Characteristics of Selected Antioxidative and Bioactive Compounds in Meat and Animal Origin Products


Abstract
:1. Introduction
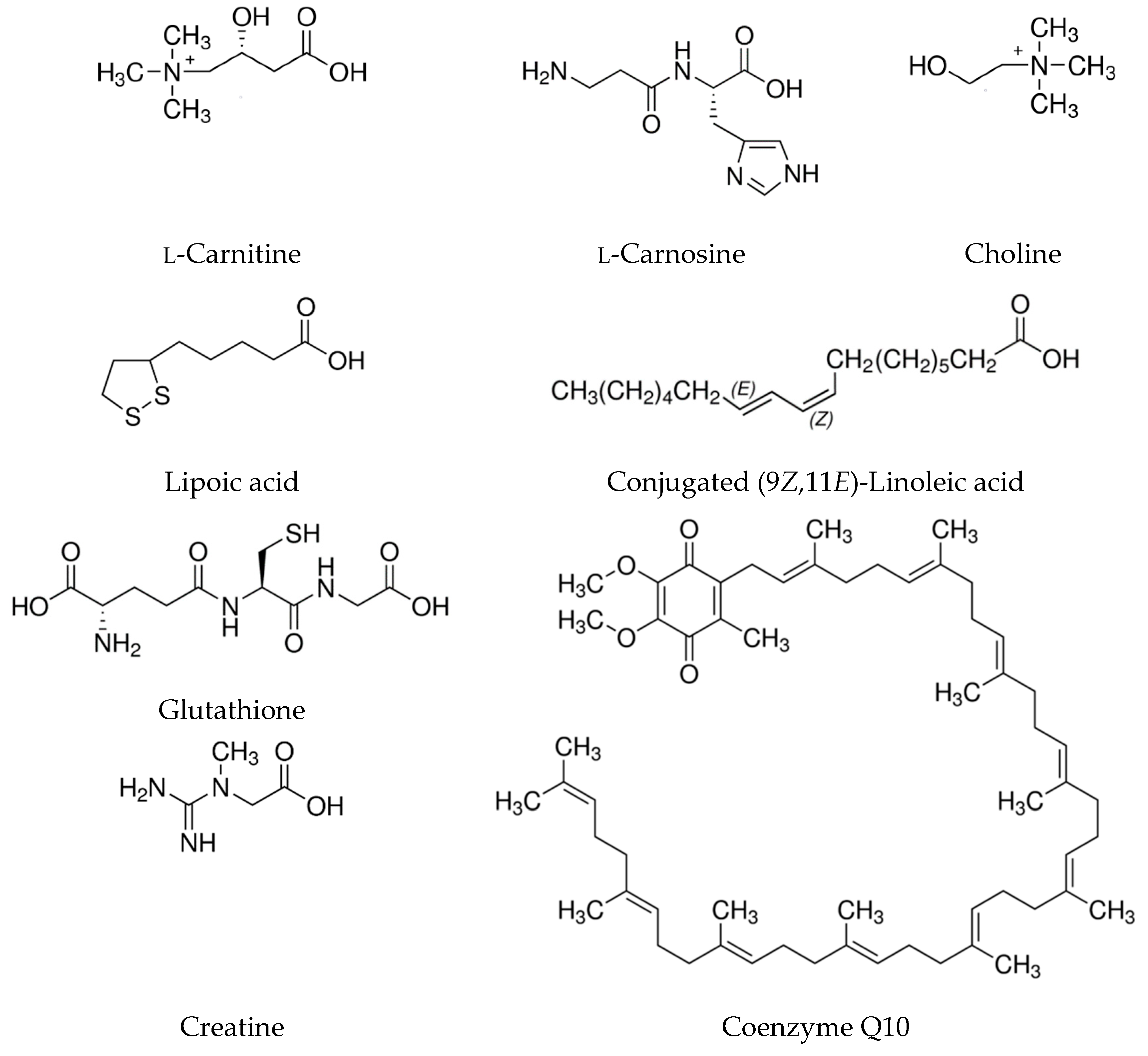
1.1. l-carnitine
1.2. l-carnosine
1.3. Choline
1.4. Alpha-Lipoic Acid
1.5. Conjugated Linoleic Acid (CLA)
1.6. Glutathione
1.7. Taurine
1.8. Bioactive Peptides
1.9. Coenzyme Q10
1.10. Creatine
1.11. Meat Consumption and Health Controversy
2. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pereira, P.M.; Vicente, A.F. Meat nutritional composition and nutritive role in the human diet. Meat Sci. 2013, 93, 586–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biesalski, H.K. Meat as a component of a healthy diet—are there any risks or benefits if meat is avoided in the diet? Meat Sci. 2005, 70, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Sarriés, M.V.; Tateo, A.; Polidori, P.; Franco, D.; Lanzae, M. Carcass characteristics, meat quality and nutritional value of horsemeat: A review. Meat Sci. 2014, 96, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Schmid, A. Bioactive substances in meat and meat products. Fleischwirtschaft Int. 2010, 2, 127–133. [Google Scholar]
- Biesalski, H.K.; Dragsted, L.O.; Elmadfa, I.; Grossklaus, R.; Müller, M.; Schrenk, D.; Walter, P.; Weber, P. Bioactive compounds: Definition and assessment of activity. Nutrients 2009, 25, 1202–1205. [Google Scholar] [CrossRef] [PubMed]
- Kumar, Y.; Yadav, D.N.; Ahmad, T.; Narsaiah, K. Recent trends in the use of natural antioxidants for meat and meat products. Compr. Rev. Food Sci. Food Saf. 2015, 14, 796–812. [Google Scholar] [CrossRef]
- Kwasek, M. Tendencje w spożyciu mięsa na świecie [Global meat consumption trends]. Ekon. Kuj.-Pom. Szk. Wyż. Bydg. 2013, 6, 265–284. [Google Scholar]
- OECD/FAO: OECD-FAO Agricultural Outlook. OECD Agriculture Statistics (database) 2015. Available online: https://www.oecd-ilibrary.org/agriculture-and-food/oecd-fao-agricultural-outlook-2015_agr_outlook-2015-en (accessed on 17 May 2019).
- Parvanova, A.; Trillini, M.; Podestà, M.A.; Iliev, I.P.; Aparicio, C.; Perna, A.; Peraro, F.; Rubis, N.; Gaspari, F.; Cannata, A.; et al. Blood pressure and metabolic effects of acetyl-l-carnitine in type 2 diabetes: DIABASI Randomized Controlled Trial. J. Endocr. Soc. 2018, 2, 420–436. [Google Scholar] [CrossRef]
- El-sheikh, H.M.; El-Haggar, S.M.; Elbedewy, T.A. Comparative study to evaluate the effect of l-carnitine plus glimepiride versus glimepiride alone on insulin resistance in type 2 diabetic patients. Diabetes Metab. Syndr. 2019, 13, 167–173. [Google Scholar] [CrossRef]
- Lee, B.J.; Lin, J.S.; Lin, Y.C.; Lin, P.T. Antiinflammatory effects of l-carnitine supplementation [1000 mg/d] in coronary artery disease patients. Nutrients 2015, 31, 475–479. [Google Scholar] [CrossRef]
- Lee, B.J.; Lin, J.S.; Lin, Y.C.; Lin, P.T. Effects of l-carnitine supplementation on oxidative stress and antioxidant enzymes activities in patients with coronary artery disease: A randomized, placebo-controlled trial. Nutr. J. 2014, 13, 79. [Google Scholar] [CrossRef] [PubMed]
- Mohammed-Jawad, N.K.; Al-Sabbagh, M.; Al-Jezaeri, K.A. Role of l-carnitine and coenzyme Q10 as adjuvant therapy in patients with type 2 diabetes mellitus. Am. J. Pharmacol. Sci. 2014, 2, 82–86. [Google Scholar] [CrossRef]
- Zhang, J.J.; Wu, Z.B.; Cai, Y.J.; Ke, B.; Huang, Y.J.; Qiu, C.P.; Yang, Y.B.; Shi, L.Y.; Qin, J. l-carnitine ameliorated fasting-induced fatigue, hunger, and metabolic abnormalities in patients with metabolic syndrome: A randomized controlled study. Nutr. J. 2014, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Baye, E.; Ukropec, J.; de Courten, M.P.J.; Mousa, A.; Kurdiova, T.; Johnson, J.; Wilson, K.; Plebanski, M.; Aldini, G.; Ukropcova, B.; et al. Carnosine supplementation improves serum resistin concentrations in overweight or obese otherwise healthy adults: A pilot randomized trial. Nutrients 2018, 10, 1258. [Google Scholar] [CrossRef] [PubMed]
- Elbarbary, N.S.; Ismail, E.A.R.; El-Naggar, A.R.; Hamouda, M.H.; El-Hamamsy, M. The effect of 12 weeks carnosine supplementation on renal functional integrity and oxidative stress in pediatric patients with diabetic nephropathy: A randomized placebo-controlled trial. Ediatr. Diabetes. 2018, 19, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Houjeghani, S.; Kheirouri, S.; Faraji, E.; Jafarabadi, M.A. l-Carnosine supplementation attenuated fasting glucose, triglycerides, advanced glycation end products, and tumor necrosis factor—α levels in patients with type 2 diabetes: A double-blind placebo-controlled randomized clinical trial. Nutr. Res. 2018, 49, 96–106. [Google Scholar] [CrossRef]
- Houjeghani, S.; Kheirouri, S.; Faraji, E.; Jafarabadi, M.A.; Jabbari, M. Antioxidant status, lipid peroxidation and protein oxidation in type 2 diabetic patients: Beneficial effects of supplementation with carnosine: A randomized, double-blind, placebo-controlled trial. Iran. Red. Crescent. Med. J. 2018, 20, e64116. [Google Scholar] [CrossRef]
- Fallah, M.; Askari, G.; Soleimani, A.; Feizi, A.; Asemi, Z. Clinical trial of the effects of coenzyme Q10 supplementation on biomarkers of inflammation and oxidative stress in diabetic hemodialysis patients. Int. J. Prev. Med. 2019, 10, 12. [Google Scholar]
- Zhang, P.; Yang, C.; Guo, H.; Wang, J.; Lin, S.; Li, H.; Yang, Y.; Ling, W. Treatment of coenzyme Q10 for 24 weeks improves lipid and glycemic profile in dyslipidemic individuals. J. Clin. Lipidol. 2018, 12, 417–427. [Google Scholar] [CrossRef]
- Lee, B.; Tseng, Y.; Yen, C.; Lin, P. Effects of coenzyme Q10 supplementation [300 mg/day] on antioxidation and anti-inflammation in coronary artery disease patients during statins therapy: A randomized, placebo-controlled trial. Nutr. J. 2013, 12, 142. [Google Scholar] [CrossRef]
- Gaullier, J.M.; Halse, J.; Høye, K.; Kristiansen, K.; Fagertun, H.; Vik, H.; Gudmundsen, O. Conjugated linoleic acid supplementation for 1 y reduces body fat mass in healthy overweight humans. Am. J. Clin. Nutr. 2004, 79, 1118–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmair, E.M.; Wood, S.G.; Keizer, H.G.; Horgan, G.W.; Ford, I.; de Roos, B. Supplementation with a 9c,11t-rich conjugated linoleic acid blend shows no clear inhibitory effects on platelet function in healthy subjects at low and moderate cardiovascular risk: A randomized controlled trial. Mol. Nutr. Food Res. 2015, 59, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Bulut, S.; Bodur, E.; Colak, R.; Turnagol, H. Effects of conjugated linoleic acid supplementation and exercise on post-heparin lipoprotein lipase, butyrylcholinesterase, blood lipid profile and glucose metabolism in young men. Chem. Biol. Interact. 2013, 203, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, R.F.; Uehara, S.K.; Rosa, G. Microencapsulated conjugated linoleic acid associated with hypocaloric diet reduces body fat in sedentary women with metabolic syndrome. Vasc. Health. Risk. Manag. 2012, 8, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.C.; Lin, Y.H.; Huang, H.P.; Hsu, W.L.; Houng, J.Y.; Huang, C.K. Effect of conjugated linoleic acid supplementation on weight loss and body fat composition in a Chinese population. Nutrition 2012, 28, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.V.; Jacques, H.; Plourde, M.; Mitchell, P.L.; McLeod, R.S.; Jones, P.J. Conjugated linoleic acid supplementation for 8 weeks does not affect body composition, lipid profile, or safety biomarkers in overweight, hyperlipidemic men. J. Nutr. 2011, 141, 1286–1291. [Google Scholar] [CrossRef] [PubMed]
- Gualano, B.; De Salles Painneli, V.; Roschel, H.; Artioli, G.G.; Neves, M., Jr.; De Sá Pinto, A.L.; Da Silva, M.E.; Cunha, M.R.; Otaduy, M.C.; Leite Cda, C.; et al. Creatine in type 2 diabetes: A randomized, double-blind, placebo-controlled trial. Med. Sci. Sports Exerc. 2011, 43, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Richie, J.P.; Nichenametla, S.; Neidig, W.; Calcagnotto, A.; Haley, J.S.; Schell, T.D.; Muscat, J.E. Randomized controlled trial of oral glutathione supplementation on body stores of glutathione. Eur. J. Nutr. 2015, 54, 251–263. [Google Scholar] [CrossRef]
- Sinha, R.; Sinha, I.; Calcagnotto, A.; Trushin, N.; Haley, J.S.; Schell, T.D.; Richie, J.P., Jr. Oral supplementation with liposomal glutathione elevates body stores of glutathione and markers of immune function. Eur. J. Clin. Nutr. 2018, 72, 105–111. [Google Scholar] [CrossRef]
- Campolo, J.; Bernardi, S.; Cozzi, L.; Rocchiccioli, S.; Dellanoce, C.; Cecchettini, A.; Tonini, A.; Parolini, M.; De Chiara, B.; Micheloni, G.; et al. Medium-term effect of sublingual l-glutathione supplementation on flow-mediated dilation in subjects with cardiovascular risk factors. Nutrients 2017, 38, 41–47. [Google Scholar] [CrossRef]
- Honda, Y.; Kessoku, T.; Sumida, Y.; Kobayashi, T.; Kato, T.; Ogawa, Y.; Tomeno, W.; Imajo, K.; Fujita, K.; Yoneda, M.; et al. Efficacy of glutathione for the treatment of nonalcoholic fatty liver disease: An open-label, single-arm, multicenter, pilot study. BMC Gastroenterol. 2017, 17, 96. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.; Bradley, R.D. Effects of oral glutathione supplementation on systemic oxidative stress biomarkers in human volunteers. J. Altern. Complement. Med. 2011, 17, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Atmaca, H.U.; Akbas, F. The effect of short term alpha lipoic acid administration on adiponectin and body weight in type 2 diabetes mellitus patients. Acta Endocrinol. 2017, 4, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Hosseinpour-Arjmand, S.; Amirkhizi, F.; Ebrahimi-Mameghani, M. The effect of alpha-lipoic acid on inflammatory markers and body composition in obese patients with non-alcoholic fatty liver disease: A randomized, double-blind, placebo-controlled trial. J. Clin. Pharm. Ther. 2019, 44, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Aslfalah, H.; Jamilian, M.; Khosrowbeygi, A. Elevation of the adiponectin/leptin ratio in women with gestational diabetes mellitus after supplementation with alpha-lipoic acid. Gynecol. Endocrinol. 2018, 35, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, V.; Khorvash, F.; Feizi, A.; Askari, G. Does alpha-lipoic acid supplementation modulate cardiovascular risk factors in patients with stroke? A randomized, double-blind clinical trial. Int. J. Prev. Med. 2018, 9, 34. [Google Scholar] [PubMed]
