Carotenoids and Markers of Oxidative Stress in Human Observational Studies and Intervention Trials: Implications for Chronic Diseases


Abstract
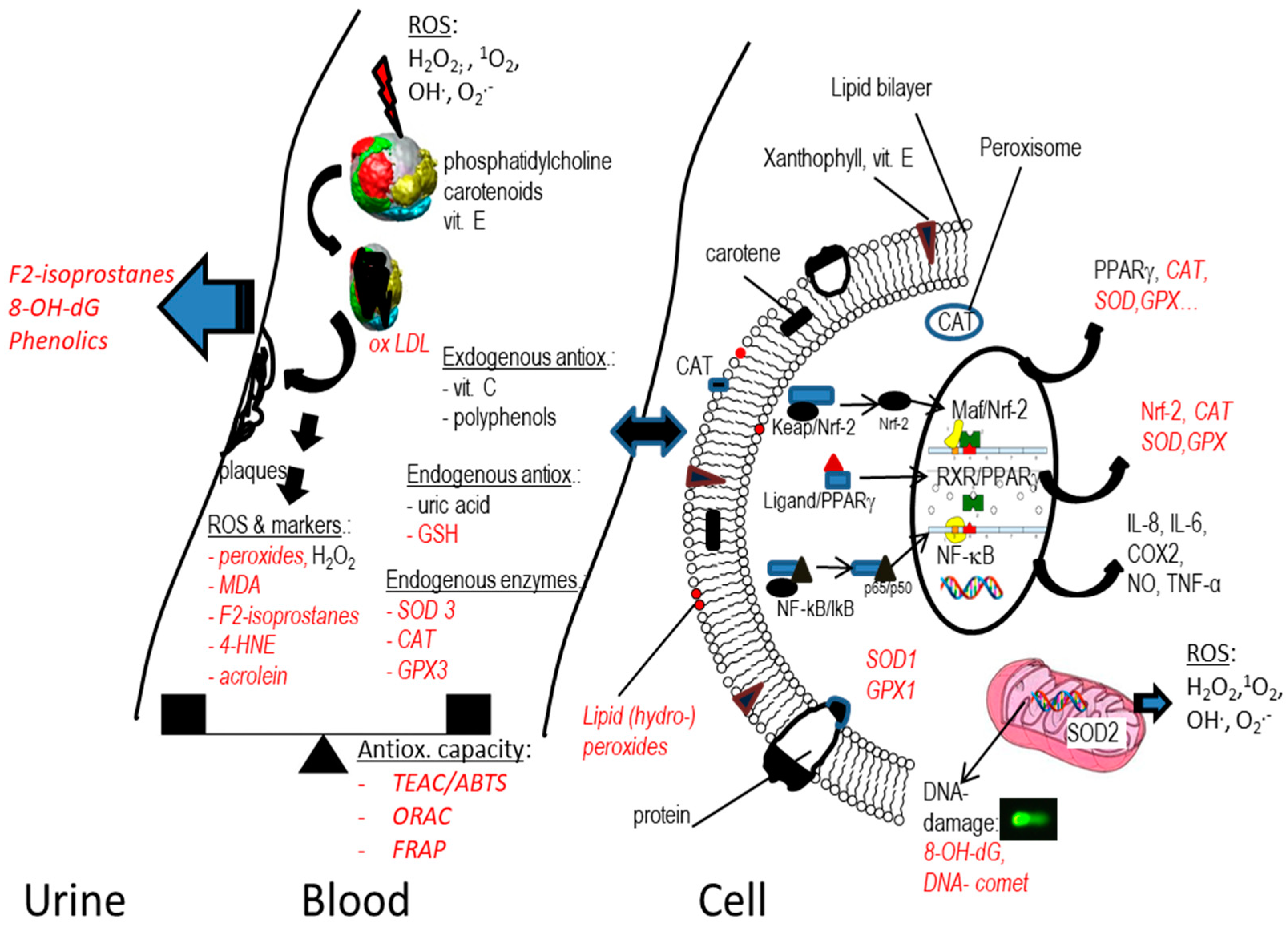
:1. Introduction

{kind=link}
| Name | Chemical Formula | Found in | Typical Conc. in Food (µg/100 g) | Dietary Intake £ (mg/d) | Conc. in Plasma/Serum £ (µM) |
|---|---|---|---|---|---|
| Beta-carotene | C40H56 | carrots | 8285 ± 1082 [29] | 4.1 ± 1.7 | 0.50 ± 0.14 |
| leafy vegetables (spinach) | 5626 ± 766 [29] | ||||
| sweet potatoes | 5219 [29] | ||||
| pumpkin | 3100 [29] | ||||
| broccoli | 361 ± 7 [29] | ||||
| Alpha-carotene | C40H56 | carrots | 3477 ± 531 [29] | 0.7 ± 0.5 | 0.10 ± 0.04 |
| leafy vegetables (spinach) | |||||
| sweet potatoes | |||||
| pumpkin | 4016 [29] | ||||
| broccoli | 25 ± 3 [29] | ||||
| Zeta-carotene | C40H60 | corn | No data | No data | 0.12–0.14 [30] |
| yellow tomatoes | |||||
| Lutein | C40H56O2 | leafy vegetables (spinach) | 12,198 ± 1930 * [29] | 2.2 ± 0.8 * | 0.33 ± 0.10 * |
| eggs | 835 * [29] $ | ||||
| broccoli | 1403 ± 40 * [29] | ||||
| Zeaxanthin | C40H56O2 | corn | 1355 * [29] | see lutein | see lutein |
| eggs | 51 ± 6 [31] | ||||
| leafy vegetables (spinach) | 445 ± 21 [31] | ||||
| Beta-cryptoxanthin | C40H56O | citrus fruits (oranges) | 1275 ± 73 [31] | 0.3 ± 0.4 | 0.23 ± 0.09 |
| loquat | Ca. 25–510 [32] | ||||
| papaya | 589 ± 160 [29] | ||||
| Violaxanthin | C40H56O4 | leafy vegetables (spinach) | 2765 ± 242 [31] | 1.2 [31] | not detectable |
| bell peppers | 457 ± 9 [31] | ||||
| Neoxanthin | C40H56O4 | leafy vegetables (spinach) | 445 ± 21 [31] | 0.5 [31] | not detectable |
| bell peppers | 361 ± 19 [31] | ||||
| Astaxanthin | C40H52O4 | Salmon | 2.6–3.8% $ [33] | No general data | Normally not detectable, but 0.2 after suppl. with 6 mg every other d for 10 d [34] |
| green algae (Haematococcus pluvialis) | 3% $ [33] | ||||
| Fucoxanthin | C42H58O6 | brown algae | 200–2000 [35] | No general data | not detectable |
| Cantaxanthin | C40H52O2 | crabs | No data | no data | |
| salmon | 120 [36] | ||||
| Strassburger sausages | 1500 [37] | 6.1 µg/kg [38] (~0.4 mg/d) | |||
| Lycopene | C50H56 | tomatoes | 2573 ± 54 [29] | 4.6 ± 2.4 | 0.59 ± 0.27 |
| tomato products (ketchup) | 12062 ± 445 [29] | ||||
| watermelon | 4532 ± 386 [29] | ||||
| pink grapefruit | 1419 ± 525 [29] | ||||
| Phytoene | C40H64 | tomatoes | 1388 ± 156 [31] | 2.0 [31] | 0.10–0.11 [30] |
| tomato products (ketchup) | 3494 ± 279 [31] | ||||
| watermelon | 1172 ± 77 [31] | ||||
| pink grapefruit | 617 ± 60 [31] | ||||
| Phytofluene | C40H62 | tomatoes, | 401 ± 54 [31] | 0.7 [31] | 0.27–0.30 [30] |
| tomato products (ketchup) | 1034 ± 97 [31] | ||||
| watermelon | 443 ± 26 [31] | ||||
| pink grapefruit | 208 ± 34 [31] |
2. Markers of Oxidative Stress Relevant for Carotenoids
2.1. Introduction
2.2. Markers Related to Lipid Peroxidation and Free Radicals
2.3. Markers Related to Antioxidant Capacity/Status
2.4. Markers Related to Nuclear Factors and Inflammation
| Marker | Matrix | Marker of | Disadvantage | Advantages | Ref. |
|---|---|---|---|---|---|
| Lipid Peroxidation Related | |||||
| Lipid peroxides (LOOH) | Plasma | Oxidative stress, measured, e.g., as phosphatidylcholine hydroperoxides by chemiluminescence-based LC, other fluorescence probes, or linoleates by GC-MS | Presumably unstable in matrix, to be measured in fresh samples, though not much data at present, analytically challenging | Accepted by EFSA [72] | [97] |
| Malon-dialdehyde (MDA) | Urine, plasma * | Enzymatic (COX-2) and non-enzymatic lipid peroxidation of PUFAs | Limited stability during storage, a few weeks at −20 °C | Relatively high concentration, 0.1–3 µM | [48] |
| F2-isoprostanes | Urine, plasma | Non-enzymatic lipid peroxidation of PUFAs | Limited storage stability, esp. thaw/freeze cycles, 6 months at −80 °C [57], further formation ex vivo, low concentration | Accepted marker for oxidative stress, accepted by EFSA | [56] |
| 4-hydroxy-nonenal (4-HNE) | Urine, plasma | Enzymatic (COX-2) and non-enzymatic lipid peroxidation of omega-6 fatty acids | Limited storage stability (20% losses at –80 °C over 22 months [51], low conc. (70–110 nM), reactive in vivo | Reactive compound plausibly related to further oxidative stress in vivo | [98] |
| Acrolein | Urine, plasma | Lipid peroxidation | Limited stability, most reactive of the lipid peroxide markers | Toxic and relevant product of lipid peroxidation | [99] |
| Protein carbonyls | Plasma | Oxidation of proteins | Several measurement techniques | Related to several diseases, relatively stable products, several months at −80 °C [100] | [101,102] |
| 8-hydroxy-deoxyguanosine (8-OH-dG) | Urine, plasma | Oxidation of DNA (reactive N and O species), especially 1O2, also marker of cancerogenic risk | Enzymatic kits with cross-reactivity to various DNA/RNA breakdown products | Quite stable ex vivo, 6 y at −20 °C for urine [65], same for plasma (2 y) for −80 °C [103] | [63] |
| Oxidized LDL (ox-LDL) | plasma | Marker of oxidative stress, atherosclerosis | Various methods, e.g., AB against oxidized phosphatidylcholine, different units, low ng/mg range, stability may be limited to some months at −80 °C, but lack of data | Accepted marker for atherosclerosis, accepted by EFSA | [69] |
| Copper-induced oxidation of LDL particles | Plasma | Lipid peroxides present in LDL particles versus present antioxidant (e.g., vitamin E), marker of atherosclerosis risk | Ox. kinetics and formation of conjugated dienes depend on Cu conc. employed, questionable physiol. relevance | Possible relevant marker of carotenoid presence in lipoproteins | [104,105] |
| Antioxidant Capacity | |||||
| Ferric-reducing antioxidant power assay (FRAP) | Plasma | Antioxidant capacity, based on electron transfer | Limited to aqueous systems | Ease of use | [77] |
| Oxygen radical absorbance capacity (ORAC) | Plasma | Antioxidant capacity, hydrogen transfer | Limited to peroxyl radicals, temperature dependence | Integral over time is measured, less susceptible to altered kinetics | [76] |
| ABTS | Plasma | Antioxidant capacity, based on hydrogen and electron transfer | Several antioxidants may not react with ABTS | Ease of use, stability of ABTS | [78] |
| DPPH | Plasma | Antioxidant capacity, based on hydrogen and electron transfer | Several antioxidants may not react with DPPH | Ease of use, stability of DPPH | [79,106] |
| Antioxidant Enzymes and Endogenous Antioxidant | |||||
| Superoxide dismutase (SOD) | Plasma, tissues | Responsible for removal of OH−. via H2O2 dismutation, defence against ROS, low conc. associated with disbalanced ROS | Not always clear dose–response relationship | Low levels linked to several chronic diseases | [86,107] |
| Glutathione peroxidase (GPx) | Plasma, tissues | Removal of peroxides especially in cytosol | Also related to ageing | Reduced conc. related to various diseases and increased ROS | [108,109] |
| Reduced glutathione (GSH) | Tissues, plasma | Most abundant cytosolic non-protein thiol, ROS scavenger | Limited predictability alone, pH dependence | Relatively high concentrations (mM) | [108] |
| Catalase (CAT) | Tissues, blood cells | H2O2 disproportionation, marker of ROS, removal of peroxinitrate and NO, low conc. associated with disbalanced ROS | Limited susceptibility to dietary interventions | Low levels linked to several chronic diseases | [110,111,112] |
| Inflammation: Pro-Inflammatory Cytokines including Chemokines and Acute Phase Proteins | |||||
| TNF-α | Plasma | Activation of immune cells, related to fever, apoptosis | Unclear cut-offs for health determinant | Stable for at least 3 y (−80 °C) [113], established marker of chronic disease | [114] |
| IF-γ | Plasma | Produced by killer cells, part of innate immune response, macrophage activation | Rather related to immune system activation | Related to autoimmune diseases, stable for 2 y at −80 °C [113] | [115] |
| IL-1 | Plasma | Related to fever, bone marrow cell differentiation | Many molecular aspects of activation not understood | Stable for at least 2 y (−80 °C) [113] | [116] |
| IL-6 | Plasma | Marker related to fever and acute phase response | Can be both pro- and anti-inflammatory, interpretation more difficult | Good storage stability, some years at −80 °C [117], 2 y [113] | [118] |
| IL-8 | Plasma | Chemotaxis, attraction of neutrophils, phagocytosis | Strong link also to cancer, rather non-specific due to the relation to many diseases | Likely stable for several y at −80 °C [119], widely used marker associated with chronic diseases | [120] |
| C-reactive protein (CRP) | Plasma | Acute phase protein, regulator of inflammation, complement activation | Various isoforms existing | High conc., good stability, 11 y at −80 °C [121] | [122] |
| Serum amyoloid A | Plasma | Acute phase protein | Limited interpretability and comparability, few data | More novel marker | [123] |
| Transcription Factors including Nuclear Receptors | |||||
| NF-κB | Cells | Activator of pro-inflammatory processes | Requires qPCR or WB, analytically not so easy | Accepted marker of inflammation | [18] |
| Nrf-2 | Cells | Activator of anti-oxidant enzymes | Requires qPCR or WB, analytically not easy | Marker clearly related to antioxidant enzyme activity | [18] |
| RAR/RXR | Cells | Related to vitamin A metabolism and apo-carotenoid status | Difficult to interpret expression directly | Accepted marker related to vitamin A activity | [91] |
| PPAR/RXR | Cells | Marker of lipid metabolism and adipocyte differentiation | Difficult to interpret expression directly | Accepted marker related to adipocyte/ lipid metabolism | [92] |
2.5. Conclusions
3. Results from Human Observational Studies
3.1. Studies with Non-Healthy Subjects
3.2. Studies with Rather Healthy Subjects
3.3. Studies with Hard Endpoints
| Outcome Measured | Study Design | Participants | Findings | Comment | Ref. |
|---|---|---|---|---|---|
| Hard Endpoints (Mortality and Morbidity) | |||||
| Type 2 diabetes (T2D) incidence | Meta-analysis of nine prospective cohort studies, dietary intake and plasma conc. of carotenoids | N = 140,000, mean follow up 13 y | Sign. inverse assoc. with total plasma carotenoid and T2D, RR = 0.761 (0.585–0.990) | Antioxidants as index of oxidative damage/ox. capacity | [161] |
