Anti-Microbiological, Anti-Hyperglycemic and Anti-Obesity Potency of Natural Antioxidants in Fruit Fractions of Saskatoon Berry

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Determination of Color
2.3. Determination of Monophosphate Nucleotides
2.4. Determination of Free Amino Acids
2.5. Determination of Polyphenolic Compounds
2.6. Determination of Anti-Microbiological Potency
2.7. Determination of Enzyme Inhibition Potency
2.8. Determination of Antioxidant Activity
2.9. Statistical Analysis
3. Results and Discussion
3.1. Determination of Color
3.2. Determination of Monophosphate Nucleotides
3.3. Determination of Free Amino Acids
3.4. Determination of Polyphenolic Compounds
3.5. Determination of Antibacterial Properties
3.6. Determination of Antioxidant Activity
3.7. Determination of Enzyme Inhibition Properties
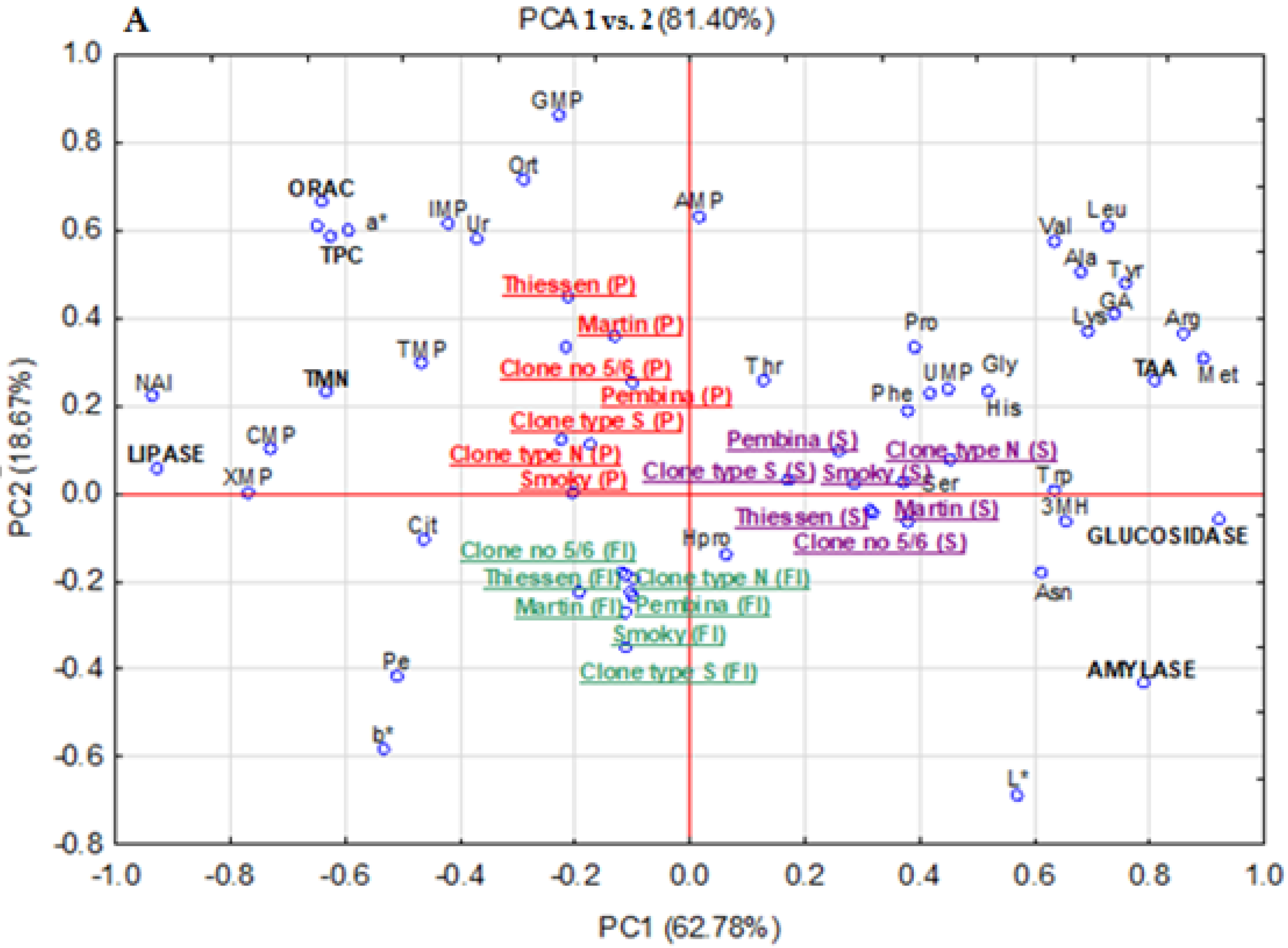
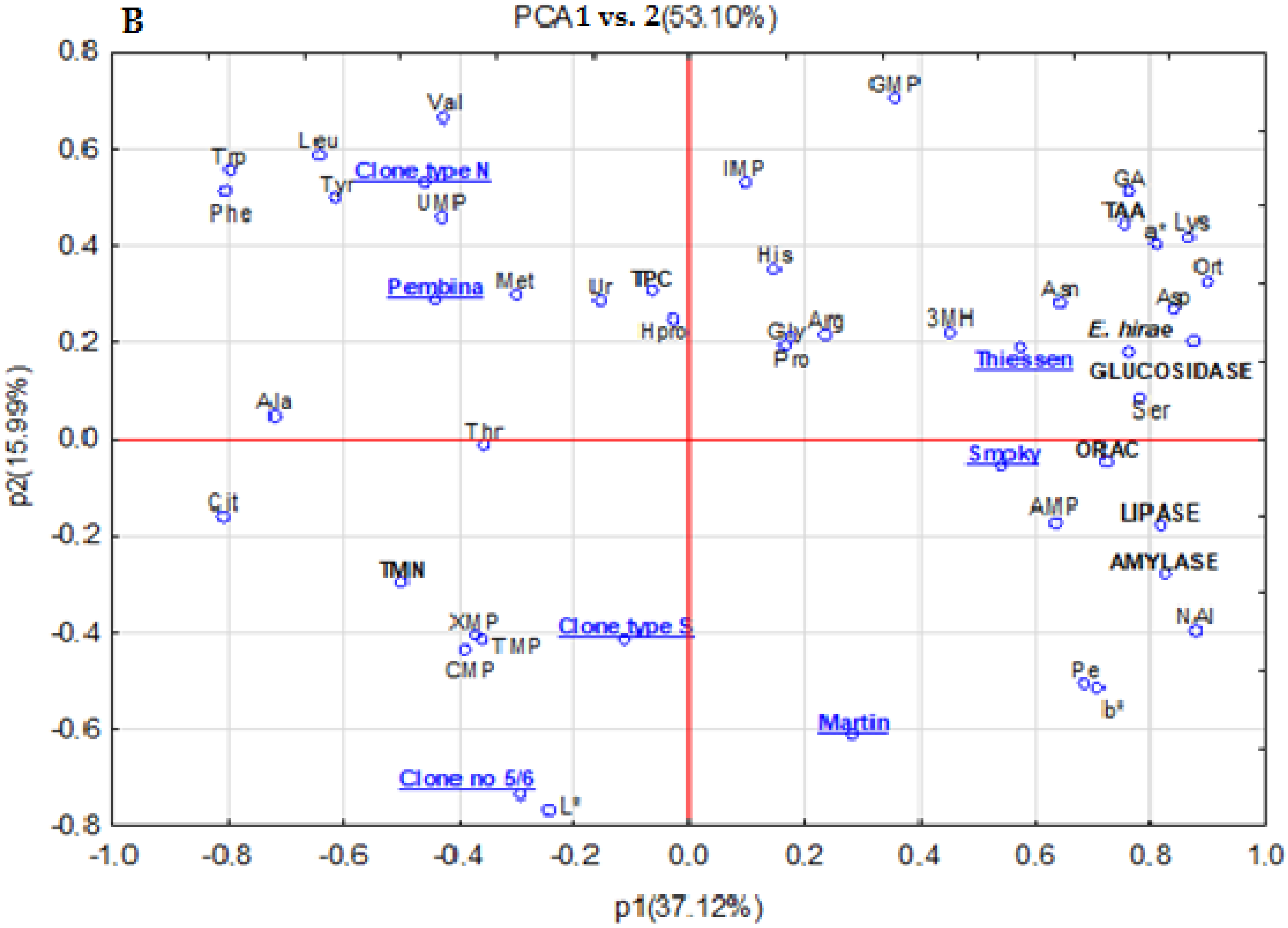
3.8. Principal Components Analysis (PCA)
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Juríková, T.; Balla, S.; Sochor, J.; Pohanka, M.; Mlcek, J.; Baron, M. Flavonoid profile of saskatoon berries (Amelanchier alnifolia Nutt.) and their health promoting effects. Molecules 2013, 18, 12571–12586. [Google Scholar]
- Oz, A.A.; Kafkas, E. Phytochemicals in fruits and vegetables. In Super Food and Functional Food—An Overview of Their Processing and Utilization; BoD–Books on Demand: Norderstedt, Germany, 2017; pp. 175–184. [Google Scholar]
- Escarpa, A.; Gonzalez, M.C. Evaluation of high-performance liquid chromatography for determination of phenolic compounds in pear horticultural cultivars. Chromatographia 2000, 51, 37–43. [Google Scholar] [CrossRef]
- Szakiel, A.; Pączkowski, C.; Huttunen, S. Triterpenoid content of berries and leaves of bilberry Vaccinium myrtillus from Finland and Poland. J. Agric. Food Chem. 2012, 60, 11839–11849. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ding, X. Hypotensive and physiological effect of angiotensin converting enzyme inhibitory peptides derived from soy protein on spontaneously hypertensive rats. J. Agric. Food Chem. 2001, 49, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Drywien, M.M.; Dzwigala, J.; Staszewska-Skurczynska, M. Znaczenie aminokwasów rozgałęzionych w żywieniu człowieka oraz profilaktyce i przebiegu niektórych chorób. Medycyna Ogólna i Nauki o Zdrowiu 2013, 19, 379–384. [Google Scholar]
- Ranogajec, A.; Beluhan, S.; Šmit, Z. Analysis of nucleosides and monophosphate nucleotides from mushrooms with reversed-phase HPLC. J. Sep. Sci. 2010, 33, 1024–1033. [Google Scholar] [CrossRef]
- Moutinho, A.; Hussey, P.P.; Trewavas, A.A.; Malho, R. cAMP acts as a second messenger in pollen tube growth and reorientation. Proc. Natl. Acad. Sci. USA 2001, 98, 10481–10486. [Google Scholar] [CrossRef] [Green Version]
- Baker, D.D.; Kelly, J.J. Structure, function and evolution of microbial adenylyl and guanylyl cyclases. Mol. Microbiol. 2004, 52, 1229–1242. [Google Scholar] [CrossRef]
- Kalia, D.; Merey, G.; Nakayama, S.; Zheng, Y.; Zhou, J.; Luo, Y.; Guo, M.; Roembke, B.T.; Sintim, H.H. Nucleotide, c-di-GMP, c-di-AMP, cGMP, cAMP,(p) ppGpp signaling in bacteria and implications in pathogenesis. Chem. Soc. Rev. 2013, 42, 305–341. [Google Scholar] [CrossRef]
- Lavelli, V.; Harsha, P.P.; Ferranti, P.; Scarafoni, A.; Iametti, S. Grape skin phenolics as inhibitors of mammalian α-glucosidase and α-amylase–effect of food matrix and processing on efficacy. Food Funct. 2016, 7, 1655–1663. [Google Scholar] [CrossRef]
- Podsedek, A.; Majewska, I.; Redzynia, M.; Sosnowska, D.; Koziołkiewicz, M. In vitro inhibitory effect on digestive enzymes and antioxidant potential of commonly consumed fruits. J. Agric. Food Chem. 2014, 62, 4610–4617. [Google Scholar] [CrossRef] [PubMed]
- Unuofin, J.J.; Otunola, G.G.; Afolayan, A.A. In vitro α-amylase, α-glucosidase, lipase inhibitory and cytotoxic activities of tuber extracts of Kedrostis africana (L.) Cogn. Heliyon 2018, 4, e00810. [Google Scholar] [CrossRef] [PubMed]
