Dendropanax morbifera Protects against Renal Fibrosis in Streptozotocin-Induced Diabetic Rats

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of DP Extract from the Leaf and Stem of D. Morbifera
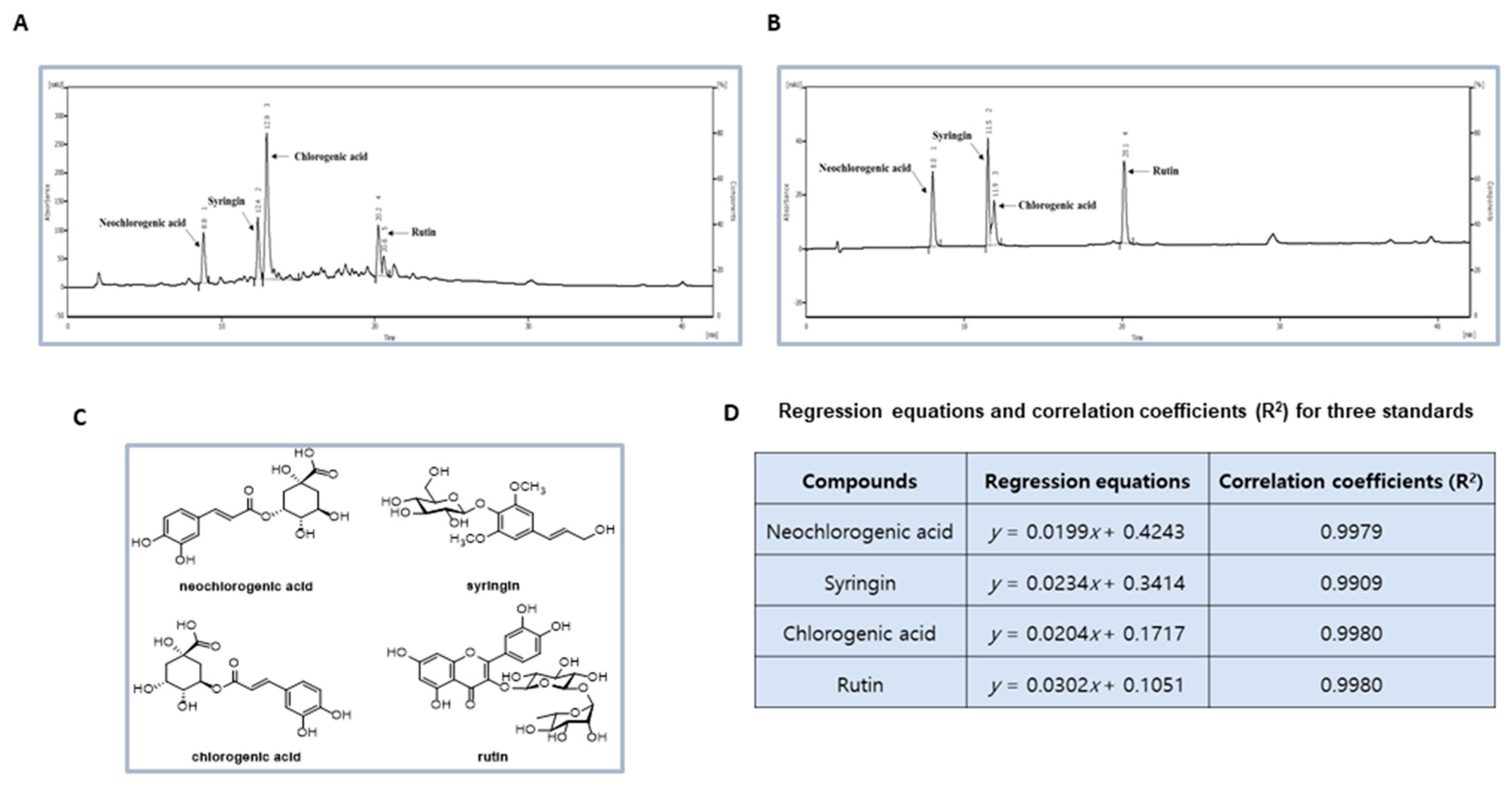
2.3. HPLC Analysis of the DP Extract
2.4. Assessment of Acute Toxicity of DP extract
2.5. Experimental Design
2.6. Assessment of Biochemical Parameters
2.7. Urinary Parameters Analysis
2.8. Histopathological Examination of Liver, Kidney, and Pancreas
2.9. Immunohistochemical Analysis
2.10. Analysis of Advanced Glycation end Products
2.11. Determination of 3-Indoxyl Sulfate (3-IS)
2.12. Measurement of 4-Hydroxyproline Content
2.13. Determination of Antioxidant Enzymes Activity and Oxidative Stress
2.14. Determination of Inflammatory Cytokines in the Serum
2.15. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL) Assay
2.16. Western Blot Analysis
2.17. Statistical Analysis
3. Results
3.1. Phytochemcials Characterization of DP Extract
3.2. Acute Oral Toxicity Study of DP Extract
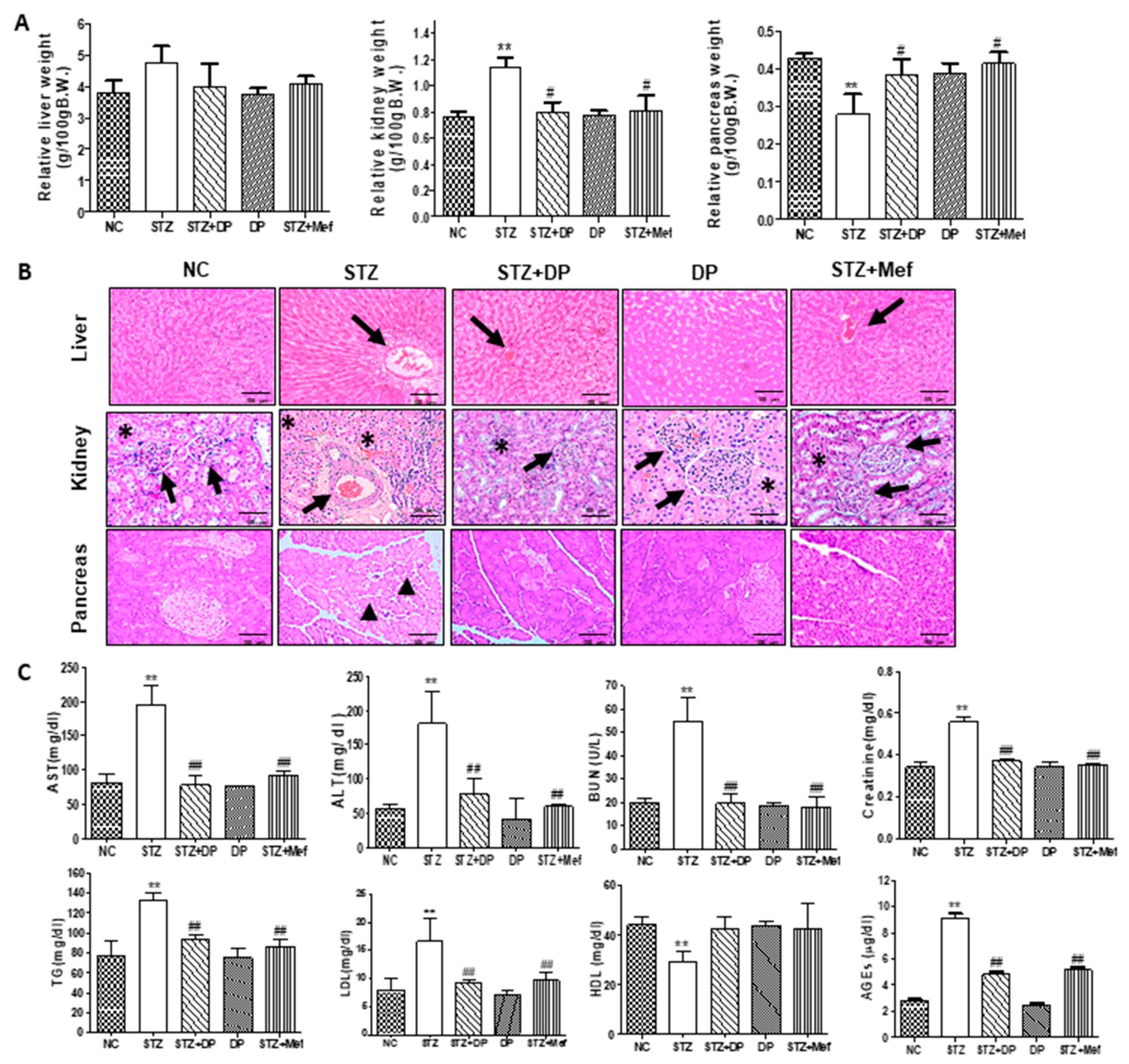
3.3. Effect of DP Extract on Body Weight, Glucose Concentration, Organ Weight, and Histology in STZ-Induced Diabetic Rats
3.4. Effect of DP Extract on Biochemical Parameters in Diabetic Rats
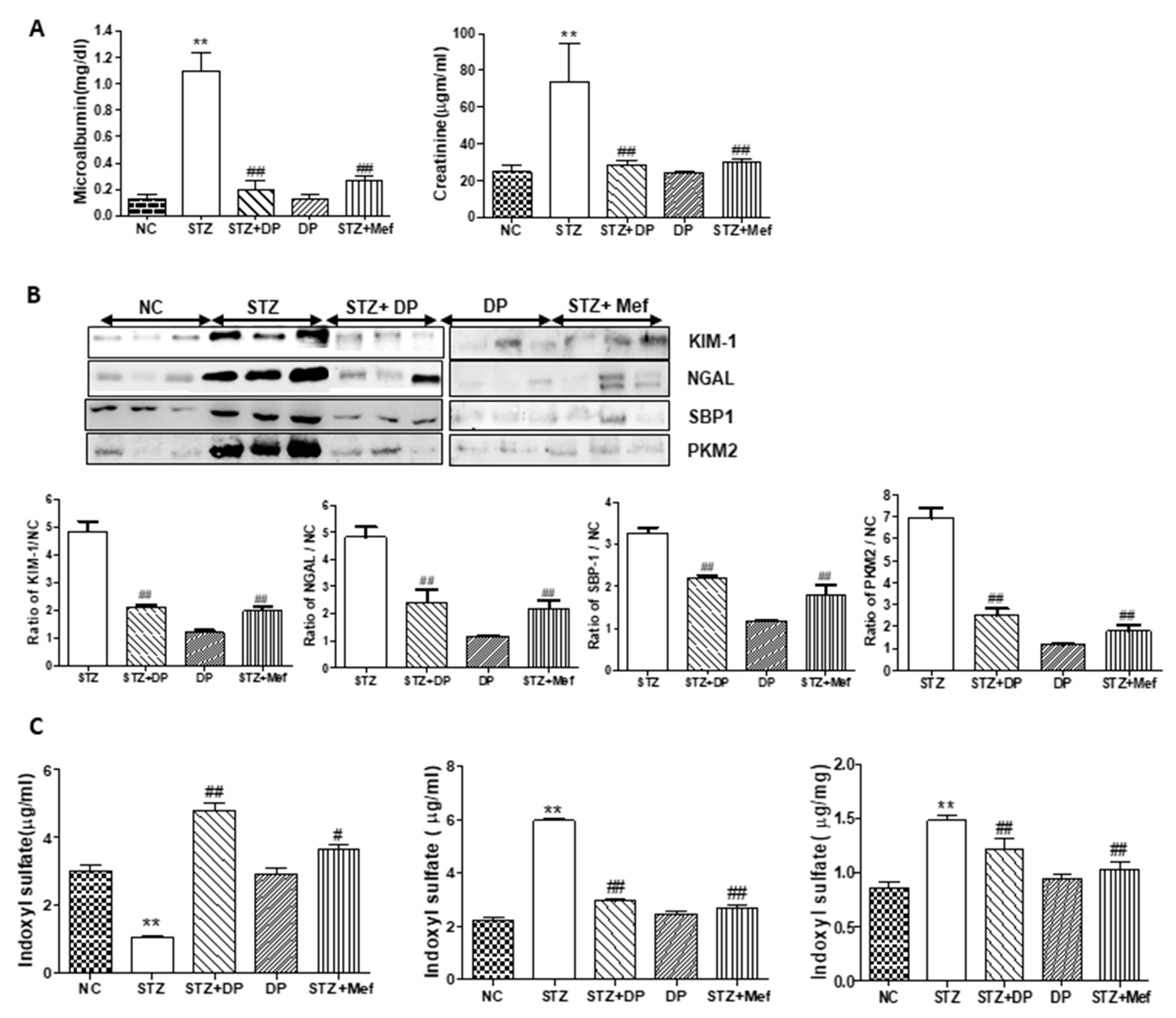
3.5. Effect of DP Extract on Urinary Excretion of Kidney Injury Biomarkers in Diabetic Rats
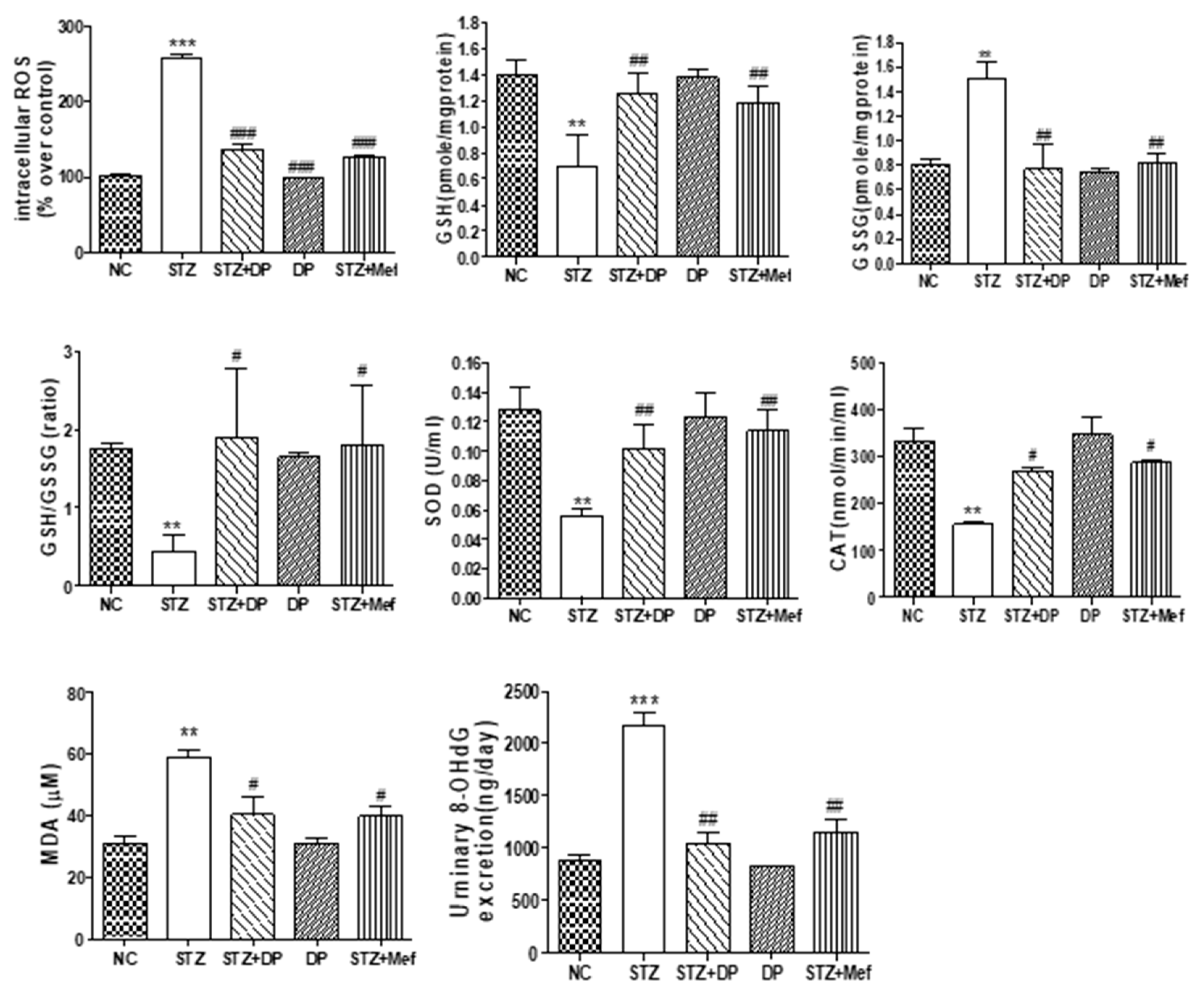
3.6. Effect of DP Extract on ROS Production, Antioxidant Enzyme Activity, and Oxidative Stress in the Kidney of Diabetic Rats
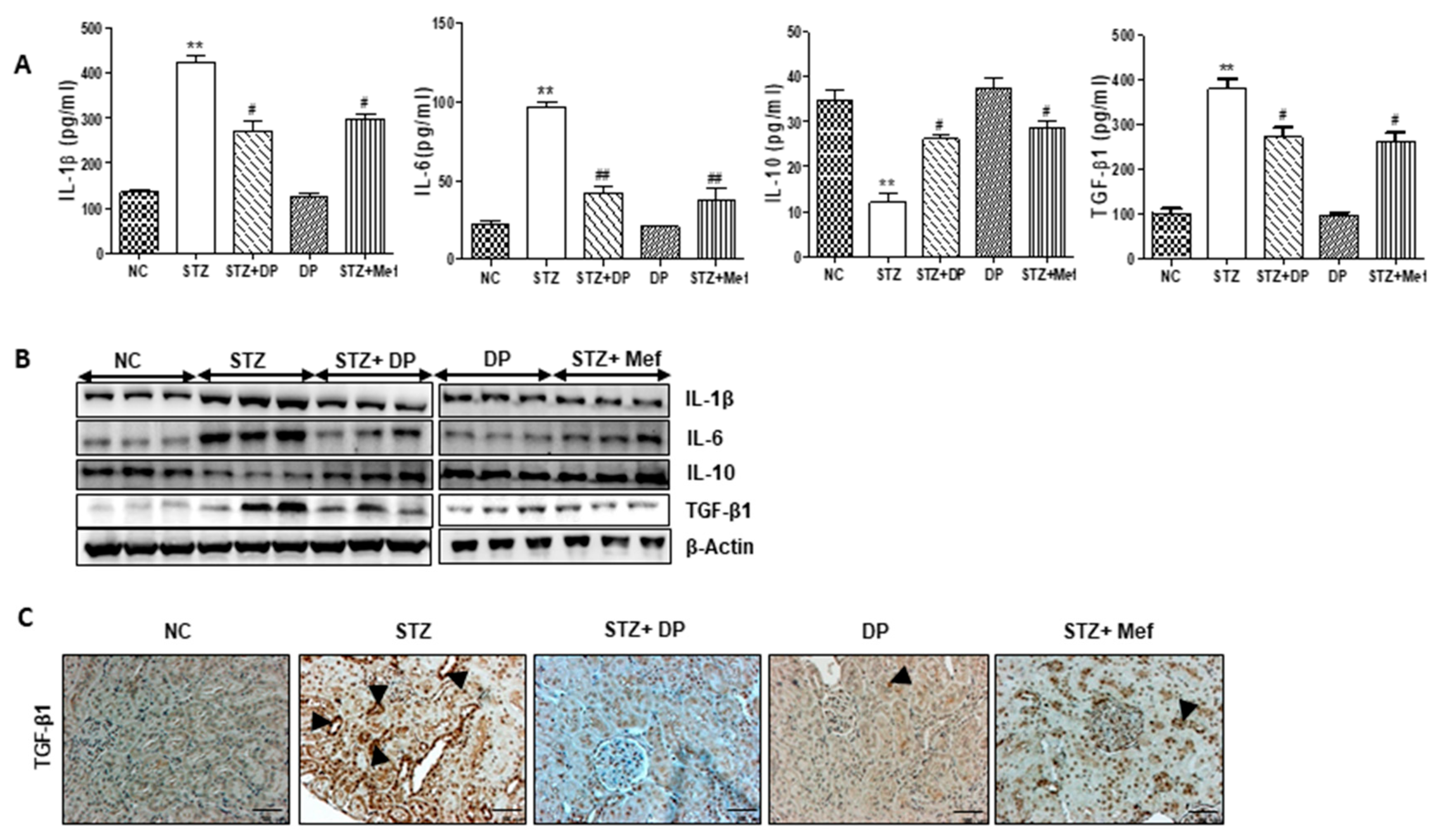
3.7. Effect of DP Extract on Production of Pro-Inflammatory Cytokines in Diabetic Rats
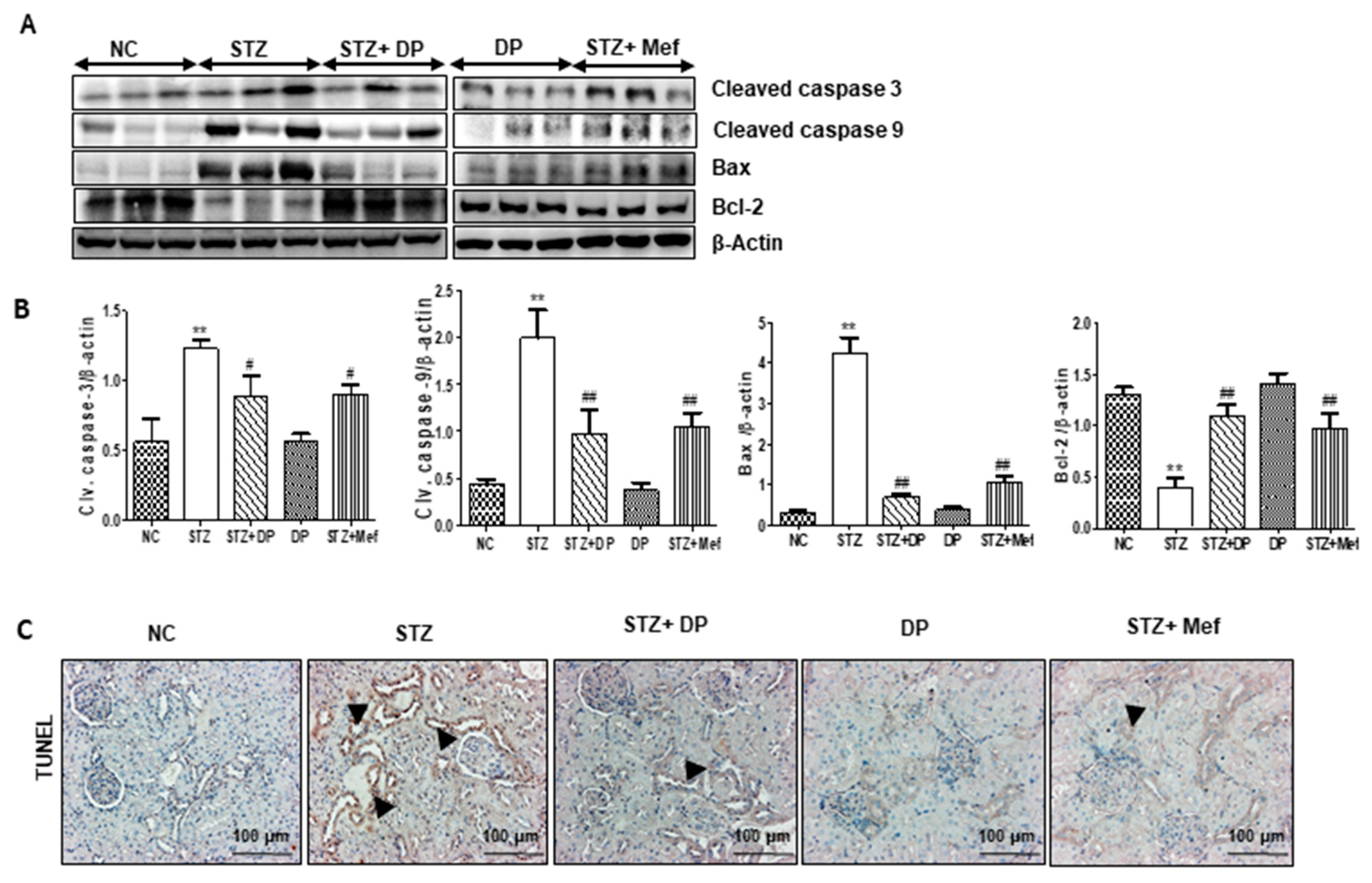
3.8. Effect of DP Extract on Apoptosis in the Kidney of Diabetic Rats
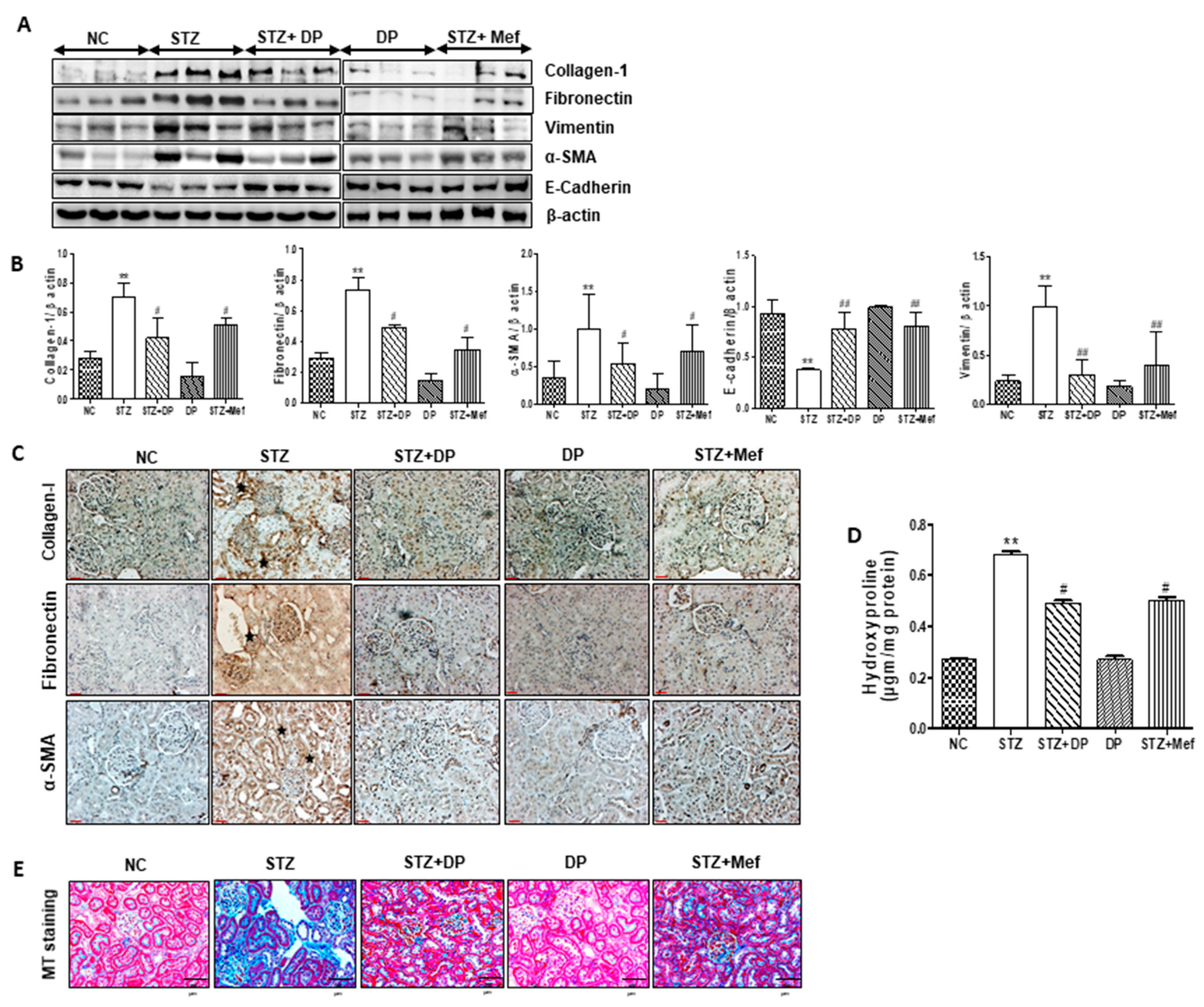
3.9. Effect of DP Extract on the Composition of ECM in the Kidney of Diabetic Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hameed, I.; Masoodi, S.R.; Mir, S.A.; Nabi, M.; Ghazanfar, K.; Ganai, B.A. Type 2 diabetes mellitus: From a metabolic disorder to an inflammatory condition. World J. Diabetes 2015, 6, 598–612. [Google Scholar] [CrossRef]