| Metabolic syndrome (MetS) | Meta-analysis of 11 cross-sectional studies of total plasma carotenoids and MetS | N = 45,000 subjects | inverse assoc. between total carotenoids & MetS (OR 0.66; 95%CI, 0.56–0.78). Sig. assoc. for beta-CAR, alpha-CAR and beta-CRY and LYC | Anti-atherosclerotic properties of carotenoids emphasized | [20] |
| All-cause mortality | Meta-analysis of prospective cohort studies and carotene plasma levels | N = >4000 elderly men and women Follow-up: 10 y | Higher carotene plasma levels sign. associated with lower mortality, by 38%: RR = 0.72 (0.59; 0.87) | Relation to inflammation and CRP discussed as potential cause | [19] |
| All-cause mortality | Meta-analysis of prospective cohort studies and beta-CAR dietary intake and plasma conc. | N = 150,000 (intake); N = 25,000 (plasma) participants, 2–26 y follow up | Circulating: Highest vs. lowest group had lower risk of total mortality (RR = 0.69, 95%CI: 0.59–0.80). Intake: RR = 0.83 (0.78–0.88). | Causes discussed include effects on immune system, antioxidant function andvitamin A | [163] |
| All-cause mortality | Prospective cohort study of serum beta-CAR and overall and cause-specific mortality | N = 29,000, follow up 31 y | Men with higher serum beta-CAR had sig. lower all-cause mortality (HR = 0.81, 0.71, 0.69, and 0.64 for quintile 2 (Q2)–Q5 versus Q1, resp. | Antiox. activity, arterial wall protection, vasomotor function, platelet aggregation and thrombosis mentioned | [164] |
| Markers Related to ROS, Non-Enzymatic Antioxidant Activity and Lipid Peroxidation | |||||
| Lipid peroxides | Case-control, type 1 diabetes (T1D) | n= 54 T1D, and n = 40 healthy controls | T1D subjects had much higher lipid peroxides (lipid normalized) and MDA vs. healthy controls, while beta-CAR was much lower (<50%). GSH, GPx were also lower, SOD higher | No direct association, hyperglycemia-caused oxidative stress discussed | [128] |
| Lipid hydro-peroxides | Cross sectional study with healthy subjects | N = 36 healthy subjects | Inverse assoc. of hydroperoxides with some carotenoids: alpha-CAR, beta-CRY, ZEA, not beta-CAR, LUT, LYC | Similar effects for tocopherols | [129] |
| Lipid peroxides (LOOH) | Cross-sectional study on depression, LOOH measured by hemoglobin-methylene blue method | N = 75 healthy females | LOOH conc. sign. positively corr. With depression scores, LOOH sign. inversely related to beta-CAR plasma conc. (r = −0.26) | Other carotenoids ns. neg. associated with LOOH Increased polymorpho-nuclear leukocytes in depression? | [157] |
| Conjugated dienes, lipid peroxides | Cross-sectional, comparison across two countries | n = 22 young adults from Naples vs. n =26 from Bristol | beta-CAR 4.74 vs. 2.85 µM, associated with sign. lower conjugated dienes & lipid peroxides: 29.0 vs. 41.5 and 1.24 vs. 4.58 µM resp. | Prone to many confounders such as physical activity, similar trend for vitamin E | [154] |
| MDA | Cross-sectional, relation of ox. stress and cognitive function | 1389 healthy elderly, 59–71 y | Low level of total carotenoids (<1.86 µM) associated with poor cognitive performance, not MDA | No effect of other antioxidant parameters (SOD, GPx) | [158] |
| MDA | Cross-sectional, correlation between various plasma carotenoids and MDA in urine, plasma and eryhtrocytes | Approximately N = 1200 healthy and self-reported Korean subjects suffering from ox. stress | No sign. assoc. with plasma and urine MDA (except inverse corr. with ZEA, sign. positive assoc. of erythrocyte MDA with all carotenoids (β = 0.247) | Higher requirement of erythrocytes for endogenous antioxidants? | [148] |
| MDA | Cross-sectional, correlation between LYC and MDA in plasma | Approximately N =1560 apparently healthy subjects | Positive sig. association of plasma LYC & MDA (r = 0.159) | LYC rather diet rich in tomato products (ketchup, pasta), thus unhealthy eating patterns | [153] |
| MDA | Case control, painters exposed to organic solvents, correlation between plasma MDA & LYC and beta-CAR | n = 42 subjects and 28 controls | Sign. correlation between MDA & LYC (r = −0.26; & MDA & beta-CAR (r = −0.27) | Also, lower GSH, higher SOD and CAT in exposed group though corr. with carotenoids not specified | [141] |
| MDA | Case control study, with multiple sclerosis (MS) subjects | n = 24 subjects, 24 gender and age matched controls | inverse correlation between serum levels of beta-CAR and MDA (r = −0.83) in patients | Similar correlation between ascorbic acid and MDA | [165] |
| MDA | Case-control hemodialysis subjects versus healthy controls | n = 29 subjects, n = 20 healthy controls | Lycopene levels correlated with MDA (r = −0.50 | LYC ns. in a more complex regression model adjusted for age, gender, CAT, SOD, GSH, GPx | [140] |
| MDA | Case-control, type I diabetic subjects (T1D) | n = 20 children vs. 22 obese children vs. 16 healthy controls | T1D & obese children had higher MDA conc. than controls and beta-CAR was lower in T1D subjects, but not in obese where levels were higher than controls | Also, higher lipoperoxides in obese and T1D children, obese children had higher GPx, no corr. analysis done | [166] |
| MDA | Case-control study, smoking women | n = 20 smoking and 20 non-smoker pregnant women | Lower beta-CAR plasma levels in smokers (ns.), but MDA not different | High variability, low number of subjects | [149] |
| MDA | Cross-sectional, Thai healthy elderly | N = 207 healthy subjects aged 60–91 y | Sign. inverse correlation between MDA and lycopene (r = −0.88) | No sign. corr. with α-tocopherol. Smokers & male had higher MDA conc. than non-smokers and females | [150] |
| MDA | Case-control, subjects with acute tonsillitis (AT) vs. acute otitis media (AOM) | n = 23 children with AOM, 27 with AT and 29 healthy control | beta-CAR & GSH sign. decreased, in both patient groups, MDA was higher | No direct corr. analysis carried out | [145] |
| MDA, protein carbonyls, sialic acid | Case-control study, acute coronary syndromes (ACS) | n = 102 patients with ACS & 45 controls | MDA and protein carbonyls sign. increased, total carotenoids decreased going from unstable angina pectoris to MI | Sign. corr. of carotenoids to MDA, protein carbonyls and sialic acid: r = −0.51, −0.32, −0.22, resp. | [133] |
| F2-isoprostanes (8,12-isoprostane F(2alpha)-VI), SOD | Case-control, subjects with congestive heart failure (CHF) | n = 30 patients and n= 30 controls | F2-isoprostanes higher in class III than in class II NYHA patients, inverse corr. between F2-isoprostanes and plasma LUT (r = −0.68), LYC (r= −0.61), ZEA (r = −0.59), alpha-CAR (r = –0.44),beta-CAR (r = –0.41), and SOD (r = −0.42) in patients | Similar corr. with vitamin C, E, A | [167] |
| F2-isoprostanes (8-iso-PGF2α) | Cross-sectional, correlation between F2-isoprostanes in urine and plasma and tissue carotenoids | N = 69 patients at risk of liver cancer | Neg. sign. correlation of plasma beta-CAR and F2-isoprostanes | Also neg. correlations with LYC and LUT, but ns., same for tissue carotenoids | [135] |
| F2-isoprostanes | Longitudinal study, FFQ and plasma carotenoids as well as free plasma F2-isoprostanes | Healthy premenopausal women studied over 2 cycles (N = 258) | Higher beta-CAR, beta-CRY and LUT due to higher dietary intakes also had lower plasma F2-isoprostane conc., LYC intake slightly pos. associated with F2-isoprostanes | Ketchup as major dietary source of LYC; though no direct associations between F2-isoprostanes & carotenoids given | [152] |
| F2-isoprostanes, MDA | Case-control study with atherosclerotic patients | n = 30 patients and 62 healthy controls | Independent of fruit & vegetable intake, sign. lower plasma levels of all carotenoids except beta-CRY vs, controls. Plasma F2- isoprostane doubled; MDA increased one-third | Similar fruit and vegetable intake between groups | [127] |
| F2-isoprostanes (15-isoprostane F2t) | Case-control study of lung cancer and urinary 15-isoprostane F2t | n = 207 subjects and 414 controls | Lower levels of carotenoids, higher levels of F2-isoprostanes pos. assoc. with higher lung cancer risk | No direct corr. shown but assumed | [136] |
| oxLDL, lipid peroxides | Cross-sectional study with healthy women using oral contraceptives (OCs) vs. non- contraception users (NCU) and intrauterine (hormonal and copper) device users (IUD) | N = 897 healthy volunteers | Sign. increase in lipid peroxides in OCU compared to NCU and IUD users and lower beta-CAR conc. (39% lower), no effect on oxLDL | Adjusted for smoking, systolic BP and BMI; estrogen intake associated with sign. altered ROS, likely due to increased Cu conc. in plasma | [155] |
| 8-OH-dG, MDA | Case-control with lung-cancer subjects | n = 39 patients with lung cancer and 31 healthy controls | Trend for higher beta-CAR correlated with lower 8-OH-dG in patients (p = 0.065), no trend for MDA | MDA pos. cor. with 8-OH-dG, but lower variability, thus ns. correlated with beta-CAR | [137] |
| Markers Related to Enzymatic Antioxidant Defence | |||||
| GPx, SOD, GST, GSH, FRAP | Cross-sectional, being vegetarian | n = 31 vegetarians (including six vegans) and 58 omnivores, non-smokers | Vegetarians ~15% higher levels of plasma carotenoids vs. omnivores, incl. LUT, alpha-CRY, LYC, alpha-CAR, beta-CAR (latter 3 ns). Levels of all antiox. markers similar between groups | No sign. association between carotene and antiox. makers in multilinear regression models | [147] |
| SOD and GPx | Cross-sectional study on sickle cell patients | n = 26 Nigerian and 30 British subjects and healthy controls (30, 15) | Higher beta-CAR in controls vs. subjects, together with higher plasma SOD and GPx, approximately 30% increase for all | Small-scale study, high variability between subjects | [142] |
| SOD, GPx and xanthine oxidase | Case-control study in newly diagnosed non-small cell lung cancer subjects | Stage III (IIIA + IIIB, n = 27) and stage IV (n = 23) and 16 healthy controls | GPx, SOD, CAT sign. reduced, XO activity sign. elevated in NSCLC patients, beta-CAR sign. increased in advanced stage compared to healthy controls (!) | Unclear why advanced NSCLC subjects had higher beta-CAR levels (supplements?), no corr. given | [138] |
| CAT, GPx, GSH, MDA | Case-control, patients with Pemphigus Vulgaris (PV) | n = 18 non-smoking PV and equal number of age- and gender-matched, healthy control subjects | Sign. lower conc. of plasma antiox. vitamins (E, A, and beta-CAR), lower antiox. enzymes (CAT in RBC and plasma, GSH-Px in RBC, and resp. GSH activities in both RBC and plasma, increased MDA (RBC, plasma) | Dietary intake not determined | [168] |
