Polyphenols in Farm Animals: Source of Reproductive Gain or Waste?




Abstract
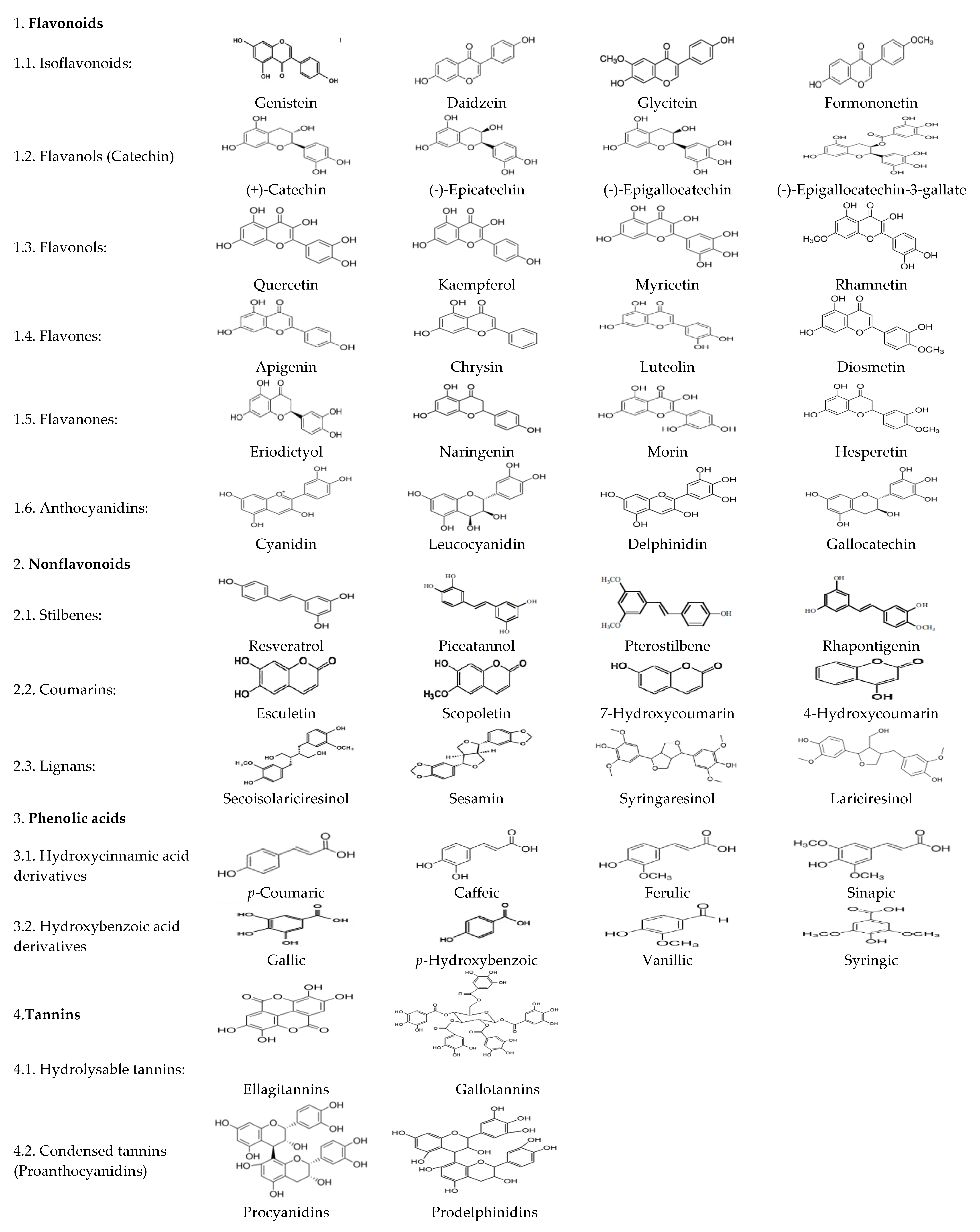
:1. Introduction
2. Polyphenols Sources in Animal Diets
3. Intake, Absorption, Bioavailability and Bioactivity of Polyphenols
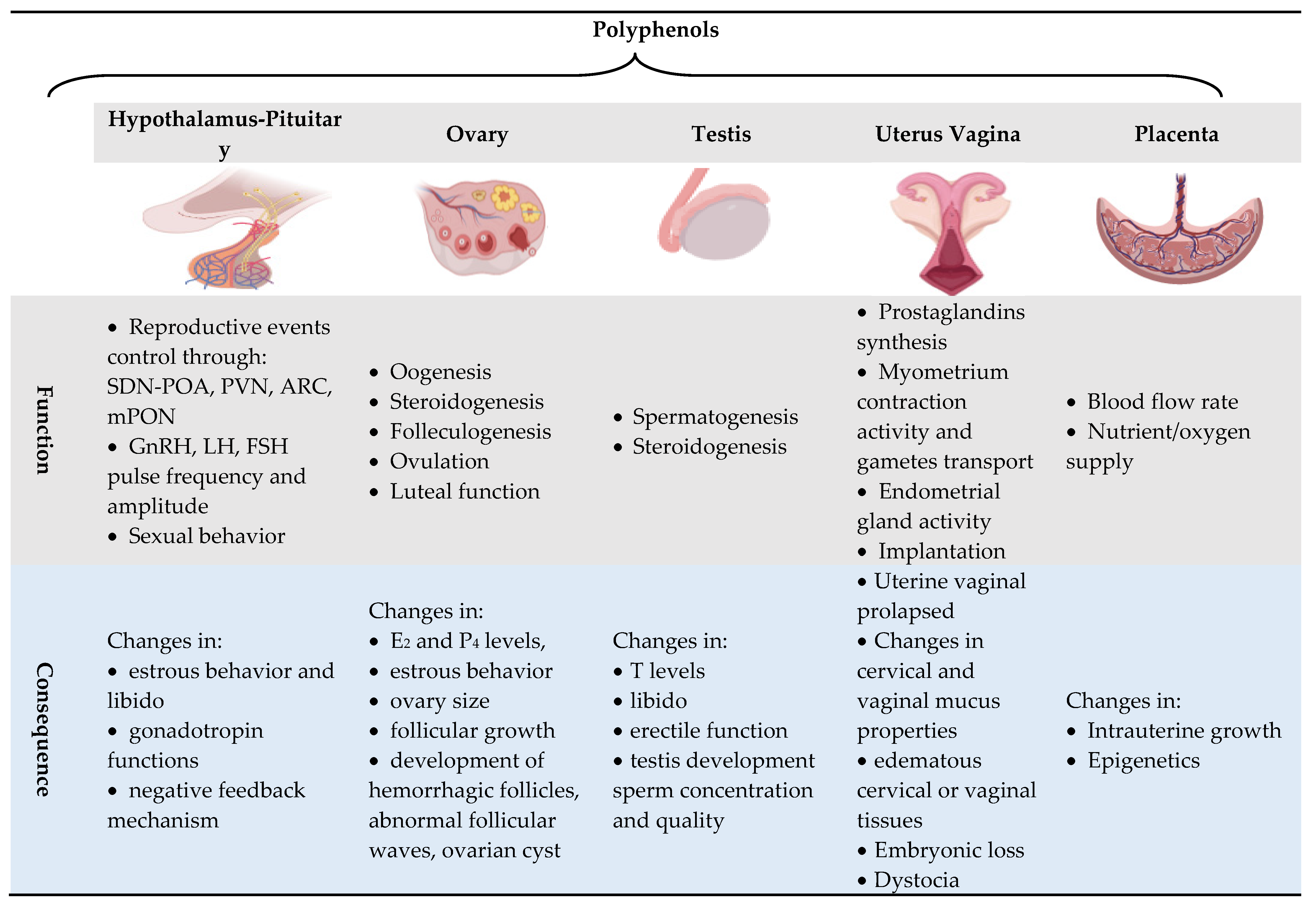
4. Polyphenols and In Vivo Reproductive Events
5. Polyphenols and In Vivo Reproductive Events
5.1. Sexual Behavior
5.2. Hormone Secretion and Function
5.3. Gametogenesis
5.4. Pregnancy and Fetal Programming
6. Polyphenols and In Vitro Assisted Reproduction Techniques
7. Polyphenols of Animal Origin and Human Health
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Mergeduš, A.; Pšenková, M.; Janžekovič, M. Tannins and their Effect on Production Efficiency of Ruminants. Agricultura 2020, 12, 1–11. [Google Scholar] [CrossRef]
- Basini, G.; Bianco, F.; Grasselli, F. Epigallocatechin-3-gallate from green tea negatively affects swine granulosa cell function. Domest. Anim. Endocrinol. 2005, 28, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, R.; Barone, C.M.A.; Claps, S.; Varricchio, E.; Rufrano, D.; Caroprese, M.; Albenzio, M.; De Palo, P.; Campanile, G.; Neglia, G. Effects of dietary supplementation with polyphenols on meat quality in Saanen goat kids. BMC Vet. Res. 2018, 14, 181. [Google Scholar] [CrossRef]
- Balazi, A.; Sirotkin, A.V.; Foldesiova, M.; Makovicky, P.; Chrastinova, L.; Makovicky, P.; Chrenek, P. Green tea can supress rabbit ovarian functions in vitro and in vivo. Theriogenology 2019, 127, 72–79. [Google Scholar] [CrossRef]
- Denninger, T.M.; Schwarm, A.; Birkinshaw, A.; Terranova, M.; Dohme-Meier, F.; Münger, A.; Eggerschwiler, L.; Bapst, B.; Wegmann, S.; Clauss, M.; et al. Immediate effect of Acacia mearnsii tannins on methane emissions and milk fatty acid profiles of dairy cows. Anim. Feed Sci. Technol. 2020, 261, 114388. [Google Scholar] [CrossRef]
- Correddu, F.; Lunesu, M.F.; Buffa, G.; Atzori, A.S.; Nudda, A.; Battacone, G.; Pulina, G. Can Agro-Industrial By-Products Rich in Polyphenols be Advantageously Used in the Feeding and Nutrition of Dairy Small Ruminants? Animals 2020, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Cabiddu, A.; Contini, S.; Gallo, A.; Lucini, L.; Bani, P.; Decandia, M.; Molle, G.; Piluzza, G.; Sulas, L. In vitro fermentation of cardoon seed press cake-A valuable byproduct from biorefinery as a novel supplement for small ruminants. Ind. Crop. Prod. 2019, 130, 420–427. [Google Scholar] [CrossRef]
- Sobczak, P.; Zawiślak, K.; Starek, A.; Żukiewicz-Sobczak, W.; Sagan, A.; Zdybel, B.; Andrejko, D. Compaction Process as a Concept of Press-Cake Production from Organic Waste. Sustainability 2020, 12, 1567. [Google Scholar] [CrossRef] [Green Version]
- Kallela, K.; Heinonen, K.; Saloniemi, H. Plant oestrogens; the cause of decreased fertility in cows. A case report. Nord. Vet. 1984, 36, 124–129. [Google Scholar]
- Mustonen, E.; Taponen, S.; Andersson, M.; Sukura, A.; Katila, T.; Taponen, J. Fertility and growth of nulliparous ewes after feeding red clover silage with high phyto-oestrogen concentrations. Animal 2014, 8, 1699–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baheg, R.; El-Bahrawy, K.; El-Azrak, K.; Samak, M.; Sallam, S. Effect of condensed tannins and saponin supplementation on reproductive performance in Barki ewes. Egypt. J. Nutr. Feed. 2017, 20, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Hashem, N.M.; El-Azrak, K.M.; Nour El-Din, A.N.M.; Sallam, S.M.; Taha, T.A.; Salem, M.H. Effects of Trifolium alexandrinum phytoestrogens on oestrous behaviour, ovarian activity and reproductive performance of ewes during the non-breeding season. Anim. Reprod. Sci. 2018, 196, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Amir, A.A.; Kelly, J.M.; Kleemann, D.O.; Durmic, Z.; Blache, D.; Martin, G.B. Extracts of forage plants affect the developmental competence of ovine oocytes in vitro. Anim. Prod. Sci. 2019, 59, 1814. [Google Scholar] [CrossRef]
- Wang, Z.; Fu, C.; Yu, S. Effects of green tea polyphenols, insulin-like growth factor I and glucose on developmental competence of bovine oocytes. Rev. Bras. Zootec. 2012, 41, 2418–2423. [Google Scholar] [CrossRef] [Green Version]
- Piras, A.-R.; Menéndez-Blanco, I.; Soto-Heras, S.; Catalá, M.-G.; Izquierdo, D.; Bogliolo, L.; Paramio, M.-T. Resveratrol supplementation during in vitro maturation improves embryo development of prepubertal goat oocytes selected by brilliant cresyl blue staining. J. Reprod. Dev. 2019, 65, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Tava, A.; Pecio, Ł.; Lo Scalzo, R.; Stochmal, A.; Pecetti, L. Phenolic Content and Antioxidant Activity in Trifolium Germplasm from Different Environments. Molecules 2019, 24, 298. [Google Scholar] [CrossRef] [Green Version]
- Hloucalova, P.; Skladanka, J.; Horky, P.; Klejdus, B.; Pelikan, J.; Knotova, D. Determination of Phytoestrogen Content in Fresh-Cut Legume Forage. Animals 2016, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Blache, D.; Maloney, S.K.; Revell, D.K. Use and limitations of alternative feed resources to sustain and improve reproductive performance in sheep and goats. Anim. Feed Sci. Technol. 2008, 147, 140–157. [Google Scholar] [CrossRef]
- Reed, K. Fertility of Herbivores Consuming Phytoestrogen-containing Medicago and Trifolium Species. Agriculture 2016, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Herchi, W.; Arráez-Román, D.; Trabelsi, H.; Bouali, I.; Boukhchina, S.; Kallel, H.; Segura-Carretero, A.; Fernández-Gutierrez, A. Phenolic Compounds in Flaxseed: A Review of Their Properties and Analytical Methods. An Overview of the Last Decade. J. Oleo Sci. 2014, 63, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolodziejczyk-Czepas, J.; Sieradzka, M.; Moniuszko-Szajwaj, B.; Nowak, P.; Oleszek, W.; Stochmal, A. Phenolic fractions from nine Trifolium species modulate the coagulant properties of blood plasma in vitro without cytotoxicity towards blood cells. J. Pharm. Pharm. 2018, 70, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.K.; Rana, Z.H.; Islam, S.N.; Akhtaruzzaman, M. Comparative assessment of nutritional composition, polyphenol profile, antidiabetic and antioxidative properties of selected edible wild plant species of Bangladesh. Food Chem. 2020, 320, 126646. [Google Scholar] [CrossRef]
