Up-Regulation of Superoxide Dismutase 2 in 3D Spheroid Formation Promotes Therapeutic Potency of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. UCB-MSCs Culture
2.2. Spheroid Formation of UCB-MSCs
2.3. 2-D Electrophoresis
2.4. In-Gel Digestion and Mass Spectrometry Analysis
2.5. Reverse Transcription Polymerase Chain Reaction and RNA Interference
2.6. Western Blot Analysis
2.7. Immunofluorescent Staining
2.8. Superoxide Dismutase 2 Activity Assay
2.9. Acquisition and Treatment of Spheroid with Hyaluronic Acid/Synovial Fluid
2.10. Live/Dead Staining
2.11. Determination of ROS Levels
2.12. Animal Models and Histopathological Analysis
2.13. Statistical Analyses
3. Results
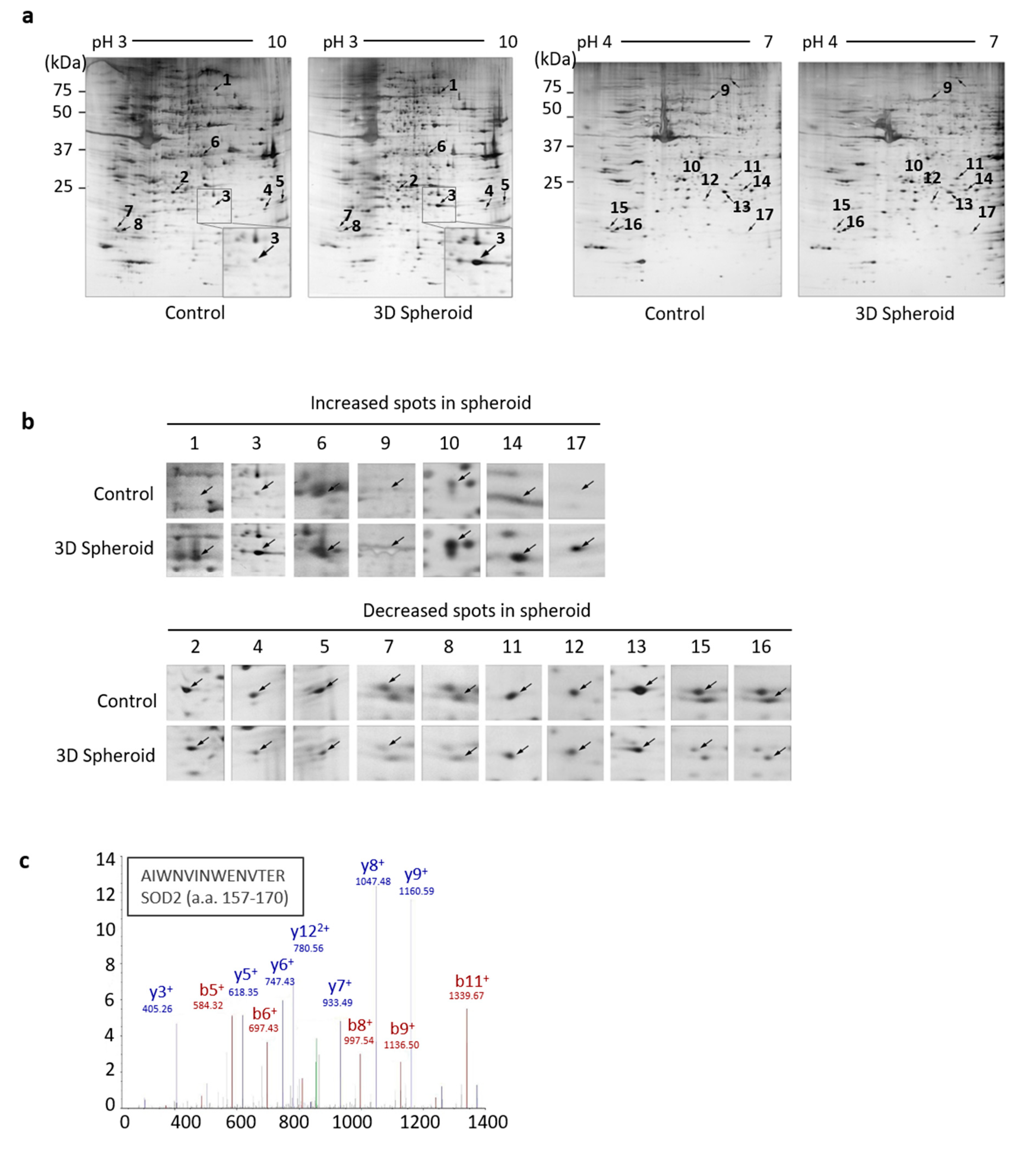
3.1. Identification of SOD2 in 3D Spheroid Form of UCB-MSCs
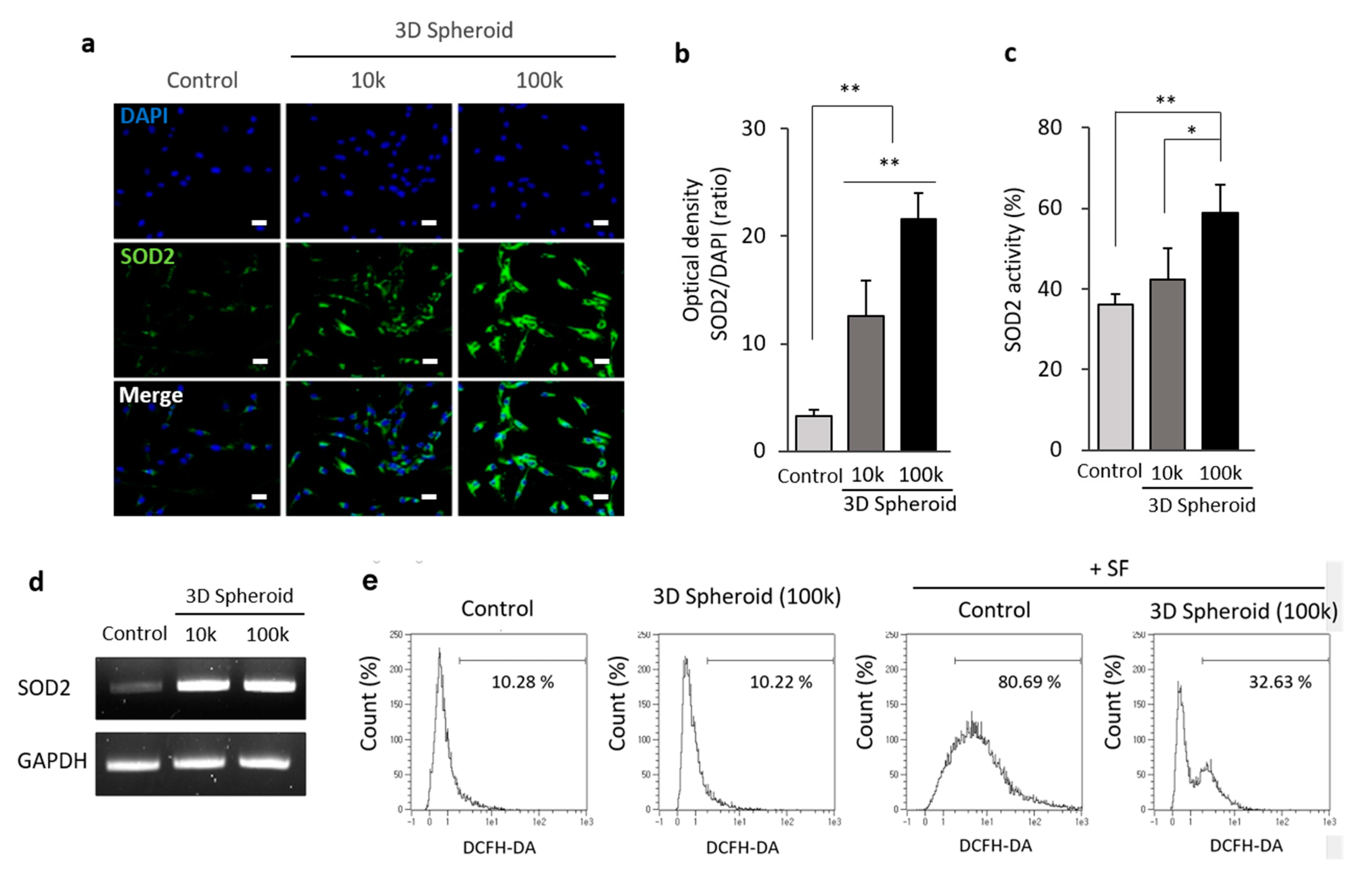
3.2. The Increased Expression of SOD2 in 3D Spheroid Formation
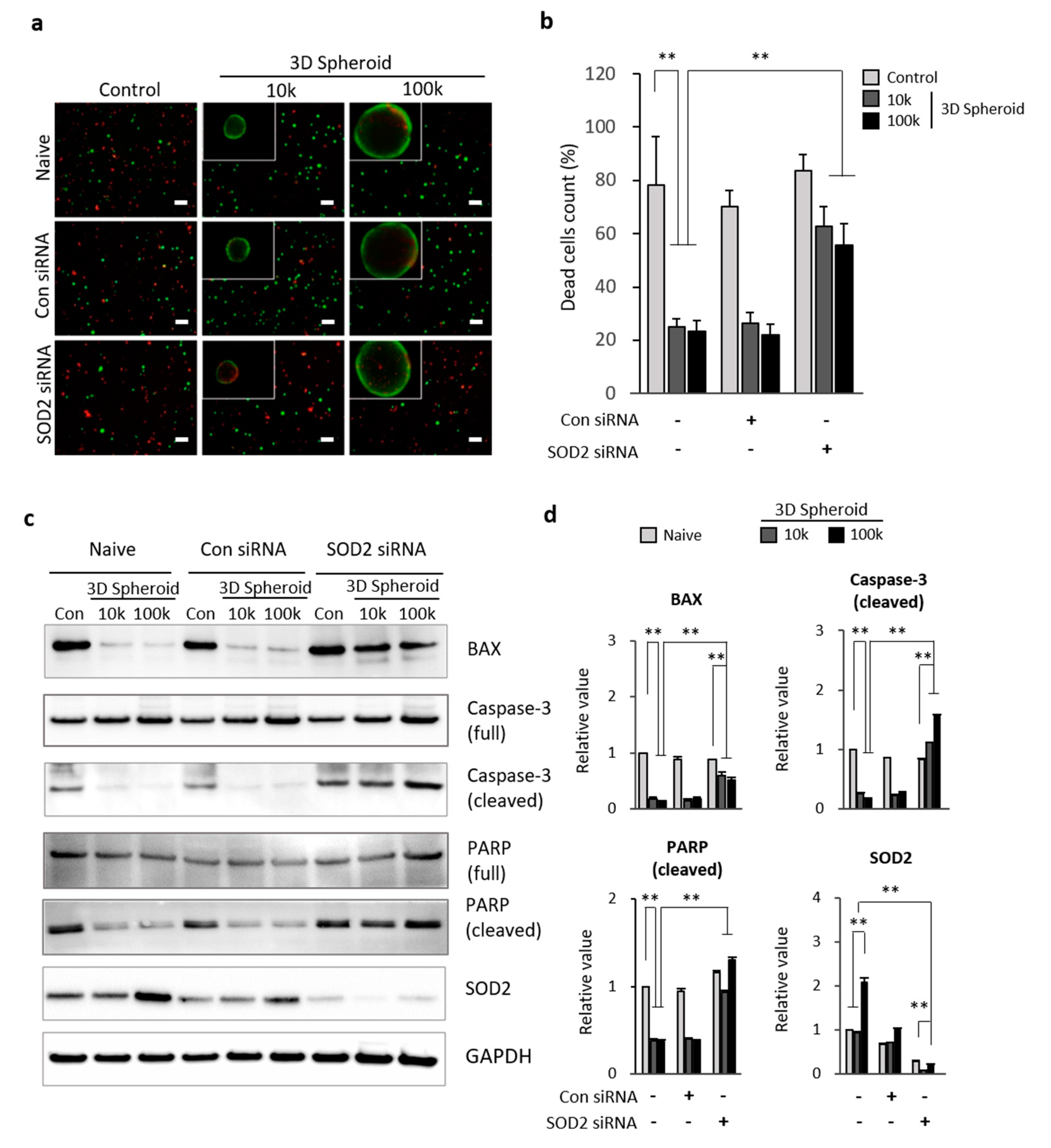
3.3. SOD2 in 3D Spheroid MSCs Related to Apoptosis
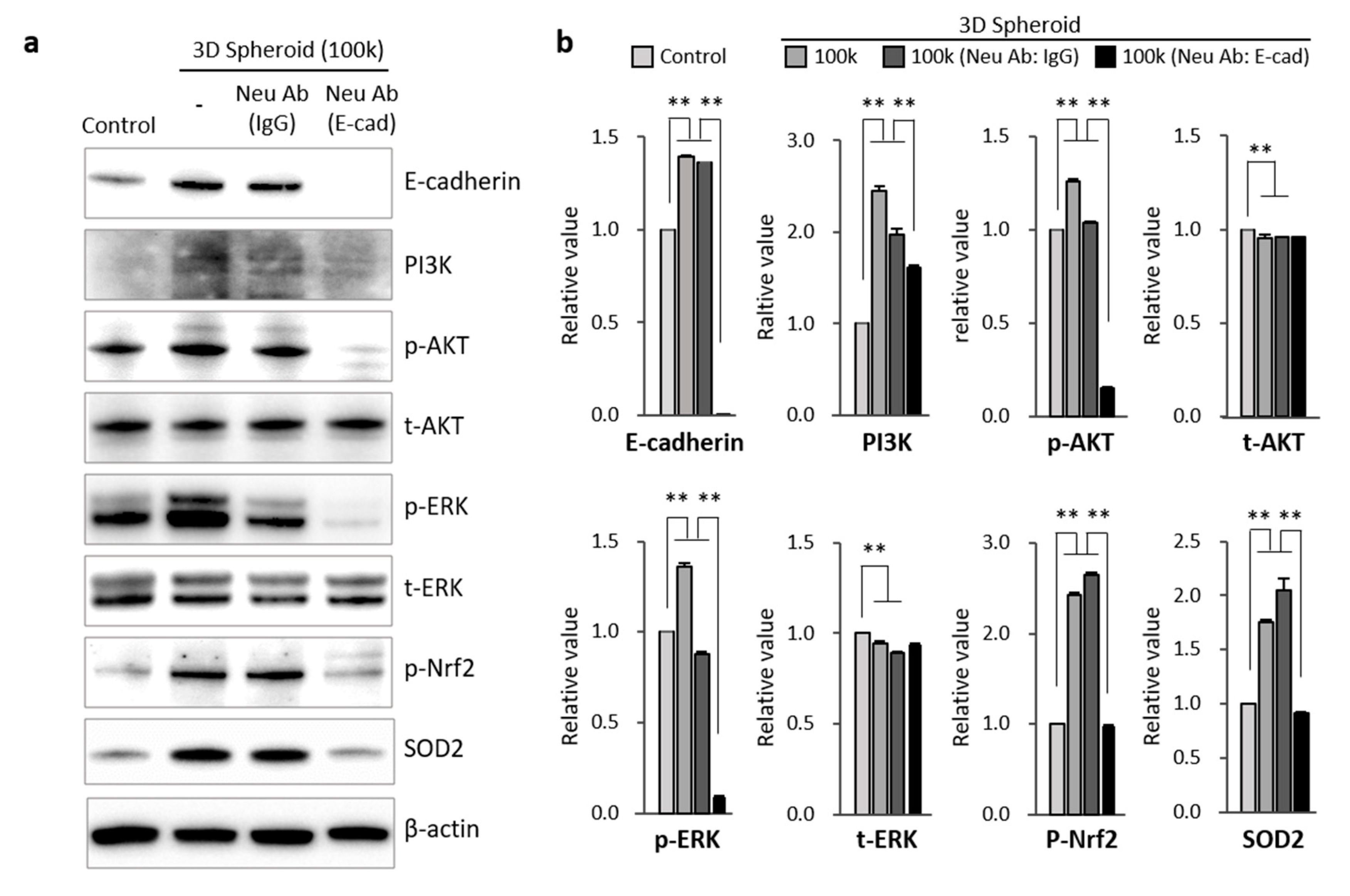
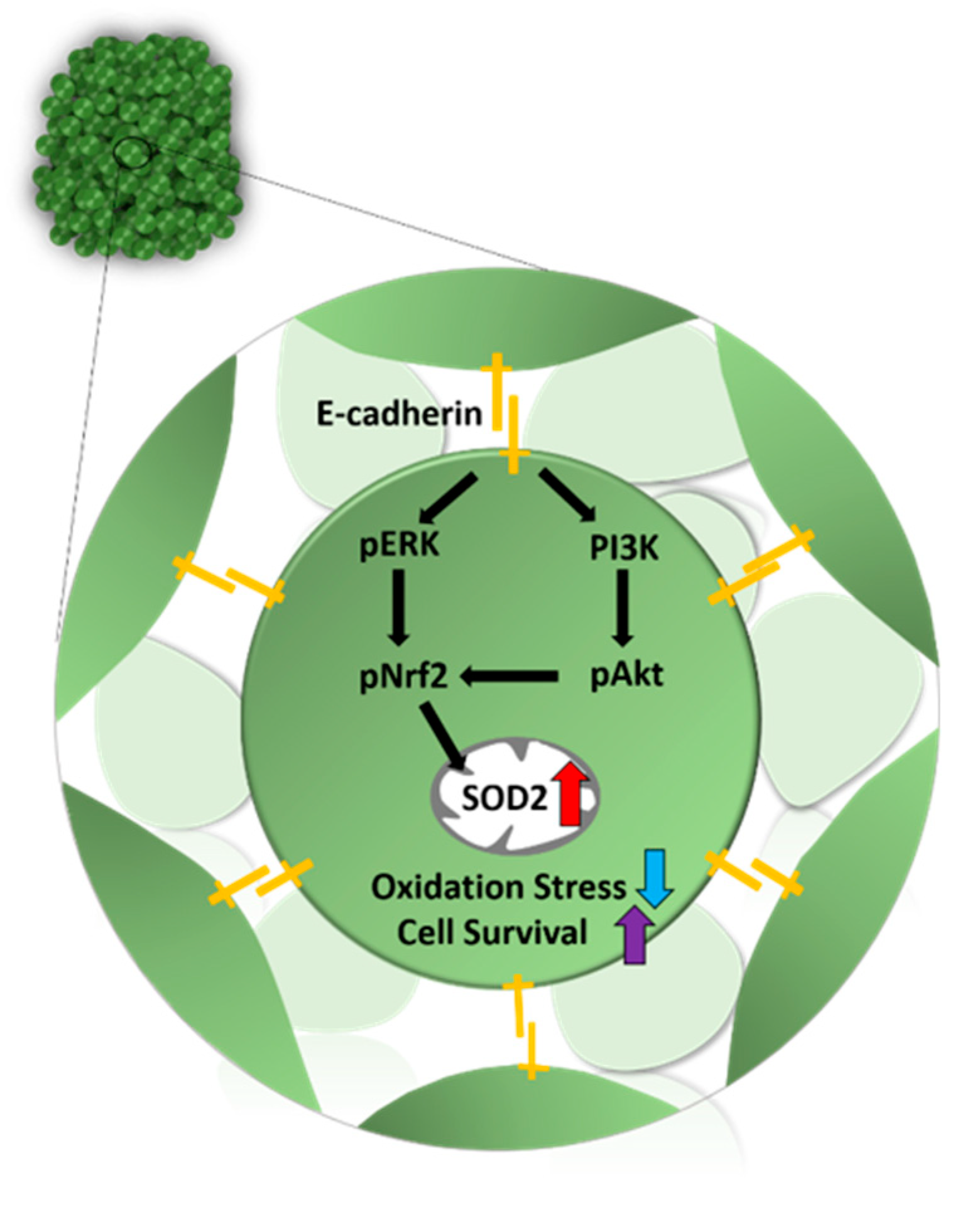
3.4. Mechanism Related to SOD2 Mediated by E-Cadherin in 3D Spheroid Form of MSCs
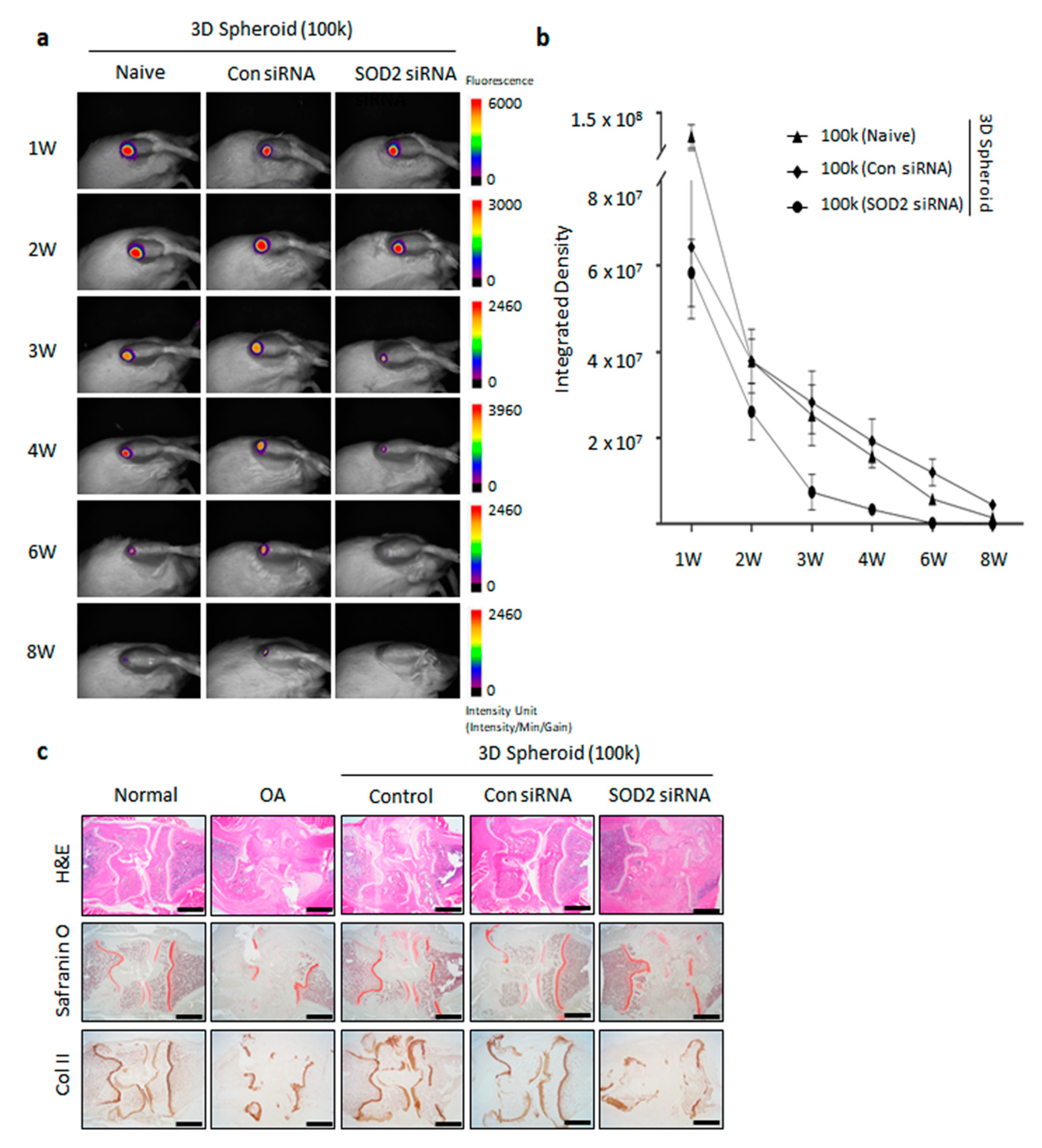
3.5. Enhanced Cartilage Repair in Spheroid-Injected OA Animal Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| UCB-MSC | Umbilical cord blood-derived MSCs |
| 2D | Two-dimensional |
| 3D | Three-dimensional |
| RGD | Arginylglycylaspartic acid |
| ACSC | Adipose-derived stem cells |
| BMSCs | Bone marrow-derived stem cells |
| WJCs | Wharton’s jelly cells |
| SOD2 | Superoxide dismutase 2 |
| MIA | Monosodium iodoacetate |
| OA | Osteoarthritis |
| MNCs | Mononuclear cells |
| SR | Serum replacement |
| PBS | Phosphate-buffered saline |
| DE | Dimensional electrophoresis |
| SDS-PAGE | Sodium dodecylsulfate polyacrylamide gel |
| LC-MS/MS | Liquid chromatography-tandem mass spectrometry |
| MALDI-TOF | Matrix assisted laser desorption ionization time-of-flight |
| SF | Synovial fluid |
| HA | Hyaluronic acid |
| BALF | Bronchoalveolar lavage fluid |
| EMT | Epithelial-mesenchymal transition |
| PI3K | Phosphoinositide 3-kinase |
| Akt | Alpha serine/threonine-protein kinase |
| Nrf | Nuclear respiratory factor |
| ERK | Extracellular signal-regulated kinases |
References