- Shang, G.; Gao, P.; Zhao, Z.; Chen, Q.; Jiang, T.; Zhang, N.; Li, H. 3,5-Diiodo-l-thyronine ameliorates diabetic nephropathy in streptozotocin-induced diabetic rats. Biochim. Biophys. Acta 2013, 1832, 674–684. [Google Scholar] [CrossRef] [Green Version]
- Gheith, O.; Farouk, N.; Nampoory, N.; Halim, M.A.; Al-Otaibi, T. Diabetic kidney disease: Worldwide difference of prevalence and risk factors. J. Nephropharmacol. 2016, 5, 49–56. [Google Scholar] [CrossRef]
- Park, J.I.; Baek, H.; Jung, H.H. Prevalence of chronic kidney disease in Korea: The Korean national health and nutritional examination survey 2011–2013. J. Korean Med. Sci. 2016, 31, 915–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnudi, L.; Coward, R.J.M.; Long, D.A. Diabetic nephropathy: Perspective on novel molecular mechanisms. Trends Endocrinol. Metab. 2016, 27, 820–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keane, W.F.; Brenner, B.M.; de Zeeuw, D.; Grunfeld, J.P.; McGill, J.; Mitch, W.E.; Ribeiro, A.B.; Shahinfar, S.; Simpson, R.L.; Snapinn, S.M.; et al. The risk of developing end-stage renal disease in patients with type 2 diabetes and nephropathy: The RENAAL study. Kidney Int. 2003, 63, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Roshan, B.; Stanton, R.C. A story of microalbuminuria and diabetic nephropathy. J. Nephropathol. 2013, 2, 234–240. [Google Scholar] [PubMed]
- Alsaad, K.O.; Herzenberg, A.M. Distinguishing diabetic nephropathy from other causes of glomerulosclerosis: An update. J. Clin. Pathol. 2007, 60, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brownlee, M. The pathobiology of diabetic complications: A unifying mechanism. Diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef] [Green Version]
- Cooper, M.E. Importance of advanced glycation end products in diabetes-associated cardiovascular and renal disease. Am. J. Hypertens. 2004, 17, S31–S38. [Google Scholar] [CrossRef]
