c | 37.137a | 31.464b | 56.040c | 84.640a | 72.671b |
| kaempferol | 0.340a | 0.020b | 0.020b | 0.040b | 0.240a | nd | 3.600c | 4.300b | 7.900a | 4.800c | 7.360a | 5.780b |
| quercetin | 0.820 | nd | nd | nd | 1.640 | nd | nd | 0.020a | 0.020a | 36.580a | 21.300b | 14.160c |
| apigenin | 0.240a | 0.040b | 0.020c | nd | 0.080a | 0.040b | nd | 4.580b | 7.040a | 2.260c | 3.710b | 8.080a |
| Total polyphenols | 1166.730c | 1316.740b | 1538.880a | 3609.090b | 1196.770c | 4083.540a | 1201.823b | 2180.034a | 2181.005a | 2200.908b | 4829.259a | 4950.433a |
| Fatty Acids | Seeds | Hypocotyls | Cotyledons | Leaves | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gigante | Spagnolo | B. Avorio | Gigante | Spagnolo | B. Avorio | Gigante | Spagnolo | B. Avorio | Gigante | Spagnolo | B. Avorio | |
| Pentadeconoic (C15:0) | 0.123c | 0.283a | 0.173b | 0.172b | 0.182a | 0.143c | 1.852a | 0.578c | 0.711b | 0.985a | 0.441c | 0.611b |
| Palmitic (C16:0) | 59.868a | 44.265b | 56.708a | 49.361a | 47.722a | 40.791b | 63.320a | 44.594c | 50.348b | 44.484b | 34.330c | 61.643a |
| Margaric (C17:0) | 0.694b | 1.25a | 0.797b | 0.109a | 0.117a | 0.098b | 2.253a | 0.639b | 0.642b | 0.882a | 0.571b | 0.608b |
| Nonadecanoic (C19:0) | 0.013b | 0.024a | 0.01b4 | n.d. | n.d. | n.d. | n.d. | 0.252 | n.d. | 0.289 | n.d. | 0.187 |
| Arachidic (C20:0) | 4.446ab | 4.427a | 3.4b | n.d. | n.d. | 0.659 | n.d. | 8.176a | 5.157b | 7.635a | 3.852c | 5.309b |
| Behenic (C22:0) | 0.634a | 0.635a | 0.489b | 0.271b | 0.431a | 0.315b | 1.976b | 3.767a | 1.459c | 4.162a | 2.103c | 3.463b |
| Lignoceric (C24:0) | 0.457a | 0.246b | 0.432a | 0.326 | n.d. | n.d. | 2.508b | 5.408a | 0.979c | 5.219b | n.d. | 5.706a |
| Cerotic (C26:0) | 0.052 | n.d. | n.d. | n.d. | n.d. | n.d. | 0.465a | 4.414a | n.d. | 3.977 | n.d. | n.d. |
| Melissic (C30:0) | 0.015 | 0.03 | n.d. | n.d. | n.d. | 0.282 | n.d. | n.d. | 0.557 | n.d. | n.d. | n.d. |
| Palmitoleic (C16:1) | 0.372b | 0.816a | 0.348b | n.d. | 0.135 | n.d. | 0.607a | n.d. | 0.439b | 1.452a | 0.420c | 0.516b |
| Hexadicadienoic (C16:2) | 0.006b | 1.389a | 0.009b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.088 | n.d. |
| Hexadicatrienoic (C16:3) | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 9.774 | n.d. | n.d. | n.d. | n.d. | n.d. |
| Oleic (C18:1) | 19.271a | 18.395b | 15.828c | 6.54c | 7.869b | 12.278a | n.d. | 9.737a | 8.240b | 10.524b | 15.519a | 6.592c |
| Linoleic (C18:2) | 13.684c | 27.34a | 21.212b | 37.892b | 38.954b | 45.434a | 8.935c | 12.997b | 24.714a | 12.806b | 37.460a | 9.197c |
| Linolenic (C18:3) | 0.355a | 0.441a | 0.021b | 5.259a | 4.588b | n.d. | 8.312ab | 9.438a | 6.644b | 7.584a | 5.215c | 6.169b |
| Nonadecenoic (C19:1) | 0.01 | 0.018 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Gadoleic (C20:1) | n.d. | 0.441b | 0.566a | 0.07 | n.d. | n.d. | n.d. | n.d. | 0.110 | n.d. | n.d. | n.d. |
| GENOTYPE | DPPH | ABTS | FRAP | |
|---|---|---|---|---|
| mmol trolox kg−1 | mmol trolox kg−1 | mmol trolox kg−1 | ||
| Seeds | GIGANTE | 49.492 ± 0.275c | 87.237 ± 3.312c | 49.833 ± 1.283c |
| SPAGNOLO | 143.871 ± 0.942a | 135.413 ± 1.119b | 59.848 ± 3.340b | |
| BIANCO AVORIO | 137.946 ± 0.265b | 148.037 ± 1.127a | 107.728 ± 1.303a | |
| Hypocotyls | GIGANTE | 57.149 ± 0.377b | 667.365 ± 1.036a | 72.000 ± 0.954b |
| SPAGNOLO | 37.543 ± 0.311c | 317.206 ± 2.071c | 57.939 ± 1.043c | |
| BIANCO AVORIO | 60.866 ± 0.106a | 631.482 ± 8.285b | 79.758 ± 2.694a | |
| Cotyledons | GIGANTE | 19.192 ± 0.123c | 487.277 ± 2.111b | 47.776 ± 0.987c |
| SPAGNOLO | 63.951 ± 0.543a | 412.113 ± 0.112c | 79.148 ± 2.876a | |
| BIANCO AVORIO | 55.126 ± 0.156b | 543.012 ± 0.765a | 57.122 ± 1.832b | |
| Leaves | GIGANTE | 27.167 ± 0.551c | 510.315 ± 0.765c | 62.450 ± 0.432c |
| SPAGNOLO | 81.543 ± 0.111b | 654.216 ± 0.981b | 97.659 ± 0.876a | |
| BIANCO AVORIO | 91.856 ± 0.116a | 771.412 ± 0.665a | 91.675 ± 0.761b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graziani, G.; Docimo, T.; Palma, M.D.; Sparvoli, F.; Izzo, L.; Tucci, M.; Ritieni, A. Changes in Phenolics and Fatty Acids Composition and Related Gene Expression during the Development from Seed to Leaves of Three Cultivated Cardoon Genotypes. Antioxidants 2020, 9, 1096. https://doi.org/10.3390/antiox9111096
Graziani G, Docimo T, Palma MD, Sparvoli F, Izzo L, Tucci M, Ritieni A. Changes in Phenolics and Fatty Acids Composition and Related Gene Expression during the Development from Seed to Leaves of Three Cultivated Cardoon Genotypes. Antioxidants. 2020; 9(11):1096. https://doi.org/10.3390/antiox9111096
Chicago/Turabian StyleGraziani, Giulia, Teresa Docimo, Monica De Palma, Francesca Sparvoli, Luana Izzo, Marina Tucci, and Alberto Ritieni. 2020. "Changes in Phenolics and Fatty Acids Composition and Related Gene Expression during the Development from Seed to Leaves of Three Cultivated Cardoon Genotypes" Antioxidants 9, no. 11: 1096. https://doi.org/10.3390/antiox9111096