- Ankrum, J.A.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Mahla, R.S. Stem Cells Applications in Regenerative Medicine and Disease Therapeutics. Int. J. Cell Biol. 2016, 2016, 6940283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Han, M.A.; Shin, J.Y.; Jeon, J.H.; Lee, S.J.; Yoon, M.Y.; Kim, H.J.; Choi, E.J.; Do, S.H.; Yang, V.C.; et al. Intra-articular delivery of synovium-resident mesenchymal stem cells via BMP-7-loaded fibrous PLGA scaffolds for cartilage repair. J. Control. Release 2019, 302, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, Z.; Yang, A.Y.; Gao, J.L.; Velez, D.S.; Cho, E.J.; McDermott, D.H.; Murphy, P.M. Mechanisms of Sustained Neutrophilia in Patient WHIM-09, Cured of WHIM Syndrome by Chromothripsis. J. Clin. Immunol. 2018, 38, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Bian, L.; Zhai, D.Y.; Tous, E.; Rai, R.; Mauck, R.L.; Burdick, J.A. Enhanced MSC chondrogenesis following delivery of TGF-beta3 from alginate microspheres within hyaluronic acid hydrogels in vitro and in vivo. Biomaterials 2011, 32, 6425–6434. [Google Scholar] [CrossRef] [Green Version]
- Huey, D.J.; Hu, J.C.; Athanasiou, K.A. Unlike bone, cartilage regeneration remains elusive. Science 2012, 338, 917–921. [Google Scholar] [CrossRef] [Green Version]
- March, L.; Smith, E.U.R.; Hoy, D.G.; Cross, M.J.; Sanchez-Riera, L.; Blyth, F.; Buchbinder, R.; Vos, T.; Woolf, A.D. Burden of disability due to musculoskeletal (MSK) disorders. Best Pract. Res. Clin. Rheumatol. 2014, 28, 353–366. [Google Scholar] [CrossRef]
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Leduc, T.; Hervieu, M.; Legendre, F.; Bouyoucef, M.; Gruchy, N.; Poulain, L.; de Vienne, C.; Herlicoviez, M.; Demoor, M.; Galera, P. Chondrogenic commitment of human umbilical cord blood-derived mesenchymal stem cells in collagen matrices for cartilage engineering. Sci. Rep. 2016, 6, 32786. [Google Scholar] [CrossRef] [Green Version]
- Huey, D.J.; Hu, J.C.; Athanasiou, K.A. Chondrogenically tuned expansion enhances the cartilaginous matrix-forming capabilities of primary, adult, leporine chondrocytes. Cell Transplant. 2013, 22, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Su, P.; Xu, C.; Yang, J.; Yu, W.; Huang, D. Chondrogenic differentiation of human mesenchymal stem cells: A comparison between micromass and pellet culture systems. Biotechnol. Lett. 2010, 32, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Zhu, S.; Tremblay, M.S.; Payette, J.N.; Wang, J.; Bouchez, L.C.; Meeusen, S.; Althage, A.; Cho, C.Y.; Wu, X.; et al. A stem cell-based approach to cartilage repair. Science 2012, 336, 