- Mohammadi, V.; Khorvash, F.; Feizi, A.; Askari, G. Does alpha-lipoic acid comsumption improve lipid profile in patients with stroke? A randomized, double blind, placebo-controlled clinical trial. Iran Red Crescent Med. J. 2017, 19, e58765. [Google Scholar] [CrossRef]
- Panda, P.; Panda, S.K.; Mishra, T. Effect of alpha lipoic acid in treatment of type 2 diabetes. Asian. J. Pharm. Clin. Res. 2017, 10, 204–207. [Google Scholar] [CrossRef]
- Aslfalah, H.; Jamilian, M.; Rafiei, F.; Khosrowbeygi, A. Reduction in maternal serum values of glucose and gamma-glutamyltransferase after supplementation with alpha-lipoic acid in women with gestational diabetes mellitus. J. Obstet. Gynaecol. Res. 2019, 45, 313–317. [Google Scholar] [CrossRef]
- Sun, Q.; Wang, B.; Li, Y.; Sun, F.; Li, P.; Xia, W.; Zhou, X.; Li, Q.; Wang, X.; Chen, J.; et al. Taurine supplementation lowers blood pressure and improves vascular function in prehypertension: Randomized, double-blind, placebo-controlled study. Hypertension 2016, 67, 541–549. [Google Scholar] [CrossRef]
- Schwarzer, R.; Kivaranovic, D.; Mandorfer, M.; Paternostro, R.; Wolrab, D.; Heinisch, B.; Reiberger, T.; Ferlitsch, M.; Gerner, C.; Trauner, M.; et al. Randomised clinical study: The effects of oral taurine 6g/day vs placebo on portal hypertension. Aliment. Pharmacol. Ther. 2018, 47, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, Y.; Hagiwara, H.; Nishimatsu, S.I.; Hirakawa, A.; Kamimura, N.; Ohtsubo, H.; Fukai, Y.; Murakami, T.; Koga, Y.; Goto, Y.I.; et al. Taurine supplementation for prevention of stroke-like episodes in MELAS: A multicentre, open-label, 52-week phase III trial. J. Neurol. Neurosurg. Psychiatry 2018, 90, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Sepand, M.R.; Razavi-Azarkhiavi, K.; Omidi, A.; Zirak, M.R.; Sabzevari, S.; Kazemi, A.R.; Sabzevari, O. Effect of acetyl-l-carnitine on antioxidant status, lipid peroxidation, and oxidative damage of arsenic in rat. Biol. Trace. Elem. Res. 2016, 171, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Keskin, E.; Uluisik, D.; Altin, M. Antioxidant effect of l-carnitine in rats fed cholesterol rich diet. Anim. Vet. Sci. 2015, 3, 113–116. [Google Scholar] [CrossRef]
- Elgazzar, U.B.; Ghanema, I.I.A.; Kalaba, Z.M. Effect of dietary l-carnitine supplementation on the concentration of circulating serum metabolites in growing New Zealand rabbits. Aust. J. Basic App. Sci. 2012, 6, 80–84. [Google Scholar]
- Hamza, R.Z.; Al-Eisa, R.A.; Mehana, A.E.; El-Shenawy, N.S. Effect of l-carnitine on aspartame-induced oxidative stress, histopathological changes, and genotoxicity in liver of male rats. J. Basic Clin. Physiol. Pharmacol. 2019, 30, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Li, Q.; Zhong, W.; Dong, J.; Wang, Z.; Wang, C. l-carnitine ameliorated fatty liver in high-calorie diet/STZ-induced type 2 diabetic mice by improving mitochondrial function. Diabetol. Metab. Syndr. 2011, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Aydın, A.F.; Küçükgergin, C.; Çoban, J.; Doğan-Ekici, I.; Doğru-Abbasoğlu, S.; Uysal, M.; Koçak-Toker, N. Carnosine prevents testicular oxidative stress and advanced glycation end product formation in d-galactose-induced aged rats. Andrologia 2018, 50, e12939. [Google Scholar] [CrossRef] [PubMed]
- Aydın, A.F.; Bingül, İ.; Küçükgergin, C.; Doğan-Ekici, I.; Doğru Abbasoğlu, S.; Uysal, M. Carnosine decreased oxidation and glycation products in serum and liver of high-fat diet and low-dose streptozotocin-induced diabetic rats. Int. J. Exp. Pathol. 2017, 98, 278–288. [Google Scholar] [CrossRef]
- Aydın, A.F.; Küçükgergin, C.; Bingül, İ.; Doğan-Ekici, I.; Doğru-Abbasoğlu, S.; Uysal, M. Effect of Carnosine on renal function, oxidation and glycation products in the kidneys of high-fat diet/streptozotocin-induced diabetic rats. Exp. Clin. Endocrinol. Diabetes. 2017, 125, 282–289. [Google Scholar]
- Bingül, İ.; Yılmaz, Z.; Aydın, A.F.; Çoban, J.; Doğru-Abbasoğlu, S.; Uysal, M. Antiglycation and anti-oxidant efficiency of carnosine in the plasma and liver of aged rats. Geriatr. Gerontol. Int. 2017, 17, 2610–2614. [Google Scholar] [CrossRef] [PubMed]
- El-Haleem, M.R.; Yassen, O.I.; Raafat, N. Protective role of coenzyme Q 10 against high cholesterol diet induced histological and biochemical changes in cerebellar cortex of adult albino rats. IOSR J. Dent. Med. Sci. 2014, 13, 50–61. [Google Scholar] [CrossRef]
- Deminice, R.; da Silva, R.P.; Lamarre, S.G.; Brown, C.; Furey, G.N.; McCarter, S.A.; Jordao, A.A.; Kelly, K.B.; King-Jones, K.; Jacobs, R.L.; et al. Creatine supplementation prevents the accumulation of fat in the livers of rats fed a high-fat diet. J. Nutr. 2011, 141, 1799–1804. [Google Scholar] [CrossRef] [PubMed]
- Deminice, R.; de Castro, G.S.; Francisco, L.V.; da Silva, L.E.; Cardoso, J.F.; Frajacomo, F.T.; Teodoro, B.G.; Dos Reis Silveira, L.; Jordao, A.A. Creatine supplementation prevents fatty liver in rats fed choline-deficient diet: A burden of one-carbon and fatty acid metabolism. J. Nutr. Biochem. 2015, 26, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.P.; Leonard, K.A.; Jacobs, R.L. Dietary creatine supplementation lowers hepatic triacylglycerol by increasing lipoprotein secretion in rats fed high-fat diet. J. Nutr. Biochem. 2017, 50, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.B.; Choudhary, R.; Vishwakarma, P.K.; Singh, A.; Shree, J.; Bodakhe, S.H. Protective effect of alpha-lipoic acid on progression of cataract formation in fructose-induced experimental cataract. PharmaNutrition 2017, 5, 127–132. [Google Scholar] [CrossRef]
- Jamor, P.; Ahmadvand, H.; Ashoory, H.; Babaeenezhad, E. Effect of alpha-lipoic acid on antioxidant gene expression and kidney injury in alloxan-induced diabetic rats. J. Nephropathol. 2019, 8, e06. [Google Scholar] [CrossRef]
- Panzhinskiy, E.; Bashir, R.; Bagchi, D.; Nair, S. Effect of curcumin and α-lipoic acid in attenuating weight gain and adiposity. J. Am. Coll. Nutr. 2019, 1–6. [Google Scholar] [CrossRef]
- Jamor, P.; Ahmadvand, H.; Birjandi, M.; Sharafabad, B.E. Activity of serum paraoxonase 1, lipid profile and atherogenic indexes in diabetic induced rats treated with alpha lipoic acid. J. Nephropathol. 2018, 7, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Sena, C.; Cipriano, M.; Botelho, M.; Seiça, R. Lipoic acid prevents high-fat diet-induced hepatic steatosis in Goto Kakizaki rats by reducing oxidative stress through Nrf2 activation. Int. J. Mol. Sci. 2018, 19, 2706. [Google Scholar] [CrossRef]
- Rosenblat, M.; Volkova, N.; Coleman, R.; Aviram, M. Anti-oxidant and anti-atherogenic properties of liposomal glutathione: Studies in vitro, and in the atherosclerotic apolipoprotein E-deficient mice. Atherosclerosis 2007, 195, e61–e68. [Google Scholar] [CrossRef] [PubMed]
- Lauver, D.A.; Kaissarian, N.M.; Lucchesi, B.R. Oral pretreatment with liposomal glutathione attenuates reperfusion injury in rabbit isolated hearts. J. Cardiovasc. Pharmacol. 2013, 61, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Mirdhayati, I.; Hermanianto, J.; Wijaya, C.H.; Sajuthi, D.; Arihara, K. Angiotensin converting enzyme (ACE) inhibitory and antihypertensive activities of protein hydrolysate from meat of Kacang goat (Capra aegagrus hircus). J. Sci. Food Agr. 2016, 96, 3536–3542. [Google Scholar] [CrossRef] [PubMed]
- Escudero, E.; Aristoy, M.C.; Nishimura, H.; Arihara, K.; Toldrá, F. Antihypertensive effect and antioxidant activity of peptide fractions extracted from Spanish dry-cured ham. Meat Sci. 2012, 91, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Escudero, E.; Toldrá, F.; Sentandreu, M.A.; Nishimura, H.; Arihara, K. Antihypertensive activity of peptides identified in the in vitro gastrointestinal digest of pork meat. Meat Sci. 2012, 91, 382–384. [Google Scholar] [CrossRef] [PubMed]
- Muguruma, M.; Ahhmed, A.M.; Katayama, K.; Kawahara, S.; Maruyama, M.; Nakamura, T. Identification of pro-drug type ACE inhibitory peptide sourced from porcine myosin B: Evaluation of its antihypertensive effects in vivo. Food Chem. 2009, 114, 516–522. [Google Scholar] [CrossRef]
- Nakade, K.; Kamishima, R.; Inoue, Y.; Ahhmed, A.; Kawahara, S.; Nakayama, T.; Maruyama, M.; Numata, M.; Ohta, K.; Aoki, T.; et al. Identification of an antihypertensive peptide derived from chicken bone extract. J. Anim. Sci. 2008, 79, 710–715. [Google Scholar] [CrossRef]
- Saiga, A.I.; Iwai, K.; Hayakawa, T.; Takahata, Y.; Kitamura, S.; Nishimura, T.; Morimatsu, F. Angiotensin I-converting enzyme-inhibitory peptides obtained from chicken collagen hydrolysate. J. Agr. Food Chem. 2008, 56, 9586–9591. [Google Scholar] [CrossRef]
- Yousef, H.N.; Aboelwafa, H.R. The potential protective role of taurine against 5-fluorouracil-induced nephrotoxicity in adult male rats. Exp. Toxicol. Pathlol. 2017, 69, 265–274. [Google Scholar] [CrossRef]
- Nandhini, A.T.; Anuradha, C.V. Hoe 140 abolishes the blood pressure lowering effect of taurine in high fructose-fed rats. Amino Acids 2004, 26, 299–303. [Google Scholar] [CrossRef]
- Murakami, S.; Kondo, Y.; Toda, Y.; Kitajima, H.; Kameo, K.; Sakono, M.; Fukuda, N. Effect of taurine on cholesterol metabolism in hamsters: Up-regulation of low density lipoprotein [LDL] receptor by taurine. Life Sci. 2002, 70, 2355–2366. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Dessouki, A.A.; Abdel-Rahman, H.G.; Eltaysh, R.; Alkahtani, S. Hepatorenal protective effects of taurine and N-acetylcysteine against fipronil-induced injuries: The antioxidant status and apoptotic markers expression in rats. Sci. Total Environ. 2019, 650, 2063–2073. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Chowdhury, S.; Das, A.K.; Sil, P.C. Taurine ameliorates oxidative stress induced inflammation and ER stress mediated testicular damage in STZ-induced diabetic Wistar rats. Food Chem. Toxicol. 2019, 124, 64–80. [Google Scholar] [CrossRef] [PubMed]
- Heidari, R.; Behnamrad, S.; Khodami, Z.; Ommati, M.M.; Azarpira, N.; Vazin, A. The nephroprotective properties of taurine in colistin-treated mice is mediated through the regulation of mitochondrial function and mitigation of oxidative stress. Biomed. Pharmacother. 2019, 109, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Adedara, I.A.; Alake, S.E.; Adeyemo, M.O.; Olajide, L.O.; Ajibade, T.O.; Farombi, E.O. Taurine enhances spermatogenic function and antioxidant defense mechanisms in testes and epididymis of l-NAME-induced hypertensive rats. Biomed. Pharmacother. 2018, 97, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Ince, S.; Arslan-Acaroz, D.; Demirel, H.H.; Varol, N.; Ozyurek, H.A.; Zemheri, F.; Kucukkurt, I. Taurine alleviates malathion induced lipid peroxidation, oxidative stress, and proinflammatory cytokine gene expressions in rats. Biomed. Pharmacother. 2017, 96, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, N.A.; Gawad, H.A. Taurine dietary supplementation attenuates brain, thyroid, testicular disturbances and oxidative stress in streptozotocin-induced diabetes mellitus in male rats. Beni-Suef Univ. J. Basic Appl. Sci. 2017, 6, 247–252. [Google Scholar] [CrossRef]