- Mekoya, A.; Oosting, S.J.; Fernandez-Rivera, S.; Tamminga, S.; Tegegne, A.; Van der Zijpp, A.J. Effect of supplementation of Sesbania sesban on reproductive performance of sheep. Livest. Sci. 2009, 121, 117–125. [Google Scholar] [CrossRef]
- Shokry, D.M.; Badr, M.R.; Orabi, S.H.; Khalifa, H.K.; El-Seedi, H.R.; Abd Eldaim, M.A. Moringa oleifera leaves extract enhances fresh and cryopreserved semen characters of Barki rams. Theriogenology 2020, 153, 133–142. [Google Scholar] [CrossRef]
- Leone, A.; Spada, A.; Battezzati, A.; Schiraldi, A.; Aristil, J.; Bertoli, S. Cultivation, Genetic, Ethnopharmacology, Phytochemistry and Pharmacology of Moringa oleifera Leaves: An Overview. Int. J. Mol. Sci. 2015, 16, 12791–12835. [Google Scholar] [CrossRef]
- Blaise, S.; Cartier, D.; Reynaud, J.; JAY, M. Evolution and differentiation of Lotus corniculatus/Lotus alpinus populations from French south-western Alps. I: Morphologic and cyanogenic variations. II: Contribution of phenolic metabolism markers. III: Conclusions. Evol. Trends Plants 1991, 5, 137–148. [Google Scholar]
- Jancic, D.; Todorovic, V.; Sircelj, H.; Dodevska, M.; Beljkas, B.; Znidarcic, D.; Sobajic, S. Biologically active compounds and antioxidant capacity of Cichorium intybus L. leaves from Montenegro. Ital. J. Food Sci. 2017, 29. [Google Scholar] [CrossRef]
- Nyman, T.; Julkunen-Tiitto, R. Manipulation of the phenolic chemistry of willows by gall-inducing sawflies. Proc. Natl. Acad. Sci. USA 2000, 97, 13184–13187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengistu, G.; Karonen, M.; Salminen, J.P.; Hendriks, W.H.; Pellikaan, W.F. In vitro fermentation of browse species using goat rumen fluid in relation to browse polyphenol content and composition. Anim. Feed Sci. Technol. 2017, 231, 1–11. [Google Scholar] [CrossRef]
- Alañón, M.E.; Castro-Vázquez, L.; Díaz-Maroto, M.C.; Gordon, M.H.; Pérez-Coello, M.S. A study of the antioxidant capacity of oak wood used in wine ageing and the correlation with polyphenol composition. Food Chem. 2011, 128, 997–1002. [Google Scholar] [CrossRef] [Green Version]
- Nudda, A.; Buffa, G.; Atzori, A.; Cappai, M.; Caboni, P.; Fais, G.; Pulina, G. Small amounts of agro-industrial byproducts in dairy ewes diets affects milk production traits and hematological parameters. Anim. Feed Sci. Technol. 2019, 251, 76–85. [Google Scholar] [CrossRef]
- Pan, Z.; Zhang, R.; Zicari, S. Integrated Processing Technologies for Food and Agricultural By-Products; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Leouifoudi, I.; Harnafi, H.; Zyad, A. Olive mill waste extracts: Polyphenols content, antioxidant, and antimicrobial activities. Adv. Pharmacol. Sci. 2015, 2015. [Google Scholar] [CrossRef]
- Ozturk, B.; Parkinson, C.; Gonzalez-Miquel, M. Extraction of polyphenolic antioxidants from orange peel waste using deep eutectic solvents. Sep. Purif. Technol. 2018, 206, 1–13. [Google Scholar] [CrossRef]
- Elfalleh, W.; Hannachi, H.; Tlili, N.; Yahia, Y.; Nasri, N.; Ferchichi, A. Total phenolic contents and antioxidant activities of pomegranate peel, seed, leaf and flower. J. Med. Plants Res. 2012, 6, 4724–4730. [Google Scholar] [CrossRef]
- Gullon, B.; Pintado, M.E.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Assessment of polyphenolic profile and antibacterial activity of pomegranate peel (Punica granatum) flour obtained from co-product of juice extraction. Food Control 2016, 59, 94–98. [Google Scholar] [CrossRef]
- Razzaghi, A.; Naserian, A.A.; Valizadeh, R.; Ebrahimi, S.H.; Khorrami, B.; Malekkhahi, M.; Khiaosa-ard, R. Pomegranate seed pulp, pistachio hulls, and tomato pomace as replacement of wheat bran increased milk conjugated linoleic acid concentrations without adverse effects on ruminal fermentation and performance of Saanen dairy goats. Anim. Feed Sci. Technol. 2015, 210, 46–55. [Google Scholar] [CrossRef]
- Zhao, C.N.; Tang, G.Y.; Cao, S.Y.; Xu, X.Y.; Gan, R.Y.; Liu, Q.; Mao, Q.Q.; Shang, A.; Li, H.B. Phenolic Profiles and Antioxidant Activities of 30 Tea Infusions from Green, Black, Oolong, White, Yellow and Dark Teas. Antioxidants 2019, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Platat, C.; Habib, H.; Al Maqbali, F.; Jaber, N.; Ibrahim, W. Identification of date seeds varieties patterns to optimize nutritional benefits of date seeds. J. Nutr. Food Sci. S 2014, 8, 2. [Google Scholar] [CrossRef]
- Oses, S.M.; Marcos, P.; Azofra, P.; de Pablo, A.; Fernandez-Muino, M.A.; Sancho, M.T. Phenolic Profile, Antioxidant Capacities and Enzymatic Inhibitory Activities of Propolis from Different Geographical Areas: Needs for Analytical Harmonization. Antioxidants 2020, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatab, A.; Hashem, N.; El-Kodary, L.; Lotfy, F.; Hassan, G. Evaluation of the effects of cypermethrin on female reproductive function by using rabbit model and of the protective role of Chinese propolis. Biomed. Environ. Sci. 2016, 29, 762–766. [Google Scholar] [PubMed]
- Hashem, N.M.; Abo-Elsoud, M.A.; Nour El-Din, A.N.M.; Kamel, K.I.; Hassan, G.A. Prolonged exposure of dietary phytoestrogens on semen characteristics and reproductive performance of rabbit bucks. Domest. Anim. Endocrinol. 2018, 64, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Abo-Elsoud, M.A.; Hashem, N.M.; Nour El-Din, A.N.M.; Kamel, K.I.; Hassan, G.A. Soybean isoflavone affects in rabbits: Effects on metabolism, antioxidant capacity, hormonal balance and reproductive performance. Anim. Reprod. Sci. 2019, 203, 52–60. [Google Scholar] [CrossRef]
- Mueller-Harvey, I.; Bee, G.; Dohme-Meier, F.; Hoste, H.; Karonen, M.; Kölliker, R.; Lüscher, A.; Niderkorn, V.; Pellikaan, W.F.; Salminen, J.-P.; et al. Benefits of Condensed Tannins in Forage Legumes Fed to Ruminants: Importance of Structure, Concentration, and Diet Composition. Crop. Sci. 2019, 59, 861–885. [Google Scholar] [CrossRef] [Green Version]
- Gloria, A.; Contri, A.; Grotta, L.; Carluccio, A.; Robbe, D.; Ianni, A.; Vignola, G.; Martino, G. Effect of dietary grape marc on fresh and refrigerated boar semen. Anim. Reprod. Sci. 2019, 205, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Idowu, O.O. Green tea extract and reproduction: A review. E3 J. Med. Res. 2017, 6, 001–006. [Google Scholar] [CrossRef] [Green Version]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Ly, C.; Yockell-Lelievre, J.; Ferraro, Z.M.; Arnason, J.T.; Ferrier, J.; Gruslin, A. The effects of dietary polyphenols on reproductive health and early development. Hum. Reprod. Update 2015, 21, 228–248. [Google Scholar] [CrossRef] [Green Version]