- De Souza, D.D.; Willems, J.J.; Low, N.N. Phenolic Composition and Antioxidant Activities of Saskatoon Berry Fruit and Pomace. Food Chem. 2019, 290, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Egea, I.; Sánchez-Bel, P.; Romojaro, F.; Pretel, M.M. Six edible wild fruits as potential antioxidant additives or nutritional supplements. Plant Foods Hum. Nutr. 2010, 65, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Lachowicz, S.; Oszmiański, J.; Pluta, S. The composition of bioactive compounds and antioxidant activity of Saskatoon berry (Amelanchier alnifolia Nutt.) genotypes grown in central Poland. Food Chem. 2017, 235, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Lachowicz, S.; Oszmiański, J.; Seliga, Ł.; Pluta, S. Phytochemical composition and antioxidant capacity of seven saskatoon berry (Amelanchier alnifolia Nutt.) Genotypes Grown in Poland. Molecule 2017, 22, 853. [Google Scholar] [CrossRef] [PubMed]
- Ochmian, I.; Oszmiański, J.; Lachowicz, S.; Krupa-Małkiewicz, M. Rootstock effect on physico-chemical properties and content of bioactive compounds of four cultivars Cornelian cherry fruits. Sci. Hortic. 2019, 256, 108588. [Google Scholar] [CrossRef]
- Yamaoka, N.; Kudo, Y.; Inazawa, K.; Inagawa, S.; Yasuda, M.; Mawatari, K.K.; Nakagomi, K.; Kaneko, K. Simultaneous determination of nucleosides and nucleotides in dietary foods and beverages using ion-pairing liquid chromatography–electrospray ionization-mass spectrometry. J. Chromatogr. B 2010, 878, 2054–2060. [Google Scholar] [CrossRef]
- Roucher, V.F.; Desnots, E.; Naël, C.; Agnoux, A.M.; Alexandre-Gouabau, M.C.; Darmaun, D.; Boquien, C.Y. Use of UPLC-ESI-MS/MS to quantitate free amino acid concentrations in micro-samples of mammalian milk. SpringerPlus 2013, 2, 622. [Google Scholar] [CrossRef]
- Armenta, J.M.; Cortes, D.F.; Pisciotta, J.M.; Shuman, J.L.; Blakeslee, K.; Rasoloson, D.; Ogunbiyi, O.; Sullivan, D.J., Jr.; Shulaev, V. Sensitive and rapid method for amino acid quantitation in malaria biological samples using AccQ• Tag ultra performance liquid chromatography-electrospray ionization-MS/MS with multiple reaction monitoring. Anal. Chem. 2009, 82, 548–558. [Google Scholar] [CrossRef]
- Xianggun, G.; Ohlander, M.; Jeppson, N.; Bjork, L. Changes in antioxidant effects and their relationship to phytonutrients in fruits of sea buckthorn during maturation. J. Agric. Food Chem. 2000, 48, 1485–1490. [Google Scholar]
- Nakai, M.; Fukui, Y.; Asami, S.; Toyoda-Ono, Y.; Iwashita, T.; Shibata, H.; Mitsunaga, T.; Hashimoto, F.; Kiso, Y. Inhibitory effects of oolong tea polyphenols on pancreatic lipase in vitro. J. Agric. Food Chem. 2005, 53, 4593–4598. [Google Scholar] [CrossRef] [PubMed]
- Nickavar, B.; Yousefian, N. Evaluation of α-amylase inhibitory activities of selected antidiabetic medicinal plants. J. Verbrauch. Lebensm. 2011, 6, 191–195. [Google Scholar] [CrossRef]
- Kapusta, I.; Cebulak, T.; Oszmiański, J. Characterization of Polish wines produced from the interspecific hybrid grapes grown in south-east Poland. Eur. Food Res. Technol. 2018, 244, 441–455. [Google Scholar] [CrossRef]
- Oszmiański, J.; Wojdylo, A. Aronia melanocarpa phenolics and their antioxidant activity. Eur. Food Res. Technol. 2005, 221, 809–813. [Google Scholar] [CrossRef]
- Pérez-Magariño, S.; González-Sanjosé, M.M. Application of absorbance values used in wineries for estimating CIELAB parameters in red wines. Food Chem. 2003, 81, 301–306. [Google Scholar] [CrossRef]
- Horszwald, A.; Julien, H.; Andlauer, W. Characterisation of Aronia powders obtained by different drying processes. Food Chem. 2013, 141, 2858–2863. [Google Scholar] [CrossRef] [PubMed]
- Lachowicz, S.; Oszmiański, J.; Wiśniewski, R.; Seliga, Ł.; Pluta, S. Chemical parameters profile analysis by liquid chromatography and antioxidative activity of the Saskatoon berry fruits and their components. Eur. Food Res. Technol. 2019, 245, 2007–2015. [Google Scholar] [CrossRef] [Green Version]
- Zielinska, M.; Michalska, A. Microwave-assisted drying of blueberry (Vaccinium corymbosum, L.) fruits: Drying kinetics, polyphenols, anthocyanins, antioxidant capacity, colour and texture. Food Chem. 2016, 212, 671–680. [Google Scholar] [CrossRef]
- Guo, S.; Duan, J.J.; Zhang, Y.; Qian, D.; Tang, Y.; Zhu, Z.; Wang, H. Contents changes of triterpenic acids, nucleosides, nucleobases, and saccharides in Jujube (Ziziphus jujuba) fruit during the drying and steaming process. Molecules 2015, 20, 22329–22340. [Google Scholar] [CrossRef]
- Manninen, H.; Rotola-Pukkila, M.; Aisala, H.; Hopia, A.; Laaksonen, T. Free amino acids and 5′-nucleotides in Finnish forest mushrooms. Food Chem. 2018, 247, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jiang, Y.; Luo, Y.; Jiang, W. A simple and rapid determination of ATP, ADP and AMP concentrations in pericarp tissue of litchi fruit by high performance liquid chromatography. Food Technol. Biotechnol. 2006, 44, 531–534. [Google Scholar]