- Reeta, K.H.; Singh, D.; Gupta, Y.K. Chronic treatment with taurine after intracerebroventricular streptozotocin injection improves cognitive dysfunction in rats by modulating oxidative stress, cholinergic functions and neuroinflammation. Neurochem. Int. 2017, 108, 146–156. [Google Scholar] [CrossRef]
- Czeczot, H.; Ścibior, D. Rola l-karnitiny w przemianach, żywieniu i terapii [The role of l-carnitine in metabolism, nutrition and therapy]. Postępy Hig. Med. Dośw. 2005, 59, 9–19. [Google Scholar]
- Kapoor, P.; Kapoor, A.K. Coenzyme Q10—A novel molecule. J. Indian Acad. Clin. Med. 2013, 14, 37–45. [Google Scholar]
- Kalpana, A. Effects of l-carnitine [Neutraceutical] in weight management among overweight and obese adults of age between 20–45 yrs—A comparative study in Chennai and Tirupathi. Int. J. Sci. Res Pub. 2012, 2, 1–5. [Google Scholar]
- Flanagan, J.L.; Simmons, P.A.; Vehige, J.; Willcox, M.D.; Garrett, Q. Role of carnitine in disease. Nutr. Metab. 2010, 7, 1–14. [Google Scholar] [CrossRef]
- Rospond, B.; Chłopicka, J. Funkcje biologiczne l-karnityny i jej zawartość w wybranych products spożywczych [Biological functions of l-carnitine and its content in selected food products]. Przegl. Lek. 2013, 70, 85–91. [Google Scholar] [PubMed]
- Dayanand, C.D.; Krishnamurthy, N.; Ashakiran, S.; Shashidhar, K.N. Carnitine: A novel health factor—An overview. Int. J. Pharm. Biomed. Res. 2011, 2, 79–89. [Google Scholar]
- Lourenco, R.; Camilo, M.E. Taurine: A conditionally essential amino acid in humans? An overview in health and disease. Nutr. Hosp. 2002, 17, 262–270. [Google Scholar] [PubMed]
- Manhiani, P.S.; Northcutt, J.K.; Han, I.; Bridges, W.C.; Dawson, P.L. Antioxidant activity of carnosine extracted from various poultry tissues. Poult. Sci. 2013, 92, 444–453. [Google Scholar] [CrossRef]
- Kiliś-Pstrusińska, K. Karnozyna i karnozynaza a choroby nerek [Carnosine and carnosinase and kidney disease]. Postepy. Hig. Med. Dosw. 2012, 66, 215–221. [Google Scholar] [CrossRef]
- Hipkiss, A.R.; Cartwright, S.P.; Bromley, C.; Gross, S.R.; Bill, R.M. Carnosine: Can understanding its actions on energy metabolism and protein homeostasis inform its therapeutic potential? Chem. Cent. J. 2013, 7, 38. [Google Scholar] [CrossRef]
- Budzeń, S.; Rymaszewska, J. The biological role of carnosine and its possible applications in medicine. Adv Clin. Exp. Med. 2013, 22, 739–744. [Google Scholar]
- Zięba, R. Karnozyna-aktywność biologiczna i perspektywy zastosowania w farmakoterapii [Carnosine—biological activity and prospects for applications in pharmacotherapy]. Wiad. Lek. 2007, 60, 73–79. [Google Scholar]
- Aristoy, M.C.; Toldra, F. Histidine dipeptides HPLC-based test for the detection of mammalian origin proteins in feeds for ruminants. Meat Sci. 2004, 67, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Kim, E.J.; Kim, Y.; Choi, C.; Lee, B. Effects of alpha-lipoic acid and l-carnosine supplementation on antioxidant activities and lipid profiles in rats. Nutr. Res. Pract. 2011, 5, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Baykara, B.; Micili, S.C.; Tugyan, K.; Tekmen, I.; Bagriyanik, H.; Sonmez, U.; Sonmez, A.; Oktay, G.; Yener, N.; Ozbal, S. The protective effects of carnosine in alcohol-induced hepatic injury in rats. Toxicol. Ind. Health 2014, 30, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Bellia, F.; Vecchio, G.; Cuzzocrea, S.; Calabrese, V.; Rizzarelli, E. Neuroprotective features of carnosine in oxidative driven diseases. Mol. Aspects. Med. 2011, 32, 258–266. [Google Scholar] [CrossRef]
- Aldini, G.; Orioli, M.; Rossoni, G.; Savi, F.; Braidotti, P.; Vistoli, G.; Yeum, K.J.; Negrisoli, G.; Carini, M. The carbonyl scavenger carnosine ameliorates dyslipidaemia and renal function in Zucker obese rats. J. Cell. Mol. Med. 2011, 6, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Wurtman, R.; Cansev, M.; Ulus, I. Choline and its products acetylcholine and phosphatidylcholine. In Handbook of Neurochemistry and Molecular Neurobiology: Neural Lipids, 3rd ed.; Tettamani, G., Goracci, G., Eds.; Springer: New York, NY, USA, 2010; pp. 443–500. [Google Scholar]
- Patterson, K.Y.; Bhagwat, S.A.; Williams, J.R.; Howe, J.C.; Holden, J.M. USDA Database for the choline content of common foods, release two. Nutrient Data Laboratory, Beltsville Human Nutrition Research Center, ARS, USDA (2008). 2008. Available online: https://data.nal.usda.gov/dataset/usda-database-choline-content-common-foods-release-2-2008 (accessed on 17 July 2019).
- Ueland, P.M. Choline and betaine in health and disease. J. Inherit. Metab. Dis. 2011, 34, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Wiedeman, A.M.; Barr, S.I.; Green, T.J.; Xu, Z.; Innis, S.M.; Kitts, D.D. Dietary choline intake: Current state of knowledge across the life cycle. Nutrients 2018, 10, 1513. [Google Scholar] [CrossRef]
- Zhu, J.; Wu, Y.; Tang, Q.; Leng, Y.; Cai, W. The effects of choline on hepatic lipid metabolism, mitochondrial function and antioxidative status in human hepatic C3A cells exposed to excessive energy substrates. Nutrients 2014, 9, 2552–2571. [Google Scholar] [CrossRef] [PubMed]
- Shaw, G.M.; Carmichael, S.L.; Yang, W.; Selvin, S.; Schaffer, D.M. Periconceptional dietary intake of choline and betaine and neural tube defects in offspring. Am. J. Epidemiol. 2004, 160, 102–109. [Google Scholar] [CrossRef]
- Detopoulou, P.; Panagiotakos, D.B.; Antonopoulou, S.; Pitsavos, C.; Stefanadis, C. Dietary choline and betaine intakes in relation to concentrations of inflammatory markers in healthy adults: The ATTICA study. Am. J. Clin. Nutr. 2008, 87, 424–430. [Google Scholar] [CrossRef]
- Bjelland, I.; Tell, G.S.; Vollset, S.E.; Konstantinova, S.V.; Ueland, P.M. Choline in anxiety and depression: The Hordaland Health Study. Am. J. Clin. Nutr. 2009, 90, 1056–1060. [Google Scholar] [CrossRef]
- Golbidi, S.; Badran, M.; Laher, I. Diabetes and alpha lipoic acid. Front. Pharmacol. 2011, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Skibska, B.; Goraca, A. The protective effect of lipoic acid on selected cardiovascular diseases caused by age-related oxidative stress. Oxid. Med. Cell. Longev. 2015, 2, 1–11. [Google Scholar] [CrossRef]
- Gomes, M.B.; Negrato, C.A. Alpha-lipoic acid as a pleiotropic compound with potential therapeutic use in diabetes and other chronic diseases. Diabetol. Metab. Syndr. 2014, 6, 80. [Google Scholar] [CrossRef]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential. Biochim. Biophys. Acta. 2009, 1790, 1149–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, N.; Yan, X.; Xue, J.; Wang, H. The protective effects of alpha-lipoic acid on kidneys in type 2 diabetic goto-kakisaki rats via reducing oxidative stress. Int. J. Mol. Sci. 2013, 14, 6746–6756. [Google Scholar] [CrossRef] [PubMed]
- Hussein, S.A.; Abdel-mageid, A.D.; Abu-ghazalla, A.M. Biochemical study on the effect of alpha-lipoic acid on lipid metabolism of rats fed high fat diet. Banha Vet. Med. J. 2015, 28, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Murali, P.; Sherin, G.; Dominic, G. Dietary supplementation of alpha lipoic acid on serum lipid profile of broiler chicken fed with animal fat diet. J. Genet. Eng. Biotechnol. 2014, 5, 23–28. [Google Scholar]
- Yang, Y.; Li, W.; Liu, Y.; Li, Y.; Gao, L.; Zhao, J.J. Alpha-lipoic acid attenuates insulin resistance and improves glucose metabolism in high fat diet-fed mice. Acta. Pharmacol. Sin. 2014, 35, 1285–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T.S.; Kim, S.K.; Shin, H.J.; Jeon, B.T.; Hahm, J.R.; Roh, G.S. Alpha-lipoic acid prevents non-alcoholic fatty liver disease in OLETF rats. Liver. Int. 2012, 32, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Morakinyo, A.O.; Awobajo, F.O.; Adegoke, O.A. Effects of alpha lipoic acid on blood lipids, renal indices, antioxidant enzymes, insulin and glucose level in streptozotocin-diabetic rats. Biol. Med. 2013, 5, 26–33, eISSN: 09748369. [Google Scholar]
- Thirunavukkarasu, V.; Nandhini, A.T.A.; Anuradha, C.V. Lipoic acid attenuates hypertension and improves insulin sensitivity, kallikrein activity and nitrite levels in high fructose-fed rats. J. Comp. Physiol. B. 2004, 174, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Morini, M.; occatagliata, L.; Dell’Eva, R.; Pedemonte, E.; Furlan, R.; Minghelli, S.; Giunti, D.; Pfeffer, U.; Marchese, M.; Noonan, D.; et al. Alpha-lipoic acid is effective in prevention and treatment of experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2004, 148, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Koba, K.; Yanagita, T. Health benefits of conjugated linoleic acid [CLA]. Obes. Res. Clin. Pract. 2014, 8, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Janczy, A.; Steinka, I. Conjugated linoleic acid and the atherosclerosis in animal models—Review. Zesz. Nauk. Akad. Mor. 2012, 74, 23–28. [Google Scholar]
- Sun, J.H.; Boo, P.; Seon, T.J. Biological activities of conjugated linoleic acid [CLA] and effects of CLA on animal products. Livest. Sci. 2007, 110, 221–229. [Google Scholar]
- Lehnen, T.E.; Silva, M.R.; Camacho, A.; Marcadenti, A.; Lehnen, A.M. A review on effects of conjugated linoleic fatty acid [CLA] upon body composition and energetic metabolism. J. Int. Soc. Sports. Nutr. 2015, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Chen, H.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, Y.Q.; Chen, W. Review of the roles of conjugated linoleic acid in health and disease. J. Funct. Foods. 2015, 15, 314–325. [Google Scholar] [CrossRef]
- Zhao, W.S.; Zhai, J.J.; Wang, Y.H.; Xie, P.S.; Yin, X.J.; Li, L.X.; Cheng, K.L. Conjugated linoleic acid supplementation enhances antihypertensive effect of ramipril in Chinese patients with obesity-related hypertension. Am. J. Hypertens. 2009, 22, 680–686. [Google Scholar] [CrossRef]