- Sieniawska, E. Activities of tannins–From in vitro studies to clinical trials. Nat. Prod. Commun. 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef]
- Woclawek-Potocka, I.; Mannelli, C.; Boruszewska, D.; Kowalczyk-Zieba, I.; Wasniewski, T.; Skarzynski, D.J. Diverse effects of phytoestrogens on the reproductive performance: Cow as a model. Int. J. Endocrinol. 2013, 2013, 650984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milojevic, V.; Sinz, S.; Kreuzer, M.; Chiumia, D.; Marquardt, S.; Giller, K. Partitioning of fatty acids into tissues and fluids from reproductive organs of ewes as affected by dietary phenolic extracts. Theriogenology 2020, 144, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.O.; Wang, C.C.; Chu, C.Y.; Chan, K.P.; Rogers, M.S.; Choy, K.W.; Pang, C.P. Pharmacokinetic studies of green tea catechins in maternal plasma and fetuses in rats. J. Pharm. Sci. 2006, 95, 1372–1381. [Google Scholar] [CrossRef] [PubMed]
- Yousif, A.N. Effect of Flaxseed on some hormonal profile and genomic DNA concentration in Karadi lambs. IOP Conf. Ser. Earth Environ. Sci. 2019, 388, 012035. [Google Scholar] [CrossRef]
- Hashem, N.M.; Sallam, S.M.A. Sexual and ovarian activity of crossbred ewes fed different types of roughage during seasonal anestrus. Small Rumin. Res. 2012, 107, 136–140. [Google Scholar] [CrossRef]
- Hashem, N.M.; El-Azrak, K.M.; Sallam, S.M. Hormonal concentrations and reproductive performance of Holstein heifers fed Trifolium alexandrinum as a phytoestrogenic roughage. Anim. Reprod. Sci. 2016, 170, 121–127. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; van der Saag, P.T.; van der Burg, B.; Gustafsson, J.A. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef]
- Yildiz, H.B.; Kiralp, S.; Toppare, L.; Yagci, Y. Immobilization of tyrosinase in poly (ethyleneoxide) electrodes and determination of phenolics in red wines. React. Funct. Polym. 2005, 63, 155–161. [Google Scholar] [CrossRef]
- Lorand, T.; Vigh, E.; Garai, J. Hormonal action of plant derived and anthropogenic non-steroidal estrogenic compounds: Phytoestrogens and xenoestrogens. Curr. Med. Chem. 2010, 17, 3542–3574. [Google Scholar] [CrossRef] [PubMed]
- Cipolletti, M.; Solar Fernandez, V.; Montalesi, E.; Marino, M.; Fiocchetti, M. Beyond the antioxidant activity of dietary polyphenols in cancer: The modulation of estrogen receptors (ers) signaling. Int. J. Mol. Sci. 2018, 19, 2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinaci, M.; Volpe, S.; De Ambrogi, M.; Tamanini, C.; Galeati, G. Effects of epigallocatechin-3-gallate (EGCG) on in vitro maturation and fertilization of porcine oocytes. Theriogenology 2008, 69, 877–885. [Google Scholar] [CrossRef]
- Macias-Cruz, U.; Vicente-Perez, R.; Lopez-Baca, M.A.; Gonzalez-Rios, H.; Correa-Calderon, A.; Arechiga, C.F.; Avendano-Reyes, L. Effects of dietary ferulic acid on reproductive function and metabolism of pre-pubertal hairbreed ewes during the anestrous season. Theriogenology 2018, 119, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Zarepourfard, H.; Riasi, A.; Frouzanfar, M.; Hajian, M.; Nasr Esfahani, M.H. Pomegranate seed in diet, affects sperm parameters of cloned goats following freezing-thawing. Theriogenology 2019, 125, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.A.; Hamed, R.S.; Bovera, F.; Abd El, A.E.-H.E.; Al-Harthi, M.A.; Shahba, H.A. Semen quality, antioxidant status and reproductive performance of rabbits bucks fed milk thistle seeds and rosemary leaves. Anim. Reprod. Sci. 2017, 184, 178–186. [Google Scholar] [CrossRef]
- El-Desoky, N.I.; Hashem, N.M.; Elkomy, A.; Abo-Elezz, Z.R. Physiological response and semen quality of rabbit bucks supplemented with Moringa leaves ethanolic extract during summer season. Animal 2017, 11, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Jin, Y.; Du, M.; Liu, W.; Ren, Y.; Zhang, C.; Zhang, J. The effect of dietary grape pomace supplementation on epididymal sperm quality and testicular antioxidant ability in ram lambs. Theriogenology 2017, 97, 50–56. [Google Scholar] [CrossRef]
- Kaya, M.; Cenesiz, M.; Onder, F.; Ucar, O.; Uzun, M.; Yildiz, S. GnRH-Induced LH Secretion in Prepubertal female fat-tailed Lambs fed with tannin-rich Oak Leaves (Quercus hartwissiana). Rev. Méd. Vét. 2006, 157, 387. [Google Scholar]
- Sallam, S.M.; El-Gendy, M.N.; Anwar, M.M.; Fahmy, W.G.; El-Zarkouny, S.Z.; Hashem, N.M.; El-Din, A.N.N.; Attia, M.F.; El-wakeel, E.-S.A.; Zeitoun, M.M. Productive and reproductive traits of sheep fed Acacia saligna Leaves-based diets. Int. J. Environ. Agric. Biotechnol. 2017, 2, 238973. [Google Scholar]
- Ramírez-Restrepo, C.A.; Barry, T.N.; López-Villalobos, N.; Kemp, P.D.; Harvey, T.G. Use of Lotus corniculatus containing condensed tannins to increase reproductive efficiency in ewes under commercial dryland farming conditions. Anim. Feed Sci. Technol. 2005, 121, 23–43. [Google Scholar] [CrossRef]
- Földešiová, M.; Baláži, A.; Chrastinová, Ľ.; Pivko, J.; Kotwica, J.; Harrath, A.H.; Chrenek, P.; Sirotkin, A.V. Yucca schidigera can promote rabbit growth, fecundity, affect the release of hormones in vivo and in vitro, induce pathological changes in liver, and reduce ovarian resistance to benzene. Anim. Reprod. Sci. 2017, 183, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Sallam, S.M.A.; Attia, M.F.A.; Nour El-Din, A.N.M.; El-Zarkouny, S.Z.; Saber, A.M.; El-Zaiat, H.M.; Zeitoun, M.M. Involvement of Quebracho tannins in diet alters productive and reproductive efficiency of postpartum buffalo cows. Anim. Nutr. 2019, 5, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Nawab, A.; Tang, S.; Gao, W.; Li, G.; Xiao, M.; An, L.; Wu, J.; Liu, W. Tannin Supplementation in Animal Feeding; Mitigation Strategies to Overcome the Toxic Effects of Tannins on Animal Health: A Review. J. Agric. Sci. 2020, 12, 217. [Google Scholar] [CrossRef] [Green Version]
- Hashem, N.M.; El-Hady, A.A.; Hassan, O. Effect of vitamin E or propolis supplementation on semen quality, oxidative status and hemato-biochemical changes of rabbit bucks during hot season. Livest. Sci. 2013, 157, 520–526. [Google Scholar] [CrossRef]
- Melaku, S.; Peters, K.J.; Tegegne, A. Supplementation of Menz ewes with dried leaves of Lablab purpureus or graded levels of Leucaena pallida 14203 and Sesbania sesban 1198: Effects on feed intake, live weight gain and estrous cycle. Anim. Feed Sci. Technol. 2004, 113, 39–51. [Google Scholar] [CrossRef]