| SOD, CAT, GPx, GSH, conjugated dienes | Case-control, patients with cardiogenic shock that complicate acute myocardial infarction (AMI) | n = 25 patients with AMI vs. n = 25 healthy controls | Patients had higher MDA, conjugated dienes and reduced activities/conc. of erythrocyte antiox. enzymes: SOD, CAT, GPx, erythrocyte and in plasma GSH, and beta-CAR | No corr. analysis between carotenoids & markers of ox. stress | [131] |
| Markers Related to Inflammation | |||||
| CRP, MDA | Cross sectional, various complications, i.e., prostate hyperplasia and prostate cancer (PC) | n = 14 healthy controls, n = 20 patients with benign prostate hyperplasia, n = 40 with local, n = 38 with metastatic (PC) | In PC patients: CRP not corr. with antioxidants or MDA. Neg. corr. Between MDA and LUT (r = −0.263) and LYC (r = −0.269). Lower conc. of carotenoids in patients vs. controls | No correlation with alpha- and beta-carotene | [139] |
| CRP | Case-control study for stroke | n = 68 patients with acute ischemic stroke vs. 41 normal controls | Plasma LYC, alpha- and beta-CAR conc. lower and conc. of inflamm. markers higher in patients with acute ischemic stroke vs. normal controls. alpha- and beta-CAR and LYC in patients with stroke neg. associated with CRP (R = −0.29, −0.41, −0.28 resp.) | na | [134] |
| CRP, TNF-α, MDA | Case-control, Crohn’s disease (CD) | n = 16 active CD patients, 27 clinically stable patients, and 15 healthy controls | beta-CAR levels in sera from all CD patients lower than controls (down to One-third in active CD compared to controls). Patients with active CD had higher GPx (30%), TNF-α (2×), MDA (2×) and CRP (20×) than controls | Conc. unclear, spectrophot. not specific for β-CAR, rather sum of carotenoids, no corr. given, decreased ratio of antioxidants: ROS rather result of acute inflammation, not due to lower intake | [144] |
3.4. Conclusions
4. Results from Human Intervention Trials
4.1. Studies with Non-Healthy Subjects
4.2. Studies with Healthy Subjects
4.3. Studies with Hard Endpoints
4.4. Conclusions
| Marker Measured | Study Design | Subjects | Carotenoid Intake | Findings | Comment | Ref. |
|---|---|---|---|---|---|---|
| Supplements | ||||||
| Lipid peroxides | Randomized double-blind controlled trial | n= 25 smokers and 38 non-smokers | 20 mg beta-CAR or placebo daily for 4 weeks | BPO decreased sign. only in smokers receiving beta-CAR | No effect in healthy non-smokers | [174] |
| Total hydro-peroxides (TH), advanced oxidation protein products (AOPP), FRAP | Randomized controlled, double-blinded clinical trial | N = 150 new-borns, 47 controls, n = 103 supple-mented | LUT or placebo at 6 h and 36 h after birth, each 0.28 mg | No sign. change of TH and AOPP, enhanced FRAP in lutein group versus no change in control group | Slight reduction of TH | [191] |
| Lipid peroxides | randomised, double-blind, placebo-controlled trial | N = 30 middle-aged and senior subjects | 12-week ASX supple-mentation (6 or 12 mg/d) | Erythrocyte peroxide conc. lower in ASX groups than in placebo group (up to 50%), in plasma, lower peroxide levels | Plasma: higher dose more strongly reduced peroxides, erythrocytes: comparable reduction | [195] |
| Phosphatidyl hydro-peroxide (PCOOH) | Randomized, double-blind, placebo-controlled cross-over trial | N = 24 healthy subjects | ASX supplement 6 mg/d (and 10 mg sesamin) for 4 weeks vs. placebo | Reduced PCOOH during ASX suppl. vs. control during mental tasks | Faster recovery from mental fatigue also, carotenoid not given alone | [34] |
| Lipid peroxides in serum | Carotenoid free diet for 2 weeks, followed by beta-CAR administration | N = 15 healthy male subjects | After 2 weeks of intervention: either 7 or 120 (n = 8) mg beta-CAR/d for 4 weeks with same carotenoid- free liquid diet | After repletion with beta-CAR, lipid peroxide levels sign. decreased in both groups, r = −0.60, r = −0.58 | Beta-CAR did not lower serum lipid peroxides, no changes in neutrophil superoxide production | [187] |
| Total hydroperoxides (TH), FRAP * | Randomized, double-blind, placebo-controlled, single-centre study | N = 20 term new-borns (10 controls, 10 supplemented) | LUT or placebo at 12 and 36 h after birth each 0.28 mg | No increase of TH in LUT group but sign. increase in control group; sign. FRAP increase in group receiving LUT vs. no change in controls | Large variabilities, partly non-normal distribution, small clinical changes | [192] |
| MDA | Effect of surgical weight loss on changes in beta-CAR and plasma MDA | N = 22 morbidly obese patients | Not determined | MDA sign. decreased (50%) after operation, no sign. change in beta-CAR | No dietary intake studied | [130] |
| MDA, total antioxidant capacity (ABTS) | 6 months clinical single centre trial | N = 24 patients with cystic fibrosis | 1 mg/kg/d (max. 50 mg/day) for 3 months vs. placebo | Initially raised plasma levels of MDA fell to normal, ABTS showed ns. increase with high dose | Further suppl. with 10 mg/d beta-CAR did not maintain low MDA levels | [172] |
| Cu-mediated LDL stability, SOD, GPx, GSH | Placebo-controlled single-blinded study | N = 175 healthy male volunteers | Daily suppl. of beta-CAR (15 mg), LUT (15 mg), LYC (15 mg) and placebo for 3 months | No sign. effect on any antiox. parameter | Clearly no effect on this healthy population | [189] |
| MDA (plasma), Cu-induced ex-vivo oxidation of LDL | Intervention with beta-CAR in subjects with cystic fibrosis (CF) | n= 34 CF patients, before/after 3 months of beta-CAR suppl., and 42 healthy controls | 0.5 mg/kg body weight beta-CAR | Sig. increase of lag-time after suppl., sign. reduced MDA, sig. increase of beta-CAR, regression of −0.40 µM MDA/µM beta-CAR | Beta-carotene deficiency can be reduced by beta-carotene supple- mentation | [171] |
| MDA, SOD, F2-isoprostane, total antioxidant capacity (TAC) ** | Intervention trial with healthy smokers with random assignment | Healthy smokers | 3 groups receiving ASX at doses of 5, 20 or 40 mg (n = 13, each) once/d for 3 weeks | Plasma MDA and isoprostane decreased, SOD and TAC increased in all ASX groups vs. baseline | Placebo group missing, strongest results for isprostanes (from 17 to 2 ng/mL), no strong diff. across groups | [175] |
| MDA, Cu-induced LDL stability | Supple-mentation trial | N = 20 patients with long-standing non-insulin-dependent diabetes mellitus | Dunaliella bardawil-derived beta-CAR was suppl. to patients for 3 weeks, 60 mg/d | Reduced MDA (25%), LDL susceptibility increased by 40% with 35% shorter lag time | High dose, rich in 9-cis beta-CAR | [173] |
| MDA, TEAC | Intervention study | N = 42 healthy subjects divided into 4 groups | 5 mg, 10 mg, 20 mg or 40 mg beta-CAR/d for 5 weeks | Uric acid in plasma sign. decreased in all groups, TEAC ns. decreased in all groups, MDA unchanged except for 40 mg beta-CAR group (−18%) | Small-scale study, changes rather small as subjects already healthy, no placebo group | [186] |
| MDA, ABTS, CRP, SOD; CAT, GPx, in plasma | LUT supple-mentation, randomized placebo controlled | N = 117 healthy non-smokers | 10 or 20 mg/d of LUT or placebo for 12 weeks | LUT and ABTS sign. increased in both LUT groups. Sign. MDA reduction with 20 mg LUT, dose-dependent CRP decrease, sign. CRP diff. between 20 mg LUT and placebo, CRP sign. related to change in plasma LUT (r = −0.44) and ABTS for both groups | No sign. change in other parameters measured, perhaps as subjects were healthy and well educated | [181] |
| LDL oxidizability, MDA, hydroxyl-nonenals, urinary F2-isoprostanes and 8-OH-dG and lymphocytes (comet assay) | Dose–response study with LYC on healthy subjects, double-blind, randomized, placebo- controlled | Healthy adults (N = 77, age ≥ 40 years) | LYC-restricted diet for 2 weeks, then randomized to receive 0, 6.5, 15 or 30 mg LYC/d for 8 weeks | Sign. decrease in DNA damage by comet assay and a sign. decrease in 8-OH-dG at 8 weeks vs. baseline with 30 mg LYC/d | Limited effects as subjects were healthy, less than 10% changes via comet assay, approximately 25% reduction of 8-OH-dG | [196] |
| FRAP, F2-isoprostane, oxidised phospholipid biomarker in serum (POVPC) | Intervention with supplement in Alzheimer’s disease (AD) subjects vs. healthy controls | n = 21 AD patients vs. 16 healthy age-matched controls | 10 mg meso-ZEA, 10 mg LUT, 2 mg ZEA for 6 months | IsoP conc. not related to disease or suppl. FRAP sign. lower and POVPC sign. higher in AD than healthy controls, but not changed by carotenoids | na | [178] |
| CRP | Randomized controlled multi-centre trial | N = 183 infants < 33 weeks gestational age | 210 µg/L in formula, about equal parts beta-CAR, LUT, LYC, up to until 40 weeks post-menstrual age | Suppl. infants had lower plasma CRP, 0.2 vs. 0.4 µg/mL | Infants on human milk had similar CRP values than those in LUT group | [193] |
| MDA, SOD, GPx, GR, GSH, CAT | LYC suppl. 10 weeks in subjects with ox. stress after a 2 week washout period | 45 patients age 40–60 y and 30 age and sex-matched healthy controls (not suppl., just baseline!) | Group 1 (n = 15) suppl. with 15 mg LYC capsule, group 2 (n = 15) 200 g tomato products containing 15 mg LYC, group 3 (n = 15) placebo | Patients: decreased lipid peroxidation and enhanced OS (all other markers) after various forms of LYC suppl. | Incomplete statistics, hard to evaluate results, both supplements and LYC rich foods appear equally effective, slight edge for supplements | [181] |
| plasma IL-6, monocyte-chemoattractant protein (MCP-19, serum apoE | Randomized placebo-controlled trial | Early atherosclerosis patients (N = 65) | 20 mg LUT/d, 3 months vs. placebo | Reduced IL-6, but ns. compared to control group, reduced MCP-19, reduced apoE | No other inflammation parameters investigated | [212] |
| Food Items | ||||||
| Phospholipid hydroperoxides (PLOOH) in erythrocytes and plasma | randomized, double-blind placebo-controlled with chlorella algae | n =12 normal senior subjects, n = 6/group | 2 months chlorella Suppl. (8 g chlorella/d; with 22.9 mg LUT/d | Reduced PLOOH in suppl. subject, but also in control group, thus no sign. difference between these | Subjects were healthy, underpowered study | [194] |