- Benjamin, S.; Prakasan, P.; Sreedharan, S.; Wright, A.G.; Spener, F. Pros and cons of CLA consumption: An insight from clinical evidences. Nutr. Metab. 2015, 12, 1–20. [Google Scholar] [CrossRef]
- Norris, L.E.; Collene, A.L.; Asp, M.L.; Hsu, J.C.; Liu, L.F.; Richardson, J.R.; Li, D.; Bell, D.; Osei, K.; Jackson, R.D.; et al. Comparison of dietary conjugated linoleic acid with safflower oil on body composition in obese postmenopausal women with type 2 diabetes mellitus. Am. J. Clin. Nutr. 2009, 90, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Houseknecht, K.L.; Van den Heuvel, J.P.; Moya-Camarena, S.Y.; Portocarrero, C.P.; Peck, L.W.; Nickel, K.P.; Belury, M.A. Dietary conjugated linoleic acid normalizes impaired glucose tolerance in the Zucker diabetic fatty fa/fa rat. Biochem. Biophys. Res. Commun. 1998, 244, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Nagao, K.; Inoue, N.; Wang, Y.M.; Yanagita, T. Conjugated linoleic acid enhances plasma adiponectin level and alleviates hyperinsulinemia and hypertension in Zucker diabetic fatty [fa/fa] rats. Biochem. Biophys. Res. Commun. 2003, 310, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Moloney, F.; Yeow, T.P.; Mullen, A.; Nolan, J.J.; Roche, H.M. Conjugated linoleic acid supplementation, insulin sensitivity, and lipoprotein metabolism in patients with type 2 diabetes mellitus. Am. J. Clin. Nutr. 2004, 80, 887–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumann, E.; Carpentier, Y.A.; Saebo, A.; Lassel, T.S.; Chardigny, J.M.; Sébédio, J.L.; Mensink, R.P. Cis-9, trans- 11 and trans-10, cis-12 conjugated linoleic acid [CLA] do not affect the plasma lipoprotein profile in moderately overweight subjects with LDL phenotype B. Atheroscler 2006, 188, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Valeille, K.; Férézou, J.; Amsler, G.; Quignard-Boulangé, A.; Parquet, M.; Gripois, D.; Dorovska-Taran, V.; Martin, J.C. A cis-9,trans-11-conjugated linoleic acid-rich oil reduces the outcome of atherogenic process in hyperlipidemic hamster. Am. J. Physiol. Heart. Circ. Physiol. 2005, 289, 652–659. [Google Scholar] [CrossRef]
- Bukowska, B. Glutation: Biosynteza, czynniki indukujące oraz stężenie w wybranych jednostkach chorobowych [Glutathione: Biosynthesis, inducing factors and concentration in selected medical conditions]. Med. Pr. 2004, 55, 501–509. [Google Scholar]
- Chang, Y.Y.; Chou, C.H.; Chiu, C.H.; Yang, K.T.; Lin, Y.L.; Weng, W.L.; Chen, Y.C. Preventive effects of taurine on development of hepatic steatosis induced by a high-fat/cholesterol dietary habit. J. Agric. Food Chem. 2011, 59, 450–457. [Google Scholar] [CrossRef]
- Szymański, K.; Winiarska, K. Tauryna i jej potencjalne wykorzystanie w terapii [Taurine and its potential therapeutic applications]. Postepy Hig. Med. Dosw. 2008, 62, 75–86. [Google Scholar]
- Abebe, W.; Mozaffari, M.S. Role of taurine in the vasculature: An overview of experimental and human studies. Am. J. Cardiovasc. Dis. 2011, 1, 293–311. [Google Scholar]
- Puerta, C.D.; Arrieta, F.J.; Balsa, J.A.; Botella-Carretero, J.I.; Zanarron, I.; Vazquez, C. Taurine and glucose metabolism: A review. Nutr. Hosp. 2010, 25, 910–919. [Google Scholar] [PubMed]
- Carneiro, E.M.; Latorraca, M.Q.; Araujo, E.; Beltrá, M.; Oliveras, M.J.; Navarro, M.; Berná, G.; Bedoya, F.J.; Velloso, L.A.; Soria, B.; et al. Taurine supplementation modulates glucose homeostasis and islet function. J. Nutr. Biochem. 2009, 20, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Higo, S.; Miyata, S.; Jiang, Q.Y.; Kitazawa, R.; Kitazawa, S.; Kasuga, M. Taurine administration after appearance of proteinuria retards progression of diabetic nephropathy in rats. Kobe. J. Med Sci. 2008, 54, 35–45. [Google Scholar]
- Martynowicz, H.; Skoczyńska, A.; Silber, M.; Andrzejak, R. Rola stresu oksydacyjnego w patogenezie nadciśnienia tętniczego [The role of oxidative stress in pathogenesis of arterial hypertension]. Nadciśnienie Tętnicze 2004, 8, 431–438. [Google Scholar]
- Imae, M.; Asano, T.; Murakami, S. Potential role of taurine in the prevention of diabetes and metabolic syndrome. Amino Acids 2014, 46, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Kulasek, G.; Jank, M.; Sawosz, E. Biologiczna rola tauryny u ssaków [The biological role of taurine in mammals]. Vet. Life 2004, 79, 603–608. [Google Scholar]
- Lorenzo, J.M.; Munekata, P.E.S.; Gómez, B.; Barba, F.J.; Mora, L.; Pérez-Santaescolástica, C.; Toldrá, F. Bioactive peptides as natural antioxidants in food products—A review. Trends. Food Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Baltic, Z.M.; Boskovic, M.; Ivanovic, J.; Janjic, J.; Dokmanovic, M.; Markovic, R.; Baltic, T. Bioactive peptides from meat and their influence on human health. Technologija Mesa 2014, 55, 8–21. [Google Scholar] [CrossRef]
- Borrajo, P.; Pateiro, M.; Barba, F.J.; Mora, L.; Franco, D.; Toldrá, F.; Lorenzo, J.M. Antioxidant and antimicrobial activity of peptides extracted from meat by-products: A review. Food Anal. Methods 2019, 1–15. [Google Scholar] [CrossRef]
- Lafarga, T.; Hayes, H. Bioactive peptides from meat muscle and by-products: Generation, functionality and application as functional ingredients. Meat Sci. 2014, 98, 227–239. [Google Scholar] [CrossRef]
- Stadnik, J.; Kęska, P. Meat and fermented meat products as a source of bioactive peptides. Acta Sci. Pol. Technol. Aliment. 2015, 14, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Darewicz, M.; Borawska, J.; Minkiewicz, P.; Iwaniak, A.; Starowicz, P. Biologicznie aktywne peptydy uwalniane z białek żywności [Biologically active peptides released from food-derived proteins]. Zywn-Nauk. Technol. Ja. 2015, 3, 26–41. [Google Scholar]
- Choe, J.; Seol, K.-H.; Kim, H.-J.; Hwang, J.-T.; Lee, M.; Jo, C. Isolation and identification of angiotensin I-converting enzyme inhibitory peptides derived from thermolysin-injected beef M. longissimus. Asian-Australas J. Anim Sci. 2019, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Arihara, K. Strategies for designing novel functional meat products. Meat Sci. 2006, 74, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Jang, A.; Lee, M. Purification and identification of angiotensin converting enzyme inhibitory peptides from beef hydrolysates. Meat Sci. 2005, 69, 653–661. [Google Scholar] [CrossRef]
- Ryan, J.T.; Ross, R.P.; Bolton, D.; Fitzgerald, G.F.; Stanton, C. Bioactive peptides from muscle sources: Meat and fish. Nutrients 2011, 3, 765–791. [Google Scholar] [CrossRef] [PubMed]
- Przybylski, R.; Firdaous, L.; Châtaigné, G.; Dhulster, P.; Nedjar, N. Production of an antimicrobial peptide derived from slaughterhouse by-product and its potential application on meat as preservative. Food Chem. 2016, 211, 306–313. [Google Scholar] [CrossRef]
- Di Bernardini, R.; Mullen, A.M.; Bolton, D.; Kerry, J.; O’Neil, E.; Hayes, M. Assessment of the angiotensin-I-converting enzyme [ACE-I] inhibitory and antioxidant activities of hydrolysates of bovine brisket sarcoplasmic proteins produced by papain and characterization of associated bioactive peptidic fractions. Meat Sci. 2012, 90, 226–235. [Google Scholar] [CrossRef]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef]
- Jang, A.; Jo, C.; Kang, K.; Lee, M. Antimicrobial and human cancer cell cytotoxic effect of synthetic angiotensin-converting enzyme [ACE] inhibitory peptides. Food Chem. 2008, 107, 327–336. [Google Scholar] [CrossRef]
- Mora, L.; Bolumar, T.; Heres, A.; Toldrá, F. Effect of cooking and simulated gastrointestinal digestion on the activity of generated bioactive peptides in aged beef meat. Food Funct. 2017, 8, 4347–4355. [Google Scholar] [CrossRef] [PubMed]
- Gopi, M.; Kumar, D.; Elaiyaraya, G.; Karthik, K.; Char, H.V.M.; Gautham, K.; Jaydip, R.; Purushothaman, M.R. Dietary essentiality I: Coenzyme Q10 conditionally essential-review. Asian J. Anim. Vet. Adv. 2015, 10, 461–475. [Google Scholar] [CrossRef]
- Gvozdjakova, A.; Takahashi, T.; Singh, R.B.; Meester, F.; Wilson, D.W.; Crane, F.L. New roles of coenzyme q10 in cardiovascular diseases, discovered by a single group. World Heart J. 2014, 5, 159–171. [Google Scholar]
- Siemieniuk, E.; Skrzydlewska, E. Koenzym Q10—biosynteza i znaczenie biologiczne w organizmach zwierząt i człowieka [Coenzyme Q10—biosynthesis and biological role in animal and human organisms]. Postepy. Hig. Med. Dosw. 2005, 59, 150–159. [Google Scholar]
- Shen, Q.; Pierce, J.D. Supplementation of coenzyme Q10 among patients with type 2 diabetes mellitus. Health 2015, 3, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Borekova, M.; Hojerova, J.; Koprda, V.; Bauerova, K. Nourishing and health benefits of coenzyme Q10—A review. Czech. J. Food Sci. 2008, 26, 229–241. [Google Scholar] [CrossRef]
- Garrido-Maraver, J.; Cordero, M.D.; Oropesa-Avila, M.; Vega, A.F.; Mata, M.; Pavon, A.D.; Miguel, M.; Calero, C.P.; Paz, M.V.; Cotan, D.; et al. Coenzyme Q10 therapy. Mol. Syndromol. 2014, 5, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeldt, F.L.; Haas, S.J.; Krum, H.; Hadj, A.; Ng, K.; Leong, J.Y.; Watts, G.F. Coenzyme Q10 in the treatment of hypertension: A meta-analysis of the clinical trials. J. Hum. Hypertens. 2007, 21, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Mohseni, M.; Vafa, M.R.; Hajimiresmail, S.J.; Zarrati, M.; Forushani, A.; Bitarafan, V.; Shidfar, F. Effects of coenzyme q10 supplementation on serum lipoproteins, plasma fibrinogen, and blood pressure in patients with hyperlipidemia and myocardial infarction. Iran. Red. Crescent. Med. J. 2014, 16, e16433. [Google Scholar] [CrossRef]
- Dobenecker, B.; Braun, U. Creatine and creatinine contents in different diet types for dogs—Effects of source and processing. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1017–1024. [Google Scholar] [CrossRef]
- Smith, R.N.; Agharkar, A.S.; Gonzales, E.B. A review of creatine supplementation in age-related diseases: More than a supplement for athletes. F1000 Res. 2014, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.; Naclerio, F.; Allgrove, J.; Jimenez, A. Creatine supplementation with specific view to exercise/sports performance: An update. J. Int. Soc. Sports Nutr. 2012, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- The Irish Sports Council. Creatine. Technical Document. Developed by INDI/SNIG for the Irish Sports Council, 2014. Available online: http://static1.squarespace.com/static/53296b1be4b0f59c2976d2c8/t/554b6f9ee4b0ba2f26130d46/1431007134557/Creatine+Technical+Document.pdf (accessed on 17 July 2019).