- Kolset, S.O.; Reinholt, F.P.; Jenssen, T. Diabetic nephropathy and extracellular matrix. J. Histochem. Cytochem. 2012, 60, 976–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Y.; Feldman, E.; Pennathur, S.; Kretzler, M.; Brosius, F.C. From fibrosis to sclerosis: Mechanisms of glomerulosclerosis in diabetic nephropathy. Diabetes 2008, 57, 1439–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffes, M.W.; Osterby, R.; Chavers, B.; Mauer, S.M. Mesangial expansion as a central mechanism for loss of kidney function in diabetic patients. Diabetes 1989, 38, 1077–1081. [Google Scholar] [CrossRef] [PubMed]
- Donate-Correa, J.; Martín-Núñez, E.; Muros-de-Fuentes, M.; Mora-Fernández, C.; Navarro-González, J.F. Inflammatory cytokines in diabetic nephropathy. J. Diabetes Res. 2015, 2015, 948417. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Saito, T.; Ootaka, T.; Soma, J.; Obara, K.; Abe, K.; Yoshinaga, K. The role of macrophages in diabetic glomerulosclerosis. Am. J. Kidney Dis. 1993, 21, 480–485. [Google Scholar] [CrossRef]
- Fan, Y.; Li, X.; Xiao, W.; Fu, J.; Harris, R.C.; Lindenmeyer, M.; Cohen, C.D.; Guillot, N.; Baron, M.H.; Wang, N.; et al. BAMBI elimination enhances alternative TGF-β signaling and glomerular dysfunction in diabetic mice. Diabetes 2015, 64, 2220–2233. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Hong, S.; Dong, L.; Cheng, B.; Lin, L.; Zhao, B.; Chen, Y.G.; Chen, X. Dynamic sialylation in transforming growth factor-β (TGF-β)-induced epithelial to mesenchymal transition. J. Biol. Chem. 2015, 290, 12000–12013. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Schnaper, H.W.; Matsusaka, T.; Pastan, I.; Ledbetter, S.; Hayashida, T. Anti-TGF-β antibody, 1D11, ameliorates glomerular fibrosis in mouse models after the onset of proteinuria. PLoS ONE 2016, 11, e0155534. [Google Scholar] [CrossRef]
- Kim, M.; Park, Y.J.; Lim, H.S.; Lee, H.H.; Kim, T.H.; Lee, B. The clinical effects of Dendropanax morbifera on postmenopausal symptoms: Review article. J. Menopausal Med. 2017, 23, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Hyun, T.K.; Ko, Y.J.; Kim, E.H.; Chung, I.M.; Kim, J.S. Anti-inflammatory activity and phenolic composition of Dendropanax morbifera leaf extracts. Ind. Crops Prod. 2015, 74, 263–270. [Google Scholar] [CrossRef]