- Roselli, C.E.; Stormshak, F. The ovine sexually dimorphic nucleus, aromatase, and sexual partner preferences in sheep. J. Steroid Biochem. Mol. Biol. 2010, 118, 252–256. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska, K.K.; Woclawek-Potocka, I.; Bah, M.M.; Piskula, M.K.; Pilawski, W.; Bober, A.; Skarzynski, D.J. Phytoestrogens and their metabolites inhibit the sensitivity of the bovine corpus luteum to luteotropic factors. J. Reprod. Dev. 2005, 0511010030. [Google Scholar] [CrossRef] [Green Version]
- Cools, S.; Van den Broeck, W.; Vanhaecke, L.; Heyerick, A.; Bossaert, P.; Hostens, M.; Opsomer, G. Feeding soybean meal increases the blood level of isoflavones and reduces the steroidogenic capacity in bovine corpora lutea, without affecting peripheral progesterone concentrations. Anim. Reprod. Sci. 2014, 144, 79–89. [Google Scholar] [CrossRef]
- Watzková, J.; Křížová, L.; Pavlík, A.; Schulzová, V.; Hajšlová, J.; Lojza, J. The Effect of Soybean-Derived Phytoestrogens on Concentrations of Plasma Isoflavones, 15-keto-13, 14-dihydroprostaglandin F 2α and Progesterone in Dairy Cows. Acta Vet. Brno 2011, 79, 525–532. [Google Scholar] [CrossRef]
- Kaplanski, O.; Shemesh, M.; Berman, A. Effects of phyto-oestrogens on progesterone synthesis by isolated bovine granulosa cells. J. Endocrinol. 1981, 89, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ando, R.; Shimizu, K.; Hashida, K.; Makino, R.; Ohara, S.; Kondo, R. Steroid 5α-reductase inhibitory activity of condensed tannins from woody plants. J. Wood Sci. 2008, 54, 68–75. [Google Scholar] [CrossRef]
- Lephart, E.D. Modulation of Aromatase by Phytoestrogens. Enzym. Res. 2015, 2015, 594656. [Google Scholar] [CrossRef] [Green Version]
- Basini, G.; Bussolati, S.; Baioni, L.; Grasselli, F. Gossypol, a polyphenolic aldehyde from cotton plant, interferes with swine granulosa cell function. Domest. Anim. Endocrinol. 2009, 37, 30–36. [Google Scholar] [CrossRef]
- Colitti, M.; Sgorlon, S.; Stradaioli, G.; Farinacci, M.; Gabai, G.; Stefanon, B. Grape polyphenols affect mRNA expression of PGHS-2, TIS11b and FOXO3 in endometrium of heifers under ACTH-induced stress. Theriogenology 2007, 68, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Swelum, A.A.-A.; Hashem, N.M.; Abo-Ahmed, A.I.; Abd El-Hack, M.E.; Abdo, M. The Role of Heat Shock Proteins in Reproductive Functions. In Heat Shock Proteins; Springer: Dordrecht, The Netherlands, 2020. [Google Scholar]
- Evans, B.A.J.; Griffiths, K.; Morton, M. Inhibition of 5α-reductase in genital skin fibroblasts and prostate tissue by dietary lignans and isoflavonoids. J. Endocrinol. 1995, 147, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Hadadi, M.; Mohammadi, G.; Erfani, N.; Fatemi, R. Effects Of Short And Long-Term Administration Of Alfalfa On Testicular Histomorphometry In Rats. J. Pharm. Res. Int. 2020, 10–17. [Google Scholar] [CrossRef]
- Retana- Márquez, S.; Muñoz-Gutiérrez, M.; Duarte, G.; Vielma, J.S.; Fitz-Rodríguez, G.; Keller, M. Effects of phytoestrogens on mammalian reproductive physiology. Trop. Subtrop. Agroecosyst. 2011, 15, S129–S145. [Google Scholar]
- Perez-Martinez, C.; Ferreras-Estrada, M.C.; Garcia-Iglesias, M.J.; Bravo-Moral, A.M.; Espinosa-Alvarez, J.; Escudero-Diez, A. Effects of in utero exposure to nonsteroidal estrogens on mouse testis. Can. J. Vet. Res. 1997, 61, 94. [Google Scholar] [PubMed]
- Cederroth, C.R.; Auger, J.; Zimmermann, C.; Eustache, F.; Nef, S. Soy, phyto-oestrogens and male reproductive function: A review. Int. J. Androl. 2010, 33, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Eustache, F.; Mondon, F.; Canivenc-Lavier, M.C.; Lesaffre, C.; Fulla, Y.; Berges, R.; Cravedi, J.P.; Vaiman, D.; Auger, J. Chronic Dietary Exposure to a Low-Dose Mixture of Genistein and Vinclozolin Modifies the Reproductive Axis, Testis Transcriptome, and Fertility. Environ. Health Perspect. 2009, 117, 1272–1279. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, W.N.; Patisaul, H.B.; Williams, C.J. Reproductive consequences of developmental phytoestrogen exposure. Reproduction 2012, 143, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, W.N.; Williams, C.J. Circulating levels of genistein in the neonate, apart from dose and route, predict future adverse female reproductive outcomes. Reprod. Toxicol. 2011, 31, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wang, A.; Liu, X.; Li, E. Effects of resveratrol on reducing spermatogenic dysfunction caused by high-intensity exercise. Reprod. Biol. Endocrinol. 2019, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira Braga, C.; Momentti, A.C.; Barbosa Peixoto, F.; de Fátima Ferreira Baptista, R.; dos Santos, F.A.; Fava, F.H.; Fernandes, A.A.H. Influence of treatment with quercetin on lipid parameters and oxidative stress of pregnant diabetic rats. Can. J. Physiol. Pharmacol. 2013, 91, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhao, J.; Xu, H.; Lyv, Y.; Feng, X.; Fang, Y.; Xu, Y. Maternal quercetin administration during gestation and lactation decrease endoplasmic reticulum stress and related inflammation in the adult offspring of obese female rats. Eur. J. Nutr. 2014, 53, 1669–1683. [Google Scholar] [CrossRef] [PubMed]
- Prater, M.R.; Laudermilch, C.L.; Liang, C.; Holladay, S.D. Placental Oxidative Stress Alters Expression of Murine Osteogenic Genes and Impairs Fetal Skeletal Formation. Placenta 2008, 29, 802–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, C.; Oest, M.E.; Prater, M.R. Intrauterine exposure to high saturated fat diet elevates risk of adult-onset chronic diseases in C57BL/6 mice. Birth Defects Res. Part B Dev. Reprod. Toxicol. 2009, 86, 377–384. [Google Scholar] [CrossRef]
- Santangelo, C.; Zicari, A.; Mandosi, E.; Scazzocchio, B.; Mari, E.; Morano, S.; Masella, R. Could gestational diabetes mellitus be managed through dietary bioactive compounds? Current knowledge and future perspectives. Br. J. Nutr. 2016, 115, 1129–1144. [Google Scholar] [CrossRef]
- Vanhees, K.; van Schooten, F.J.; van Waalwijk van Doorn-Khosrovani, S.B.; van Helden, S.; Munnia, A.; Peluso, M.; Briedé, J.J.; Haenen, G.R.M.M.; Godschalk, R.W.L. Intrauterine exposure to flavonoids modifies antioxidant status at adulthood and decreases oxidative stress-induced DNA damage. Free Radic. Biol. Med. 2013, 57, 154–161. [Google Scholar] [CrossRef]