- Candrow, D.G.; Chilibeck, P.D. Potential of creatine supplementation for improving aging bone health. J. Nutr. Health Aging 2010, 14, 149–153. [Google Scholar] [CrossRef]
- Nadal, M.; Domingo, J.L. Carcinogenicity of consumption of red meat and processed meat: A review of scientific news since the IARC decision. Food Chem. Toxicol. 2017, 105, 256–261. [Google Scholar]
- Domingo, J.L.; Nadal, M. Carcinogenicity of consumption of red and processed meat: What about environmental contaminants? Environ. Res. 2016, 145, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Ekmekcioglu, C.; Wallner, P.; Kundi, M.; Weisz, U.; Haas, W.; Hutter, H.P. Red meat, diseases, and healthy alternatives: A critical review. Crit. Rev. Food Sci. Nutr. 2018, 58, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Yip, C.S.C.; Lam, W.; Fielding, R. A summary of meat intakes and health burdens. Eur. J. Clin. Nutr. 2018, 72, 18–29. [Google Scholar] [CrossRef]
- Crippa, A.; Larsson, S.C.; Discacciati, A.; Wolk, A.; Orsini, N. Red and processed meat consumption and risk of bladder cancer: A dose-response meta-analysis of epidemiological studies. Eur. J. Nutr. 2018, 57, 689–701. [Google Scholar] [CrossRef] [PubMed]
- De Smet, S.; Vossen, E. Meat: The balance between nutrition and health. A review. Meat Sci. 2016, 120, 145–156. [Google Scholar] [CrossRef]
- Young, J.F.; Therkildsen, M.; Ekstrand, B.; Che, B.N.; Larsen, M.K.; Oksbjerg, N.; Stagsted, J. Novel aspects of health promoting compounds in meat. Meat Sci. 2013, 95, 904–911. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Component | Experimental Model | Study Group | Treatment | Effects | Reference |
|---|---|---|---|---|---|
| Acetyl-l-carnitine | Randomized, phase III, double-blind, placebo-controlled trial | Patients with hypertension, T2DM and dyslipidemia on background statin therapy (n = 229) were randomized to the placebo (n = 113) and acetyl-l-carnitine (n = 116) groups. | 2000 mg acetyl-l-carnitine/d (2 × 1000 mg capsules) for 6 months | BW↔, BMI↔, SBP↓, DBP↔, mean BP↓, glucose↔, HbA1c↑, insulin↔, HOMA-IR↓, GDR↔, TC↑, HDL↓, LDL↔, TG↔, Lp(a)↔, serum creatinine ↔, albuminuria↔, GFR↔ | Parvanova et al. [9] |
| l-carnitine (Eva Pharma, Egypt) | Parallel randomized controlled prospective | T2DM patients on glimepiride (n = 72) were randomized to glimepiride group (n = 34), and glimepiride + l-carnitine group (n = 38). | 2 g l-carnitine/d (1 g twice daily) for 6 months | BMI↔, SBP↔, DBP↔, fasting glucose↓, postprandial blood glucose↓, HbA1c↓, insulin↓, HOMA-IR↓, IRAPe↑, TNF-α↓, Visfatin↓, TC↓, TG↓, HDL↑, LDL↓ | El-sheikh, El-Haggar and Elbedewy [10] |
| l-carnitine commercially available capsules (New Health Taiwan Co., Ltd.). | Single blind, randomized, parallel, placebo-controlled trial | Patients with coronary artery disease (n = 47) were randomly assigned to the placebo (n = 24) and l-carnitine (n = 23) groups. | 1000 mg l-carnitine/d (2 × 500 mg capsules) for 12 weeks | • CRP↓, IL-6↓, TNF-α↓; • levels of inflammation markers were negatively correlated with the levels of LC and antioxidant enzymes activities (SOD, GPx); | Lee et al. [11] |
| L-carnitine Commercially available capsules (New Health Taiwan Co., Ltd.). | Single blind, randomized, parallel, placebo-controlled trial | Patients with coronary artery disease (n = 47) were randomly assigned to the placebo (n = 24) and L-carnitine (n = 23) groups. | 1000 mg l-carnitine/d (2 × 500 mg capsules) for 12 weeks | • CAT↑, GPx↑, SOD↑, MDA↓, l-carnitine↑ • level of L-carnitine was significantly correlated with CAT and SOD activities | Lee et al. [12] |
| l-carnitine tablets (Ultimate Nutrition Company, USA) Coenzyme Q10 soft gel (Vitane’s Nature Company, USA) | Randomized controlled single center clinical trial | Type 2 diabetes patients (n = 75) who treated with oral antidiabetic drugs metformin and sulfonylurea) were randomly assigned into l-carnitine, coenzyme Q10 and control groups. | l-carnitine 1000 mg tablet once daily for 8 weeks | glucose↓, HbA1c↔, TC↓, LDL↓, HDL↔, Lp(a)↓ | Mohammed-Jawad et al. [13] |
| 150 mg coenzyme Q10 soft gel daily (2 × 75 mg) for 8 weeks | glucose↓, HbA1c↓, TC↓, LDL↓, HDL↔, Lp(a)↓ | ||||
| l-carnitine (Lanling Pharmaceutical CO., LTD, China) | Randomized, single-blinded, placebo-controlled clinical study | Patients with MetS (n = 30) were randomly allocated into l-carnitine (n = 15) and control (n = 15) groups | 4 g l-carnitine infusion daily (2 g twice a day) for 7 days | • BW↓, BMI↓, WC↓, HC↓, WHR↓, SBP↔, DBP↔, TC↔,TG↓, HDL↓, LDL↑, ApoA1↓, ApoB↑, ApoA1/ ApoB↓, Lp(a)↑, glucose↓, insulin↓, HOMA-IR↓, CRP↔, UA↑, FFA↑, AST↑, ALT↔, GGT↓ • hunger score in the L-carnitine group was decreased. • reduction physical and mental fatigue and fatigue severity scores during starvatio: improved physical fatigue (l-carnitine vs. control, p < 0.001), mental fatigue (l-carnitine vs. control, p = 0.001), and fatigue severity (l-carnitine vs. control, p < 0.001). | Zhang et al. [14] |
| Carnosine (Flamma S.p.A, Italy) | Pilot randomized, double-blind, placebo-controlled trial | Overweight and obese, non-diabetic individuals (n = 30), were assigned to carnosine and placebo groups | 2 g/day (2 × 1 g) for 12 weeks | adipsin↔, leptin↔, resistin↓ | Baye et al. [15] |
| l-Carnosine capsules (NOW FOODS Company for Natural Products manufactured by GMP Pharma, USA) | Randomized, double-blinded, placebo-controlled trial | Patients (n = 90) with type 1 diabetes, aged 9 to 18 years with at least 5 years disease duration, active diabetic nephropathy in the form of microalbuminuria were randomly assigned into carnosine (n = 45), or matching placeb group (n = 45). Patients in both groups received oral captopril 25 mg tablet | 1 g/d (2 × 500 mg capsule) administered orally for 3 months | BW↔, BMI↔, SBP↔, DBP↔, glucose↔, TG↓, TC↓, HDL↑, HbA1c↓, creatinine↔, UACR↓, Alpha 1-microglobulin↓, TAC↑, MDA↓, serum carnosine↑ | Elbarbary et al. [16] |
| l-Carnosine capsules (Myprotein, UK and Ireland) | Double-blind, randomized, parallel-design, clinical trial | Oral agents for controlling hyperglycemia (n = 54) were randomly assigned into carnosine (n = 27) and placebo (n = 27) group | 1 g/d (2 × 500 mg capsules) after a meal for 12 weeks | BW↔, BMI↔, WC↔, BFM↓, FFM↑, SBP↓, DBP↔, glucose↓, HbA1c↓, insulin↓, HOMA-IR↔, HOMA-β↔, TG↓, TC↔, LDL↔, HDL↔, CML↓, pentosidine↓, s-RAGE↔, TNF-α↓, IL-6↓, IL-1β↔ | Houjeghani, Kheirouri, Faraji and Jafarabadi [17] |
| l-Carnosine capsules (Myprotein, UK and Ireland) | Double-blind, randomized, parallel designed, clinical trial | Patients with T2DM, using only oral agents for controlling hyperglycemia (n = 54) were randomly assigned into carnosine (n = 27) and placebo (n = 27) group | 1 g/d (2 × 500 mg capsules) after a meal for 12 weeks | glucose↓, CAT↑, SOD↔, MDA↓, PC↓ | Houjeghani, Kheirouri, Faraji et al. [18] |
| Coenzyme Q10 (Zahravi Company, Iran) | Randomized, double-blinded, placebo-controlled clinical trial | Diabetic hemodialysis patients were randomly assigned into coenzyme Q10 (n = 30) or placebo (n = 30) groups | 120 mg coenzyme Q10/d (60 mg twice a day) for 12 weeks | TAC↑, GSH↔, MDA↔, CRP↓, NO↑ | Fallah, Askari, Soleimani et al. [19] |
| Coenzyme Q10 soft gel (BY-Health Co Ltd., China) | Randomized, double-blinded, placebo-controlled trial | Dyslipidemic subjects without taking any hypoglycemic or hypolipidemic drugs (n = 101) were randomly assigned to the placebo (n = 50) or coenzyme Q10 (n = 51) groups. | 120 mg coenzyme Q10 daily (2 softgels 30 mg coenzyme Q10 each twice a day) for 24 weeks | BW↔, HC↓, WC↔, BMI↓, SBP↓, DBP↓, TC↔, TG↓, LDL↓, HDL↔, non HDL↔, ApoA1↑, ApoB↔, ApoA1/ApoB↑, glucose↓, insulin↓, HOMA-IR↓, CRP↔, TAC↑, AST↔, ALT↔, GGT↔, urea↔, creatinine↔, UA↔ | Zhang, Yang, Guoet al. [20] |
| Coenzyme Q10 commercially available capsules (New Health Taiwan Co., Ltd., Taiwan) | Single blinded, randomized, parallel, placebo-controlled study | Patients with coronary artery disease with statins therapy for at least 1 month (n = 51) were randomly assigned to the placebo (n = 24) or coenzyme Q10 (n = 27) groups. | 300 mg coenzyme Q10/d for 12 weeks | coenzyme Q10↑, vitamin E↑, SOD↑, CAT↑, GPx↑, CRP↔, TNF-α↓, IL-6↔, adiponectin↔ | Lee, Tseng, Yen and Lin [21] |
| Conjugated linoleic acid - CLA free fatty acids (FFA): cis-9, trans-11 isomer (39 g/100 g) and the trans-10, cis-12 isomer (41 g/100 g) - CLA triacylglycerols: cis-9, trans-11 isomer (38 g/100 g) and the trans-10, cis-12 isomer (38 g/100 g) (Natural Lipids, Norway) | Randomized, double-blind, placebo-controlled study | Healthy overweight volunteer men and women with BMI 25–30 kg/m2 (n = 180) were randomly assigned to placebo (n = 59), CLA-FFA (n = 61) or CLA-triacylglycerol (n = 60) | 4.5 g 80% CLA-FFA (3.6 g active CLA isomers) or 4.5 g 76% CLA triacylglycerols (3.4 g active isomers) for 12 months | BW↓, BMI↓, BFM↓, LBM↑, BMM↓, diet daily intake↓, HbA1c↑, glucose↔, TG↔, TC↔, HDL↓, LDL↓, Lp(a)↑, leukocytes↑, thrombocytes↑, ALT↔, AST↑ | Gaullier, Halse, Høye et al. [22] |
| Conjugated linoleic acid CLA80:20 capsules (Stepan Specialty Products BV, Netherlands) each containing 1 g of oil and 0.05% v/v Tocoblend TM L50 IP (IOI Loders Croklaan, NL) as anti-oxidant | Double-blind, randomized, cross-over, baseline, and placebo controlled human intervention study | Healthy subjects at low and moderate cardiovascular risk (n = 45) assigned to placebo (n = 23) or CLA (n = 22) groups | Four capsules daily for two weeks, crossing over to the other treatment arm after a wash-out of at least four weeks. The dose (4 g/day) provided 2.5 g/day 9c,11t-CLA or 1.1% of energy | • plasma FA: 16:0↔, 18:0↔, 18:1 t11↔, 18:1 c9↔, 18:2 n6↔, 9c,11t-CLA↑, 10t,12c-CLA↑, 9c,11t+10t,12c-CLA↑, 18:3 n3↔, 18:3 n6↔, 20:3 n6↔, 20:4 n6↔, 20:5 n3↔, 22:6 n3↔ | Bachmair, Wood, Keizer et al. [23] |
| Conjugated linoleic acid (Tonalin® WDP 60) cis-9, trans-11; trans-10, cis-12 CLA isomers (50:50 ratios) | Double blind, randomized and placebo controlled study | Healthy sedentary slightly overweight (n = 18), were randomly assigned to CLA (n = 9) and (n = 9) placebo groups | 3 g CLA 3 times dailyfor 30 days | WC↔, HC↔, BFM↔, LMB↔, BMI↔, VO2 peak↔, TC↔, TG↓, VLDL↓, LDL↓, HDL↔, ApoA↔, ApoB↔, ApoB/ApoA↔, leptin↓, glucose↔, insulin↓, HOMA-IR↔, BChE↓, lipoprotein lipase↑ | Bulut, Bodur, Colak and Turnagol [24] |