717–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillen, P.; Holley, S.A. Integration of cell-cell and cell-ECM adhesion in vertebrate morphogenesis. Curr. Opin. Cell Biol. 2015, 36, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrandt, C.; Buth, H.; Thielecke, H. A scaffold-free in vitro model for osteogenesis of human mesenchymal stem cells. Tissue Cell 2011, 43, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.C.; Wang, S.; Young, T.H. The influence of spheroid formation of human adipose-derived stem cells on chitosan films on stemness and differentiation capabilities. Biomaterials 2012, 33, 1748–1758. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhou, Y.; Wang, S.; Wu, Y. Epigenetic changes of mesenchymal stem cells in three-dimensional (3D) spheroids. J. Cell. Mol. Med. 2014, 18, 2009–2019. [Google Scholar] [CrossRef]
- Bhang, S.H.; Lee, S.; Shin, J.Y.; Lee, T.J.; Kim, B.S. Transplantation of cord blood mesenchymal stem cells as spheroids enhances vascularization. Tissue Eng. Part A 2012, 18, 2138–2147. [Google Scholar] [CrossRef]
- Cesarz, Z.; Tamama, K. Spheroid Culture of Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 9176357. [Google Scholar] [CrossRef] [Green Version]
- Petrenko, Y.; Sykova, E.; Kubinova, S. The therapeutic potential of three-dimensional multipotent mesenchymal stromal cell spheroids. Stem Cell Res. Ther. 2017, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Bartosh, T.J.; Ylostalo, J.H.; Bazhanov, N.; Kuhlman, J.; Prockop, D.J. Dynamic compaction of human mesenchymal stem/precursor cells into spheres self-activates caspase-dependent IL1 signaling to enhance secretion of modulators of inflammation and immunity (PGE2, TSG6, and STC1). Stem Cells 2013, 31, 2443–2456. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, Y.Y.; Chiang, Y.L.; Wu, C.C.; Lin, S.C. Spheroid formation and neural induction in human adipose-derived stem cells on a chitosan-coated surface. Cells Tissues Organs 2012, 196, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.L.; Ong, H.S.; Zhu, Y.; Liu, S.W.; Liu, L.M.; Zhou, K.H.; Xu, Z.Q.; Gao, J.; Zhang, Y.; Ye, J.H.; et al. In Situ Release of VEGF Enhances Osteogenesis in 3D Porous Scaffolds Engineered with Osterix-Modified Adipose-Derived Stem Cells. Tissue Eng. Part A 2017, 23, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, B.; Lin, S.M.; Hwu, A.T.; Laflin, A.D.; Detamore, M.S. Stem Cells in Aggregate Form to Enhance Chondrogenesis in Hydrogels. PLoS ONE 2015, 10, e0141479. [Google Scholar] [CrossRef] [PubMed]