- Mazza, G. Compositional and functional properties of Saskatoon berry and blueberry. Int. J. Fruit Sci. 2005, 5, 101–120. [Google Scholar] [CrossRef]
- Glew, R.R.; Ayaz, F.F.; Sanz, C.; VanderJagt, D.D.; Huang, H.H.; Chuang, L.L.; Strnad, M. Changes in sugars, organic acids and amino acids in medlar (Mespilus germanica, L.) during fruit development and maturation. Food Chem. 2013, 83, 363–369. [Google Scholar] [CrossRef]
- Silva, B.B.; Andrade, P.P.; Ferreres, F.; Seabra, R.R.; Beatriz, M.; Oliveira, P.P.; Ferreira, M.M. Composition of quince (Cydonia oblonga Miller) seeds: Phenolics, organic acids and free amino acids. Nat. Prod. Res. 2005, 19, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, P.; Cheng, L. Developmental changes of carbohydrates, organic acids, amino acids, and phenolic compounds in ‘Honeycrisp’ apple flesh. Food Chem. 2010, 123, 1013–1018. [Google Scholar] [CrossRef]
- Yang, J.H.; Lin, H.C.; Mau, J.L. Non-volatile taste components of several commercial mushrooms. Food Chem. 2001, 72, 465–471. [Google Scholar] [CrossRef]
- Rop, O.; Mlcek, J.; Jurikova, T.; Sochor, J.; Kizek, R. Antioxidant properties of saskatoon berry (Amelanchier alnifolia Nutt.) fruits. Fruits 2013, 68, 435–444. [Google Scholar] [CrossRef]
- Lavola, A.; Karjalainen, R.; Julkunen-Tiitto, R. Bioactive polyphenols in leaves, stems, and berries of Saskatoon (Amelanchier alnifolia Nutt.) cultivars. J. Agric. Food Chem. 2012, 60, 1020–1027. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, Y.; Cao, L.; Lu, J. Phenolic compounds and antioxidant properties of different grape cultivars grown in China. Food Chem. 2010, 119, 1557–1565. [Google Scholar] [CrossRef]
- Pantelić, M.M.; Zagorac, D.Č.D.; Davidović, S.S.; Todić, S.S.; Bešlić, Z.Z.; Gašić, U.U.; Natić, M.M. Identification and quantification of phenolic compounds in berry skin, pulp, and seeds in 13 grapevine varieties grown in Serbia. Food Chem. 2016, 211, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Inglett, G.E.; Chen, D. Contents of phenolics and flavonoids and antioxidant activities in skin, pulp, and seeds of miracle fruit. J. Food Sci. 2011, 76, C479–C482. [Google Scholar] [CrossRef] [PubMed]
- Puupponen-Pimiä, R.; Nohynek, L.; Meier, C.; Kähkönen, M.; Heinonen, M.; Hopia, A.; Oksman-Caldentey, K.M. Antimicrobial properties of phenolic compounds from berries. J. Appl. Microbiol. 2001, 90, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Puganen, A.; Alakomi, H.H.; Uusitupa, A.; Saarela, M.; Yang, B. Antioxidative and antibacterial activities of aqueous ethanol extracts of berries, leaves, and branches of berry plants. Food Res. Int. 2018, 106, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Y.; Göksel, Z.; Erdoğan, S.S.; Öztürk, A.; Atak, A.; Özer, C. Antioxidant Activity and Phenolic Content of Seed, Skin and Pulp Parts of 22 Grape (Vitis vinifera, L.) Cultivars (4 Common and 18 Registered or Candidate for Registration). J. Food Process. Preserv. 2015, 39, 1682–1691. [Google Scholar] [CrossRef]
- Hu, C.; Kwok, B.H.L.; Kitts, D.D. Saskatoon berries (Amelanchier alnifolia Nutt.) scavenge free radicals and inhibit intracellular oxidation. Food Res. Int. 2005, 38, 1079–1085. [Google Scholar] [CrossRef]
- Wang, J.; Mazza, G. Inhibitory effects of anthocyanins and other phenolic compounds on nitric oxide production in LPS/IFN-γ-activated RAW 264.7 macrophages. J. Agric. Food Chem. 2002, 50, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Burns Kraft, T.T.; Dey, M.; Rogers, R.R.; Ribnicky, D.D.; Gipp, D.D.; Cefalu, W.W.; Lila, M.M. Phytochemical composition and metabolic performance-enhancing activity of dietary berries traditionally used by native North Americans. J. Agric. Food Chem. 2008, 56, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Boath, A.A.; Stewart, D.; McDougall, G.G. Berry components inhibit α-glucosidase in vitro: Synergies between acarbose and polyphenols from black currant and rowanberry. Food Chem. 2012, 135, 929–936. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Fruit Fraction | Genotypes | Parameters of Color | NAI | ||
|---|---|---|---|---|---|
| L* | a* | b* | |||
| Fruit | Thiessen | 32.71 ± 0.03 1 | 33.31 ± 0.03 | 23.54 ± 0.02 | 0.70 ± 0.00 |
| Smoky | 39.74 ± 0.04 | 34.55 ± 0.03 | 24.03 ± 0.02 | 0.71 ± 0.00 | |
| Martin | 42.71 ± 0.04 | 24.25 ± 0.02 | 23.83 ± 0.02 | 0.64 ± 0.00 | |
| Pembina | 33.47 ± 0.03 | 25.22 ± 0.02 | 18.10 ± 0.02 | 0.61 ± 0.00 | |