| Cu-induced LDL oxidation ex vivo, SOD, GPx, GSH, | Intervention study with fruits and vegetables for 2 weeks | Smokers (n = 11) and age and gender-matched non-smokers (n = 11) | 30 mg/d via diet | Carotenoids increased in smokers 23% and 11% in non-smokers, LDL resistance to ox. increased by 14% in smokers and 28% in non-smokers | Very small-scale study, short-term, no sig. effect on other antiox. markers | [177] |
| DNA strand breaks, ox. DNA damage, plasma MDA and F2-isoprostanes, TEAC | Randomized controlled trial | N= 64 non-smoking healthy male subjects | Either low (2 servings/d), medium (5 servings/d), or high (8 servings/d) intake of VF for another 4 wks. | No changes in observed parameters | Already healthy subjects, little changes | [197] |
| MDA, SOD, GPx, GR, GSH | Intervention study | hypertensive subjects (n = 40) | 60 d of tomato supple-mentation, 200 g ripe tomatoes/d | Sign. reduction in MDA, sign. increases in other parameters | No intake and plasma levels of lycopene determined | [182] |
| MDA, SOD, CAT, GPx, protein thiols | Dietary intervention with diet low in LYC for 1 month | N = 23 healthy post-menopausal women | Reduction from 3.50 mg/d to 0.13 mg/d | Sig. decreased serum LYC (1170 to 495 nM), LUT/ZEA, alpha-& beta-CAR. GPx, lipid and protein ox. increased (ns.), while CAT & SOD sign. decreased | General reduction of fruits and vegetables as major confounder | [200] |
| Amyloid A (apolipoprotein, marker of systemic and HDL–associated inflammation) | Randomized, controlled intervention trial | Middle-aged, overweight adults, n = 54 | LYC-rich diet (224–350 mg) or supplements (70 mg/wk), for 12 wks | Reduced (30%) serum-amyloid A for LYC supplement group only | No change in lycopene rich diet without supplements | [213] |
| MDA (erythrocytes), F2-isoprostanes (urine) | Intervention study with red palm oil (RPO) or vitamin E | N = 60 patients (mean age 62 y, range 54–75) with child A/B, genotype 1 hepatitis C virus-related cirrhosis | Not determined, 15 g RPO for 8 weeks | Both treatments sign. decreased erythrocyte MDA and isoprostane output, carotenoid treatment stronger, by ~30% | RPO also sign. affected macrophage-colony stimulating factor and monocyte tissue factor | [180] |
| MDA | Single-blind, placebo-controlled trial | N = 31 subjects with grade-1 hyper-tension | 8-week treatment period with tomato extract, 250 mg (15 mg LYC/d) | MDA decreased from 4.58 to 3.81 nmol/mg | Marginal changes in MDA, but 10 mm Hg reduction in systolic BP | [199] |
| MDA, GSH | Randomized controlled trial | N = 41 healthy post-meno-pausal women, 6 months | Either hormone replacement therapy (HRT, n = 21) or LycoRed (n = 20) containing 4 mg LYC/d | MDA sign. decreased by 16.3% and 13.3%, GSH increased sign. by 5.9% and 12.5% in HRT and LycoRed groups, resp. | Limited effects, likely due to the generally healthy subjects | [198] |
| MDA, 8-OH-dG | Double-blind, randomized, placebo-controlled trial | 105 African men, veterans, recommended for prostate biopsy | Tomato oleorosin with 30 mg/d LYC vs. placebo | No sign. changes in MDA in plasma. Tissue 8-OH-dG lower but ns. vs. controls | Too large variability in tissue 8-OH-dG | [185] |
| Lymphocyte 8-OH-DhG, urinary urinary F2-isopostanes | Dietary intervention with diets rich in botanicals, 8–10 servings of VF/d in low (LB) or high (HB) botanical variety study, for 14 d | n = 2 × 53 healthy women | µg/d: alpha-CAR: 108, 7200; beta-CAR: 7200, 9100, LUT: 12100, 5800; LYC: 8900; 7300. beta-CRY: 900, 200 | Sign. decrease for both diets for 8-OH-dG, −0.03 and −0.81; and 8-iso-PGF2a: −0.05 µmol, −0.13 | LB slightly less effective than HB, both diets of same macronutrient comp., more diverse phyto- chemicals in smaller amounts superior? | [205] |
| 8-OH-dG, lipid peroxides | double-blinded, randomized, placebo-controlled study, 28 d | Healthy Japanese adults, N = 60 | Capsules with juice powder conc. from apple, orange, pineapple, papaya, cranberry, acerola cherry, peach, carrot, parsley, beetroot, broccoli, cabbage, spinach, tomato, kale, barley & oat bran, with ca. 234 mg vit. C, 32 mg vit. E, 7.5 mg beta- CAR equivalents and 160 mg bioflavonoids/d. | Measures of ox. stress decreased with serum lipid peroxides declining −10.5% and urine 8-OH-dG decreased −21.1% | Similar improvements in smokers vs. non-smokers | [188] |
| Urinary F2-isoprostanes, 8-OH-dG in urine and lymphocytes | Dietary intervention for 2 weeks, receiving 12 portions fruits/ vegetables per day | N = 37 healthy women at risk for breast cancer | Not determined | Sign. corr. between all markers and plasma carotenoids, sign. corr. between change in plasma xanthophylls and lymphocyte 8-OH-dG, r = −0.45 | No other sign. correlations observed during 2 week study, too short intervention | [204] |
| TNF-α, basal lymphocyte DNA damage (comet assay), F2-isoprostanes (urine) | Placebo-controlled crossover trial | N = 26 healthy subjects | Lyc-o-Mato drink: 5.7 mg Lyc, 3.7 mg PHY, 2.7 mg PHF, 1 mg beta-CAR, 1.8 mg α-TOC for 26 d | TNF-αsecretion decreased 34% compared to control | No effect on other parameters | [214] |
| CRP | Lycopene: Randomized controlled trial | n = 22, men and women in patients with heart failure vs. control (n = 18) | 30 mg /d for 30 d within V8 tomato juice | CRP↓ (25%) in women (not men)↓ | Gender-specific effects | [215] |
| IL-2, IL-4, TNF-α of cultured PBMCs | Randomized controlled trial | N = 55 elderly | 330 mL/d tomato juice (47.1 mg/d LYC) for 8 weeks | No effect compared to control group | [216] | |
5. Summary and Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Steiger, S.; Perez-Fons, L.; Fraser, P.D.; Sandmann, G. Biosynthesis of a novel C30 carotenoid in bacillus firmus isolates. J. Appl. MicroBiol. 2012, 113, 888–895. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids database: Structures, chemical fingerprints and distribution among organisms. Database 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, M.S.; Lee, B.Y.; Lee, P.C. Generation of structurally novel short carotenoids and study of their biological activity. Sci. Rep. 2016, 6, 21987. [Google Scholar] [CrossRef]
- Squillaci, G.; Parrella, R.; Carbone, V.; Minasi, P.; La Cara, F.; Morana, A. Carotenoids from the extreme halophilic archaeon Haloterrigena turkmenica: Identification and antioxidant activity. Extremophiles 2017, 21, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.W.; Hsu, S.H.; Lin, M.T.; Hsu, Y.H. Mass production of C50 carotenoids by haloferax mediterranei in using extruded rice bran and starch under optimal conductivity of brined medium. BioProc.ess Biosyst. Eng. 2015, 38, 2361–2367. [Google Scholar] [CrossRef] [PubMed]
- Heider, S.A.E.; Peters-Wendisch, P.; Wendisch, V.F.; Beekwilder, J.; Brautaset, T. Metabolic engineering for the microbial production of carotenoids and related products with a focus on the rare C50 carotenoids. Appl. MicroBiol. Biotechnol. 2014, 98, 4355–4368. [Google Scholar] [CrossRef]
- Hou, J.; Cui, H.L. In vitro antioxidant, antihemolytic, and anticancer activity of the carotenoids from halophilic archaea. Curr. MicroBiol. 2018, 75, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Böhm, V.; Borel, P.; Corte-Real, J. From carotenoid intake to carotenoid/metabolite blood and tissue concentrations—Implications for dietary intake recommendations. Nutr. Rev. 2018. under review. [Google Scholar]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, R.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef]
- Kancheva, V.D.; Kasaikina, O.T. Bio-antioxidants—A chemical base of their antioxidant activity and beneficial effect on human health. Curr. Med. Chem. 2013, 20, 4784–4805. [Google Scholar] [CrossRef]
- Erdman, J.W., Jr.; Ford, N.A.; Lindshield, B.L. Are the health attributes of lycopene related to its antioxidant function? Arch. BioChem. Biophys. 2009, 483, 229–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briviba, K.; Schnabele, K.; Rechkemmer, G.; Bub, A. Supplementation of a diet low in carotenoids with tomato or carrot juice does not affect lipid peroxidation in plasma and feces of healthy men. J. Nutr. 2004, 134, 1081–1083. [Google Scholar] [CrossRef] [PubMed]
- Rühl, R. Effects of dietary retinoids and carotenoids on immune development. Proc. Nutr. Soc. 2007, 66, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonet, M.L.; Canas, J.A.; Ribot, J.; Palou, A. Carotenoids in adipose tissue biology and obesity. Subcell. BioChem. 2016, 79, 377–414. [Google Scholar] [PubMed]
- FAO. Enrichment of dried skim milk with special reference to vitamin A. Food Nutr. 1977, 3, 2–7. [Google Scholar]
- Linnewiel, K.; Ernst, H.; Caris-Veyrat, C.; Ben-Dor, A.; Kampf, A.; Salman, H.; Danilenko, M.; Levy, J.; Sharoni, Y. Structure activity relationship of carotenoid derivatives in activation of the electrophile/antioxidant response element transcription system. Free Radic. Biol. Med. 2009, 47, 659–667. [Google Scholar] [CrossRef]
- Linnewiel-Hermoni, K.; Motro, Y.; Miller, Y.; Levy, J.; Sharoni, Y. Carotenoid derivatives inhibit nuclear factor kappa B activity in bone and cancer cells by targeting key thiol groups. Free Radic. Biol. Med. 2014, 75, 105–120. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Carotenoids, inflammation, and oxidative stress—Implications of cellular signaling pathways and relation to chronic disease prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef]
- Buijsse, B.; Feskens, E.J.; Schlettwein-Gsell, D.; Ferry, M.; Kok, F.J.; Kromhout, D.; de Groot, L.C. Plasma carotene and alpha-tocopherol in relation to 10-y all-cause and cause-specific mortality in European elderly: The survey in Europe on nutrition and the elderly, a concerted action (SENECA). Am. J. Clin. Nutr. 2005, 82, 879–886. [Google Scholar]
- Beydoun, M.A.; Chen, X.; Jha, K.; Beydoun, H.A.; Zonderman, A.B.; Canas, J.A. Carotenoids, vitamin A, and their association with the metabolic syndrome: A systematic review and meta-analysis. Nutr. Rev. 2019, 77, 32–45. [Google Scholar] [CrossRef]
- Donaldson, M.S. A carotenoid health index based on plasma carotenoids and health outcomes. Nutrients 2011, 3, 1003–1022. [Google Scholar] [CrossRef] [PubMed]
- CDC. National Report on Biochemical Indicators of Diet and Nutrition in the U.S. Population 1999–2002; CDC: Atlanta, GA, USA, 2008.