- Cao, L.; Tan, C.; Meng, F.; Liu, P.; Reece, E.A.; Zhao, Z. Amelioration of intracellular stress and reduction of neural tube defects in embryos of diabetic mice by phytochemical quercetin. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Bonacasa, B.; Siow, R.C.M.; Mann, G.E. Impact of Dietary Soy Isoflavones in Pregnancy on Fetal Programming of Endothelial Function in Offspring. Microcirculation 2011, 18, 270–285. [Google Scholar] [CrossRef]
- Do, G.-M.; Jung, U.J.; Park, H.-J.; Kwon, E.-Y.; Jeon, S.-M.; McGregor, R.A.; Choi, M.-S. Resveratrol ameliorates diabetes-related metabolic changes via activation of AMP-activated protein kinase and its downstream targets in db/db mice. Mol. Nutr. Food Res. 2012, 56, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Smoliga, J.M.; Baur, J.A.; Hausenblas, H.A. Resveratrol and health—A comprehensive review of human clinical trials. Mol. Nutr. Food Res. 2011, 55, 1129–1141. [Google Scholar] [CrossRef]
- Singh, C.K.; Kumar, A.; LaVoie, H.A.; DiPette, D.J.; Singh, U.S. Diabetic complications in pregnancy: Is resveratrol a solution? Exp. Biol. Med. 2013, 238, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, C.K.; Kumar, A.; Hitchcock, D.B.; Fan, D.; Goodwin, R.; LaVoie, H.A.; Nagarkatti, P.; DiPette, D.J.; Singh, U.S. Resveratrol prevents embryonic oxidative stress and apoptosis associated with diabetic embryopathy and improves glucose and lipid profile of diabetic dam. Mol. Nutr. Food Res. 2011, 55, 1186–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.R.; Pires, K.M.P.; Nalbones-Barbosa, M.N.; dos Santos Valença, S.; Resende, Â.C.; de Moura, R.S. Grape skin extract-derived polyphenols modify programming-induced renal endowment in prenatal protein-restricted male mouse offspring. Eur. J. Nutr. 2015, 55, 1455–1464. [Google Scholar] [CrossRef]
- Bourque, S.L.; Dolinsky, V.W.; Dyck, J.R.B.; Davidge, S.T. Maternal resveratrol treatment during pregnancy improves adverse fetal outcomes in a rat model of severe hypoxia. Placenta 2012, 33, 449–452. [Google Scholar] [CrossRef]
- Doerge, D.R. Bioavailability of soy isoflavones through placental/lactational transfer and soy food. Toxicol. Appl. Pharmacol. 2011, 254, 145–147. [Google Scholar] [CrossRef]
- Shahzad, H.; Giribabu, N.; Karim, K.; Kassim, N.; Muniandy, S.; Kumar, K.E.; Salleh, N. Quercetin interferes with the fluid volume and receptivity development of the uterus in rats during the peri-implantation period. Reprod. Toxicol. 2017, 71, 42–54. [Google Scholar] [CrossRef]
- Johnson, J.R.; Makaji, E.; Ho, S.; Boya, X.; Crankshaw, D.J.; Holloway, A.C. Effect of Maternal Raspberry Leaf Consumption in Rats on Pregnancy Outcome and the Fertility of the Female Offspring. Reprod. Sci. 2009, 16, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Bulotta, S.; Celano, M.; Lepore, S.M.; Montalcini, T.; Pujia, A.; Russo, D. Beneficial effects of the olive oil phenolic components oleuropein and hydroxytyrosol: Focus on protection against cardiovascular and metabolic diseases. J. Transl. Med. 2014, 12, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chango, A.; Pogribny, I. Considering Maternal Dietary Modulators for Epigenetic Regulation and Programming of the Fetal Epigenome. Nutrients 2015, 7, 2748–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhees, K.; Coort, S.; Ruijters, E.J.B.; Godschalk, R.W.L.; Schooten, F.J.; Doorn-Khosrovani, S.B.v.W. Epigenetics: Prenatal exposure to genistein leaves a permanent signature on the hematopoietic lineage. FASEB J. 2010, 25, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.-M.; Cheong, A.; Adgent, M.A.; Veevers, J.; Suen, A.A.; Tam, N.N.C.; Leung, Y.-K.; Jefferson, W.N.; Williams, C.J. Environmental factors, epigenetics, and developmental origin of reproductive disorders. Reprod. Toxicol. 2017, 68, 85–104. [Google Scholar] [CrossRef] [Green Version]
- Vanhees, K.; van Schooten, F.J.; Moonen, E.J.; Maas, L.M.; Barjesteh van, S.; Godschalk, R.W.L. Maternal intake of quercetin during gestation alters ex vivo benzo[a]pyrene metabolism and DNA adduct formation in adult offspring. Mutagenesis 2012, 27, 445–451. [Google Scholar] [CrossRef]
- Lanoue, L.; Green, K.K.; Kwik-Uribe, C.; Keen, C.L. Dietary factors and the risk for acute infant leukemia: Evaluating the effects of cocoa-derived flavanols on DNA topoisomerase activity. Exp. Biol. Med. 2010, 235, 77–89. [Google Scholar] [CrossRef]
- Vanhees, K.; de Bock, L.; Godschalk, R.W.L.; van Schooten, F.J.; van Waalwijk van Doorn-Khosrovani, S.B. Prenatal Exposure to Flavonoids: Implication for Cancer Risk. Toxicol. Sci. 2011, 120, 59–67. [Google Scholar] [CrossRef]
- Puangpraphant, S.; De Mejia, E.G. Saponins in yerba mate tea (Ilex paraguariensis A. St.-Hil) and quercetin synergistically inhibit iNOS and COX-2 in lipopolysaccharide-induced macrophages through NFκB pathways. J. Agric. Food Chem. 2009, 57, 8873–8883. [Google Scholar] [CrossRef]
- Majed, B.H.; Khalil, R.A.; Garland, C.J. Molecular Mechanisms Regulating the Vascular Prostacyclin Pathways and Their Adaptation during Pregnancy and in the Newborn. Pharmacol. Rev. 2012, 64, 540–582. [Google Scholar] [CrossRef] [Green Version]
- Tanir, H.M.; Sener, T.; Inal, M.; Akyuz, F.; Uzuner, K.; Sivri, E. Effect of quercetine and glutathione on the level of superoxide dismutase, catalase, malonyldialdehyde, blood pressure and neonatal outcome in a rat model of pre-eclampsia induced by NG-nitro-l-arginine-methyl ester. Eur. J. Obstet. Gynecol. Reprod. Biol. 2005, 118, 190–195. [Google Scholar] [CrossRef]
- Zielinsky, P.; Piccoli, A.L.; Manica, J.L.L.; Nicoloso, L.H.S. New insights on fetal ductal constriction: Role of maternal ingestion of polyphenol-rich foods. Expert Rev. Cardiovasc. Ther. 2010, 8, 291–298. [Google Scholar] [CrossRef]
- Zielinsky, P.; Busato, S. Prenatal effects of maternal consumption of polyphenol-rich foods in late pregnancy upon fetal ductus arteriosus. Birth Defects Res. Part C Embryo Today Rev. 2013, 99, 256–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, M.; Baierle, M.; Charão, M.F.; Bubols, G.B.; Gravina, F.S.; Zielinsky, P.; Arbo, M.D.; Cristina Garcia, S. Polyphenol-rich food general and on pregnancy effects: A review. Drug Chem. Toxicol. 2017, 40, 368–374. [Google Scholar] [CrossRef]