| Clone no 5/6 | 49.31 ± 0.04 | 18.33 ± 0.02 | 24.06 ± 0.02 | 0.65 ± 0.00 | |
| Clone type S | 45.76 ± 0.04 | 21.92 ± 0.02 | 21.98 ± 0.02 | 0.65 ± 0.00 | |
| Clone type N | 39.01 ± 0.04 | 23.53 ± 0.02 | 19.71 ± 0.02 | 0.57 ± 0.00 | |
| Flesh | Thiessen | 55.54 ± 0.05 | 9.67 ± 0.01 | 30.73 ± 0.03 | 0.57 ± 0.00 |
| Smoky | 51.41 ± 0.05 | 17.62 ± 0.02 | 40.74 ± 0.04 | 0.58 ± 0.00 | |
| Martin | 55.26 ± 0.05 | 12.37 ± 0.01 | 40.41 ± 0.04 | 0.52 ± 0.00 | |
| Pembina | 43.30 ± 0.04 | 12.86 ± 0.01 | 30.69 ± 0.03 | 0.50 ± 0.00 | |
| Clone no 5/6 | 63.80 ± 0.06 | 9.35 ± 0.01 | 40.80 ± 0.04 | 0.53 ± 0.00 | |
| Clone type S | 59.20 ± 0.05 | 11.18 ± 0.01 | 37.27 ± 0.03 | 0.54 ± 0.00 | |
| Clone type N | 50.47 ± 0.05 | 12.00 ± 0.01 | 33.42 ± 0.03 | 0.62 ± 0.00 | |
| Peel | Thiessen | 19.07 ± 0.02 | 46.36 ± 0.04 | 26.85 ± 0.02 | 0.83 ± 0.00 |
| Smoky | 23.17 ± 0.02 | 48.09 ± 0.04 | 29.82 ± 0.03 | 0.85 ± 0.00 | |
| Martin | 24.90 ± 0.02 | 33.75 ± 0.03 | 29.57 ± 0.03 | 0.76 ± 0.00 | |
| Pembina | 19.51 ± 0.02 | 35.10 ± 0.03 | 22.46 ± 0.02 | 0.73 ± 0.00 | |
| Clone no 5/6 | 28.75 ± 0.03 | 25.51 ± 0.02 | 29.86 ± 0.03 | 0.77 ± 0.00 | |
| Clone type S | 26.68 ± 0.02 | 30.51 ± 0.03 | 27.28 ± 0.02 | 0.78 ± 0.00 | |
| Clone type N | 23.38 ± 0.02 | 48.07 ± 0.04 | 24.46 ± 0.02 | 0.83 ± 0.00 | |
| Seeds | Thiessen | 51.26 ± 0.05 | 11.88 ± 0.01 | 20.96 ± 0.02 | 0.00 ± 0.00 |
| Smoky | 56.70 ± 0.05 | 12.72 ± 0.01 | 26.43 ± 0.02 | 0.00 ± 0.00 | |
| Martin | 60.94 ± 0.05 | 8.92 ± 0.01 | 26.21 ± 0.02 | 0.00 ± 0.00 | |
| Pembina | 47.76 ± 0.04 | 9.28 ± 0.01 | 19.91 ± 0.02 | 0.00 ± 0.00 | |
| Clone no 5/6 | 70.36 ± 0.06 | 6.75 ± 0.01 | 26.46 ± 0.02 | 0.00 ± 0.00 | |
| Clone type S | 65.29 ± 0.06 | 8.07 ± 0.01 | 24.18 ± 0.02 | 0.00 ± 0.00 | |
| Clone type N | 55.66 ± 0.05 | 8.66 ± 0.01 | 21.68 ± 0.02 | 0.00 ± 0.00 | |
| Fruit 3 | 40.39 c2 | 25.87 b | 22.18 d | 0.65 b | |
| Flesh | 54.14 b | 12.15 c | 36.29 a | 0.55 c | |
| Peel | 23.64 d | 38.20 a | 27.19 b | 0.79 a | |
| Seeds | 58.28 a | 9.47 d | 23.69 c | 0.00 d | |
| Fruit Fraction | Genotypes | Monophosphate Nucleotides * | |||||||
|---|---|---|---|---|---|---|---|---|---|
| IMP | UMP | CMP | GMP | XMP | TMP | AMP | SUM | ||
| Fruit | Thiessen | 1.04 ± 0.01 1 | 5.13 ± 0.04 | 9.04 ± 0.07 | 1.25 ± 0.01 | 10.66 ± 0.09 | 1.74 ± 0.01 | 1.36 ± 0.01 | 30.22 ± 0.24 |
| Smoky | 1.06 ± 0.01 | 4.09 ± 0.03 | 10.69 ± 0.09 | 0.72 ± 0.01 | 13.39 ± 0.11 | 1.16 ± 0.01 | 0.98 ± 0.01 | 32.09 ± 0.26 | |
| Martin | 1.25 ± 0.01 | 4.68 ± 0.04 | 10.81 ± 0.09 | 1.66 ± 0.01 | 12.42 ± 0.10 | 1.33 ± 0.01 | 1.64 ± 0.01 | 33.80 ± 0.27 | |
| Pembina | 0.55 ± 0.00 | 4.14 ± 0.03 | 4.10 ± 0.03 | 0.52 ± 0.00 | 4.81 ± 0.04 | 0.69 ± 0.01 | 0.74 ± 0.01 | 15.55 ± 0.12 | |
| Clone no 5/6 | 1.07 ± 0.01 | 4.90 ± 0.04 | 10.91 ± 0.09 | 1.28 ± 0.01 | 12.31 ± 0.10 | 1.57 ± 0.01 | 1.23 ± 0.01 | 33.27 ± 0.27 | |
| Clone type S | 0.53 ± 0.00 | 2.53 ± 0.02 | 8.88 ± 0.07 | 0.50 ± 0.00 | 10.12 ± 0.08 | 0.73 ± 0.01 | 0.77 ± 0.01 | 24.06 ± 0.19 | |
| Clone type N | 0.77 ± 0.01 | 2.93 ± 0.02 | 7.58 ± 0.06 | 0.85 ± 0.01 | 9.03 ± 0.07 | 0.83 ± 0.01 | 0.92 ± 0.01 | 22.92 ± 0.18 | |
| Flesh | Thiessen | 0.57 ± 0.00 | 6.03 ± 0.05 | 7.68 ± 0.06 | 0.97 ± 0.01 | 8.59 ± 0.07 | 0.92 ± 0.01 | 1.18 ± 0.01 | 25.93 ± 0.21 |
| Smoky | 0.89 ± 0.01 | 4.39 ± 0.04 | 10.69 ± 0.09 | 1.34 ± 0.01 | 10.92 ± 0.1 | 1.73 ± 0.01 | 1.00 ± 0.01 | 30.96 ± 0.26 | |
| Martin | 1.02 ± 0.01 | 3.80 ± 0.01 | 3.18 ± 0.01 | 0.57 ± 0.00 | 2.26 ± 0.01 | 0.67 ± 0.01 | 0.72 ± 0.01 | 12.22 ± 0.05 | |
| Pembina | 1.07 ± 0.01 | 4.01 ± 0.03 | 2.78 ± 0.10 | 1.77 ± 0.01 | 3.89 ± 0.11 | 1.33 ± 0.01 | 1.20 ± 0.01 | 16.05 ± 0.29 | |
| Clone no 5/6 | 0.47 ± 0.00 | 3.55 ± 0.03 | 4.85 ± 0.04 | 0.55 ± 0.00 | 5.38 ± 0.04 | 0.69 ± 0.01 | 0.54 ± 0.00 | 16.03 ± 0.13 | |
| Clone type S | 1.04 ± 0.01 | 5.06 ± 0.04 | 11.76 ± 0.09 | 1.27 ± 0.01 | 7.08 ± 0.10 | 1.59 ± 0.01 | 0.95 ± 0.01 | 28.75 ± 0.28 | |
| Clone type N | 0.50 ± 0.00 | 4.52 ± 0.00 | 7.81 ± 0.01 | 0.39 ± 0.00 | 0.82 ± 0.01 | 0.77 ± 0.00 | 0.63 ± 0.01 | 15.49 ± 0.03 | |
| Peel | Thiessen | 0.76 ± 0.01 | 3.03 ± 0.02 | 8.17 ± 0.07 | 0.84 ± 0.01 | 9.59 ± 0.08 | 0.84 ± 0.01 | 0.72 ± 0.01 | 23.94 ± 0.19 |
| Smoky | 0.56 ± 0.00 | 6.22 ± 0.05 | 8.27 ± 0.07 | 0.96 ± 0.01 | 9.12 ± 0.07 | 0.93 ± 0.01 | 0.91 ± 0.01 | 26.98 ± 0.22 | |
| Martin | 0.97 ± 0.01 | 4.80 ± 0.04 | 11.21 ± 0.09 | 1.17 ± 0.01 | 12.53 ± 0.1 | 2.81 ± 0.02 | 1.21 ± 0.01 | 34.70 ± 0.28 | |
| Pembina | 1.02 ± 0.01 | 2.80 ± 0.01 | 5.18 ± 0.01 | 0.57 ± 0.00 | 7.26 ± 0.01 | 0.67 ± 0.01 | 0.72 ± 0.01 | 18.22 ± 0.05 | |