- Winter, A.; Breit, S.; Parsch, D.; Benz, K.; Steck, E.; Hauner, H.; Weber, R.M.; Ewerbeck, V.; Richter, W. Cartilage-like gene expression in differentiated human stem cell spheroids: A comparison of bone marrow-derived and adipose tissue-derived stromal cells. Arthritis Rheumatol. 2003, 48, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.H.; Bhang, S.H.; Shin, J.Y.; Shin, J.; Kim, B.S. Enhanced cartilage formation via three-dimensional cell engineering of human adipose-derived stem cells. Tissue Eng. Part A 2012, 18, 1949–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baptista, L.S.; Kronemberger, G.S.; Cortes, I.; Charelli, L.E.; Matsui, R.A.M.; Palhares, T.N.; Sohier, J.; Rossi, A.M.; Granjeiro, J.M. Adult Stem Cells Spheroids to Optimize Cell Colonization in Scaffolds for Cartilage and Bone Tissue Engineering. Int. J. Mol. Sci. 2018, 19, 1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Guo, G.; Li, L.; Chen, F.; Bao, J.; Shi, Y.J.; Bu, H. Three-dimensional spheroid culture of human umbilical cord mesenchymal stem cells promotes cell yield and stemness maintenance. Cell Tissue Res. 2015, 360, 297–307. [Google Scholar] [CrossRef]
- Talaei-Khozani, T.; Borhani-Haghighi, M.; Ayatollahi, M.; Vojdani, Z. An in vitro model for hepatocyte-like cell differentiation from Wharton’s jelly derived-mesenchymal stem cells by cell-base aggregates. Gastroenterol. Hepatol. Bed Bench 2015, 8, 188–199. [Google Scholar]
- Lee, E.J.; Park, S.J.; Kang, S.K.; Kim, G.H.; Kang, H.J.; Lee, S.W.; Jeon, H.B.; Kim, H.S. Spherical bullet formation via E-cadherin promotes therapeutic potency of mesenchymal stem cells derived from human umbilical cord blood for myocardial infarction. Mol. Ther. 2012, 20, 1424–1433. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wang, Y.; Zhou, L.; Zou, Y.; Huang, G.; Gao, G.; Ting, S.; Lei, X.; Ding, X. Spheroid-cultured human umbilical cord-derived mesenchymal stem cells attenuate hepatic ischemia-reperfusion injury in rats. Sci. Rep. 2018, 8, 2518. [Google Scholar] [CrossRef]
- Pias, E.K.; Ekshyyan, O.Y.; Rhoads, C.A.; Fuseler, J.; Harrison, L.; Aw, T.Y. Differential effects of superoxide dismutase isoform expression on hydroperoxide-induced apoptosis in PC-12 cells. J. Biol. Chem. 2003, 278, 13294–13301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becuwe, P.; Ennen, M.; Klotz, R.; Barbieux, C.; Grandemange, S. Manganese superoxide dismutase