- Vanhees, K.; Godschalk, R.W.; Sanders, A.; van Waalwijk van Doorn-Khosrovani, S.B.; van Schooten, F.J. Maternal quercetin intake during pregnancy results in an adapted iron homeostasis at adulthood. Toxicology 2011, 290, 350–358. [Google Scholar] [CrossRef]
- Biondi, C.; Pavan, B.; Dalpiaz, A.; Medici, S.; Lunghi, L.; Vesce, F. Expression and characterization of vitamin C transporter in the human trophoblast cell line HTR-8/SVneo: Effect of steroids, flavonoids and NSAIDs. MHR Basic Sci. Reprod. Med. 2007, 13, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Martel, F.; Monteiro, R.; Calhau, C. Effect of polyphenols on the intestinal and placental transport of some bioactive compounds. Nutr. Res. Rev. 2010, 23, 47–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, H.; Jahan, S.; Ullah, H.; Ullah, F.; Salman, M.M. The addition of resveratrol in tris citric acid extender ameliorates post-thaw quality parameters, antioxidant enzymes levels, and fertilizing capability of buffalo (Bubalus bubalis) bull spermatozoa. Theriogenology 2020, 152, 106–113. [Google Scholar] [CrossRef]
- Al-Mutary, M.G.; Al-Ghadi, M.Q.; Ammari, A.A.; Al-Himadi, A.R.; Al-Jolimeed, A.H.; Arafah, M.W.; Amran, R.A.; Aleissa, M.S.; Swelum, A.A. Effect of different concentrations of resveratrol on the quality and in vitro fertilizing ability of ram semen stored at 5 degrees C for up to 168 h. Theriogenology 2020, 152, 139–146. [Google Scholar] [CrossRef]
- Arando, A.; Delgado, J.; Fernández-Prior, A.; León, J.; Bermúdez-Oria, A.; Nogales, S.; Pérez-Marín, C. Effect of different olive oil-derived antioxidants (hydroxytyrosol and 3, 4-dihydroxyphenylglycol) on the quality of frozen-thawed ram sperm. Cryobiology 2019, 86, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.; Li, Y.; Feng, T.; Du, Y.; Ren, F.; Zhang, L.; Han, N.; Ma, S.; Li, F.; Wang, P. Grape Seed Procyanidin Extract (GSPE) Improves Goat Sperm Quality When Preserved at 4 °C. Animals 2019, 9, 810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, C.; Larbi, A.; Wu, G.; Hong, Q.; Quan, G. Improving the quality of cryopreserved goat semen with a commercial bull extender supplemented with resveratrol. Anim. Reprod. Sci. 2019, 208, 106127. [Google Scholar] [CrossRef] [PubMed]
- Spinaci, M.; Muccilli, V.; Bucci, D.; Cardullo, N.; Gadani, B.; Tringali, C.; Tamanini, C.; Galeati, G. Biological effects of polyphenol-rich extract and fractions from an oenological oak-derived tannin on in vitro swine sperm capacitation and fertilizing ability. Theriogenology 2018, 108, 284–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longobardi, V.; Zullo, G.; Salzano, A.; De Canditiis, C.; Cammarano, A.; De Luise, L.; Puzio, M.V.; Neglia, G.; Gasparrini, B. Resveratrol prevents capacitation-like changes and improves in vitro fertilizing capability of buffalo frozen-thawed sperm. Theriogenology 2017, 88, 1–8. [Google Scholar] [CrossRef]
- Gadani, B.; Bucci, D.; Spinaci, M.; Tamanini, C.; Galeati, G. Resveratrol and Epigallocatechin-3-gallate addition to thawed boar sperm improves in vitro fertilization. Theriogenology 2017, 90, 88–93. [Google Scholar] [CrossRef]
- Menzel, V.A.; Hinsch, E.; Hagele, W.; Hinsch, K.D. Effect of genistein on acrosome reaction and zona pellucida binding independent of protein tyrosine kinase inhibition in bull. Asian J. Androl. 2007, 9, 650–658. [Google Scholar] [CrossRef]
- Spinaci, M.; Bucci, D.; Muccilli, V.; Cardullo, N.; Nerozzi, C.; Galeati, G. A polyphenol-rich extract from an oenological oak-derived tannin influences in vitro maturation of porcine oocytes. Theriogenology 2019, 129, 82–89. [Google Scholar] [CrossRef]
- Zabihi, A.; Shabankareh, H.K.; Hajarian, H.; Foroutanifar, S. Resveratrol addition to in vitro maturation and in vitro culture media enhances developmental competence of sheep embryos. Domest. Anim. Endocrinol. 2019, 68, 25–31. [Google Scholar] [CrossRef]
- Amir, A.A.; Kelly, J.M.; Kleemann, D.O.; Durmic, Z.; Blache, D.; Martin, G.B. Phyto-oestrogens affect fertilisation and embryo development in vitro in sheep. Reprod. Fertil. Dev. 2018, 30, 1109. [Google Scholar] [CrossRef]
- Kuhnle, G.G.; Dell’Aquila, C.; Aspinall, S.M.; Runswick, S.A.; Mulligan, A.A.; Bingham, S.A. Phytoestrogen content of foods of animal origin: Dairy products, eggs, meat, fish, and seafood. J. Agric. Food Chem. 2008, 56, 10099–10104. [Google Scholar] [CrossRef]
- Mustonen, E.A.; Tuori, M.; Saastamoinen, I.; Taponen, J.; Wahala, K.; Saloniemi, H.; Vanhatalo, A. Equol in milk of dairy cows is derived from forage legumes such as red clover. Br. J. Nutr. 2009, 102, 1552–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blahova, L.; Kohoutek, J.; Prochazkova, T.; Prudikova, M.; Blaha, L. Phytoestrogens in milk: Overestimations caused by contamination of the hydrolytic enzyme used during sample extraction. J. Dairy Sci. 2016, 99, 6973–6982. [Google Scholar] [CrossRef] [PubMed]
- Tsen, S.Y.; Siew, J.; Lau, E.K.L.; Afiqah bte Roslee, F.; Chan, H.M.; Loke, W.M. Cow’s milk as a dietary source of equol and phenolic antioxidants: Differential distribution in the milk aqueous and lipid fractions. Dairy Sci. Technol. 2014, 94, 625–632. [Google Scholar] [CrossRef]
- Di Trana, A.; Bonanno, A.; Cecchini, S.; Giorgio, D.; Di Grigoli, A.; Claps, S. Effects of Sulla forage (Sulla coronarium L.) on the oxidative status and milk polyphenol content in goats. J. Dairy Sci. 2015, 98, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chávez-Servín, J.L.; Andrade-Montemayor, H.M.; Velázquez Vázquez, C.; Aguilera Barreyro, A.; García-Gasca, T.; Ferríz Martínez, R.A.; Olvera Ramírez, A.M.; de la Torre-Carbot, K. Effects of feeding system, heat treatment and season on phenolic compounds and antioxidant capacity in goat milk, whey and cheese. Small Rumin. Res. 2018, 160, 54–58. [Google Scholar] [CrossRef]
- Antignac, J.P.; Cariou, R.; Le Bizec, B.; Cravedi, J.P.; Andre, F. Identification of phytoestrogens in bovine milk using liquid chromatography/electrospray tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Steinshamn, H.; Purup, S.; Thuen, E.; Hansen-Møller, J. Effects of clover-grass silages and concentrate supplementation on the content of phytoestrogens in dairy cow milk. J. Dairy Sci. 2008, 91, 2715–2725. [Google Scholar] [CrossRef]
- Andersen, C.; Nielsen, T.S.; Purup, S.; Kristensen, T.; Eriksen, J.; Soegaard, K.; Sorensen, J.; Frette, X.C. Phyto-oestrogens in herbage and milk from cows grazing white clover, red clover, lucerne or chicory-rich pastures. Animal 2009, 3, 1189–1195. [Google Scholar] [CrossRef] [Green Version]