| Clone no 5/6 | 1.17 ± 0.01 | 4.38 ± 0.04 | 13.39 ± 0.11 | 1.55 ± 0.01 | 14.59 ± 0.12 | 2.16 ± 0.02 | 1.46 ± 0.01 | 38.71 ± 0.31 | |
| Clone type S | 0.52 ± 0.00 | 3.88 ± 0.03 | 5.09 ± 0.04 | 0.48 ± 0.00 | 5.65 ± 0.05 | 1.11 ± 0.01 | 0.65 ± 0.01 | 17.38 ± 0.14 | |
| Clone type N | 0.83 ± 0.01 | 4.78 ± 0.04 | 10.99 ± 0.09 | 1.03 ± 0.01 | 12.02 ± 0.1 | 1.29 ± 0.01 | 0.52 ± 0.00 | 31.45 ± 0.25 | |
| Seeds | Thiessen | 1.02 ± 0.01 | 4.80 ± 0.01 | 10.18 ± 0.01 | 0.57 ± 0.00 | 9.26 ± 0.01 | 0.67 ± 0.01 | 0.72 ± 0.01 | 33.44 ± 0.05 |
| Smoky | 1.00 ± 0.01 | 4.36 ± 0.03 | 13.13 ± 0.11 | 1.37 ± 0.01 | 14.01 ± 0.11 | 0.99 ± 0.01 | 0.62 ± 0.00 | 35.47 ± 0.28 | |
| Martin | 0.44 ± 0.00 | 3.86 ± 0.03 | 4.99 ± 0.04 | 0.43 ± 0.00 | 5.42 ± 0.04 | 0.51 ± 0.00 | 0.28 ± 0.00 | 15.92 ± 0.13 | |
| Pembina | 0.95 ± 0.01 | 0.00 ± 0.00 | 9.29 ± 0.07 | 1.18 ± 0.01 | 10.66 ± 0.09 | 1.98 ± 0.02 | 0.52 ± 0.00 | 24.59 ± 0.20 | |
| Clone no 5/6 | 0.47 ± 0.00 | 6.62 ± 0.05 | 6.71 ± 0.05 | 0.63 ± 0.01 | 7.66 ± 0.06 | 1.39 ± 0.01 | 0.54 ± 0.00 | 24.02 ± 0.19 | |
| Clone type S | 0.84 ± 0.01 | 3.56 ± 0.03 | 6.93 ± 0.06 | 1.08 ± 0.01 | 8.56 ± 0.07 | 1.31 ± 0.01 | 0.49 ± 0.00 | 22.77 ± 0.18 | |
| Clone type N | 0.77 ± 0.01 | 5.59 ± 0.04 | 3.86 ± 0.03 | 1.15 ± 0.01 | 4.26 ± 0.03 | 0.69 ± 0.01 | 1.46 ± 0.01 | 17.77 ± 0.14 | |
| Fruit 3 | 0.97 b2 | 4.15 b | 10.56 a | 1.22 b | 11.88 a | 1.81 a | 0.97 b | 31.56 a | |
| Flesh | 0.80 c | 2.31 d | 4.38 d | 0.57 d | 5.11 d | 0.82 c | 0.73 d | 14.71 d | |
| Peel | 0.98 a | 3.85 c | 10.40 b | 1.30 a | 11.73 b | 1.26 b | 1.01 a | 30.52 b | |
| Seeds | 0.56 d | 4.75 a | 5.55 c | 0.72 c | 6.18 c | 0.79 d | 0.82 c | 19.37 c | |
| Fruit Fraction | Genotypes | Arg | His | Met | Lys | Leu | Thr | Val | Phe | PE | Trp | Ur | 3MH |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fruit | Thiessen | 8.10 ± 0.06 1 | 0.63 ± 0.01 | 0.02 ± 0.00 | 0.69 ± 0.01 | 0.28 ± 0.00 | 0.01 ± 0.00 | 0.14 ± 0.00 | 2.80 ± 0.02 | 0.65 ± 0.01 | 0.32 ± 0.00 | 0.07 ± 0.00 | 0.12 ± 0.00 |
| Smoky | 3.25 ± 0.03 | 0.99 ± 0.01 | 0.02 ± 0.00 | 0.32 ± 0.00 | 0.09 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 3.48 ± 0.03 | 0.85 ± 0.01 | 0.33 ± 0.00 | 0.07 ± 0.00 | 0.08 ± 0.00 | |
| Martin | 8.65 ± 0.07 | 1.23 ± 0.01 | 0.04 ± 0.00 | 0.66 ± 0.01 | 0.44 ± 0.00 | 0.01 ± 0.00 | 0.29 ± 0.00 | 3.95 ± 0.03 | 0.57 ± 0.00 | 0.24 ± 0.00 | 0.14 ± 0.00 | 0.04 ± 0.00 | |
| Pembina | 9.15 ± 0.15 | 2.61 ± 0.02 | 0.11 ± 0.00 | 0.90 ± 0.01 | 0.57 ± 0.00 | 0.01 ± 0.00 | 0.37 ± 0.00 | 6.24 ± 0.05 | 0.43 ± 0.00 | 3.88 ± 0.03 | 0.07 ± 0.00 | 0.57 ± 0.00 | |
| Clone no 5/6 | 8.66 ± 0.07 | 2.23 ± 0.02 | 0.02 ± 0.00 | 0.57 ± 0.00 | 0.23 ± 0.00 | 0.01 ± 0.00 | 0.22 ± 0.00 | 3.38 ± 0.03 | 0.73 ± 0.01 | 0.56 ± 0.00 | 0.07 ± 0.00 | 0.28 ± 0.00 | |
| Clone type S | 5.92 ± 0.01 | 0.60 ± 0.00 | 0.02 ± 0.00 | 0.20 ± 0.00 | 0.15 ± 0.00 | 0.01 ± 0.00 | 0.13 ± 0.00 | 1.85 ± 0.01 | 0.64 ± 0.01 | 0.11 ± 0.00 | 0.10 ± 0.00 | 0.02 ± 0.00 | |
| Clone type N | 5.19 ± 0.01 | 0.25 ± 0.00 | 0.02 ± 0.00 | 0.26 ± 0.00 | 0.14 ± 0.00 | 0.01 ± 0.00 | 0.11 ± 0.00 | 1.83 ± 0.01 | 0.55 ± 0.00 | 0.15 ± 0.00 | 0.07 ± 0.00 | 0.03 ± 0.00 | |
| Flesh | Thiessen | 12.52 ± 0.1 | 5.29 ± 0.01 | 0.10 ± 0.00 | 1.35 ± 0.01 | 0.64 ± 0.01 | 0.02 ± 0.00 | 0.36 ± 0.00 | 9.35 ± 0.07 | 0.33 ± 0.00 | 1.11 ± 0.01 | 0.07 ± 0.00 | 0.22 ± 0.00 |
| Smoky | 4.66 ± 0.04 | 3.31 ± 0.03 | 0.02 ± 0.00 | 0.47 ± 0.00 | 0.34 ± 0.00 | 0.01 ± 0.00 | 0.33 ± 0.00 | 4.52 ± 0.04 | 0.60 ± 0.00 | 1.06 ± 0.01 | 0.07 ± 0.00 | 0.36 ± 0.00 | |
| Martin | 2.87 ± 0.01 | 5.25 ± 0.04 | 0.02 ± 0.00 | 0.22 ± 0.00 | 0.11 ± 0.00 | 0.01 ± 0.00 | 0.12 ± 0.00 | 5.61 ± 0.04 | 0.79 ± 0.01 | 1.09 ± 0.01 | 0.07 ± 0.00 | 0.23 ± 0.00 | |
| Pembina | 4.98 ± 0.04 | 6.49 ± 0.05 | 0.03 ± 0.00 | 0.45 ± 0.00 | 0.54 ± 0.00 | 0.01 ± 0.00 | 0.71 ± 0.01 | 6.38 ± 0.05 | 0.53 ± 0.00 | 0.8 ± 0.01 | 0.13 ± 0.00 | 0.13 ± 0.00 | |
| Clone no 5/6 | 3.02 ± 0.09 | 4.82 ± 0.11 | 0.10 ± 0.00 | 0.62 ± 0.00 | 0.70 ± 0.01 | 0.01 ± 0.00 | 0.61 ± 0.01 | 3.61 ± 0.03 | 0.39 ± 0.00 | 1.76 ± 0.01 | 0.07 ± 0.00 | 1.70 ± 0.01 | |
| Clone type S | 5.40 ± 0.04 | 3.28 ± 0.01 | 0.03 ± 0.00 | 0.38 ± 0.00 | 0.46 ± 0.00 | 0.01 ± 0.00 | 0.27 ± 0.00 | 5.49 ± 0.04 | 0.53 ± 0.00 | 1.67 ± 0.01 | 0.08 ± 0.00 | 0.10 ± 0.00 | |
| Clone type N | 9.17 ± 0.02 | 4.03 ± 0.02 | 0.03 ± 0.00 | 0.18 ± 0.00 | 0.15 ± 0.00 | 0.01 ± 0.00 | 0.22 ± 0.00 | 6.81 ± 0.05 | 0.69 ± 0.01 | 1.73 ± 0.01 | 0.08 ± 0.00 | 0.06 ± 0.00 | |