- Lim, A.K.H. Diabetic nephropathy—Complications and treatment. Int. J. Nephrol. Renov. Dis. 2014, 7, 361–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.Y.; Min, B.S.; Oh, S.R.; Kim, J.H.; Kim, T.J.; Kim, D.H.; Bae, K.H.; Lee, H.K. Isolation and anticomplement activity of compounds from Dendropanax morbifera. J. Ethnopharmacol. 2004, 90, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Kim, M.Y.; Park, S.D.; Park, W.H.; Moon, H.I. In vitro evaluation of the antiplasmodial activity of Dendropanax morbifera against chloroquine-sensitive strains of Plasmodium falciparum. Phytother. Res. 2009, 23, 1634–1637. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Seo, S.H.; Kang, E.Y.; Park, S.D.; Park, W.H.; Moon, H.I. Chemical composition and larvicidal effects of essential oil of Dendropanax morbifera against Aedes aegypti L. Biochem. Syst. Ecol. 2009, 37, 470–473. [Google Scholar] [CrossRef]
- Moon, H.I. Antidiabetic effects of dendropanoxide from leaves of Dendropanax morbifera Leveille in normal and streptozotocin-induced diabetic rats. Hum. Exp. Toxicol. 2011, 30, 870–875. [Google Scholar] [CrossRef]
- Achi, N.K.; Ohaeri, O.C.; Ijeh, I.I.; Eleazu, C. Modulation of the lipid profile and insulin levels of streptozotocin induced diabetic rats by ethanol extract of Cnidoscolus aconitifolius leaves and some fractions: Effect on the oral glucose tolerance of normoglycemic rats. Biomed. Pharmacother. 2017, 86, 562–569. [Google Scholar] [CrossRef]
- Lou, J.; Gu, X.; Xing, Y.; Cui, J.; Lv, W.; Zhang, Y. Chlorogenic acid slows down proteinuria and renal fibrosis in 5/6-nephrectomized rats by anti-oxidation and inhibiting accumulation of extracellular matrix. Int. J. Clin. Exp. Med. 2016, 9, 15719–15727. [Google Scholar]
- Kaur, N.; Kishore, L.; Singh, R. Dillenia indica L. attenuates diabetic nephropathy via inhibition of advanced glycation endproducts accumulation in STZ-nicotinamide induced diabetic rats. J. Tradit. Complementary Med. 2017, 8, 226–238. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′ -deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Won, A.J.; Kim, S.; Kim, Y.G.; Kim, K.B.; Choi, W.S.; Kacew, S.; Kim, K.S.; Jung, J.H.; Lee, B.M.; Kim, S.; et al. Discovery of urinary metabolomic biomarkers for early detection of acute kidney injury. Mol. Biosyst. 2016, 12, 133–144. [Google Scholar] [CrossRef]