| Conjugated linoleic acid CLA mixture containing 38.57% of cis-9, trans-11 isomers, and 39.76% of trans-10, cis-12 isomers, in an equal proportion (50:50) (Idealfarma, Brazil) | Placebo-controlled and randomized clinical trial | Women with diagnosed MetS (n = 14) assigned to placebo (n = 7) or glutathione (n = 7) groups | 3 g CLA/day added to strawberry jam for 90 days | glucose↔, insulin↓, HOMA-IR↔, TG↔, TC↔, LDL↔, HDL↔, SBP↔, DBP↔, BFM↓, BW↔, BMI↔, WC↓ | Carvalho, Uehara and Rosa [25] |
| Conjugated linoleic acid CLA mixture of the bioactive isomers 50% cis-9, trans-11 and 50% trans-10, cis-12 (Tonalin) | Randomized, double-blind, placebo-controlled trial | Overweight and grade I obese subjects (n = 80) divided to CLA (n = 40) and placebo (n = 40) groups | 1.7 g CLA in 200 mL of sterilized milk twice a day for 12 weeks | BW↓, BMI↓, LBM↔, BFM↓, WHR↓, internal organ fat↔, SFM↓, GOT↔, GPT↔, TC↔, TG↔, HDL↔, LDL↔, glucose↔, SBP↔, DBP↔ | Chen, Lin, Huang et al. [26] |
| Conjugated linoleic acid - 50:50 mixture of trans 10, cis 12 and cis 9, trans 11 CLA (Clarinol® G-80, Lipid Nutrition) - cis 9, trans 11 (Lipid Nutrition) | Double-blinded, 3-phase crossover clinical trial, placebo-controlled trial | Healthy, overweight, hypercholesterolemic, male volunteers (n = 28) | - 3.5 g/d of a 50:50 mixture of t10, c12 and c9, t11 CLA oil (Clarinol G-80, containing 2.8 g of total CLA) - 3.5 g/d of c9, t11 CLA (c9, t11 CLA oil, containing 2.7 g of total CLA) 3 treatment phases of 8 consecutive weeks each alternated with 4 weeks washout periods | BW↔, BMI↔, BFM↔, LBM↔, TC↔, TG↔, VLDL↔, LDL↔, HDL↔, CRP↔, TNF-α↔, IL-6↔, HOMA-IR↔, adiponectin↔, Ox-LDL↔ | Joseph, Jacques Plourde et al. [27] |
| Creatine monohydrate | Double-blind, randomized, parallel-group, placebo-controlled trial | Men and women prediagnosed with T2DM, physically inactive for at least 1 yr (n = 28), were randomly assigned to the placebo (n = 14) and creatine (n = 14) groups | 5 g/d single dose during lunch for 12 weeks | • HbA1c↓, glucose↓, AUC glucose↓, insulin↔, C-peptide↔, total GLUT-4↔, membrane GLUT-4↑, membrane/total GLUT-4↑, glucose/insulin↔, HOMA-IR↔, HOMA- β ↔, TC↔, TG↔, VLDL↔, LDL↔, HDL↔, apoA1↔, apoA2↔, apoE↔, apoB↔, L(a)↔ • no significant differences were observed between the groups for any physical capacity variable | Gualano, De Salles Painneli, Roschel et al. [28] |
| Glutathione Setria® capsules (Kyowa Hakko USA) | Randomized, double-blinded, placebo-controlled trial | Healthy non-smokers, not taking antioxidant supplements for at least 1 month (n = 61) were randomly assigned to one of three treatment GSH low dose (n = 20), GSH high dose (n = 20) and placebo (n = 21) | 250 mg/d orally (2 × 125 mg capsules) or 1000 mg/day orally (2 × 500 mg capsules) for 6 months | whole-blood GSH↑, erythrocyte GSH↑, plasma GSH↑, lymphocytes GSH↑, buccal cells GSH↑, (GSSG+GSSP):GSH ratio↓, NK cells cytotoxicity↑, lymphocyte proliferation↔, respiratory burst↔, neutrophil phagocytosis↔ | Richie, Nichenametl, Neidig et al. [29] |
| Liposomal Glutathione Tri-Fortify Orange (phosphatidylcholine liposome GSH) (Researched Nutritionals, USA) | Pilot clinical study | Healthy nonsmokers, 50–80 years of age, had no antioxidant supplementation for ≥ 1 month (n = 12). Subjects were randomly assigned to low-dose (n = 6) or high-dose (n = 6) groups | 500 mg, per os 1000 mg per os for 4 weeks | whole-blood GSH↑, erythrocyte GSH↑, plasma GSH, PBMC GSH↑, (GSSG+GSSP):GSH ratio↓, 8-isoprostane↓, NK cell cytotoxicity↑, lymphocyte proliferation↑ | Sinha, Sinha, Calcagnotto et al. [30] |
| l-Glutathione Oxition (NTCPharma, Italy) | Double-blinded, randomized placebo controlled crossover study | Healthy male volunteers with one or more cardiovascular risk factors (n = 16) randomized to the AB (n = 8) and BA (n = 8) groups | Oxition 100 mg twice daily for 4 weeks. Each intervention phase lasted 4 weeks with 4 weeks washout period between the two treatments for a total of 12 weeks | ALT↔, GGT↔, TC↓, TG↔, HDL↓, LDL↓, glucose↑, CysGly plasma↑, CysGly reduced blood↔, GSH plasma↔, GSH reduced plasma↔, GSH total blood↔, GSH reduced blood↔, 3-NT↔, MDA↔, PAS↔, PAD↔, HR↔, RHI↔, FRHI↔, augmentation index↔, augmentation index standardized for heart rate of 75 bpm | Campolo, Bernardi, Cozzi et al. [31] |
| l-Glutathione (KOHJIN Life Sciences, Japan) | Open label, single arm, multicenter, pilot trial | NAFLD patients (n = 34) | 300 mg/d for 4 months by oral administration | BMI↔, glucose↔, IRI↔, HbA1c↑, HDL↔, LDL↔, TG↓, NEFA↓, AST↔, ALT↓, GGT↔, ferritin↓, platelet count↔, type IV collagen 7 s↔, GSH in protein fraction↓, GSH in deproteinized fraction↔, CAP↔, LSM↔ | Honda, Kessoku, Sumida et al. [32] |
| l-Glutathione capsules (KOHJIN Co. Ltd., Japan) | Randomized, double-blind, placebo-controlled clinical trial | Healthy, nonsmoking subjects (n = 40) men and women assigned to the placebo (n = 20) or glutathione (n = 20) groups. | 1 g (2 × 500 mg/d) administered 15 min before breakfast and dinner for 4 weeks | F2-isoP↔, 8-OHdG↔, GSH↔, GSSG↔, | Allen and Bradley [33] |
| α-Lipoic acid | Randomised, double-blind, placebo controlled, prospective study | T2DM patients (n = 23) with diabetic neuropathy and control group- healthy people (n = 21) | 600 mg lipoic acid/d, 30 min prior to meals for 6 weeks | glucose↔, HbA1c↔, TC↔, TG↔, HDL↔, LDL↔, CRP↔, insulin↔, adiponectin↔ | Atmaca, Akbas et al. [34] |
| α-Lipoic acid | Double-blind, placebo-controlled, randomized, clinical trial | Obese patients with NAFLD (n = 50) were randomly allocated to the lipoic acid (n = 25) and placebo (n = 25) groups. | 1200 mg/d (2 × 600 mg capsule, one capsule 20 min before breakfast and one capsule 20 min before dinner) plus 400 mg vitamin E/d for 12 weeks | BW↓, BMI↓, WC↓, HC↓, BFM↓, visceral fat↓, total body water↑, free fat mass↑, bone mass↔, ALT↓, AST↓, glucose↓, insulin↓, QUICKI↓, adiponectin↑, MCP-1↔, IL-6↓, ferritin↓, grade of liver steatosis↓ | Hosseinpour-Arjmand, Amirkhizi, and Ebrahimi-Mameghani [35] |
| α-Lipoic acid capsules (Puritan’s Pride, USA) | Randomized double-blind placebo-controlled clinical trial study | Pregnant women newly diagnosed with gestational diabetes mellitus (n = 60) were divided into drug (n = 30) and placebo (n = 30) groups | 100 mg capsule/d for 8 weeks with lunch | glucose↓, insulin↔, HOMA-IR↓, QUICKI↑, lipoic acid ↑, adiponectin↑, leptin↔, MDA/TAC↓, | Aslfalah, Jamilian, and Khosrowbeygi [36] |
| α-Lipoic acid (produced by Karen Company and capsulated in the School of Pharmacy, Isfahan University of Medical Sciences, Iran) | Randomized, double blind, placebo-controlled clinical trial | Patients with stroke (n = 80) were randomly assigned into lipoic acid (n = 40) or placebo (n = 40) groups | 600 mg lipoic acid/d for 12 weeks | SBP↓, DBP↓, glucose↓, insulin↔ | Mohammadi, Khorvash, Feizi Askari [37] |
| α-Lipoic acid (produced by Karen Company and capsulated in the School of Pharmacy, Isfahan University of Medical Sciences, Iran) | Randomized, double blind, placebo-controlled clinical trial | Patients who experienced a stroke (n = 80) were randomized to the placebo (n = 40) and lipoic acid (n = 40) groups. | 1 capsule containing 600 mg lipoic acid, 1 h before or 2 h after lunch daily for 12 weeks | TG↓, TC↓, LDL↓, HDL↑ | Mohammadi, Khorvash, Feizi, Askari [38] |
| α-Lipoic acid capsules | Randomized, double blind, placebo-controlled clinical trial | Patients with T2DM (n = 35) were included in lipoic acid group and healthy participants (n = 35) were taken as control group | 300 mg/d (2 capsules) for 6 months | BW↔, BMI↔, fasting blood glucose↓, postprandial blood glucose↓, HbA1c↓, LDL↓, HDL↑, VLDL↓, TG↓, TC↓, MDA↓, GSH↑, NO↑ | Panda, Panda, and Mishra [39] |
| Lipoic acid capsules (Puritan’s Pride, USA) | Randomized double-blind placebo-controlled clinical trial study | Women with gestational diabetes mellitus (n = 60) were divided into drug (n = 30) and placebo (n = 30) groups randomly | 100 mg capsule/d for 8 weeks with lunch | glucose↓, α-lipoic acid ↑, ALT↓, AST↔, ALP↔, GGT↓, urea↑, creatinine↔, UA↔, MDA/TAC↓ | Aslfalah, Jamilian, Rafiei and Khosrowbeygi [40] |
| Taurine (independent third-party pharmacy) | Single-center, double-blind, randomized, placebo-controlled trial | Untreated participants (n = 120) with prehypertension assigned to placebo (n = 60) taurine (n = 60) groups and age-matched normotensive control subjects without taurine supplementation (n = 58) | 1.6 g/d for 12 weeks | clinic SBP↓, clinic DBP↓, 24 h ambulatory SBP↓, 24 h ambulatory DBP↓, FMD↑, NMD↑, plasma: H2S↑, taurine↑ | Sun, Wang, Li et al. [41] |
| Taurine capsules (Landesapotheke, Austria) | Randomized, controlled, double blind trial | Patients with hepatic venous pressure gradient (HVPG) (n = 30) were randomly assigned into taurine (n = 15) or placebo (n = 15) groups | 6 g (6 capsules a 1000 mg) for 4 weeks | HVPG↓, FHVP↔, WHVP↔, creatinine↔, BUN↔, bilirubin↔, albumin↔, AST↔, ALT↔, GGT↔, PPT↔, CRP | Schwarzer, Kivaranovic, Mandorfer et al. [42] |
| Taurine (Taisho Pharmaceutical, Japan) | Multicentre, open-label, phase III trial | 10 patients with MELAS (mitochondrial myopathy, encephalopathy, lactic acidosis and stroke-like episodes) | 9 g/d (participants 25–39 kg BW) or 12 g/d (participants ≥ 40 kg BW) for 52 weeks | • plasma taurine↑, CSF taurine↑, serum lactate↔, CSF lactate↔, serum pyruvate↔, CSF pyruvate↔ • reduction of the annual relapse rate of stroke-like episodes from 2.22 to 0.72 • five patients showed a significant increase in the taurine modification of mitochondrial tRNALeu(UUR) from peripheral blood leukocytes | Ohsawa, Hagiwara, Nishimatsu et al. [43] |
| Component | Experimental Model | Treatment | Effects | Reference |
|---|---|---|---|---|
| Acetyl-l-Carnitine (Sigma-Tau, Italy) | Pathogen-free male Wistar rats with oxidative stress induced by NaAsO2 intoxication (20 mg/kg) | Orally administered 300 mg Acetyl-l-Carnitine/kg, 1 h prior to NaAsO2 for 28 days. | • AST↓, ALT↓, LDH↓, bilirubin↓; • oxidant/antioxidant organs status (kidney, liver, heart, lung, brain): GST↑, SOD↑, CAT↑, TBARS↓, -SH↓; • significantly suppressed oxidative organs damage; | Sepand, Razavi-Azarkhiavi, Omidi at al. [44] |
| l-carnitine (Solgar Vitamin and Herb, USA) | Male Wistar Albino rats fed cholesterol rich diet (7.5% cholesterol) | L-carnitine aqueous solution 75 mg/L for 40 days. | TBARS↓, GSH↑, SOD↑, GPx↔, CAT↔ | Keskin, Uluisik and Altin [45] |
| l-carnitine (MEPACO, Egypt) | New Zealand rabbits | Diets contained 25, 50 and 100 mg l-carnitine/kg for 4 weeks. | • blood constituents: TC↓, TG↓, HDL↑, LDL↓, VLDL↓, glucose↑; • metabolites: creatinine↑ • plasma enzymes activity: AST↓, ALT↓, ALP↓; • electrolytes: Na↓, K↑, Cl↑; • hormones: T3↑, T4↑, cortisol↓. | Elgazzar, Ghanema and Kalaba [46] |