in breast cancer: From molecular mechanisms of gene regulation to biological and clinical significance. Free Radic. Biol. Med. 2014, 77, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Shen, H.; Zhao, M.; Shao, P.; Liu, C.; Cui, J.; Wang, J.; Wang, C.; Guo, N.; Kang, L.; et al. Inflammation-mediated SOD-2 upregulation contributes to epithelial-mesenchymal transition and migration of tumor cells in aflatoxin G1-induced lung adenocarcinoma. Sci. Rep. 2017, 7, 7953. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.H.; Kim, M.; Bae, Y.K.; Kim, G.H.; Choi, S.J.; Oh, W.; Um, S.; Jin, H.J. Decorin Secreted by Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Induces Macrophage Polarization via CD44 to Repair Hyperoxic Lung Injury. Int. J. Mol. Sci. 2019, 20, 4815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyrek, I.; Mathew, B.; Fischer, S.C.; Lissek, S.M.; Becker, S.; Stelzer, E.H.K. E-cadherin, actin, microtubules and FAK dominate different spheroid formation phases and important elements of tissue integrity. Biol. Open 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Capra, J.; Eskelinen, S. Correlation between E-cadherin interactions, survivin expression, and apoptosis in MDCK and ts-Src MDCK cell culture models. Lab. Investig. 2017, 97, 1453–1470. [Google Scholar] [CrossRef]
- Zhang, Q.; Nguyen, A.L.; Shi, S.; Hill, C.; Wilder-Smith, P.; Krasieva, T.B.; Le, A.D. Three-dimensional spheroid culture of human gingiva-derived mesenchymal stem cells enhances mitigation of chemotherapy-induced oral mucositis. Stem Cells Dev. 2012, 21, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Bartosh, T.J.; Ylostalo, J.H.; Mohammadipoor, A.; Bazhanov, N.; Coble, K.; Claypool, K.; Lee, R.H.; Choi, H.; Prockop, D.J. Aggregation of human mesenchymal stromal cells (MSCs) into 3D spheroids enhances their antiinflammatory properties. Proc. Natl. Acad. Sci. USA 2010, 107, 13724–13729. [Google Scholar] [CrossRef] [Green Version]
- Chatterjea, A.; LaPointe, V.L.; Barradas, A.; Garritsen, H.; Yuan, H.; Renard, A.; van Blitterswijk, C.A.; de Beor, J. Cell aggregation enhances bone formation by human mesenchymal stromal cells. Eur. Cell Mater. 2017, 33, 121–129. [Google Scholar] [CrossRef]
- Potapova, I.A.; Gaudette, G.R.; Brink, P.R.; Robinson, R.B.; Rosen, M.R.; Cohen, I.S.; Doronin, S.V. Mesenchymal stem cells support migration, extracellular matrix invasion, proliferation, and survival of endothelial cells in vitro. Stem Cells 2007, 25, 1761–1768. [Google Scholar] [CrossRef]