- Zhao, L.; Xu, L.; Tao, X.; Han, X.; Yin, L.; Qi, Y.; Peng, J. Protective effect of the total flavonoids from Rosa laevigata Michx fruit on renal ischemia-reperfusion injury through suppression of oxidative stress and inflammation. Molecules 2016, 21, 952. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liao, Q.; Luo, Y.; Qing, Z.; Zhang, Q.; He, G. Renal protective effect of Rosa laevigata Michx. by the inhibition of oxidative stress in streptozotocin-induced diabetic rats. Mol. Med. Rep. 2012, 5, 1548–1554. [Google Scholar] [PubMed] [Green Version]
- Han, F.; Xue, M.; Chang, Y.; Li, X.; Yang, Y.; Sun, B.; Chen, L. Triptolide suppresses glomerular mesangial cell proliferation in diabetic nephropathy is associated with inhibition of PDK1/Akt/mTOR pathway. Int. J. Biol. Sci. 2017, 13, 1266–1275. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.X.; Xu, X.X.; Wang, K.; Qi, X.M.; Wu, Y.G. Paeoniflorin attenuates incipient diabetic nephropathy in streptozotocin-induced mice by the suppression of the Toll-like receptor-2 signaling pathway. Drug Des. Devel. Ther. 2017, 11, 3221–3233. [Google Scholar] [CrossRef] [Green Version]
- Domitrović, R.; Cvijanović, O.; Šušnić, V.; Katalinić, N. Renoprotective mechanisms of chlorogenic acid in cisplatin-induced kidney injury. Toxicology 2014, 324, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Chen, S.; Chen, L.; Zhang, L.; Meng, F.; Sha, S.; Ai, C.; Tai, J. Chlorogenic acid ameliorates lead-induced renal damage in mice. Biol. Trace Elem. Res. 2019, 189, 109–117. [Google Scholar] [CrossRef]
- Arjumand, W.; Seth, A.; Sultana, S. Rutin attenuates cisplatin induced renal inflammation and apoptosis by reducing NFκB, TNF-α and caspase-3 expression in wistar rats. Food Chem. Toxicol. 2011, 49, 2013–2021. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, X.; Feng, T.; Jin, G.; Li, Z. Rutin prevents high glucose-induced renal glomerular endothelial hyperpermeability by inhibiting the ROS/Rhoa/ROCK signaling pathway. Planta Med. 2016, 82, 1252–1257. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.Y.; Kim, K.S.; Lee, Y.H.; Park, J.H.; Kim, J.H.; Lee, S.Y.; Kim, Y.M.; Kim, I.S.; Kacew, S.; Lee, B.M.; et al. Dendropanax morbifera ameliorates thioacetamide- induced hepatic fibrosis via TGF-β1/Smads pathways. Int. J. Biol. Sci. 2019, 15, 800–811. [Google Scholar] [CrossRef] [Green Version]
- Fiseha, T. Urinary biomarkers for early diabetic nephropathy in type 2 diabetic patients. Biomark. Res. 2015, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S.; Yang, H.Y.; Song, H.; Kang, Y.R.; Kwon, J.; An, J.; Son, J.Y.; Kwack, S.J.; Kim, Y.M.; Bae, O.N.; et al. Identification of a sensitive urinary biomarker, selenium-binding protein 1, for early detection of acute kidney injury. J. Toxicol. Environ. Health A 2017, 80, 453–464. [Google Scholar] [CrossRef]
- Parikh, C.R.; Edelstein, C.L.; Devarajan, P.; Cantley, L. Biomarkers of acute kidney injury: Early diagnosis, pathogenesis, and recovery. J. Investig. Med. 2007, 55, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.M.; Coughlan, M.T.; Cooper, M.E. Oxidative stress as a major culprit in kidney disease in diabetes. Diabetes 2008, 57, 1446–1454. [Google Scholar] [CrossRef] [Green Version]
- Ha, H.; Hwang, I.A.; Park, J.H.; Lee, H.B. Role of reactive oxygen species in the pathogenesis of diabetic nephropathy. Diabetes Res. Clin. Pract. 2008, 82, S42–S45. [Google Scholar] [CrossRef] [PubMed]
- King, G.L.; Loeken, M.R. Hyperglycemia-induced oxidative stress in diabetic complications. Histochem. Cell Biol. 2004, 122, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, O.; Yan, S.D.; Ogawa, S.; Kuwabara, K.; Matsumoto, M.; Stern, D.; Schmidt, A.M. The receptor for advanced glycation end-products on the development of vascular disease in diabetes mellitus. Nephrol. Dial. Transplant. 1996, 11, 13–16. [Google Scholar] [CrossRef]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B. Diabetes, oxidative Stress, and antioxidants: A review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef]