| l-carnitine | Rats with oxidative stress induced by aspartame intoxication (75 mg/kg or 150 mg/kg) | Oral dose 10 mg l-carnitine/kg for 30 days | • TG↓, TC↓, HDL↑, LDL↓, VLDL↓, ALT↓, AST↓, ALP↓, LDH↓, GGT↓, total proteins↑, albumin↑, CRP↓, TNF-α↓, IL-6↓; hepatic: MDA↓, SOD↑, CAT↑, GPx↑, GSH↑; serum hepatic: MPO↓, XO↓ • more percentage of intact liver cells with undamaged DNA and fewer comet cells • decrased area of damaged cells in liver, obvious improvement liver histology | Hamza, Al-Eisa, Mehana, El-Shenawy et al. [47] |
| l-carnitine (Northeast Pharmaceutical Factory, China) | Male Kunming SPF mice with induced diabetes by high-calorie diet (20% sugar, 18% lard) and two low doses of STZ (100 mg/kg, i.p.) at age of 6 and 9 weeks. | High 250 mg l-carnitine/kg i.g. dose or low 125 mg l-carnitine/kg i.g. dose for 3 weeks. | • BW↓, liver weight↓; • liver: FFA↔, TG↓, L-carnitine↔, Acetyl-L-carnitine↓; • plasma: TG↔; • reduced numer lipid droplet deposits in hepatocytes • recovered mitochondrial damage | Xia, Li, Zhong et al. [48] |
| l-Carnosine (Sigma-Aldrich, USA) | Male Wistar rats with mimic natural agening induced by applying d-galactose subcutaneously as 300 mg/kg, 5 days/week for 2 months | 250 mg/kg, i.p. 5 days/week for 2 months | total testosterone↔; testicular: ROS↓, TBARS↓, DC↓, PC↓, AOPP↓, AGE↓, FRAP↔, GSH↔, SOD↔, GPx↔, GST↔ | Aydın, Küçükgergin, Çoban et al. [49] |
| l-Carnosine (Sigma-Aldrich, USA) | Male Wistar rats with induced diabetes by high fat diet (60% of total calories from fat) and single STZ injection at a dose of 40 mg/kg BW | 250 mg/kg BW i.p. 5 times a Week for last 4 weeks of study | • BW↔, liver weight↔ • serum: glucose↔, HbA1c↔, TG↓, TC↓, ALT↓, AST↓, LDH↓ • serum/plasma: ROS↓, MDA↔, i-MDA↓, AOPP↓, AGE↓, FRAP↔ • hepatic: TG↓, TC↔, ROS↓, MDA↓, PC↓, AOPP↔, AGE↓, FRAP↔, GSH↔, SOD↔, CAT↔, GPx↔; mRNA expression of hepatic SOD↔, GPx↔ • liver histopathologic scoring steatosis↓, lobular inflammation↔ and hepatocyte ballooning↓ | Aydın, Bingül, Küçükgergin et al. [50] |
| l-Carnosine (Sigma-Aldrich, USA) | Male Wistar rats with induced diabetes by high fat diet (34.3–60% fat of total calories) and STZ injection at a dose of 40 mg/kg BW | 250 mg/kg BW i.p. 5 times a week for 4 weeks | • BW↔, kidney weight↔ • blood: glucose↔, HbA1c↔, TG↓, TC↓, • serum: BUN↓, creatinine↓, total protein↔, albumin↔ • kidney: ROS↓, MDA↓, PC↓, AOPP↓, AGE↓, FRAP↔, GSH↔, SOD↔, CAT↔, GPx↔; mRNA expression of kidney SOD↔, GPx↔ • Histopathologic examination of kidney tissue showed normal appearance of glomeruli and tubules in all rat groups | Aydın, Küçükgergin, Bingül et al. [51] |
| l-Carnosine (Sigma-Aldrich, USA) | Aged (20 months-of-age) male Wistar rats | 250 mg/kg/5 days per week; i.p. for 2 months | • serum/plasma: AGE↓, PC↓, AOPP↓, MDA↓, FRAP↔, ROS↓ • liver: AGE↓, PC↓, AOPP↓, MDA↓, FRAP↔, ROS↓ | Bingül, Yılmaz, Aydın et al. [52] |
| Coenzyme-Q10 (Mepaco company, Egypt). | Male albino rats fed cholesterol rich diet (5% cholesterol) | 1mg coenzyme Q10/rat by oral gavage for 4 months | • TG↓, TC↓, HDL↑, LDL↓, SOD↑, CAT↑, GPx↑, MDA↓, • amelioration histological and biochemical structure of cerebellal cortex | El-Haleem, Yassen, and Raafat [53] |
| Creatine monohydrate | Male Sprague-Dawley rats with NAFLD induced by high-fat liquid diet with 71% of energy derived from fat | Free access to food diet with 1% (w/v) creatine monohydrate throughout the 3 weeks | • BW↔, • liver: fat↓, TG↓, TC↓, TBARS↓, SAM↑, SAH↔, SAM/SAH↑, phosphatidylcholine↔, phosphatidylethanolamine↑; mRNA levels: Pemt↔, PPARα↑, CD36↓, CPT1a↑, LCAD↑, Bhmt↓, Gnmt↓, MGAT↓ • plasma: glucose↔, insulin↔, creatine↑, GAA↓, Hcy↔, Cys↑, • kidney: AGAT↓ | Deminice, da Silva, Lamarre et al. [54] |
| Creatine monohydrate | Male Wistar rats with nonalcoholic steatohepatitis (NASH) induced by choline-deficient diet | 2% (w/v) creatine monohydrate in diet (free access to food) for 4 weeks | • BW↔, food intake↔ • plasma: creatine↑, Hcy↓, methionine↔, Cys↔, phosphatidylcholine↔, ALT↓, TNF-α↓ • liver: fat↓, TG↓, TC↓, creatinie↑, SAM↔, SAH↑, SAM/SAH↔, phosphatidylcholine↔, MDA↓, GSH↑, GSH/GSSG↑, TNF-α↓, PPARγ↔, • mRNA genes expression: - methionine metabolism: Bhmt1↑, Cbs↔, Pemt↔, Gnmt↑ - phospholipids metabolism: Chka↔, Chkb↔, ChDh↓, Pcyt1a↔ - MTP↔ - transcription factors: PPARα↓, PPARγ↔, - fatty acid oxidation genes: UCP2↓, PGC1a↔, LCAD↑, CPT1a↓, FABP3↔, HAD↔ • kidney: AGAT↓ | Deminice, de Castro, Francisco et al. [55] |
| Creatine monohydrate | Sprague–Dawley rats with NAFLD induced by HFD (0.82 kcal/g protein, 3.24 kcal/g fat and 1.43 kcal/g carbohydrate for a total of 5.49 kcal/g) | 2% creatine monohydrate in diet (20 g/kg) for 4 weeks | • BW↔, calorie intake↔, • liver: weight↔, TG↓, cholesterol ester↓, MTTP↔, • liver cytokines: Eotaxin↔, EGF↔, Fractalkine↔, IFN-γ↔, IL-1α↔, IL-1β↔, IL-2↔, IL-4↔, IL-5↔, IL-6↔, IL-10↓, IL-12(p70) ↔, IL-13↔, IL-17A↔, IL-18↔, IP-10↔, GRO/KC↔, TNF-α↔, G-CSF↔, GM-CSF↔, MCP-1↔, leptin↔, LIX↔, MIP-1α↔, MIP-2↔, RANTES↔, VEGF↔. • plasma: appearance over time TG↔, ApoB48↑, ApoB100↔; fasting: TG↔, ApoB48↔, ApoB100↔; AUC: TG↔, ApoB48↔, ApoB100↔. • mitochondrial respiratory chain complexes: VDAC loading control↑, VDAC loading control: complex I↔, II↔, III↔, IV↔, V↔; PDI loading control: complex I↔, II↑, III↔, IV↔, V↔; ND6 DNA↔, ATP6 DNA↔ | da Silva, Leonard and Jacobs [56] |
| α-Lipoic acid (Hi-Media chemicals, India) | Male Sprague-Dawley albino rats with fructose-induced experimental cataract (10% w/v fructose solution in drinking water-equivalent to a diet containing 48–57% fructose) for 8 weeks | 20 or 40 mg lipoic acid/kg/d orally by gavage for 8 weeks | • MAP↓, glucose↓, lens: GPx↑, CAT↑, SOD↑, GSH↑, MDA↓, total proteins↑, Ca2+ ATPase activity↑, Ca2+↓, • potentially reduced progression of cataract formation: stage of cataract↓, delayed progression of cataract formation | Khan, Choudhary, Vishwakarma et al. [57] |
| α-Lipoic acid powder (Sigma, USA) | Wistar rats with alloxan induced diabete | 100 mg lipoic acid/kg/d BW i.p. injection for 6 weeks | • serum: GPx↑, CAT↑, MPO↓, MDA↓, GSH↑, glucose↓, urea↓, creatinine↓ • liver: : GPx↔, CAT↑, MPO↓, MDA↓, GSH↑, • kidney: : GPx↑, CAT↑, MPO↓, MDA↓, GSH↑, mRNA levels: SOD↑, CAT↑ GPx↑, • histopathological lesions such as increased glomerularvolume and lymphocyte infiltration were attenuated | Jamor, Ahmadvand, Ashoory and Babaeenezhad [58] |
| α-Lipoic acid | C57BL6 mice with obesity induced by high-fat diet (60% kcal% fat) | 0.2% lipoic acid in diet for 12 weeks | BW↓, food intake↓, caloric intake↓, % body fat↔, LBM↓, BFM↓ | Panzhinskiy, Bashir, Bagchi and Nair [59] |
| α-Lipoic acid powder (Sigma, USA) | Male Sprague Dawley rats with diabetes inducted with injection of 100 mg/kg alloxan | 100 mg lipoic acid/kg was injected i.p. daily for 6 weeks | glucose↓, TG↓, TC↓, HDL↑, LDL↓, VLDL↓, PON1↑ | Jamor, Ahmadvand, Birjandi and Sharafabad [60] |
| α-Lipoic acid | Diabetic Goto-Kakizaki rats fed HFD (7.5% cocoa butter, and 1.25% cholesterol) | 50 mg/kg BW i.p., 3 days/week for 3 months | • BW↔, liver weight↓, fasting blood glucose↓, blood glucose 2 h after load↔, TC↓, non-HDL↓, TG↓, albumin↔, T-Bilirubin↔, AST↓, ALT↔, ALP↓, GGT↓, HEF↑, MDA↓, 8-OHdG↓, UA↓ • liver: TC↓, TG↓, GPx↑, GRd↑, MDA↓, GSH↑, Nrf2↑, TNF-α↓, | Sena, Cipriano, Botelho and Seiça [61] |
| Liposomal Glutathione (8.25% GSH (84.5 mg/mL), 75.15% deionized water, 15% glycerin, 1.5% lecithin, and 0.1% potassium sorbate (% w/w) | Atherosclerotic apolipoprotein E-deficient (E0) mice | 12.5 or 50 mg/kg/d in drinking water for 2 months | • TC↓, HDL↓, TG↑, glucose↔, AAPH induced serum lipid peroxidation↓, • mouse peritoneal macrophages (MPM): GSH↑, PON2 lactonase activity↑, total peroxides↓, LDL uptake↓, Ox-LDL uptake↓, cholesterol biosynthesis↓, HDL-mediated macrophage cholesterol efflux↑, TC↓, atherosclerotic lesion area↓ | Rosenblat, Volkova, Coleman and Aviram [62] |
| Liposomal Glutathione “ReadiSorb” glutathione (Your Energy Systems, LLC, USA) | Male, New Zealand white rabbits | Orally administered 5 mL of liposomal glutathione (containing approximately 428.8 mg of GSH) for 3, 7 or 14 days | LVEDP↓; LVDP↓; CPP↓; total GSH: heart↑, liver↑, brain↔; cTnI↔; heart MDA↔ | Lauver, Kaissarian and Lucchesi [63] |
| Peptides (protein hydrolysate, Phe-Gln-Pro and Phe-Gln-Pro-Ser) protein hydrolysate from meat of Kacang goat (Capra aegagrus hircus) was obtained by Protamex® and Flavourzyme® digestion | Male SHR | Single oral administration: - 0.01 or 0.1 g hydrolysate Kacang goat meat/kg BW - 0.00195 g Phe-Gln-Pro/kg BW - 0.00239 g Phe-Gln-Pro-Ser/kg BW | • after administering 0.01 g or 0.1 g hydrolysate/kg BW highest reduction of SBP was 19.3 or 26.9 mmHg, occurred at 6 h after administration. SBP was still significantly lower than that of the control group after 24 h. • Phe-Gln-Pro showed the highest reduction of SBP by 12.6 mm Hg at 6 h • Phe-Gln-Pro-Ser showed the highest reduction of SBP by 10.6 mmm Hg at 8 h after administration • SBP 24 h after pure peptides administration was not different to the controls | Mirdhayati, Hermanianto, Wijaya et al. [64] |
| Peptides Three sample extracts of pooled fractions from Spanish dry-cured hams | Male SHRs | Single oral administration 4.56 mg of sample 1/kg BW or 1.48 mg of sample 2/kg BW or 8.7 mg of sample 3/kg BW by gastric intubation with a metal tube | • All samples decrase SBP: - sample 1 by 33.1 mm Hg and 38.38 mm Hg after 4 and 6 h; - sample 2 by 27.48 mm Hg after 6 h - sample 3 by 23.56 mm Hg at 6 h after oral administration. • In all cases SBP returned to pretreatment values after 24 h. | Escudero, Aristoy, Nishimura et al. [65] |
| Peptides (RPR, KAPVA and PTPVP) peptides identified in pork meat hydrolysate after in vitro digestion | Male SHRs | Single administration of distilled water peptide suspension 1 mg peptide/kg of BW by gastric intubation. | • analysed peptides decrase mean SBP compared with the control SHRs: - RPR decrease 33.21, 28.81 and 21.16 mm Hg at 6, 8 and 4 h after administration - KAPVA decrease 19.1 and 33.72 at 4 and 6 h after administration - PTPVP decreased by 24.52 and 25.66 mm Hg at 4 and 6 h after administration • in all cases SBP returned to pretreatment values after 24 h. | Escudero, Toldrá, Sentandreu et al. [66] |