| Peel | Thiessen | 8.76 ± 0.05 | 2.51 ± 0.02 | 0.05 ± 0.00 | 0.36 ± 0.00 | 0.73 ± 0.01 | 0.02 ± 0.00 | 0.58 ± 0.00 | 7.75 ± 0.06 | 0.47 ± 0.00 | 1.27 ± 0.01 | 0.15 ± 0.00 | 0.04 ± 0.00 |
| Smoky | 9.75 ± 0.10 | 4.34 ± 0.04 | 0.05 ± 0.00 | 0.49 ± 0.00 | 0.94 ± 0.01 | 0.02 ± 0.00 | 0.75 ± 0.01 | 4.38 ± 0.04 | 0.35 ± 0.00 | 2.78 ± 0.02 | 0.08 ± 0.00 | 0.47 ± 0.00 | |
| Martin | 7.05 ± 0.05 | 1.05 ± 0.01 | 0.02 ± 0.00 | 0.26 ± 0.00 | 0.34 ± 0.00 | 0.02 ± 0.00 | 0.18 ± 0.00 | 4.14 ± 0.03 | 0.68 ± 0.01 | 0.80 ± 0.01 | 0.07 ± 0.00 | 0.11 ± 0.00 | |
| Pembina | 4.43 ± 0.02 | 2.67 ± 0.01 | 0.02 ± 0.00 | 0.12 ± 0.00 | 0.11 ± 0.00 | 0.01 ± 0.00 | 0.10 ± 0.00 | 5.14 ± 0.04 | 0.90 ± 0.01 | 0.83 ± 0.01 | 0.07 ± 0.00 | 0.07 ± 0.00 | |
| Clone no 5/6 | 6.46 ± 0.05 | 2.06 ± 0.02 | 0.04 ± 0.00 | 0.25 ± 0.00 | 0.54 ± 0.00 | 0.02 ± 0.00 | 0.38 ± 0.00 | 5.84 ± 0.05 | 0.60 ± 0.00 | 0.61 ± 0.00 | 0.13 ± 0.00 | 0.04 ± 0.00 | |
| Clone type S | 9.29 ± 0.11 | 4.39 ± 0.04 | 0.10 ± 0.00 | 0.34 ± 0.00 | 0.69 ± 0.01 | 0.02 ± 0.00 | 0.49 ± 0.00 | 9.23 ± 0.07 | 0.45 ± 0.00 | 9.77 ± 0.08 | 0.07 ± 0.00 | 0.51 ± 0.00 | |
| Clone type N | 8.08 ± 0.06 | 0.86 ± 0.01 | 0.02 ± 0.00 | 0.38 ± 0.00 | 0.17 ± 0.00 | 0.02 ± 0.00 | 0.16 ± 0.00 | 3.47 ± 0.03 | 0.57 ± 0.00 | 0.65 ± 0.01 | 0.07 ± 0.00 | 0.14 ± 0.00 | |
| Seeds | Thiessen | 8.84 ± 0.02 | 1.36 ± 0.01 | 0.02 ± 0.00 | 1.18 ± 0.00 | 0.06 ± 0.00 | 0.02 ± 0.00 | 0.10 ± 0.00 | 4.31 ± 0.03 | 0.75 ± 0.01 | 1.68 ± 0.01 | 0.07 ± 0.00 | 0.09 ± 0.00 |
| Smoky | 7.56 ± 0.06 | 1.69 ± 0.01 | 0.03 ± 0.00 | 0.36 ± 0.00 | 0.27 ± 0.00 | 0.02 ± 0.00 | 0.34 ± 0.00 | 4.90 ± 0.04 | 0.51 ± 0.00 | 1.50 ± 0.00 | 0.13 ± 0.00 | 0.05 ± 0.00 | |
| Martin | 7.56 ± 0.06 | 1.69 ± 0.01 | 0.03 ± 0.00 | 0.36 ± 0.00 | 0.27 ± 0.00 | 0.02 ± 0.00 | 0.34 ± 0.00 | 4.90 ± 0.04 | 0.51 ± 0.00 | 0.50 ± 0.00 | 0.13 ± 0.00 | 0.05 ± 0.00 | |
| Pembina | 7.19 ± 0.07 | 1.81 ± 0.01 | 0.03 ± 0.00 | 0.38 ± 0.00 | 0.48 ± 0.00 | 0.01 ± 0.00 | 0.35 ± 0.00 | 6.27 ± 0.05 | 0.52 ± 0.00 | 1.88 ± 0.02 | 0.07 ± 0.00 | 0.16 ± 0.00 | |
| Clone no 5/6 | 3.40 ± 0.01 | 0.19 ± 0.00 | 0.02 ± 0.00 | 0.22 ± 0.00 | 0.20 ± 0.00 | 0.01 ± 0.00 | 0.14 ± 0.00 | 1.80 ± 0.01 | 0.33 ± 0.00 | 0.33 ± 0.00 | 0.08 ± 0.00 | 0.02 ± 0.00 | |
| Clone type S | 5.29 ± 0.01 | 0.12 ± 0.00 | 0.02 ± 0.00 | 0.22 ± 0.00 | 0.26 ± 0.00 | 0.01 ± 0.00 | 0.13 ± 0.00 | 1.76 ± 0.01 | 0.32 ± 0.00 | 0.31 ± 0.00 | 0.07 ± 0.00 | 0.03 ± 0.00 | |
| Clone type N | 8.82 ± 0.17 | 1.50 ± 0.01 | 0.17 ± 0.00 | 1.25 ± 0.02 | 1.00 ± 0.01 | 0.02 ± 0.00 | 0.68 ± 0.01 | 5.00 ± 0.04 | 0.42 ± 0.00 | 1.12 ± 0.02 | 0.07 ± 0.00 | 0.18 ± 0.00 | |
| Fruit 3 | 7.02 b2 | 1.59 d | 0.02 c | 0.45 b | 0.33 c | 0.01 a | 0.23 c | 4.30 c | 0.61 b | 0.99 b | 0.07 d | 0.18 b | |
| Flesh | 2.13 d | 1.64 c | 0.02 c | 0.20 d | 0.12 d | 0.01 a | 0.05 d | 4.14 d | 0.71 a | 0.73 c | 0.08 c | 0.08 c | |
| Peel | 5.13 c | 2.05 b | 0.03 b | 0.37 c | 0.42 b | 0.02 a | 0.36 b | 4.63 b | 0.51 c | 0.55 d | 0.12 a | 0.05 d | |
| Seeds | 7.95 a | 4.38 a | 0.11 a | 0.90 a | 0.69 a | 0.02 a | 0.56 a | 6.10 a | 0.41 d | 1.04 a | 0.08 b | 0.53 a | |
| Fruit Fraction | Genotypes | Glu | Asp | Pro | Hpro | Ser | Gly | Ala | Tyr | Cit | Ort | Asn | Arg |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fruit | Thiessen | 5.62 ± 0.04 | 13.37 ± 0.11 | 1.37 ± 0.01 | 0.00 ± 0.00 | 4.26 ± 0.03 | 0.03 ± 0.00 | 0.18 ± 0.00 | 0.36 ± 0.00 | 0.05 ± 0.00 | 0.84 ± 0.01 | 10.38 ± 0.12 | 5.62 ± 0.04 |
| Smoky | 3.88 ± 0.03 | 13.89 ± 0.11 | 1.48 ± 0.00 | 0.02 ± 0.00 | 3.51 ± 0.03 | 0.04 ± 0.00 | 0.11 ± 0.00 | 0.21 ± 0.00 | 0.08 ± 0.00 | 0.51 ± 0.00 | 10.73 ± 0.09 | 3.88 ± 0.03 | |
| Martin | 4.87 ± 0.04 | 9.80 ± 0.08 | 1.34 ± 0.01 | 0.02 ± 0.00 | 3.21 ± 0.03 | 0.04 ± 0.00 | 0.21 ± 0.00 | 0.66 ± 0.01 | 0.08 ± 0.00 | 1.93 ± 0.05 | 8.77 ± 0.05 | 4.87 ± 0.04 | |
| Pembina | 6.32 ± 0.05 | 8.86 ± 0.07 | 1.61 ± 0.01 | 0.03 ± 0.00 | 3.67 ± 0.03 | 0.06 ± 0.00 | 0.32 ± 0.00 | 1.09 ± 0.01 | 0.05 ± 0.00 | 0.60 ± 0.00 | 12.40 ± 0.11 | 6.32 ± 0.05 | |
| Clone no 5/6 | 4.95 ± 0.04 | 10.17 ± 0.08 | 1.79 ± 0.01 | 0.03 ± 0.00 | 4.17 ± 0.03 | 0.00 ± 0.00 | 0.12 ± 0.00 | 0.35 ± 0.00 | 0.05 ± 0.00 | 0.78 ± 0.01 | 11.46 ± 0.11 | 4.95 ± 0.04 | |
| Clone type S | 2.04 ± 0.02 | 9.15 ± 0.07 | 0.89 ± 0.01 | 0.02 ± 0.00 | 3.47 ± 0.03 | 0.03 ± 0.00 | 0.15 ± 0.00 | 0.11 ± 0.00 | 0.08 ± 0.00 | 0.14 ± 0.00 | 9.76 ± 0.07 | 2.04 ± 0.02 | |
| Clone type N | 2.30 ± 0.02 | 7.20 ± 0.06 | 1.04 ± 0.01 | 0.03 ± 0.00 | 2.67 ± 0.02 | 0.03 ± 0.00 | 0.18 ± 0.00 | 0.13 ± 0.00 | 0.05 ± 0.00 | 0.53 ± 0.00 | 9.46 ± 0.04 | 2.30 ± 0.02 | |