- Albanes, D.; Heinonen, O.P.; Taylor, P.R.; Virtamo, J.; Edwards, B.K.; Rautalahti, M.; Hartman, A.M.; Palmgren, J.; Freedman, L.S.; Haapakoski, J.; et al. Alpha-Tocopherol and beta-carotene supplements and lung cancer incidence in the alpha-tocopherol, beta-carotene cancer prevention study: Effects of base-line characteristics and study compliance. J. Natl. Cancer Inst. 1996, 88, 1560–1570. [Google Scholar] [CrossRef] [PubMed]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L., Jr.; Valanis, B.; Williams, J.H., Jr.; et al. Effects of a combination of beta carotene and vitamin A on lung cancer and cardiovascular disease. N. Engl. J. Med. 1996, 334, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Cullen, M.R.; Barnett, M.J.; Balmes, J.R.; Cartmel, B.; Redlich, C.A.; Brodkin, C.A.; Barnhart, S.; Rosenstock, L.; Goodman, G.E.; Hammar, S.P.; et al. Predictors of lung cancer among asbestos-exposed men in the {beta}-carotene and retinol efficacy trial. Am. J. Epidemiol. 2005, 161, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Albanes, D.; Virtamo, J.; Taylor, P.R.; Rautalahti, M.; Pietinen, P.; Heinonen, O.P. Effects of supplemental beta-carotene, cigarette smoking, and alcohol consumption on serum carotenoids in the alpha-tocopherol, beta-carotene cancer prevention study. Am. J. Clin. Nutr. 1997, 66, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T. Carotenoids, chronic disease prevention and dietary recommendations. Int. J. Vitam. Nutr. Res. 2018, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive oxygen species in metabolic and inflammatory signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- USDA (2015) National Nutrient Database for Standard Reference Release 28. Available online: http://ndb.nal.usda.gov/ndb/foods/show (accessed on 5 November 2015).
- Paetau, I.; Khachik, F.; Brown, E.D.; Beecher, G.R.; Kramer, T.R.; Chittams, J.; Clevidence, B.A. Chronic ingestion of lycopene-rich tomato juice or lycopene supplements significantly increases plasma concentrations of lycopene and related tomato carotenoids in humans. Am. J. Clin. Nutr. 1998, 68, 1187–1195. [Google Scholar] [CrossRef]
- Biehler, E.; Alkerwi, A.; Hoffmann, L.; Krause, E.; Guillaume, M.; Lair, M.L.; Bohn, T. Contribution of violaxanthin, neoxanthin, phytoene and phytofluene to total carotenoid intake: Assessment in luxembourg. J. Food Compos. Anal. 2012, 25, 56–65. [Google Scholar] [CrossRef]
- Zhou, C.H.; Xu, C.J.; Sun, C.D.; Li, X.; Chen, K.S. Carotenoids in white-and red-fleshed loquat fruits. J. Agric. Food Chem. 2007, 55, 7822–7830. [Google Scholar] [CrossRef]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef] [PubMed]
- Imai, A.; Oda, Y.; Ito, N.; Seki, S.; Nakagawa, K.; Miyazawa, T.; Ueda, F. Effects of dietary supplementation of astaxanthin and sesamin on daily fatigue: A randomized, double-blind, placebo-controlled, two-way crossover study. Nutrients 2018, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Timmermans, B. Fucoxanthin: A Promising Bioproduct to Be Derived from Algae? Rijksuniversiteit: Groningen, The Netherlands, 2016. [Google Scholar]
- Brizio, P.; Benedetto, A.; Righetti, M.; Prearo, M.; Gasco, L.; Squadrone, S.; Abete, M.C. Astaxanthin and canthaxanthin (xanthophyll) as supplements in rainbow trout diet: In vivo assessment of residual levels and contributions to human health. J. Agric. Food Chem. 2013, 61, 10954–10959. [Google Scholar] [CrossRef] [PubMed]
- Esatbeyoglu, T.; Rimbach, G. Canthaxanthin: From molecule to function. Mol. Nutr. Food Res. 2017, 61, 1600469. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion on the re-evaluation of canthaxanthin (E 161 g) as a food additive. EFSA J. 2010, 8, 1852. [Google Scholar] [CrossRef]
- Sy, C.; Gleize, B.; Dangles, O.; Landrier, J.F.; Veyrat, C.C.; Borel, P. Effects of physicochemical properties of carotenoids on their bioaccessibility, intestinal cell uptake, and blood and tissue concentrations. Mol. Nutr. Food Res. 2012, 56, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.W.; Ingold, K.U. Beta-Carotene: An unusual type of lipid antioxidant. Science 1984, 224, 569–573. [Google Scholar] [CrossRef]
- Gruszecki, W.I.; Strzalka, K. Carotenoids as modulators of lipid membrane physical properties. Biochim. Biophys. Acta 2005, 1740, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef]
- El-Agamey, A.; Lowe, G.M.; McGarvey, D.J.; Mortensen, A.; Phillip, D.M.; Truscott, T.G.; Young, A.J. Carotenoid radical chemistry and antioxidant/pro-oxidant properties. Arch. BioChem. Biophys. 2004, 430, 37–48. [Google Scholar] [CrossRef]
- Schaich, K.M.; Borg, D.C. Fenton reactions in lipid phases. Lipids 1988, 23, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Conn, P.F.; Lambert, C.; Land, E.J.; Schalch, W.; Truscott, T.G. Carotene-oxygen radical interactions. Free Radic. Res. Commun. 1992, 16, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Chiste, R.C.; Freitas, M.; Mercadante, A.Z.; Fernandes, E. Carotenoids inhibit lipid peroxidation and hemoglobin oxidation, but not the depletion of glutathione induced by ROS in human erythrocytes. Life Sci. 2014, 99, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Ito, F.; Sono, Y.; Ito, T. Measurement and clinical significance of lipid peroxidation as a biomarker of oxidative stress: Oxidative stress in diabetes, atherosclerosis, and chronic inflammation. AntiOxid.ants 2019, 8, 72. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.A.; Gescher, A.; Plastaras, J.P.; Leuratti, C.; Singh, R.; Gallacher-Horley, B.; Offord, E.; Marnett, L.J.; Steward, W.P.; Plummer, S.M. Cyclooxygenase-2, malondialdehyde and pyrimidopurinone adducts of deoxyguanosine in human colon cells. Carcinogenesis 2001, 22, 1557–1560. [Google Scholar] [CrossRef]
- Nam, T.G. Lipid peroxidation and its toxicological implications. Toxicol. Res. 2011, 27, 1–6. [Google Scholar] [CrossRef]
- Spies-Martin, D.; Sommerburg, O.; Langhans, C.D.; Leichsenring, M. Measurement of 4-hydroxynonenal in small volume blood plasma samples: Modification of a gas chromatographic-mass spectrometric method for clinical settings. J. Chromatogr. B 2002, 774, 231–239. [Google Scholar] [CrossRef]
- Il’yasova, D.; Scarbrough, P.; Spasojevic, I. Urinary biomarkers of oxidative status. Clin. Chim. Acta 2012, 413, 1446–1453. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Dhillon, B.S.; Rao, D.N.; Menon, G.; Shankar, H.; Dhaliwal, L.K.; Leema, M.; Chandhiok, N.; Kumar, N.; Sehgal, R.; et al. Temporal trends of malondialdehyde in stored human plasma. Indian J. Clin. Biochem. 2012, 27, 405–409. [Google Scholar] [CrossRef]
- Tsikas, D.; Suchy, M.T.; Niemann, J.; Tossios, P.; Schneider, Y.; Rothmann, S.; Gutzki, F.M.; Frölich, J.C.; Stichtenoth, D.O. Glutathione promotes prostaglandin H synthase (cyclooxygenase)-dependent formation of malondialdehyde and 15(S)-8-iso-prostaglandin F2alpha. FEBS Lett. 2012, 586, 3723–3730. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.S.; Bilodeau, J.F.; Julien, P.; Rudkowska, I. Dietary fats and F2-isoprostanes: A review of the clinical evidence. Crit. Rev. Food Sci. Nutr. 2017, 57, 3929–3941. [Google Scholar] [CrossRef] [PubMed]
- Kaviarasan, S.; Muniandy, S.; Qvist, R.; Ismail, I.S. F(2)-isoprostanes as novel biomarkers for type 2 diabetes: A review. J. Clin. Biochem. Nutr. 2009, 45, 1–8. [Google Scholar] [CrossRef]
- Kitano, S.; Hisatomi, H.; Hibi, N.; Kawano, K.; Harada, S. Improved method of plasma 8-Isoprostane measurement and association analyses with habitual drinking and smoking. World J. Gastroenterol. 2006, 12, 5846–5852. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Junger, W.G. Measurement of oxidative burst in neutrophils. Methods Mol. Biol. 2012, 844, 115–124. [Google Scholar] [PubMed]
- Slauch, J.M. How does the oxidative burst of macrophages kill bacteria? Still an open question. Mol. MicroBiol. 2011, 80, 580–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, D.B.; Boff, J.M. Chemistry and reaction of singlet oxygen in foods. Compr. Rev. Food Sci. Food Saf. 2002, 1, 58–72. [Google Scholar] [CrossRef]
- Ojima, F.; Sakamoto, H.; Ishiguro, Y.; Terao, J. Consumption of carotenoids in photosensitized oxidation of human plasma and plasma low-density lipoprotein. Free Radic. Biol. Med. 1993, 15, 377–384. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2’ -deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health Part C 2009, 27, 120–139. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Gasparutto, D.; Ravanat, J.L. Oxidative damage to DNA: Formation, measurement and biochemical features. Mutat. Res. 2003, 531, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, H.E.; Loft, S.; Prieme, H.; Vistisen, K.; Lykkesfeldt, J.; Nyyssonen, K.; Salonen, J.T. Oxidative DNA damage in vivo: Relationship to age, plasma antioxidants, drug metabolism, glutathione-S-transferase activity and urinary creatinine excretion. Free Radic. Res. 1998, 29, 565–571. [Google Scholar] [CrossRef]
- Goulinet, S.; Chapman, M.J. Plasma LDL and HDL subspecies are heterogenous in particle content of tocopherols and oxygenated and hydrocarbon carotenoids: Relevance to oxidative resistance and atherogenesis. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Romanchik, J.E.; Morel, D.W.; Harrison, E.H. Distributions of carotenoids and alpha-tocopherol among lipoproteins do not change when human plasma is incubated in vitro. J. Nutr. 1995, 125, 2610–2617. [Google Scholar] [PubMed]
- Gao, S.; Liu, J. Association between circulating oxidized low-density lipoprotein and atherosclerotic cardiovascular disease. Chr. Dis. Transl. Med. 2017, 3, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Itabe, H. Oxidized low-density lipoprotein as a biomarker of in vivo oxidative stress: From atherosclerosis to periodontitis. J. Clin. Biochem. Nutr. 2012, 51, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Itabe, H.; Ueda, M. Measurement of plasma oxidized low-density lipoprotein and its clinical implications. J. ATher.oscler. Thromb. 2007, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef]
- EFSA. Guidance on the scientific requirements for health claims related to antioxidants, oxidative damage and cardiovascular health. EFSA J. 2018, 16, 5136. [Google Scholar]
- Yamamoto, Y.; Kambayashi, Y.; Ueda, T. Assay of phospholipid hydroperoxides by chemiluminescence-based high-performance liquid chromatography. Methods Mol. Biol. 1998, 108, 63–70. [Google Scholar]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; Miyashita, K.; Camp, J.V.; Alasalvar, C.; Ismail, A.B. Antioxidant activity, total phenolics and flavonoids contents: Should we ban in vitro screening methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Garrett, A.R.; Murray, B.K.; Robison, R.A.; O’Neill, K.L. Measuring antioxidant capacity using the ORAC and TOSC assays. Methods Mol. Biol. 2010, 594, 251–262. [Google Scholar] [PubMed]
- Gülçin, İ. (2015) Fe3+–Fe2+ Transformation Method: An Important Antioxidant Assay. In Advanced Protocols in Oxidative Stress III; Armstrong, D., Ed.; Springer: New York, NY, USA, 2015; pp. 233–246. [Google Scholar]