- Xie, L.; Mao, M.; Zhou, L.; Jiang, B. Spheroid Mesenchymal Stem Cells and Mesenchymal Stem Cell-Derived Microvesicles: Two Potential Therapeutic Strategies. Stem Cells Dev. 2016, 25, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candas, D.; Li, J.J. MnSOD in oxidative stress response-potential regulation via mitochondrial protein influx. Antioxid. Redox Signal. 2014, 20, 1599–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukui, M.; Zhu, B.T. Mitochondrial superoxide dismutase SOD2, but not cytosolic SOD1, plays a critical role in protection against glutamate-induced oxidative stress and cell death in HT22 neuronal cells. Free Radic. Biol. Med. 2010, 48, 821–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.C.; Wang, Y.; Liu, W.; Zhang, X.M.; Fan, M.; Zhao, M. Protective effects of SOD2 overexpression in human umbilical cord mesenchymal stem cells on lung injury induced by acute paraquat poisoning in rats. Life Sci. 2018, 214, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, J.Z.; Wu, Y.J.; Li, W.G. Anti-inflammatory effect of recombinant human superoxide dismutase in rats and mice and its mechanism. Acta Pharmacol. Sin. 2002, 23, 439–444. [Google Scholar]
- Toygar, M.; Aydin, I.; Agilli, M.; Aydin, F.N.; Oztosun, M.; Gul, H.; Macit, E.; Karslioglu, Y.; Topal, T.; Uysal, B.; et al. The relation between oxidative stress, inflammation, and neopterin in the paraquat-induced lung toxicity. Hum. Exp. Toxicol. 2015, 34, 198–204. [Google Scholar] [CrossRef]
- Nagata, S. Apoptosis and Clearance of Apoptotic Cells. Annu. Rev. Immunol. 2018, 36, 489–517. [Google Scholar] [CrossRef]
- van Roy, F.; Berx, G. The cell-cell adhesion molecule E-cadherin. Cell. Mol. Life Sci. 2008, 65, 3756–3788. [Google Scholar] [CrossRef]
- Onder, T.T.; Gupta, P.B.; Mani, S.A.; Yang, J.; Lander, E.S.; Weinberg, R.A. Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer Res. 2008, 68, 3645–3654. [Google Scholar] [CrossRef] [Green Version]
- Aigner, T.; Hemmel, M.; Neureiter, D.; Gebhard, P.M.; Zeiler, G.; Kirchner, T.; McKenna, L. Apoptotic cell death is not a widespread phenomenon in normal aging and osteoarthritis human articular knee cartilage: A study of proliferation, programmed cell death (apoptosis), and viability of chondrocytes in normal and osteoarthritic human knee cartilage. Arthritis Rheumatol. 2001, 44, 1304–1312. [Google Scholar] [CrossRef]