- Allen, D.A.; Harwood, S.; Varagunam, M.; Raftery, M.J.; Yaqoob, M.M. High glucose-induced oxidative stress causes apoptosis in proximal tubular epithelial cells and is mediated by multiple caspases. FASEB J. 2003, 17, 908–910. [Google Scholar] [CrossRef]
- Jiménez-Osorio, A.S.; García-Niño, W.R.; González-Reyes, S.; Álvarez-Mejía, A.E.; Guerra-León, S.; Salazar-Segovia, J.; Falcón, I.; Montes de Oca-Solano, H.; Madero, M.; Pedraza-Chaverri, J. The effect of dietary supplementation with curcumin on redox status and Nrf2 activation in patients with nondiabetic or diabetic proteinuric chronic kidney disease: A pilot study. J. Ren. Nutr. 2016, 26, 237–244. [Google Scholar] [CrossRef]
- Liakopoulos, V.; Roumeliotis, S.; Bozikas, A.; Eleftheriadis, T.; Dounousi, E. Antioxidant supplementation in renal replacement therapy patients: Is there evidence? Oxidative Med. Cell. Longev. 2019, 2019, 9109473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. Inflammatory processes in renal fibrosis. Nat. Rev. Nephrol. 2014, 10, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Muragaki, Y.; Saika, S.; Roberts, A.B.; Ooshima, A. Targeted disruption of TGF-β1/Smad3 signaling protects against renal tubulointerstitial fibrosis induced by unilateral ureteral obstruction. J. Clin. Invest. 2003, 112, 1486–1494. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.B.; Andreoli, T.E. Transforming growth factor β contributes to progressive diabetic nephropathy. Proc. Natl. Acad. Sci. USA 2000, 97, 7667–7669. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | Day 6 | Day 7 |
|---|---|---|---|---|---|---|---|
| Group 1 (100 mg/kg/day) | 120.67 ± 0.58 | 120 ± 2.65 | 118.33 ± 5.13 | 121.67 ± 1.154 | 119 ± 1 | 114.67 ± 3.05 | 121 ± 2 |
| Group 2 (500 mg/kg/day) | 119.67 ± 1.163 | 115.33 ± 1.02 | 121.33 ± 3.05 | 120 ± 1.13 | 119 ± 2 | 116.67 ± 1.71 | 121 ± 3.11 |
| Group 3 (1000 mg/kg/day) | 121.67 ± 1.51 | 122.33 ± 2.30 | 120 ± 1 | 120.67 ± 0.57 | 117 ± 2.54 | 120.33 ± 2.21 | 121 ± 1.08 |
| Group 4 (3000 mg/kg/day) | 120.33 ± 2.23 | 118 ± 2 | 120.33 ± 1.154 | 119.33 ± 1.01 | 119 ± 1 | 119.33 ± 1.50 | 120.68 ± 2.51 |
| Group | Week 0 | Week 1 | Week 2 | Week 3 | Week 4 | Relative Growth Rate (%) |