| Flesh | Thiessen | 5.56 ± 0.04 | 6.24 ± 0.05 | 2.32 ± 0.02 | 0.03 ± 0.00 | 4.37 ± 0.03 | 0.17 ± 0.00 | 0.52 ± 0.00 | 0.89 ± 0.01 | 0.13 ± 0.00 | 0.62 ± 0.00 | 14.98 ± 0.12 | 5.56 ± 0.04 |
| Smoky | 4.81 ± 0.04 | 13.64 ± 0.11 | 2.21 ± 0.02 | 0.00 ± 0.00 | 3.26 ± 0.03 | 0.04 ± 0.00 | 0.19 ± 0.00 | 0.43 ± 0.00 | 0.06 ± 0.00 | 0.60 ± 0.00 | 21.45 ± 0.17 | 4.81 ± 0.04 | |
| Martin | 3.32 ± 0.03 | 14.17 ± 0.11 | 0.77 ± 0.01 | 0.01 ± 0.00 | 2.68 ± 0.02 | 0.05 ± 0.00 | 0.13 ± 0.00 | 0.25 ± 0.00 | 0.10 ± 0.00 | 0.36 ± 0.00 | 14.97 ± 0.12 | 3.32 ± 0.03 | |
| Pembina | 4.17 ± 0.03 | 9.99 ± 0.08 | 2.17 ± 0.02 | 0.01 ± 0.00 | 2.46 ± 0.02 | 0.05 ± 0.00 | 0.47 ± 0.00 | 0.80 ± 0.01 | 0.10 ± 0.00 | 1.24 ± 0.03 | 9.45 ± 0.08 | 4.17 ± 0.03 | |
| Clone no 5/6 | 5.41 ± 0.04 | 9.03 ± 0.07 | 2.61 ± 0.02 | 0.01 ± 0.00 | 2.81 ± 0.02 | 0.07 ± 0.00 | 0.72 ± 0.01 | 1.32 ± 0.01 | 0.06 ± 0.00 | 0.43 ± 0.00 | 18.69 ± 0.15 | 5.41 ± 0.04 | |
| Clone type S | 3.74 ± 0.03 | 6.47 ± 0.05 | 0.42 ± 0.00 | 0.01 ± 0.00 | 3.07 ± 0.02 | 0.00 ± 0.00 | 0.17 ± 0.00 | 0.68 ± 0.01 | 0.08 ± 0.00 | 0.40 ± 0.00 | 10.45 ± 0.08 | 3.74 ± 0.03 | |
| Clone type N | 2.58 ± 0.02 | 6.72 ± 0.05 | 0.15 ± 0.00 | 0.05 ± 0.00 | 2.53 ± 0.02 | 0.01 ± 0.00 | 0.12 ± 0.00 | 0.40 ± 0.00 | 0.13 ± 0.00 | 0.24 ± 0.00 | 7.29 ± 0.06 | 2.58 ± 0.02 | |
| Peel | Thiessen | 3.23 ± 0.03 | 4.74 ± 0.04 | 0.42 ± 0.00 | 0.05 ± 0.00 | 2.31 ± 0.02 | 0.01 ± 0.00 | 0.42 ± 0.00 | 1.27 ± 0.01 | 0.13 ± 0.00 | 2.81 ± 0.02 | 4.60 ± 0.04 | 3.23 ± 0.03 |
| Smoky | 4.20 ± 0.03 | 4.28 ± 0.03 | 0.50 ± 0.00 | 0.07 ± 0.00 | 2.64 ± 0.02 | 0.01 ± 0.00 | 0.64 ± 0.01 | 2.09 ± 0.02 | 0.08 ± 0.00 | 0.28 ± 0.00 | 9.11 ± 0.07 | 4.20 ± 0.03 | |
| Martin | 3.02 ± 0.02 | 5.22 ± 0.04 | 2.98 ± 0.02 | 0.01 ± 0.00 | 3.10 ± 0.02 | 0.00 ± 0.00 | 0.25 ± 0.00 | 0.49 ± 0.00 | 0.08 ± 0.00 | 0.19 ± 0.00 | 5.67 ± 0.05 | 3.02 ± 0.02 | |
| Pembina | 2.08 ± 0.02 | 5.43 ± 0.04 | 1.04 ± 0.01 | 0.03 ± 0.00 | 2.56 ± 0.02 | 0.01 ± 0.00 | 0.04 ± 0.00 | 0.29 ± 0.00 | 0.13 ± 0.00 | 0.11 ± 0.00 | 3.95 ± 0.03 | 2.08 ± 0.02 | |
| Clone no 5/6 | 2.61 ± 0.02 | 3.83 ± 0.03 | 2.93 ± 0.02 | 0.03 ± 0.00 | 2.34 ± 0.02 | 0.01 ± 0.00 | 0.62 ± 0.00 | 0.90 ± 0.01 | 0.13 ± 0.00 | 1.32 ± 0.01 | 2.50 ± 0.02 | 2.61 ± 0.02 | |
| Clone type S | 3.39 ± 0.03 | 3.46 ± 0.03 | 3.51 ± 0.03 | 0.04 ± 0.00 | 2.68 ± 0.02 | 0.01 ± 0.00 | 0.96 ± 0.01 | 1.49 ± 0.01 | 0.08 ± 0.00 | 0.13 ± 0.00 | 4.94 ± 0.04 | 3.39 ± 0.03 | |
| Clone type N | 2.36 ± 0.02 | 6.56 ± 0.05 | 0.90 ± 0.01 | 0.01 ± 0.00 | 1.91 ± 0.02 | 0.03 ± 0.00 | 0.11 ± 0.00 | 0.19 ± 0.00 | 0.07 ± 0.00 | 0.22 ± 0.00 | 12.84 ± 0.1 | 2.36 ± 0.02 | |
| Seeds | Thiessen | 4.63 ± 0.01 | 6.81 ± 0.05 | 0.32 ± 0.00 | 0.03 ± 0.00 | 1.58 ± 0.01 | 0.04 ± 0.00 | 0.12 ± 0.00 | 0.11 ± 0.00 | 0.11 ± 0.00 | 0.13 ± 0.00 | 8.96 ± 0.07 | 4.63 ± 0.01 |
| Smoky | 2.04 ± 0.02 | 4.81 ± 0.04 | 0.89 ± 0.01 | 0.03 ± 0.00 | 1.44 ± 0.01 | 0.04 ± 0.00 | 0.28 ± 0.00 | 0.35 ± 0.00 | 0.11 ± 0.00 | 1.56 ± 0.01 | 5.65 ± 0.05 | 2.04 ± 0.02 | |
| Martin | 2.04 ± 0.02 | 4.81 ± 0.04 | 0.89 ± 0.01 | 0.03 ± 0.00 | 1.44 ± 0.01 | 0.04 ± 0.00 | 0.28 ± 0.00 | 0.35 ± 0.00 | 0.11 ± 0.00 | 1.56 ± 0.01 | 5.65 ± 0.05 | 2.04 ± 0.02 | |
| Pembina | 2.64 ± 0.03 | 4.86 ± 0.04 | 0.68 ± 0.01 | 0.02 ± 0.00 | 2.19 ± 0.02 | 0.02 ± 0.00 | 0.26 ± 0.00 | 0.61 ± 0.00 | 0.07 ± 0.00 | 0.21 ± 0.00 | 7.23 ± 0.06 | 3.64 ± 0.03 | |
| Clone no 5/6 | 2.15 ± 0.02 | 3.37 ± 0.03 | 0.31 ± 0.00 | 0.03 ± 0.00 | 1.56 ± 0.01 | 0.04 ± 0.00 | 0.28 ± 0.00 | 0.27 ± 0.00 | 0.08 ± 0.00 | 0.09 ± 0.00 | 4.85 ± 0.04 | 2.15 ± 0.02 | |
| Clone type S | 2.24 ± 0.02 | 3.41 ± 0.03 | 0.27 ± 0.00 | 0.03 ± 0.00 | 1.18 ± 0.01 | 0.04 ± 0.00 | 0.32 ± 0.00 | 0.22 ± 0.00 | 0.08 ± 0.00 | 0.09 ± 0.00 | 6.85 ± 0.03 | 2.24 ± 0.02 | |
| Clone type N | 6.05 ± 0.06 | 5.26 ± 0.04 | 0.97 ± 0.01 | 0.03 ± 0.00 | 2.62 ± 0.02 | 0.72 ± 0.01 | 0.75 ± 0.01 | 1.83 ± 0.01 | 0.01 ± 0.00 | 0.38 ± 0.00 | 9.33 ± 0.10 | 5.05 ± 0.06 | |
| Fruit 3 | 4.02 b2 | 8.61 a | 1.48 b | 0.01 c | 3.14 a | 0.02 b | 0.17 c | 0.44 c | 0.07 b | 0.46 c | 9.35 a | 4.02 b | |
| Flesh | 2.53 d | 8.51 b | 0.57 d | 0.03 b | 2.55 c | 0.03 b | 0.08 d | 0.23 d | 0.10 a | 0.23 d | 8.50 c | 2.53 d | |
| Peel | 3.07 c | 6.25 c | 1.29 c | 0.03 b | 2.23 d | 0.03 b | 0.36 b | 0.62 b | 0.10 a | 2.35 a | 5.47 d | 3.07 c | |
| Seeds | 5.00 a | 5.99 d | 1.77 a | 0.04 a | 2.89 b | 0.15 a | 0.60 a | 1.30 a | 0.07 b | 0.57 b | 9.30 b | 5.00 a | |