| Peptides (KRVITY, Lys-Arg-Val-Ile-Gln-Tyr; VKAGF, Val-Lys-Ala-Gly-Phe) identified in pork loin muscle after extraction and pepsin hydrolysis | SHRs | 10 mg KRVITY or VKAGF/kg BW with a metal oral syringe | • KRVITY decrease SBP by 12 mmHg in 3 h and 23 mmHg in 6 h after oral administration • VKAGF decrease SBP by 12 mmHg in 3 h and 17 mmHg in 6 h after oral administration | Muguruma, Ahhmed, Katayama et al. [67] |
| Peptides (YYRA, Tyr-Tyr-Arg-Ala) identified in chicken bone after extraction and hydrolysis with pepsin | SHRs | Single oral administration 10 mg/kg BW administered orally by intubation. | SBP decrase significantly over a short period of time 3 h from 3rd to 6th h | Nakade, Kamishima, Inoue et al. [68] |
| Peptides low fraction hydrolysate from chicken legs collagen obtained by extraction and digestion with protease | Male SHRs | 3 g hydrolysate/kg BW single administration or long-term administration for 4 weeks | • after single administration reduction in blood pressure was observed from 4 to 8 h • long-term administration showed that there was a reduction in from 2nd to 4th week of the study | Saiga, Iwai, Hayakawa et al. [69] |
| Taurine (Sigma Chemical Co., USA) | Male albino rats (Rattus norvegicus) i.p. injected with 5-fluorouracil (20 mg/kg BW/day) for 7 days. | 50 mg/kg BW/day for 21 days: 7 days alone, 7 days parallel with i.p. injections with 5-fluorouracil, 7 days alone | • BUN↓, creatinine↓, UA↓, SOD↑, CAT↑, GPx↑, MDA↓, GGT↑, ALP↑ • reversed most histological and ultrastructural alterations in kidney tissues | Yousef and Aboelwafa [70] |
| Taurine | Male Wistar rats fed high fructose diet (60% fructose) | 2% taurine solution ad libitum for 30 days | BW↔, SBP↓, kallikrein: heart↑, kidney↑, plasma↑, urine↑; sodium: plasma↓, urine↑ | Nandhini and Anuradha [71] |
| Taurine (Taisho Pharmaceutical, Japan) | Male Golden Syrian hamsters fed high-fat diet (0.05% cholesterol and 10% coconut oil). | Taurine dissolved in drinking water at 1% (w/v) was freely available for 14 days | • BW↔, TC↓, non-HDL↓, HDL↔, TG↓, phospholipids↓, ACAT↓, HMG–CoA reductase↔, cholesterol 7a–hydroxylase↑, • up-regulation LDL receptor activity • acceleration receptor-mediated LDL turnover | Murakami, Kondo, Toda et al. [72] |
| Taurine (Sigma Chemicals, USA) | Male Wistar rats with oxidative injuries induced by Fipronil supplementation 19.4 mg/kg for 5 days (6–10th day of the experiment). | Oral dose 50 mg/kg daily (5 days before and 5 days along with Fipronil supplementation) | • liver: MDA↓, NO↓, GSH↑, GPx↑, SOD↑, CAT↑, AST↓, ALT↓, ALP↓, LDH↓, TC↓, • kidney: MDA↓, NO↓, GSH↑, GPx↑, SOD↑, CAT↑, urea↓, creatinine↓ • amelioration and normalization of the harmful effects of Fipronil on hepatorenal injury | Abdel-Daim, Dessouki, Abdel-Rahman et al. [73] |
| Taurine (Sigma–Aldrich Chemical Company, USA) | Male Wistar rats with diabetes and testicular damage induced by one i.p. injection of 50 STZ mg/kg BW | 100 mg/kg BW daily, via oral gavage, for 6 weeks. | • glucose↓, insulin↑, testis weight/BW↑, MDA↓, protein carbonylation↓, GSH/GSSG↑, SOD↑, CAT↑, TNF-α↓, IL-1β↓, IL-6↓, MCP-1↓, ICAM-1↓, VCAM-1↓, testosterone↑, 3β-HSD↑, 17β-HSD↑, SDH↑ • testicular tissue: ER stress related pathway: calpain-1↓, cleaved Caspase-12↓, p-PERK↓, p-eIF2α/total eIF2α↓, CHOP↓, Grp78↓; NFκB mediated pathway: nuclear NFκB↓, cytosolic NFκB↑, phospho and total I ĸBα↓; mitochondria dependent apoptotic pathways: Bax/Bcl-2↓, cytosolic cytochrome-C↓, mitochondrial cytochrome-C↑, cleaved Caspase-9↓, cleaved Caspase-3↓, cleaved PARP↓ • treatment with taurine improve histological alterations like loss of spermatids, disappearance of testicular cells like Leydig and Sertoli cells, sloughing of centrally located spermatozoa and the disruption of germinal epithelium. | Ghosh, Chowdhury, Das et al. [74] |
| Taurine (Sigma-Aldrich, USA) | Male BALB/c mice with Colistin (15 mg/kg/d, i.p. for 7 consecutive days) associated renal injury | 500 or 1000 mg/kg/d, i.p for 7 consecutive days | • BUN↓, creatinine↓, kidney: ROS↓, TBARS↓, TAC↑ GSSG↓, GSH↑, GSH/GSSG↑, histopathological SQS↓ • mitochondrial: dehydrogenases↑, swelling↓, depolarization↓, ATP↑, TBARS↓, GSH↑, GSSG↓, GSH/GSSG↑ | Heidari, Behnamrad, Khodami et al. [75] |
| Taurine (Sigma Chemical Co., USA) | Male Wistar rats with hypertension induced by L-NAME at 40 mg/kg BW p.o. daily | 100 and 200 mg/kg p.o. for 28 days | • SBP↓, DBP↓, MAP↓, BW↔, OSI of the testes↔, OSI of the epididymis↔, ACP↑, ALP↑, LDH↑, LH↑, FSH↑, testosterone↑, • testes: SOD↑, CAT↑, GPx↑, GSH↑, H2O2↓, MDA↓, MPO↓, NO↑ • epididymis: SOD↔, CAT↑, GPx↑, GSH↔, H2O2↓, MDA↓, MPO↓, NO↑ • sperm: testicular sperm number↑, epididymal sperm number↑, motility↑, viability↔, abnormalities↔ | Adedara, Alake, Adeyemo et al. [76] |
| Taurine (Sigma-Aldrich, USA) | Male Wistar albino rats with malathion induced toxicity (27 mg/kg orally) | 0.5 mL taurine solution at dose of 50, 100, and 200 mg/kg orally for 30 days | • blood: MDA↓, GSH↑ • erythrocyte: SOD↓, CAT↔ • serum: AChE↑ • liver: MDA↓, GSH, SOD↓, CAT↓, AChE↑, mRNA levels: IFN-γ↓, NFĸB↓, TNF-α↓, IL-1β↓, • testis: MDA↓, GSH↑, SOD↓, CAT↓, • brain: MDA↓, GSH↑, SOD↓, CAT↓, • kidney: MDA↓, GSH↑, SOD↓, CAT↓, • preventive action against malathion-induced histopathological changes in rat tissues. | Ince, Arslan-Acaroz, Demirel et al. [77] |
| Taurine (Sigma-Aldrich, USA) | Male Wistar rats with diabetes induced by a single i.p. injection of 40 mg STZ/kg BW | 50 mg/kg BW for 60 days | AChE↓, GnRH↓, TRH↑, T3↑, T4↑, TSH↓, testosterone↑, FSH↓, LH↓, sperm count↑, abnormal sperms↓, motility↑, • brain: MDA↓, SOD↑, CAT↑, • thyroid: MDA↓, SOD↑, CAT↑, • testis: MDA↓, SOD↑, CAT↑ • marked repairing of testicular abnormalities and a maximum healing effect against STZ induced testicular damage | Mohamed and Gawad [78] |
| Taurine (Sigma-Aldrich, USA) | Male Wistar rats with cognitive impairment induced by intracerebroventricular STZ injection at a dose of 3 mg/kg | 40, 60 and 120 mg/kg p.o. by gavage for 28 days | • BW↔, • cortex: GSH↑, MDA↓, NO↓, SOD↑, AChE↓, BChE↓, TNF-α↓, IL-1β↓, ROCK-II↓, GSK-3β↔, ChAT↔ • hippocampus: GSH↑, MDA↓, NO↓, SOD↑, AChE↓, BChE↓, TNF-α↓, IL-1β↓, ROCK-II↓, GSK-3β↔, ChAT↑ • improved behavioural parameters: escape latency↓, time spent in target quadrant↑, retention transfer latency in elevated plus maze test↓, transfer latency in passive avoidance test↑ | Reeta, Singh and Gupta [79] |
| Product | Content (mg/100 g) | |
|---|---|---|
| l-carnitine Kalpana [82]; Dayanand et al. [85] | Taurine Lourenco and Camilo [86] | |
| Kangaroo meat | 637 | n.e. |
| Horse meat | 423 | n.e. |
| Lamb | 190 | 43.8 |
| Veal | n.e. | 39.8 |
| Steak tartare | 183 | n.e. |
| Beef | 139–143 | 43.1 |
| Pork | 25–60.8 | 61.2 |
| Beef ribs | 226 | n.e. |
| Pork ribs | 40.2 | n.e. |
| Duck, fillet | 73.2 | n.e. |
| Turkey fillet | 51.4–200 | 29.5 |
| Chicken fillet | 13–34.4 | 17.8 |
| Pheasant breast | 13.5 | n.e. |
| Beef liver | 15.6 | 69 |
| Pork liver | 10.7 | 89 |
| Poultry liver | n.e. | 110 |
| Eggs | 0.8 | n.e. |
| Cow milk, 1.5% fat | 40 | 1 |
| Goat milk | n.e. | 7 |
| Buttermilk | 38.5 | n.e. |
| Sour cream | 19.7 | n.e. |
| Hard cheese | 2.8 | n.e. |
| Brie | 5.85 | n.e. |
| Feta | 14.9 | n.e. |
| Tuna | n.e. | 68 |
| Cod | n.e. | 31 |
| Atlantic salmon | n.e. | 130 |
| Oysters | n.e. | 396 |
| Products | Content (mg/100 g) Aristoy and Toldra [92] |
|---|---|
| Pork loin | 313 |
| Pork ham | 449 |
| Beef loin | 375 |
| Lamb shoulder | 39.3 |
| Chicken breast | 180 |
| Chicken thigh | 63 |
| Turkey wings | 66 |
| Salmon | 0.53 |
| Trout | 1.6 |
| Sardine | 0.1 |
| Product | Choline (mg/100 g) Patterson et al. [98] | Alpha-Lipoic Acid (mg/100 g) Schmid [4] | CLA (mg/g fat) Koba and Yanagita [117] |
|---|---|---|---|
| Poultry, chicken liver | 190 | n.e. | n.e. |
| Poultry, turkey liver | 220 | n.e. | n.e. |
| Poultry, turkey heart | 130 | n.e. | n.e. |
| Poultry, turkey meat | n.e. | n.e. | 2.0–2.5 |
| Poultry, broiler meat | 66 | n.e. | 0.7–1.5 |
| Turkey sausage, fresh | 51 | n.e. | n.e. |
| Pork sausage, fresh | 53 | n.e. | n.e. |
| Poultry frankfurters | 51 | n.e. | n.e. |
| Ground pork | 69 | 0.02–0.03 | 0.6 |
| Pork, neck | 79 | 0.02–0.04 | n.e. |
| Pork, liver | n.e. | 0.06–0.08 | n.e. |
| Pork, heart | n.e. | 0.11–0.16 | n.e. |
| Lean beef, ground | 66 | n.e. | 4.3 |
| Beef, liver | 330 | 0.06–0.11 | n.e. |
| Beef, neck | 100 | n.e. | n.e. |
| Beef, heart | n.e. | 0.07–0.10 | n.e. |
| Mutton | n.e. | n.e. | 5.6 |
| Veal, muscle | n.e. | 0.01–0.02 | 2.7 |
| Veal, liver | 310 | 0.03–0.05 | n.e. |
| Veal, heart | n.e. | 0.05–0.07 | n.e. |
| Lamb, liver | n.e. | 0.07–0.08 | n.e. |
| Lamb, muscle | n.e. | 0.02–0.04 | 4.3–19.0 |
| Lamb, heart | n.e. | 0.05–0.07 | n.e. |
| Reindeer meat, ground | 150 | n.e. | n.e. |
| Whole eggs, fresh | 250 | n.e. | n.e. |
| Egg yolk | 680 | n.e. | 0.6 |
| Cheddar | 17 | n.e. | 4.0–5.3 |
| Whole milk, 3.25% fat | 14 | n.e. | 5.5 |
| Feta | n.e. | n.e. | 4.9 |
| Butter | 19 | n.e. | 4.7 |
| Cream | n.e. | n.e. | 4.6–7.5 |
| Product | Content (mg/100 g) Bukowska [130] |
|---|---|
| Cooked ham | 23.3 |
| Fried chicken | 13.1 |
| Fried beef | 17.5 |
| Fried bacon | 5 |
| Beefsteak | 12.3 |
| Tuna in oil | 1.1 |
| Spinach | 11.4 |
| Carrot | 5.9 |
| Potatoes | 11 |
| Asparagus | 28.3 |
| Oranges | 7.3 |
| Bananas | 3.3 |
| Product | Content (mg/100 g) | |
|---|---|---|
| Coenzyme Q10 Borekova et al. [159] | Creatine Schmid [4] | |
| Beef | 3.65 | n.e. |
| Beef, semitendinosus muscle | n.e. | 401 |
| Lamb | n.e. | 278–511 |
| Pork, ham | 2 | 247–374 |
| Reindeer meat | 15.7 | n.e. |
| Chicken | 1.4 | n.e. |
| Beef, heart | 11.3 | 298 |
| Beef, liver | 3.9 | 16 |
| Pork, heart | 12.6 | n.e. |
| Pork, liver | 2.27 | n.e. |
| Herring | n.e. | 650–1000 |
| Salmon | n.e. | 450 |
| Tuna | n.e. | 400 |
| Cod | n.e. | 300 |
| Eggs | 0.12 | n.e. |
| Milk, 1.5% fat | 0.01 | n.e. |
| Edam cheese | 0.12 | n.e. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulczyński, B.; Sidor, A.; Gramza-Michałowska, A. Characteristics of Selected Antioxidative and Bioactive Compounds in Meat and Animal Origin Products. Antioxidants 2019, 8, 335. https://doi.org/10.3390/antiox8090335
Kulczyński B, Sidor A, Gramza-Michałowska A. Characteristics of Selected Antioxidative and Bioactive Compounds in Meat and Animal Origin Products. Antioxidants. 2019; 8(9):335. https://doi.org/10.3390/antiox8090335
Chicago/Turabian StyleKulczyński, Bartosz, Andrzej Sidor, and Anna Gramza-Michałowska. 2019. "Characteristics of Selected Antioxidative and Bioactive Compounds in Meat and Animal Origin Products" Antioxidants 8, no. 9: 335. https://doi.org/10.3390/antiox8090335