- Olszowy, M.; Dawidowicz, A.L. Is it possible to use the DPPH and ABTS methods for reliable estimation of antioxidant power of colored compounds? Chem. Pap. 2018, 72, 393–400. [Google Scholar] [CrossRef]
- Kedare, S.B.; Singh, R.P. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prior, R.L.; Hoang, H.; Gu, L.; Wu, X.; Bacchiocca, M.; Howard, L.; Hampsch-Woodill, M.; Huang, D.; Ou, B.; Jacob, R. Assays for hydrophilic and lipophilic antioxidant capacity (oxygen radical absorbance capacity (ORAC(FL))) of plasma and other biological and food samples. J. Agric. Food Chem. 2003, 51, 3273–3279. [Google Scholar] [CrossRef]
- Cano, A.; Alcaraz, O.; Acosta, M.; Arnao, M.B. On-line antioxidant activity determination: Comparison of hydrophilic and lipophilic antioxidant activity using the ABTS*+ assay. Redox. Rep. 2002, 7, 103–109. [Google Scholar] [CrossRef]
- Abramovic, H.; Grobin, B.; Poklar Ulrih, N.; Cigić, B. Relevance and standardization of in vitro antioxidant assays: ABTS, DPPH, and Folin-Ciocalteu. J. Chem. 2018, 2018, 9. [Google Scholar] [CrossRef]
- Huang, Y.; Li, W.; Su, Z.Y.; Kong, A.N. The complexity of the Nrf2 pathway: Beyond the antioxidant response. J. Nutr. Biochem. 2015, 26, 1401–1413. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Targ. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Margaritelis, N.V.; Veskoukis, A.S.; Paschalis, V.; Vrabas, I.S.; Dipla, K.; Zafeiridis, A.; Kyparos, A.; Nikolaidis, M.G. Blood reflects tissue oxidative stress: A systematic review. Biomarkers 2015, 20, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. AntiOxid. Redox. Signal 2011, 15, 1583–1606. [Google Scholar] [CrossRef]
- McCord, J.M. Superoxide dismutase, lipid peroxidation, and bell-shaped dose response curves. Dose Response 2008, 6, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Sever, R.; Glass, C.K. Signaling by nuclear receptors. Cold Spring Harb. Perspect. Biol. 2013, 5, a016709. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T.; Desmarchelier, C.; El, S.N.; Keijer, J.; van Schothorst, E.; Rühl, R.; Borel, P. beta-Carotene in the human body: Metabolic bioactivation pathways - from digestion to tissue distribution and excretion. Proc. Nutr. Soc. 2019, 78, 68–87. [Google Scholar] [CrossRef]
- Bonet, M.L.; Canas, J.A.; Ribot, J.; Palou, A. Carotenoids and their conversion products in the control of adipocyte function, adiposity and obesity. Arch. Biochem. Biophys. 2015, 572, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Le Maire, A.; Alvarez, S.; Shankaranarayanan, P.; Lera, A.R.; Bourguet, W.; Gronemeyer, H. Retinoid receptors and therapeutic applications of RAR/RXR modulators. Curr. Top Med. Chem. 2012, 12, 505–527. [Google Scholar] [CrossRef]
- Kroker, A.J.; Bruning, J.B. Review of the structural and dynamic mechanisms of PPAR-gamma partial agonism. PPAR Res. 2015, 2015, 816856. [Google Scholar] [CrossRef]
- Aydemir, G.; Kasiri, Y.; Bartok, E.M.; Birta, E.; Fröhlich, K.; Böhm, V.; Mihaly, J.; Rühl, R. Lycopene supplementation restores vitamin A deficiency in mice and possesses thereby partial pro-vitamin A activity transmitted via RAR signaling. Mol. Nutr. Food Res. 2016, 60, 2413–2420. [Google Scholar] [CrossRef] [Green Version]
- Carrier, A. Metabolic syndrome and oxidative stress: A complex relationship. AntiOxid. Redox. Signal 2017, 26, 429–431. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Umeno, A.; Shichiri, M. Lipid peroxidation biomarkers for evaluating oxidative stress and assessing antioxidant capacity in vivo. J. Clin. Biochem. Nutr. 2013, 52, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaur, R.J.; Siems, W.; Bresgen, N.; Eckl, P.M. 4-Hydroxy-nonenal-a bioactive lipid peroxidation product. Biomolecules 2015, 5, 2247–2337. [Google Scholar] [CrossRef]
- Uchida, K.; Kanematsu, M.; Sakai, K.; Matsuda, T.; Hattori, N.; Mizuno, Y.; Suzuki, D.; Miyata, T.; Noguchi, N.; Niki, E.; et al. Protein-bound acrolein: Potential markers for oxidative stress. Proc. Natl. Acad Sci. USA 1998, 95, 4882–4887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, H.R. Antioxidants and protein oxidation. Free Radic. Res. 2000, 33, S47–S58. [Google Scholar]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein carbonylation as a major hallmark of oxidative damage: Update of analytical strategies. Mass Spectrom. Rev. 2014, 33, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Ogawa, Y.; Yoshida, R.; Shimamori, A.; Kasai, H.; Ohta, H. The stability of the oxidative stress marker, urinary 8-hydroxy-2’- deoxyguanosine (8-OHdG), when stored at room temperature. J. Occup. Health 2008, 50, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Bordone, R.; Perugini, C.; Bagnati, M.; Cau, C.; Bellomo, G. The kinetics of copper-induced LDL oxidation depend upon its lipid composition and antioxidant content. BioChem. Biophys. Res. Commun. 2000, 268, 818–822. [Google Scholar] [CrossRef]
- Ghaffari, M.A.; Ghiasvand, T. Kinetic study of low density lipoprotein oxidation by copper. Indian J. Clin. Biochem. 2010, 25, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Moniruzzaman, M.; Khalil, M.I.; Sulaiman, S.A.; Gan, S.H. Advances in the analytical methods for determining the antioxidant properties of honey: A review. Afr. J. Tradit. Complement. Altern. Med. 2012, 9, 36–42. [Google Scholar] [CrossRef]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. 2018, 12, 88–93. [Google Scholar]
- Marrocco, I.; Altieri, F.; Peluso, I. Measurement and clinical significance of biomarkers of oxidative stress in humans. Oxid. Med. Cell Longev. 2017, 2017, 6501046. [Google Scholar] [CrossRef]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. AntiOxid. Redox Signal 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [PubMed]
- Sepasi Tehrani, H.; Moosavi-Movahedi, A.A. Catalase and its mysteries. Prog. Biophys. Mol. Biol. 2018, 140, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B., 3rd. Diabetes, oxidative stress, and antioxidants: A review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef]
- Bohm, B.; Heinzelmann, S.; Motz, M.; Bauer, G. Extracellular localization of catalase is associated with the transformed state of malignant cells. Biol. Chem. 2015, 396, 1339–1356. [Google Scholar] [CrossRef] [Green Version]
- De Jager, W.; Bourcier, K.; Rijkers, G.T.; Prakken, B.J.; Seyfert-Margolis, V. Prerequisites for cytokine measurements in clinical trials with multiplex immunoassays. BMC Immunol. 2009, 10, 52. [Google Scholar] [CrossRef]
- Monastero, R.N.; Pentyala, S. Cytokines as biomarkers and their respective clinical cutoff levels. Int. J. Inflamm. 2017, 2017, 4309485. [Google Scholar] [CrossRef] [PubMed]
- Mühl, H.; Pfeilschifter, J. Anti-inflammatory properties of pro-inflammatory interferon-gamma. Int. Immunopharmacol. 2003, 3, 1247–1255. [Google Scholar] [CrossRef]
- Di Paolo, N.C.; Shayakhmetov, D.M. Interleukin 1alpha and the inflammatory process. Nat. Immunol. 2016, 17, 906–913. [Google Scholar] [CrossRef]
- Kenis, G.; Teunissen, C.; De Jongh, R.; Bosmans, E.; Steinbusch, H.; Maes, M. Stability of interleukin 6, soluble interleukin 6 receptor, interleukin 10 and CC16 in human serum. Cytokine 2002, 19, 228–235. [Google Scholar] [CrossRef]
- Hunter, C.A.; Jones, S.A. IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef]
- Berry, L.J.; Sheil, B.; Garratt, L.; Sly, P.D.; AREST, C.F. Stability of interleukin 8 and neutrophil elastase in bronchoalveolar lavage fluid following long-term storage. J. Cyst. Fibr. 2010, 9, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Doumatey, A.P.; Zhou, J.; Adeyemo, A.; Rotimi, C. High sensitivity C-reactive protein (Hs-CRP) remains highly stable in long-term archived human serum. Clin. Biochem. 2014, 47, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Sproston, N.R.; Ashworth, J.J. Role of C-reactive protein at sites of inflammation and infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef]
- Sack, G.H., Jr. Serum amyloid A—A review. Mol. Med. 2018, 24, 46. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Proinflammatory cytokines. Chest 2000, 118, 503–508. [Google Scholar] [CrossRef]
- Alfaro, C.; Sanmamed, M.F.; Rodríguez-Ruiz, M.E.; Teijeira, Á.; Oñate, C.; González, Á.; Ponz, M.; Schalper, K.A.; Pérez-Gracia, J.L.; Melero, I. Interleukin-8 in cancer pathogenesis, treatment and follow-up. Cancer Treat. Rev. 2017, 60, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Polidori, M.C.; Pratico, D.; Parente, B.; Mariani, E.; Cecchetti, R.; Yao, Y.; Sies, H.; Cao, P.; Mecocci, P.; Stahl, W. Elevated lipid peroxidation biomarkers and low antioxidant status in atherosclerotic patients with increased carotid or iliofemoral intima media thickness. J. Invest. Med. 2007, 55, 163–167. [Google Scholar] [CrossRef]
- Martin-Gallan, P.; Carrascosa, A.; Gussinye, M.; Domínguez, C. Biomarkers of diabetes-associated oxidative stress and antioxidant status in young diabetic patients with or without subclinical complications. Free Radic. Biol. Med. 2003, 34, 1563–1574. [Google Scholar] [CrossRef]
- Facchini, F.S.; Humphreys, M.H.; DoNascimento, C.A.; Abbasi, F.; Reaven, G.M. Relation between insulin resistance and plasma concentrations of lipid hydroperoxides, carotenoids, and tocopherols. Am. J. Clin. Nutr. 2000, 72, 776–779. [Google Scholar] [CrossRef] [Green Version]
- Kisakol, G.; Guney, E.; Bayraktar, F.; Yilmaz, C.; Kabalak, T.; Ozmen, D. Effect of surgical weight loss on free radical and antioxidant balance: A preliminary report. Obes. Surg. 2002, 12, 795–800. [Google Scholar] [CrossRef]
- Senthil, S.; Veerappan, R.M.; Ramakrishna Rao, M.; Pugalendi, K.V. Oxidative stress and antioxidants in patients with cardiogenic shock complicating acute myocardial infarction. Clin. Chim Acta 2004, 348, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Polidori, M.C.; Praticó, D.; Savino, K.; Rokach, J.; Stahl, W.; Mecocci, P. Increased F2 isoprostane plasma levels in patients with congestive heart failure are correlated with antioxidant status and disease severity. J. Card. Fail. 2004, 10, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Serdar, Z.; Serdar, A.; Altin, A.; Eryilmaz, U.; Albayrak, S. The relation between oxidant and antioxidant parameters and severity of acute coronary syndromes. Acta Cardiol. 2007, 62, 373–380. [Google Scholar] [CrossRef]
- Chang, C.Y.; Chen, J.Y.; Ke, D.; Hu, M.L. Plasma levels of lipophilic antioxidant vitamins in acute ischemic stroke patients: Correlation to inflammation markers and neurological deficits. Nutrition 2005, 21, 987–993. [Google Scholar] [CrossRef]
- Kataria, Y.; Deaton, R.J.; Enk, E.; Jin, M.; Petrauskaite, M.; Dong, L.; Goldenberg, J.R.; Cotler, S.J.; Jensen, D.M.; van Breemen, R.B.; et al. Retinoid and carotenoid status in serum and liver among patients at high-risk for liver cancer. BMC Gastroenterol. 2016, 16, 30. [Google Scholar] [CrossRef]
- Epplein, M.; Franke, A.A.; Cooney, R.V.; Morris, J.S.; Wilkens, L.R.; Goodman, M.T.; Murphy, S.P.; Henderson, B.E.; Kolonel, L.N.; Le Marchand, L. Association of plasma micronutrient levels and urinary isoprostane with risk of lung cancer: The multiethnic cohort study. Cancer Epidemiol. Prev. Biomark. 2009, 18, 1962–1970. [Google Scholar] [CrossRef]