- Kim, H.A.; Blanco, F.J. Cell death and apoptosis in osteoarthritic cartilage. Curr. Drug Targets 2007, 8, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.L.; Gabrielides, C.; Davidson, R.K.; Swingler, T.E.; Clark, I.M.; Wallis, G.A.; Boot-Handford, R.P.; Kirkwood, T.B.; Taylor, R.W.; Young, D.A. Superoxide dismutase downregulation in osteoarthritis progression and end-stage disease. Ann. Rheum Dis. 2010, 69, 1502–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.P.; Chen, Y.W.; Weng, T.I.; Yang, R.S.; Liu, S.H. Arsenic induces human chondrocyte senescence and accelerates rat articular cartilage aging. Arch. Toxicol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.A.; Buckwalter, J.A. Aging, articular cartilage chondrocyte senescence and osteoarthritis. Biogerontology 2002, 3, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Koike, M.; Nojiri, H.; Ozawa, Y.; Watanabe, K.; Muramatsu, Y.; Kaneko, H.; Morikawa, D.; Kobayashi, K.; Saita, Y.; Sasho, T.; et al. Mechanical overloading causes mitochondrial superoxide and SOD2 imbalance in chondrocytes resulting in cartilage degeneration. Sci. Rep. 2015, 5, 11722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Romero, C.; Calamia, V.; Mateos, J.; Carreira, V.; Martinez-Gomariz, M.; Fernandez, M.; Blanco, F.J. Mitochondrial dysregulation of osteoarthritic human articular chondrocytes analyzed by proteomics: A decrease in mitochondrial superoxide dismutase points to a redox imbalance. Mol. Cell. Proteom. 2009, 8, 172–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, M.C.; Brouillette, M.J.; Andresen, N.S.; Oberley-Deegan, R.E.; Martin, J.M. Differential Effects of Superoxide Dismutase Mimetics after Mechanical Overload of Articular Cartilage. Antioxidants 2017, 6, 98. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, H.; Yamamoto, M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef]
- Lacher, S.E.; Lee, J.S.; Wang, X.; Campbell, M.R.; Bell, D.A.; Slattery, M. Beyond antioxidant genes in the ancient Nrf2 regulatory network. Free Radic. Biol. Med. 2015, 88, 452–465. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Wu, X.; Wang, H.; Chen, W.; Zhao, X.; Yang, Y.; Chen, W. Protective Effects Of Astragalus Polysaccharides On Oxidative Stress In High Glucose-Induced Or SOD2-Silenced H9C2 Cells Based On PCR Array Analysis. Diabetes Metab. Syndr. Obes. 2019, 12, 2209–2220. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | Name | MW | PI | Score (MOWSE) | Cov % | Swiss-Prot Assession No | ±Fold/ (Control) |