- Setchell, K.D.; Clerici, C.; Lephart, E.D.; Cole, S.J.; Heenan, C.; Castellani, D.; Wolfe, B.E.; Nechemias-Zimmer, L.; Brown, N.M.; Lund, T.D. S-equol, a potent ligand for estrogen receptor β, is the exclusive enantiomeric form of the soy isoflavone metabolite produced by human intestinal bacterial flora. Am. J. Clin. Nutr. 2005, 81, 1072–1079. [Google Scholar] [CrossRef] [Green Version]
- Rangel-Huerta, O.D.; Pastor-Villaescusa, B.; Aguilera, C.M.; Gil, A. A systematic review of the efficacy of bioactive compounds in cardiovascular disease: Phenolic compounds. Nutrients 2015, 7, 5177–5216. [Google Scholar] [CrossRef] [Green Version]
- Redan, B.W.; Buhman, K.K.; Novotny, J.A.; Ferruzzi, M.G. Altered transport and metabolism of phenolic compounds in obesity and diabetes: Implications for functional food development and assessment. Adv. Nutr. 2016, 7, 1090–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowska, H.; Kalinowska, M.; Lewandowski, W.; Stępkowski, T.M.; Brzoska, K. The role of natural polyphenols in cell signaling and cytoprotection against cancer development. J. Nutr. Biochem. 2016, 32, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, S.; Huh, K.; Kim, Y.; Joung, H.; Park, M. High serum isoflavone concentrations are associated with the risk of precocious puberty in Korean girls. Clin. Endocrinol. 2011, 75, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Adgent, M.A.; Daniels, J.L.; Edwards, L.J.; Siega-Riz, A.M.; Rogan, W.J. Early-life soy exposure and gender-role play behavior in children. Environ. Health Perspect. 2011, 119, 1811–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouwers, M.M.; Van Der Zanden, L.F.; De Gier, R.P.; Barten, E.J.; Zielhuis, G.A.; Feitz, W.F.; Roeleveld, N. Hypospadias: Risk factor patterns and different phenotypes. BJU Int. 2010, 105, 254–262. [Google Scholar] [CrossRef] [PubMed]
- DeAnn Cook, J.; Davis, B.J.; Goewey, J.A.; Berry, T.D.; Walker, C.L. Identification of a sensitive period for developmental programming that increases risk for uterine leiomyoma in Eker rats. Reprod. Sci. 2007, 14, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Rossen, L.M.; Simon, A.E.; Herrick, K.A. Types of infant formulas consumed in the United States. Clin. Pediatr. 2016, 55, 278–285. [Google Scholar] [CrossRef] [Green Version]
- McCarver, G.; Bhatia, J.; Chambers, C.; Clarke, R.; Etzel, R.; Foster, W.; Hoyer, P.; Leeder, J.S.; Peters, J.M.; Rissman, E. NTP-CERHR expert panel report on the developmental toxicity of soy infant formula. Birth Defects Res. Part B Dev. Reprod. Toxicol. 2011, 92, 421–468. [Google Scholar] [CrossRef]
- Harlid, S.; Adgent, M.; Jefferson, W.N.; Panduri, V.; Umbach, D.M.; Xu, Z.; Stallings, V.A.; Williams, C.J.; Rogan, W.J.; Taylor, J.A. Soy formula and epigenetic modifications: Analysis of vaginal epithelial cells from infant girls in the IFED study. Environ. Health Perspect. 2017, 125, 447–452. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ning, Y.; Guo, X. Integrative meta-analysis of differentially expressed genes in osteoarthritis using microarray technology. Mol. Med. Rep. 2015, 12, 3439–3445. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Dietary Source (Reference) | Total Phenols, mg/g DM | Majorclass | Identified Polyphenolic Compounds |
|---|---|---|---|
| Feed concentrates | |||
| Glycine max [21] | 2.23–6.18 | Isoflavone | Genistein, daidzein, glycitein, formononetin, puerarin, coumestrol |
| Linum usitatissimum [22] | 8.0–10.0 | Lignans | Secoisolariciresinol, matairesinol, pinoresinol, pinoresinol, isolariciresinol, esterified phenolic acids, Kaempferol, herbacetin |
| Forage and shrubs | |||
| Trifolium subterraneum [17] | 24.2–114.2 | Flavonoids | Flavonoids, such as flavones, flavonols and isoflavones |
| Trifolium pratense L. [11,18] | 14.82 | Isoflavones | Formononetin, biochanin A, genistein, daidzein, ononin, sissotrin, phenolic acids |
| Trifolium alexandrinum [18,23] | 52.55 | Isoflavone | Biochanin A, formononetin, genistein, daidzein, ononin, sissotrin, clovamides (caffeic acid derivatives), phenolic acids |
| Medicago sativa [18] | 70.50 | Coumestans | Coumestans, formononetin, biochanin A, genistein, daidzein, ononin, sissotrin |
| Sesbania sesban [24,25] | 167.66 | Tannins, phenolic acids | Gallic acid, catechin hydrate, vanillic acid, caffeic acid, epicatechin, rutin ellagic acid, myricetin, kaempferol, quercetin |
| Moringa oleifera [26,27] | 96.30 | Flavonols, phenolic acids | Caffeoylquinic acids, carotenoids, lutein, kaempferol, quercetin, ellagic acid, and apigenin glucoside, myricetin, rutin, isorhamnetin, ferulic acid, coumaric acids, caffeic acid, gallic acid, ellagic acid, chlorogenic acid, epicatechin |
| Lotus corniculatus [28] | - | Flavonoids | Kaempferol, quercetin, isorhamnetin and their derivatives |
| Cichorium intybus [29] | 0.65–3.73 | Flavonoids | Chlorogenic and caffeic acids |
| Salix spp. [30] | 212.0 | Tannins | Condensed tannins, salicylates, cinnamic acid derivatives, gallic acid, caffeic acid, vanillin, p-coumaric acid, myricetin, catechin, epigallocatechin gallate, rutin, quercetin, salicin |
| Acacia etbaica [31] | - | Tannins | Proanthocyanidin, galloyl derivatives kaempferol, quercetin, myricetin |
| Quercus robur [32] | 25.30–50.95 | Tannins | Ellagitannins (roburin A,B,C,D,E; grandinin, vescalagin; castalagin), protocatechuic acid/aldehyde, gallic acid, vanillic acid, caffeic acid, vanillin, syringaldehyde, coumaric acid, scopoletin, ferulic acid, sinapic acid, coniferyl aldehyde, sinapaldehyde, ellagic acid |
| Agro-industrial byproducts and feed additives | |||
| Vitis vinifera pomace [33,34] | 14.8–70.5 | Tannins, anthocyanins | Anthocyanins, condensed tannins, catechin, epicatechin, gallic acids, cyanidin 3-glucoside, malividin 3-glucoside, cyaniding, and peonidin, resveratrol |
| Olea europaea L. cake [35] | 4.1–19.4 | Flavanone | Tyrosol, hydroxytyrosol, oleuropein, verbacoside, rutin, luteolin, apigenin, quercetin |
| Citrus sinensis peel [36] | 104–223 | Flavanone | Hesperidin, quercetin, eriocitrin, narirutin, isosakuranetin rutinoside, kaempferol, gallic acid, ferulic acid, p-coumaric, catechins, epicatechins |
| Punica granatum seed [37] | 27.2 | Tannins | Flavonoids, anthocyanins, hydrolysable tannins, gallic acid |