- Caliskan-Can, E.; Firat, H.; Ardic, S.; Simşek, B.; Torun, M.; Yardim-Akaydin, S. Increased levels of 8-hydroxydeoxyguanosine and its relationship with lipid peroxidation and antioxidant vitamins in lung cancer. Clin. Chem. Lab. Med. 2008, 46, 107–112. [Google Scholar] [CrossRef]
- Tsao, S.M.; Yin, M.C.; Liu, W.H. Oxidant stress and B vitamins status in patients with non-small cell lung cancer. Nutr. Cancer 2007, 59, 8–13. [Google Scholar] [CrossRef]
- Almushatat, A.S.; Talwar, D.; McArdle, P.A.; Williamson, C.; Sattar, N.; O’Reilly, D.S.; Underwood, M.A.; McMillan, D.C. Vitamin antioxidants, lipid peroxidation and the systemic inflammatory response in patients with prostate cancer. Int. J. Cancer 2006, 118, 1051–1053. [Google Scholar] [CrossRef]
- Roehrs, M.; Valentini, J.; Paniz, C.; Moro, A.; Charão, M.; Bulcão, R.; Freitas, F.; Brucker, N.; Duarte, M.; Leal, M.; et al. The relationships between exogenous and endogenous antioxidants with the lipid profile and oxidative damage in hemodialysis patients. BMC Nephrol. 2011, 12, 59. [Google Scholar] [CrossRef]
- Charao, M.F.; Moro, A.M.; Valentini, J.; Brucker, N.; Bubols, G.B.; Bulcão, R.P.; Baierle, M.; Freitas, F.A.; Nascimento, S.N.; Barth, A.; et al. Exogenous and endogenous antioxidants attenuate the lipid peroxidation in workers occupationally exposed to paints. Drug Chem. Toxicol. 2014, 37, 69–75. [Google Scholar] [CrossRef]
- Ren, H.; Ghebremeskel, K.; Okpala, I.; Lee, A.; Ibegbulam, O.; Crawford, M. Patients with sickle cell disease have reduced blood antioxidant protection. Int. J. Vitam. Nutr. Res. 2008, 78, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Hebbel, R.P.; Eaton, J.W.; Balasingam, M.; Steinberg, M.H. Spontaneous oxygen radical generation by sickle erythrocytes. J. Clin. Invest. 1982, 70, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Maor, I.; Rainis, T.; Lanir, A.; Lavy, A. Oxidative stress, inflammation and neutrophil superoxide release in patients with Crohn’s disease: Distinction between active and non-active disease. Dig. Dis. Sci. 2008, 53, 2208–2214. [Google Scholar] [CrossRef] [PubMed]
- Cemek, M.; Dede, S.; Bayiroglu, F.; Caksen, H.; Cemek, F.; Yuca, K. Oxidant and antioxidant levels in children with acute otitis media and tonsillitis: A comparative study. Int. J. Pediatr. Otorhinolaryngol. 2005, 69, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, J.; Bohn, T. Exogenous antioxidants—Double-edged swords in cellular redox state: Health beneficial effects at physiologic doses versus deleterious effects at high doses. Oxid. Med. Cell Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Haldar, S.; Rowland, I.R.; Barnett, Y.A.; Bradbury, I.; Robson, P.J.; Powell, J.; Fletcher, J. Influence of habitual diet on antioxidant status: A study in a population of vegetarians and omnivores. Eur. J. Clin. Nutr. 2007, 61, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, Y.J.; Lim, Y.; Oh, B.; Kim, J.Y.; Bouwman, J.; Kwon, O. Combination of diet quality score, plasma carotenoids, and lipid peroxidation to monitor oxidative stress. Oxid. Med. Cell Longev. 2018, 2018, 8601028. [Google Scholar] [CrossRef] [PubMed]
- Orhon, F.S.; Ulukol, B.; Kahya, D.; Cengiz, B.; Başkan, S.; Tezcan, S. The influence of maternal smoking on maternal and newborn oxidant and antioxidant status. Eur. J. Pediatr. 2009, 168, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Suwannalert, P.; Boonsiri, P.; Khampitak, T.; Khampitak, K.; Sriboonlue, P.; Yongvanit, P. The levels of lycopene, alpha-tocopherol and a marker of oxidative stress in healthy northeast Thai elderly. Asia Pac. J. Clin. Nutr. 2007, 16 Suppl 1, 27–30. [Google Scholar] [PubMed]
- Zhou, Y.E.; Buchowski, M.S.; Liu, J.; Schlundt, D.G.; Ukoli, F.; Blot, W.J.; Hargreaves, M.K. Plasma lycopene is associated with pizza and pasta consumption in middle-aged and older african american and white adults in the southeastern USA in a cross-sectional study. PLoS ONE 2016, 11, e0161918. [Google Scholar] [CrossRef]
- Rink, S.M.; Mendola, P.; Mumford, S.L.; Poudrier, J.K.; Browne, R.W.; Wactawski-Wende, J.; Perkins, N.J.; Schisterman, E.F. Self-report of fruit and vegetable intake that meets the 5 a day recommendation is associated with reduced levels of oxidative stress biomarkers and increased levels of antioxidant defense in premenopausal women. J. Acad. Nutr. Diet. 2013, 113, 776–785. [Google Scholar] [CrossRef]
- Weber, D.; Stuetz, W.; Toussaint, O.; Debacq-Chainiaux, F.; Dollé, M.E.T.; Jansen, E.; Gonos, E.S.; Franceschi, C.; Sikora, E.; Hervonen, A.; et al. Associations between specific redox biomarkers and age in a large European cohort: The MARK-AGE project. Oxid. Med. Cell Longev. 2017, 2017, 1401452. [Google Scholar] [CrossRef]
- Parfitt, V.J.; Rubba, P.; Bolton, C.; Marotta, G.; Hartog, M.; Mancini, M. A comparison of antioxidant status and free radical peroxidation of plasma lipoproteins in healthy young persons from Naples and Bristol. Eur. Heart J. 1994, 15, 871–876. [Google Scholar] [CrossRef]
- Pincemail, J.; Vanbelle, S.; Gaspard, U.; Collette, G.; Haleng, J.; Cheramy-Bien, J.P.; Charlier, C.; Chapelle, J.P.; Giet, D.; Albert, A.; et al. Effect of different contraceptive methods on the oxidative stress status in women aged 40 48 years from the ELAN study in the province of liege, belgium. Hum. Reprod. 2007, 22, 2335–2343. [Google Scholar] [CrossRef] [PubMed]
- Mazereeuw, G.; Herrmann, N.; Andreazza, A.C.; Khan, M.M.; Lanctôt, K.L. A meta-analysis of lipid peroxidation markers in major depression. Neuropsychiatr. Dis. Treat. 2015, 11, 2479–2491. [Google Scholar] [PubMed] [Green Version]
- Tsuboi, H.; Shimoi, K.; Kinae, N.; Oguni, I.; Hori, R.; Kobayashi, F. Depressive symptoms are independently correlated with lipid peroxidation in a female population: Comparison with vitamins and carotenoids. J. Psychosom. Res. 2004, 56, 53–58. [Google Scholar] [CrossRef]
- Berr, C.; Richard, M.J.; Roussel, A.M.; Bonithon-Kopp, C. Systemic oxidative stress and cognitive performance in the population-based EVA study. Etude du vieillissement arteriel. Free Radic. Biol. Med. 1998, 24, 1202–1208. [Google Scholar] [CrossRef]
- Rehman, K.; Akash, M.S.H. Mechanism of generation of oxidative stress and pathophysiology of type 2 diabetes mellitus: How are they interlinked? J. Cell Biochem. 2017, 118, 3577–3585. [Google Scholar] [CrossRef]
- Gill, J.G.; Piskounova, E.; Morrison, S.J. Cancer, oxidative stress, and metastasis. Cold Spring Harb. Symp. Quant. Biol. 2016, 81, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Hamer, M.; Chida, Y. Intake of fruit, vegetables, and antioxidants and risk of type 2 diabetes: Systematic review and meta-analysis. J. Hypertens. 2007, 25, 2361–2369. [Google Scholar] [CrossRef]
- Shardell, M.D.; Alley, D.E.; Hicks, G.E.; Etl-Kamary, S.S.; Miller, R.R.; Semba, R.D.; Ferrucci, L. Low-serum carotenoid concentrations and carotenoid interactions predict mortality in US adults: The third national health and nutrition examination survey. Nutr. Res. 2011, 31, 178–189. [Google Scholar] [CrossRef]
- Zhao, L.G.; Zhang, Q.L.; Zheng, J.L.; Li, H.L.; Zhang, W.; Tang, W.G.; Xiang, Y.B. Dietary, circulating beta-carotene and risk of all-cause mortality: A meta-analysis from prospective studies. Sci. Rep. 2016, 6, 26983. [Google Scholar] [CrossRef]
- Huang, J.; Weinstein, S.J.; Yu, K.; Männistö, S.; Albanes, D. Serum beta carotene and overall and cause-specific mortality. Circ. Res. 2018, 123, 1339–1349. [Google Scholar] [CrossRef]
- Besler, H.T.; Comoglu, S.; Okcu, Z. Serum levels of antioxidant vitamins and lipid peroxidation in multiple sclerosis. Nutr. Neurosci. 2002, 5, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Codoner-Franch, P.; Pons-Morales, S.; Boix-Garcia, L.; Valls-Bellés, V. Oxidant/antioxidant status in obese children compared to pediatric patients with type 1 diabetes mellitus. Pediatr. Diabetes 2010, 11, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Polidori, M.C.; Savino, K.; Alunni, G.; Freddio, M.; Senin, U.; Freddio, M.; Senin, U.; Sies, H.; Stahl, W.; Mecocci, P. Plasma lipophilic antioxidants and malondialdehyde in congestive heart failure patients: Relationship to disease severity. Free Radic. Biol. Med. 2002, 32, 148–152. [Google Scholar] [CrossRef]
- Naziroglu, M.; Kokcam, I.; Simsek, H.; Karakılçık, A.; Naziroǧlu, M. Lipid peroxidation and antioxidants in plasma and red blood cells from patients with pemphigus vulgaris. J. Basic Clin. Physiol Pharmacol. 2003, 14, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T. Bioavailabilty of non-provitamin A carotenoids. Curr. Nutr. Food Sci. 2008, 4, 240–258. [Google Scholar] [CrossRef]
- Bohn, T. Provitamin A carotenoids—Occurrence, intake, and bioavailability. In Vitamin A and Carotenoids: Chemistry, Analysis, Function and Effects (Food and Nutritional Components in Focus); Preedy, V., Ed.; RSC Publishing: London, UK, 2012. [Google Scholar]
- Winklhofer-Roob, B.M.; Puhl, H.; Khoschsorur, G.; Hof, M.A.V.; Esterbauer, H.; Shmerling, D.H. Enhanced resistance to oxidation of low density lipoproteins and decreased lipid peroxide formation during beta-carotene supplementation in cystic fibrosis. Free Radic. Biol. Med. 1995, 18, 849–859. [Google Scholar] [CrossRef]
- Renner, S.; Rath, R.; Rust, P.; Lehr, S.; Frischer, T.; Elmadfa, I.; Eichler, I. Effects of beta-carotene supplementation for six months on clinical and laboratory parameters in patients with cystic fibrosis. Thorax 2001, 56, 48–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, Y.; Zaltsberg, H.; Ben-Amotz, A.; Levy, Y.; Aviram, M. Dietary supplementation of a natural isomer mixture of beta-carotene inhibits oxidation of LDL derived from patients with diabetes mellitus. Ann. Nutr. Metab. 2000, 44, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Allard, J.P.; Royall, D.; Kurian, R.; Muggli, R.; Jeejeebhoy, K.N. Effects of beta-carotene supplementation on lipid peroxidation in humans. Am. J. Clin. Nutr. 1994, 59, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Chang, M.J.; Choi, H.D.; Youn, Y.K.; Kim, J.T.; Oh, J.M. Protective effects of Haematococcus astaxanthin on oxidative stress in healthy smokers. J. Med. Food 2011, 14, 1469–1475. [Google Scholar] [CrossRef] [PubMed]
- Naguib, Y.M. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef] [PubMed]
- Hininger, I.; Chopra, M.; Thurnham, D.I.; Laporte, F.; Richard, M.J.; Favier, A.; Roussel, A.M. Effect of increased fruit and vegetable intake on the susceptibility of lipoprotein to oxidation in smokers. Eur. J. Clin. Nutr. 1997, 51, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Ademowo, O.S.; Dias, H.K.I.; Milic, I.; Devitt, A.; Moran, R.; Mulcahy, R.; Howard, A.; Nolan, J.; Griffiths, H.; Dias, I. Phospholipid oxidation and carotenoid supplementation in Alzheimer’s disease patients. Free Radic. Biol. Med. 2017, 108, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Souci, S.W.; Fachmann, W.; Kraut, H. Food Compoisition and Nutrition Tables; CRC press: Boca Raton, FL, USA, 2000; Volume 6. [Google Scholar]