|---|---|---|---|---|---|---|---|
| 1 | No identification | 4.81 | |||||
| 2 | Guanylyl cyclase-activating protein 1 | 22,920 | 4.3 | 7341 | 29.4 | P43080 | 0.61 (–1.64) |
| 3 | Superoxide dismutase [Mn], mitochondrial | 24,722 | 8.3 | 181 | 71.0 | P04179 | 4.14 |
| 4 | Transgelin-2 | 22,392 | 8.4 | 17,935 | 36.2 | P37802 | 0.64 (–1.56) |
| 5 | No identification | 0.62 (–1.61) | |||||
| 6 | Aldose reductase | 35,854 | 6.5 | 16,298 | 24.4 | P15121 | 1.41 |
| 7 | Myosin regulatory light chain 12A | 19,794 | 4.7 | 10,303 | 40.1 | P19105 | 0.27 (–3.70) |
| 8 | Myosin regulatory light chain 12B | 19,779 | 4.7 | 25,759 | 40.1 | O14950 | 0.41 (–2.44) |
| 9 | Serum albumin (Bovin) | 69,294 | 5.8 | 5,100,000 | 19.4 | P02769 | 2.88 |
| 10 | Cathepsin D | 44,553 | 6.1 | 1,930,000 | 34.5 | P07339 | 2.95 |
| 11 | Glutathione S-transferase omega-1 | 27,566 | 6.2 | 41,480 | 25.7 | P78417 | 0.66 (–1.52) |
| 12 | Ribosomal RNA small subunit methyltransferase NEP1 | 26,720 | 9.3 | 2202 | 27.9 | Q92979 | 0.62 (–1.61) |
| 13 | Guanylyl cyclase-activating protein 1 | 22,920 | 4.3 | 7341 | 29.4 | P43080 | 0.61 (–1.64) |
| 14 | Triosephosphate isomerase | 26,670 | 6.4 | 7037 | 30.5 | P60174 | 1.54 |
| 15 | Myosin regulatory light chain 12A | 19,794 | 4.7 | 11,511 | 34.9 | P19105 | 0.27 (–3.70) |
| 16 | Myosin regulatory light chain 12B | 19,779 | 4.7 | 25,759 | 40.1 | O14950 | 0.45 (–2.22) |
| 17 | No identification | 5.60 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.; Song, B.R.; Kim, D.H.; Ha, J.; Lee, M.; Choi, S.J.; Oh, W.; Um, S.; Jin, H.J. Up-Regulation of Superoxide Dismutase 2 in 3D Spheroid Formation Promotes Therapeutic Potency of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells. Antioxidants 2020, 9, 66. https://doi.org/10.3390/antiox9010066
Lee M, Song BR, Kim DH, Ha J, Lee M, Choi SJ, Oh W, Um S, Jin HJ. Up-Regulation of Superoxide Dismutase 2 in 3D Spheroid Formation Promotes Therapeutic Potency of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells. Antioxidants. 2020; 9(1):66. https://doi.org/10.3390/antiox9010066
Chicago/Turabian StyleLee, Miyoung, Bo Ram Song, Dong Hyun Kim, Jueun Ha, Minju Lee, Soo Jin Choi, Wonil Oh, Soyoun Um, and Hye Jin Jin. 2020. "Up-Regulation of Superoxide Dismutase 2 in 3D Spheroid Formation Promotes Therapeutic Potency of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells" Antioxidants 9, no. 1: 66. https://doi.org/10.3390/antiox9010066