| Punica granatum peel [38] | 48.3 | Tannins | Gallic acid, punicalagin, punicalin, flavonoids, hydrolysable tannins, condensed tannins |
| Solanum lycopersicum [39] | 6.1–6.4 | Flavonols | Naringenin, rutin, quercetin, kaempferol |
| Citrus aurantifolia [36] | 104–223 | Flavanone | Hesperidin, quercetin, kaempferol, gallic acid, ferulic acid, p-coumaric, catechins, epicatechins, |
| Camellia sinensis [40] | 148.16–252.65 | Catechins | Catechin, epicatechin, gallocatechin, epigallocatechin, catechin gallate, epicatechin gallate, gallocatechin gallate, gallic acid, ellagic acid, quercitrin, astragalin, quercetin, kaempferol, chlorogenic acid, myricetin |
| Phoenix dactylifera pits [41] | 12.7–47.7 | Phenolic acids | Hydroxytyrosol, tyrosol oleuropein, gallic acid, ferulic acid, coumaric acids, p-hydroxybenzoic acid, flavonoids |
| Propolis [42,43] | 65.49–228.4 | Phenolic acids, flavonoids | Gallic acid, caffeic acid, catechin, chlorogenic acid, p-coumaric acid, ferulic acid, naringenin, quercetin, apigenin, baicalin, luteolin, pinocembrin, galangin |
| Animal (Reference) | Treatment | Main Results |
|---|---|---|
| Barki rams [26] | 0, 40 mg/kg diet Moringa oleifera leaves extract |
|
| Boar [47] | 0, 2, 4% of diet Vitis vinifera marc | Both levels:
|
| Lambs [57] | 0,4, 8% of diet Linum usitatissimum | At 8%:
|
| Rabbit bucks [45] | 0,5, 20 mg/kg body weight Glycine max isoflavones (5 daidzein: 1 genistein) | At both levels:
|
| Rabbit bucks [44] | Glycine max-based and Linum usitatissimum-based diets | Both diets:
|
| Cloned goat bucks [66] | 0, 8.83, 17.66% of diet Punica granatum seed for 9 weeks |
|
| Rabbit bucks [67] | 0, 5, 10 g/kg of diet milk thistle seeds and rosemary leaves | At milk thistle seeds 10 and rosemary leaves at 5 g/kg diet:
|
| Heat stressed rabbit bucks [68] | 0, 50, 100, 150 mg/kg body weight (BW) Moringa oleifera leaves extract | All levels:
|
| Ram lambs kept under restrained conditions (pen conditions) [69] | 0, 5, 10% of DM diet wine Punica granatum pomace for 74 days |
|
| Animal | Treatment | Main Results |
|---|---|---|
| Rabbit does [5] | 0, 5, 20 g/100 kg diet Camellia sinensis powder from weaning (45 days old) and throughout two consequent reproductive cycles |
|
| Buffalo cows [70] | 0, 100, 200 g/head/day Quebracho tannins |
|
| Seasonal anestrous Rahmani ewes [13] | T. alexandrinum vs. corn silage 2 weeks prepartum to 8 weeks post induced estrus |
|
| Prepubertal hair breed ewe lambs during the natural anestrous season [65] | 0, 300 mg of ferulic acid/day/head for 34 days |
|
| Barki ewes | Quebracho condensed tannins (20 g/head/day) for four weeks pre-mating to lambing |
|
| Barki, fat-tailed, Ewes [71] | 50, 100% substitution of clover hay by tannins-rich plant (Acacia saligna) |
|
| Rabbit does [73] | Yucca schidigera extract (0, 5, 20 g of Y powder extract per 100 kg diet for 350 days. |
|
| Holstein heifers [59] | Trifolium alexandrinum vs. corn silage for five month, 3 premating plus two post-mating |
|
| Cypermethrin-challenged rabbit does [43] | 50 mg/kg bodyweight propolis |
|
| Finnish Landrace ewe lamb [11] | T. pratense L. silage vs. grass silage for five month, 3 premating plus two post-mating |
|
| Prepubertal female Tuj lambs [70] | Quercus hartwissiana |
|
| Menz ewes [25] | 0.28% of diet Sesbania sesban during pre-mating, mating, pregnancy and lambing |
|
| ARTs | Source/Dose | Main Results |
|---|---|---|
| Semen frozen-thawed extender of buffalo [130] | 0, 10, 20, 50, 100 µM/mL RES | At 50, 100 µM/mL:
|
| Liquid store (cooling at 5 ◦C) extender for 168 h of ram semen [131] | 0, 200, 400 µM/mL RES | At 400 µM/mL:
|
| Semen frozen-thawed extender of rams [132] | 0, 10, 30, 50, 70 μg/mL of HT, DHPG and a mixture (MIX) | At all levels of HT and DHPG:
|
| Liquid storage (4 °C, 120 h) of goat semen [133] | 0, 10, 30, 50, and 70 mg/L GSPE | At 30 mg/L:
|
| Frozen-thawed extender of goat semen [134] | 0, 10, 50, 100, 250 μM/mL RES | At 10 or 50 μM/mL:
|
| IVF medium of boar semen [135] | 0, 1, 10, 100 µg/mL tannin (Quercus robur) and its four fractions (FA, FB, FC, FD), | At 10 µg/mL:
|
| Frozen-thawed extender of buffalo semen [136] | 0, 0.5, 1, 10 and 50 µM/mL RES | At 50 µM/mL:
|
| Post-thawing semen extender of boar for 1 h [137] | 0, 0.5, 1, 2 mM/mL RES or 0, 25, 50, 100 µM/mL EGCG |
|
| IVF medium using cryopreserved bovine spermatozoa [138] | 0, 0.074, 0.74, 7.4 µmol/L GEN |
|
| ARTs | Source/Dose | Main Results |
|---|---|---|
| IVM of ewe oocytes [5,14] | Methanolic plant extracts: 0, 50, 100 mg/mL Bituminaria bituminosa, Medicago sativa, Cichorium intybus, Trifolium (T.) subterraneum, T. pratense L, Biserrula pelecinus and Eremophila glabra | At 100 mg/L: B. pelecinus improved fertilization and embryo development.
|
| IVC of ovarian fragments from of rabbit does [5] | 0, 1, 10 or 100 mg/mL EGCG, GTPP and RSV |
|
| IVM of bovine oocytes either supplemented (IVM A) or not supplemented (IVM B) with cysteine and b-mercaptoethanol [139] | 0, 1, 5, 10, 20 µg/mL oenological Quercus robur-derived tannin | At all tested levels:
|
| IVM of prepubertal goat oocytes with high [16] (+) or low (−) quality | 0, 1 µM/mL RSV |
|
| Exp 1: IVC, alone, of sheep embryos Exp 2: IVM of oocyte and IVC of sheep embryos [140] | 0, 0.1, 0.25, 0.5, 2.0, 5.0 µM/mL RSV | Exp 1: At 0.25 and 0.5 µM/mL:
|
| IVM of oocytes and IVF in sheep [141] | 0, 2.5, 5, 10, 25 µg/mL GEN, BIO A, FOR | At 25 µg/mL:
|
| IVM of bovine oocytes [15] | 0, 10, 15, 20 μM/mL green tea catechin | At 15 μM/mL:increased cleavage and blastocyst rates and intracellular GSH concentration of oocytes |
| IVM and IVF of swine oocyte [64] | 0, 2.5, 5, 10,25 µg/mL EGCG | At ≤10 mg/mL:
|
| IVC of swine granulosa cells (From follicles >5 mm) [3] | 0, 5, 50 µg/mL EGCG |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hashem, N.M.; Gonzalez-Bulnes, A.; Simal-Gandara, J. Polyphenols in Farm Animals: Source of Reproductive Gain or Waste? Antioxidants 2020, 9, 1023. https://doi.org/10.3390/antiox9101023
Hashem NM, Gonzalez-Bulnes A, Simal-Gandara J. Polyphenols in Farm Animals: Source of Reproductive Gain or Waste? Antioxidants. 2020; 9(10):1023. https://doi.org/10.3390/antiox9101023
Chicago/Turabian StyleHashem, Nesrein M., Antonio Gonzalez-Bulnes, and Jesus Simal-Gandara. 2020. "Polyphenols in Farm Animals: Source of Reproductive Gain or Waste?" Antioxidants 9, no. 10: 1023. https://doi.org/10.3390/antiox9101023