| Fruit Fraction | Genotypes | Polyphenols (g GAE/100 g d.w.) | Antioxidative Activity | Antidiabetic Potency (IC50 (mg/mL)) | Anti-Bacterial Activity [mm] | ||
|---|---|---|---|---|---|---|---|
| ORAC (mmol vit.C/100 g d.w.) | α-Amylase | α-Glucosidase | Pancreatic Lipase | E. hirae | |||
| Fruit | Thiessen | 2.27 ± 0.02 1 | 61.02 ± 0.05 | 24.33 ± 0.02 | 38.07 ± 0.03 | 109.03 ± 0.10 | 5.88 ± 0.24 |
| Smoky | 2.04 ± 0.02 | 62.05 ± 0.06 | 31.70 ± 0.03 | 42.23 ± 0.04 | 132.63 ± 0.12 | 5.35 ± 0.01 | |
| Martin | 2.03 ± 0.02 | 40.12 ± 0.04 | 26.40 ± 0.02 | 35.30 ± 0.03 | 90.83 ± 0.08 | 4.34 ± 0.07 | |
| Pembina | 1.43 ± 0.01 | 35.37 ± 0.03 | 19.03 ± 0.02 | 27.83 ± 0.03 | 88.20 ± 0.08 | 2.57 ± 0.19 | |
| Clone no 5/6 | 1.11 ± 0.01 | 46.43 ± 0.04 | 23.43 ± 0.02 | 30.17 ± 0.03 | 95.10 ± 0.09 | 2.83 ± 0.19 | |
| Clone type S | 1.68 ± 0.01 | 47.23 ± 0.04 | 22.73 ± 0.02 | 29.87 ± 0.03 | 91.93 ± 0.08 | 8.03 ± 0.50 | |
| Clone type N | 1.98 ± 0.02 | 49.74 ± 0.04 | 18.33 ± 0.02 | 32.83 ± 0.03 | 81.93 ± 0.07 | 5.35 ± 0.32 | |
| Flesh | Thiessen | 0.96 ± 0.01 | 25.01 ± 0.02 | 33.20 ± 0.03 | 39.47 ± 0.04 | 101.24 ± 0.09 | 0.00 ± 0.00 |
| Smoky | 0.87 ± 0.01 | 28.59 ± 0.03 | 42.15 ± 0.04 | 43.79 ± 0.04 | 123.15 ± 0.11 | 0.00 ± 0.00 | |
| Martin | 0.45 ± 0.00 | 28.36 ± 0.03 | 36.03 ± 0.03 | 36.60 ± 0.03 | 84.34 ± 0.08 | 0.00 ± 0.00 | |
| Pembina | 0.49 ± 0.00 | 23.68 ± 0.02 | 25.29 ± 0.02 | 28.86 ± 0.03 | 81.90 ± 0.07 | 0.00 ± 0.00 | |
| Clone no 5/6 | 0.52 ± 0.00 | 20.70 ± 0.02 | 31.98 ± 0.03 | 31.28 ± 0.03 | 88.31 ± 0.08 | 0.00 ± 0.00 | |
| Clone type S | 0.75 ± 0.01 | 26.64 ± 0.02 | 30.23 ± 0.03 | 30.97 ± 0.03 | 85.37 ± 0.08 | 0.00 ± 0.00 | |
| Clone type N | 0.78 ± 0.01 | 33.72 ± 0.03 | 22.53 ± 0.02 | 33.87 ± 0.03 | 83.97 ± 0.08 | 0.00 ± 0.00 | |
| Peel | Thiessen | 2.86 ± 0.02 | 151.96 ± 0.14 | 14.13 ± 0.01 | 33.40 ± 0.03 | 118.46 ± 0.11 | 0.00 ± 0.00 |
| Smoky | 2.76 ± 0.02 | 105.05 ± 0.09 | 18.41 ± 0.02 | 37.06 ± 0.03 | 144.10 ± 0.13 | 0.00 ± 0.00 | |
| Martin | 1.56 ± 0.01 | 91.94 ± 0.08 | 15.34 ± 0.01 | 30.97 ± 0.03 | 98.69 ± 0.09 | 0.00 ± 0.00 | |
| Pembina | 1.11 ± 0.01 | 113.31 ± 0.1 | 11.05 ± 0.01 | 24.42 ± 0.02 | 95.83 ± 0.09 | 0.00 ± 0.00 | |
| Clone no 5/6 | 1.60 ± 0.01 | 117.65 ± 0.11 | 13.61 ± 0.01 | 26.47 ± 0.02 | 103.34 ± 0.09 | 0.00 ± 0.00 | |
| Clone type S | 1.91 ± 0.02 | 142.59 ± 0.13 | 13.21 ± 0.01 | 26.21 ± 0.02 | 99.89 ± 0.09 | 0.00 ± 0.00 | |
| Clone type N | 1.96 ± 0.02 | 136.03 ± 0.12 | 12.27 ± 0.01 | 23.60 ± 0.02 | 94.50 ± 0.09 | 0.00 ± 0.00 | |
| Seeds | Thiessen | 0.77 ± 0.01 | 14.09 ± 0.01 | 42.30 ± 0.04 | 81.90 ± 0.07 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Smoky | 0.53 ± 0.00 | 9.30 ± 0.01 | 55.12 ± 0.05 | 90.87 ± 0.08 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Martin | 0.21 ± 0.00 | 10.17 ± 0.01 | 45.90 ± 0.04 | 75.95 ± 0.07 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Pembina | 0.14 ± 0.00 | 8.96 ± 0.01 | 33.08 ± 0.03 | 59.89 ± 0.05 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Clone no 5/6 | 0.10 ± 0.00 | 12.59 ± 0.01 | 40.74 ± 0.04 | 64.90 ± 0.06 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Clone type S | 0.26 ± 0.00 | 8.99 ± 0.01 | 39.52 ± 0.04 | 64.26 ± 0.06 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Clone type N | 0.26 ± 0.00 | 18.47 ± 0.02 | 51.07 ± 0.05 | 77.70 ± 0.07 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Fruit 3 | 1.79 b2 | 48.85 b | 23.71 c | 33.76 c | 98.52 b | 4.91 a | |
| Flesh | 0.69 | 26.67 c | 31.63 b | 34.97 b | 92.61 c | 0.00 b | |
| Peel | 2.00 a | 122.65 a | 14.00 d | 28.88 d | 107.83 a | 0.00 b | |
| Seeds | 0.32 | 11.79 d | 43.96 a | 73.64 a | 0.00 d | 0.00 b | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lachowicz, S.; Wiśniewski, R.; Ochmian, I.; Drzymała, K.; Pluta, S. Anti-Microbiological, Anti-Hyperglycemic and Anti-Obesity Potency of Natural Antioxidants in Fruit Fractions of Saskatoon Berry. Antioxidants 2019, 8, 397. https://doi.org/10.3390/antiox8090397
Lachowicz S, Wiśniewski R, Ochmian I, Drzymała K, Pluta S. Anti-Microbiological, Anti-Hyperglycemic and Anti-Obesity Potency of Natural Antioxidants in Fruit Fractions of Saskatoon Berry. Antioxidants. 2019; 8(9):397. https://doi.org/10.3390/antiox8090397
Chicago/Turabian StyleLachowicz, Sabina, Rafał Wiśniewski, Ireneusz Ochmian, Katarzyna Drzymała, and Stanisław Pluta. 2019. "Anti-Microbiological, Anti-Hyperglycemic and Anti-Obesity Potency of Natural Antioxidants in Fruit Fractions of Saskatoon Berry" Antioxidants 8, no. 9: 397. https://doi.org/10.3390/antiox8090397