|---|---|---|---|---|---|---|
| NC | 211.65 ± 5.38 | 219.13 ± 8.23 | 231.67 ± 4.01 | 251.17 ± 4.57 | 267.4 ± 6.41 | 26.3 (i/c) |
| STZ | 211.67 ± 6.39 | 211.43 ± 8.01 | 202.93 ± 1.50 a | 193.63 ± 2.23 a | 184.33 ± 5.64 a | −12.9 (d/c) |
| DP | 213 ± 2.08 | 221.47 ± 1.56 | 231.87 ± 2.04 b | 250.6 ± 2.07 b | 269.66 ± 1.53 b | 26.4 (i/c) |
| STZ + DP | 212.63 ± 5.83 | 216.25 ± 1.43 | 225.98 ± 1.84 b | 245.83 ± 0.28 b | 255.19 ± 5.15 b | 20.01 (i/c) |
| STZ + Mef | 214.34 ± 4.04 | 223.3 ± 5.81 | 229.53 ± 7.51 b | 240.63 ± 1.46 b | 252.03 ± 2.61 b | 17.5 (i/c) |
| Group | Week 0 | Week 1 | Week 2 | Week 3 | Week 4 | Change (%) |
|---|---|---|---|---|---|---|
| NC | 131 ± 8.01 | 132.33 ± 1.52 | 129.33 ± 1.15 | 130.33 ± 1.527 | 131.66 ± 2.08 | 0.50 (i/c) |
| STZ | 423.33 ± 40.05 a | 515 ± 61.55 a | 563.33 ± 32.15 a | 593 ± 11.27 a | 599.33 ± 0.58 a | 41.57 (i/c) |
| DP | 134 ± 4.36 a | 127.66 ± 3.21 a,b | 128.66 ± 1.15 b | 129.66 ± 0.57 b | 129 ± 4.58 b | −3.73(d/c) |
| STZ + DP | 432 ± 44.50 b | 318.33 ± 96.02 a,b | 233.66 ± 104.04 b | 150 ± 26.46 b | 131.67 ± 9.07 b | −69.52 (d/c) |
| STZ + Mef | 422 ± 32.36 a | 347.33 ± 40.77 a,b | 258.67 ± 4.51 a,b | 171.67 ± 6.03 a,b | 139 ± 7.21 b | −67.06 (d/c) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sachan, R.; Kundu, A.; Dey, P.; Son, J.Y.; Kim, K.S.; Lee, D.E.; Kim, H.R.; Park, J.H.; Lee, S.H.; Kim, J.-H.; et al. Dendropanax morbifera Protects against Renal Fibrosis in Streptozotocin-Induced Diabetic Rats. Antioxidants 2020, 9, 84. https://doi.org/10.3390/antiox9010084
Sachan R, Kundu A, Dey P, Son JY, Kim KS, Lee DE, Kim HR, Park JH, Lee SH, Kim J-H, et al. Dendropanax morbifera Protects against Renal Fibrosis in Streptozotocin-Induced Diabetic Rats. Antioxidants. 2020; 9(1):84. https://doi.org/10.3390/antiox9010084
Chicago/Turabian StyleSachan, Richa, Amit Kundu, Prasanta Dey, Ji Yeon Son, Kyeong Seok Kim, Da Eun Lee, Hae Ri Kim, Jae Hyeon Park, Su Hyun Lee, Jung-Hwan Kim, and et al. 2020. "Dendropanax morbifera Protects against Renal Fibrosis in Streptozotocin-Induced Diabetic Rats" Antioxidants 9, no. 1: 84. https://doi.org/10.3390/antiox9010084