- Catanzaro, R.; Zerbinati, N.; Solimene, U.; Marcellino, M.; Mohania, D.; Italia, A.; Ayala, A.; Marotta, F. Beneficial effect of refined red palm oil on lipid peroxidation and monocyte tissue factor in HCV-related liver disease: A randomized controlled study. Hepatobiliary Pancreat Dis. Int. 2016, 15, 165–172. [Google Scholar] [CrossRef]
- Sarkar, P.D.; Gupt, T.; Sahu, A. Comparative analysis of lycopene in oxidative stress. J. Assoc. Physicians India 2012, 60, 17–19. [Google Scholar] [PubMed]
- Bose, K.S.; Agrawal, B.K. Effect of lycopene from tomatoes (cooked) on plasma antioxidant enzymes, lipid peroxidation rate and lipid profile in grade-I hypertension. Ann. Nutr. Metab. 2007, 51, 477–481. [Google Scholar] [CrossRef]
- Giovannucci, E. Commentary: Serum lycopene and prostate cancer progression: A re-consideration of findings from the prostate cancer prevention trial. Cancer Causes Control 2011, 22, 1055–1059. [Google Scholar] [CrossRef] [PubMed]
- Giovannucci, E.; Liu, Y.; Platz, E.A.; Stampfer, M.J.; Willett, W.C. Risk factors for prostate cancer incidence and progression in the health professionals follow-up study. Int. J. Cancer 2007, 121, 1571–1578. [Google Scholar] [CrossRef] [Green Version]
- Van Breemen, R.B.; Sharifi, R.; Viana, M.; Pajkovic, N.; Zhu, D.; Yuan, L.; Yang, Y.; Bowen, P.E.; Stacewicz-Sapuntzakis, M. Antioxidant effects of lycopene in African American men with prostate cancer or benign prostate hyperplasia: A randomized, controlled trial. Cancer Prev. Res. 2011, 4, 711–718. [Google Scholar] [CrossRef]
- Elmadfa, I.; Rust, P.; Majchrzak, D.; Wagner, K.H.; Genser, D.; Lettner, R.; Pinter, M. Effects of beta-carotene supplementation on free radical mechanism in healthy adult subjects. Int. J. Vitam. Nutr. Res. 2004, 74, 147–152. [Google Scholar] [CrossRef]
- Mobarhan, S.; Bowen, P.; Andersen, B.; Evans, M.; Stacewicz-Sapuntzakis, M.; Sugerman, S.; Simms, P.; Lucchesi, D.; Friedman, H. Effects of beta-carotene repletion on beta-carotene absorption, lipid peroxidation, and neutrophil superoxide formation in young men. Nutr. Cancer 1990, 14, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, A.; Madarame, T.; Koike, H.; Komatsu, Y.; Wise, J.A. Four week supplementation with mixed fruit and vegetable juice concentrates increased protective serum antioxidants and folate and decreased plasma homocysteine in Japanese subjects. Asia Pac. J. Clin. Nutr. 2007, 16, 411–421. [Google Scholar] [PubMed]
- Hininger, I.A.; Meyer-Wenger, A.; Moser, U.; Wright, A.; Southon, S.; Thurnham, D.; Chopra, M.; Berg, H.V.D.; Olmedilla, B.; Favier, A.E.; et al. No significant effects of lutein, lycopene or beta-carotene supplementation on biological markers of oxidative stress and LDL oxidizability in healthy adult subjects. J. Am. Coll. Nutr. 2001, 20, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.X.; Jiao, J.H.; Li, Z.Y.; Liu, R.-R.; Shi, Q.; Ma, L. Lutein supplementation reduces plasma lipid peroxidation and C-reactive protein in healthy nonsmokers. Atherosclerosis 2013, 227, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Perrone, S.; Tei, M.; Longini, M.; Santacroce, A.; Turrisi, G.; Proietti, F.; Felici, C.; Picardi, A.; Bazzini, F.; Vasarri, P.; et al. Lipid and protein oxidation in newborn infants after lutein administration. Oxid. Med. Cell Longev. 2014, 2014, 781454. [Google Scholar] [CrossRef] [PubMed]
- Perrone, S.; Longini, M.; Marzocchi, B.; Picardi, A.; Bellieni, C.V.; Proietti, F.; Rodriguez, A.; Turrisi, G.; Buonocore, G. Effects of lutein on oxidative stress in the term newborn: A pilot study. Neonatology 2010, 97, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.P.; Chan, G.M.; Barrett-Reis, B.M.; Fulton, A.B.; Hansen, R.M.; Ashmeade, T.L.; Oliver, J.S.; Mackey, A.D.; Dimmit, R.A.; Hartmann, E.E.; et al. Effect of carotenoid supplementation on plasma carotenoids, inflammation and visual development in preterm infants. J. Perinatol. 2011, 32, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, T.; Nakagawa, K.; Takekoshi, H.; Higuchi, O.; Kato, S.; Kondo, M.; Kimura, F.; Miyazawa, T. Ingestion of chlorella reduced the oxidation of erythrocyte membrane lipids in senior Japanese subjects. J. Oleo. Sci. 2013, 62, 873–881. [Google Scholar] [CrossRef]
- Nakagawa, K.; Kiko, T.; Miyazawa, T.; Kimura, F.; Satoh, A.; Miyazawa, T. Antioxidant effect of astaxanthin on phospholipid peroxidation in human erythrocytes. Br. J. Nutr. 2011, 105, 1563–1571. [Google Scholar] [CrossRef] [Green Version]
- Devaraj, S.; Mathur, S.; Basu, A.; Aung, H.H.; Vasu, V.T.; Meyers, S.; Jialal, I. A dose-response study on the effects of purified lycopene supplementation on biomarkers of oxidative stress. J. Am. Coll. Nutr. 2008, 27, 267–273. [Google Scholar] [CrossRef]
- Briviba, K.; Bub, A.; Moseneder, J.; Schwerdtle, T.; Hartwig, A.; Kulling, S.E.; Watzl, B. No differences in DNA damage and antioxidant capacity between intervention groups of healthy, nonsmoking men receiving 2, 5, or 8 servings/day of vegetables and fruit. Nutr. Cancer 2008, 60, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Misra, R.; Mangi, S.; Joshi, S.; Mittal, S.; Gupta, S.K.; Pandey, R.M. LycoRed as an alternative to hormone replacement therapy in lowering serum lipids and oxidative stress markers: A randomized controlled clinical trial. J. Obstet. Gynaecol. Res. 2006, 32, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Engelhard, Y.N.; Gazer, B.; Paran, E. Natural antioxidants from tomato extract reduce blood pressure in patients with grade-1 hypertension: A double-blind, placebo-controlled pilot study. Am. Heart J. 2006, 151, 100. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, E.S.; Rao, A.V.; Rao, L.G. Dietary restriction of lycopene for a period of one month resulted in significantly increased biomarkers of oxidative stress and bone resorption in postmenopausal women. J. Nutr. Health Aging 2011, 15, 133–138. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Anti-oxidative action of resveratrol: Implications for human health. Arab. J. Chem. 2011, 4, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Jowko, E.; Dlugolecka, B.; Makaruk, B.; Cieśliński, I. The effect of green tea extract supplementation on exercise-induced oxidative stress parameters in male sprinters. Eur. J. Nutr. 2015, 54, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Minzanova, T.S.; Mironov, F.V.; Arkhipova, M.D.; Khabibullina, A.V.; Mironova, L.G.; Zakirova, Y.M.; Milyukov, V.A. Biological activity and pharmacological application of pectic polysaccharides: A review. Polymers 2018, 10, 1407. [Google Scholar] [CrossRef]
- Haegele, A.D.; Gillette, C.; O’Neill, C.; Wolfe, P.; Heimendinger, J.; Sedlacek, S.; Thompson, H.J. Plasma xanthophyll carotenoids correlate inversely with indices of oxidative DNA damage and lipid peroxidation. Cancer Epidemiol. Biomark. Prev. 2000, 9, 421–425. [Google Scholar]
- Thompson, H.J.; Heimendinger, J.; Diker, A.; Meinecke, B.; Wolfe, P.; Zhu, Z.; Heimendinger, J.; Diker, A.; Sedlacek, S.; Jiang, W. Dietary botanical diversity affects the reduction of oxidative biomarkers in women due to high vegetable and fruit intake. J. Nutr. 2006, 136, 2207–2212. [Google Scholar] [CrossRef]
- The Alpha-tocopherol Beta-Carotene Cancer Prevention Group. The effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers. The alpha-tocopherol, beta carotene cancer prevention study group. N Engl. J. Med. 1994, 330, 1029–1035. [Google Scholar] [CrossRef]
- Druesne-Pecollo, N.; Latino-Martel, P.; Norat, T.; Barrandon, E.; Bertrais, S.; Galan, P.; Hercberg, S. Beta-carotene supplementation and cancer risk: A systematic review and metaanalysis of randomized controlled trials. Int. J. Cancer 2010, 127, 172–184. [Google Scholar] [CrossRef]
- Hennekens, C.H.; Buring, J.E.; Manson, J.E.; Stampfer, M.; Rosner, B.; Cook, N.R.; Belanger, C.; LaMotte, F.; Gaziano, J.M.; Ridker, P.M.; et al. Lack of effect of long-term supplementation with beta carotene on the incidence of malignant neoplasms and cardiovascular disease. N Engl. J. Med. 1996, 334, 1145–1149. [Google Scholar] [CrossRef] [PubMed]
- Heart Protection Study Collaborative Group. MRC/BHF heart protection study of antioxidant vitamin supplementation in 20,536 high-risk individuals: A randomised placebo-controlled trial. Lancet 2002, 360, 23–33. [Google Scholar] [CrossRef]
- Blot, W.J.; Li, J.Y.; Taylor, P.R.; Guo, W.; Dawsey, S.M.; Li, B. The Linxian trials: Mortality rates by vitamin-mineral intervention group. Am. J. Clin. Nutr. 1995, 62, 1424S–1426S. [Google Scholar] [CrossRef] [PubMed]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. Cochrane Database Syst. Rev. 2012, 14, Cd007176. [Google Scholar] [CrossRef]
- Xu, X.R.; Zou, Z.Y.; Xiao, X.; Huang, Y.M.; Wang, X.; Lin, X.M. Effects of lutein supplement on serum inflammatory cytokines, ApoE and lipid profiles in early atherosclerosis population. J. Atheroscler. Thromb. 2013, 20, 170–177. [Google Scholar] [CrossRef] [PubMed]
- McEneny, J.; Wade, L.; Young, I.S.; Masson, L.; Duthie, G.; McGinty, A.; McMaster, C.; Thies, F. Lycopene intervention reduces inflammation and improves HDL functionality in moderately overweight middle-aged individuals. J. Nutr. Biochem. 2013, 24, 163–168. [Google Scholar] [CrossRef]
- Riso, P.; Visioli, F.; Grande, S.; Guarnieri, S.; Gardana, C.; Simonetti, P.; Porrini, M. Effect of a tomato-based drink on markers of inflammation, immunomodulation, and oxidative stress. J. Agric. Food Chem. 2006, 54, 2563–2566. [Google Scholar] [CrossRef]
- Biddle, M.J.; Lennie, T.A.; Bricker, G.V.; Kopec, R.E.; Schwartz, S.J.; Moser, D.K. Lycopene dietary intervention: A pilot study in patients with heart failure. J. Cardiovasc. Nurs. 2015, 30, 205–212. [Google Scholar] [CrossRef]
- Watzl, B.; Bub, A.; Blockhaus, M.; Herbert, B.M.; Lührmann, P.M.; Neuhäuser-Berthold, M.; Rechkemmer, G. Prolonged tomato juice consumption has no effect on cell-mediated immunity of well-nourished elderly men and women. J. Nutr. 2000, 130, 1719–1723. [Google Scholar] [CrossRef]
- Lee, C. Collaborative power of Nrf2 and PPARgamma activators against metabolic and drug-induced oxidative injury. Oxid. Med. Cell Longev. 2017, 2017, 1378175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, W.E.; Hu, L.; Zhao, L.; Huang, J. Carotenoids inhibit proliferation and regulate expression of peroxisome proliferators-activated receptor gamma (PPARgamma) in K562 cancer cells. Arch. Biochem. Biophys. 2011, 512, 96–106. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bohn, T. Carotenoids and Markers of Oxidative Stress in Human Observational Studies and Intervention Trials: Implications for Chronic Diseases. Antioxidants 2019, 8, 179. https://doi.org/10.3390/antiox8060179
Bohn T. Carotenoids and Markers of Oxidative Stress in Human Observational Studies and Intervention Trials: Implications for Chronic Diseases. Antioxidants. 2019; 8(6):179. https://doi.org/10.3390/antiox8060179
Chicago/Turabian StyleBohn, Torsten. 2019. "Carotenoids and Markers of Oxidative Stress in Human Observational Studies and Intervention Trials: Implications for Chronic Diseases" Antioxidants 8, no. 6: 179. https://doi.org/10.3390/antiox8060179