Water- and Fat-Soluble Antioxidants in Human Seminal Plasma and Serum of Fertile Males


 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling of Serum and Seminal Plasma
2.2. Sample Processing and HPLC Analyses
2.3. Statistics
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bardaweel, S.K.; Gul, M.; Alzweiri, M.; Ishaqat, A.; ALSalamat, H.A.; Bashatwah, R.M. Reactive Oxygen Species: The dual role in physiological and pathological conditions of the human body. Eurasian J. Med. 2018, 50, 193–201. [Google Scholar] [CrossRef]
- Ježek, J.; Cooper, K.F.; Strich, R. Reactive Oxygen Species and mitochondrial dynamics: The Yin and Yang of mitochondrial dysfunction and cancer progression. Antioxidants 2018, 7, 13. [Google Scholar] [CrossRef]
- Di Pietro, V.; Amorini, A.M.; Tavazzi, B.; Vagnozzi, R.; Logan, A.; Lazzarino, G.; Signoretti, S.; Lazzarino, G.; Belli, A. The molecular mechanisms affecting N-acetylaspartate homeostasis following experimental graded traumatic brain injury. Mol. Med. 2014, 20, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, V.; Lazzarino, G.; Amorini, A.M.; Signoretti, S.; Hill, L.J.; Porto, E.; Tavazzi, B.; Lazzarino, G.; Belli, A. Fusion or fission: The destiny of mitochondria in traumatic brain injury of different severities. Sci. Rep. 2017, 7, 9198. [Google Scholar] [CrossRef]
- Petzold, A.; Nijland, P.G.; Balk, L.J.; Amorini, A.M.; Lazzarino, G.; Wattjes, M.P.; Gasperini, C.; van der Valk, P.; Tavazzi, B.; Lazzarino, G.; et al. Visual pathway neurodegeneration winged by mitochondrial dysfunction. Ann. Clin. Transl. Neurol. 2015, 2, 140–150. [Google Scholar]
- Zhang, R.; Xu, M.; Wang, Y.; Xie, F.; Zhang, G.; Qin, X. Nrf2-a promising therapeutic target for defensing against oxidative stress in stroke. Mol. Neurobiol. 2017, 54, 6006–6017. [Google Scholar] [CrossRef]
- Youn, S.W.; Li, Y.; Kim, Y.M.; Sudhahar, V.; Abdelsaid, K.; Kim, H.W.; Liu, Y.; Fulton, D.J.R.; Ashraf, M.; Tang, Y.; et al. Modification of cardiac progenitor cell-derived exosomes by mir-322 provides protection against myocardial infarction through Nox2-dependent angiogenesis. Antioxidants 2019, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Kamkar, N.; Ramezanali, F.; Sabbaghian, M. The relationship between sperm DNA fragmentation, free radicals and antioxidant capacity with idiopathic repeated pregnancy loss. Reprod. Biol. 2018, 18, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.; Kurpisz, M. Inflammatory mediators exert toxic effects of oxidative stress on human spermatozoa. J. Androl. 2007, 28, 325–333. [Google Scholar] [CrossRef]
- Colitti, M.; Stefanon, B.; Gabai, G.; Gelain, M.E.; Bonsembiante, F. Oxidative stress and nutraceuticals in the modulation of the immune function: Current knowledge in animals of veterinary interest. Antioxidants 2019, 8, 28. [Google Scholar] [CrossRef]
- Cesar, V.; Jozić, I.; Begović, L.; Vuković, T.; Mlinarić, S.; Lepeduš, H.; Borović Šunjić, S.; Žarković, N. Cell-type-specific modulation of hydrogen peroxide cytotoxicity and 4-hydroxynonenal binding to human cellular proteins in vitro by antioxidant Aloe vera extract. Antioxidants 2018, 7, 125. [Google Scholar] [CrossRef]
- Belardin, L.B.; Antoniassi, M.P.; Camargo, M.; Intasqui, P.; Fraietta, R.; Bertolla, R.P. Semen levels of matrix metalloproteinase (MMP) and tissue inhibitor of metallorproteinases (TIMP) protein families members in men with high and low sperm DNA fragmentation. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Sun, S.; Xie, F.; Xu, X.; Cai, Q.; Zhang, Q.; Cui, Z.; Zheng, Y.; Zhou, J. Advanced oxidation protein products induce S-phase arrest of hepatocytes via the ROS-dependent, β-catenin-CDK2-mediated pathway. Redox Biol. 2018, 14, 338–353. [Google Scholar] [CrossRef]
- Radak, Z.; Zhao, Z.; Goto, S.; Koltai, E. Age-associated neurodegeneration and oxidative damage to lipids, proteins and DNA. Mol. Asp. Med. 2011, 32, 305–315. [Google Scholar] [CrossRef]
- Cui, C.; Song, S.; Cui, J.; Feng, Y.; Gao, J.; Jiang, P. Vitamin D receptor activation influences NADPH oxidase (Nox2) activity and protects against neurological deficits and apoptosis in a rat model of traumatic brain injury. Oxid. Med. Cell Longev. 2017, 2017. [Google Scholar] [CrossRef]
- Su, B.C.; Pan, C.Y.; Chen, J.Y. Antimicrobial peptide TP4 induces ROS-mediated necrosis by triggering mitochondrial dysfunction in wild-type and mutant p53 glioblastoma cells. Cancers 2019, 11, 171. [Google Scholar] [CrossRef]
- Fresta, C.G.; Chakraborty, A.; Wijesinghe, M.B.; Amorini, A.M.; Lazzarino, G.; Lazzarino, G.; Tavazzi, B.; Lunte, S.M.; Caraci, F.; Dhar, P.; et al. Non-toxic engineered carbon nanodiamond concentrations induce oxidative/nitrosative stress, imbalance of energy metabolism, and mitochondrial dysfunction in microglial and alveolar basal epithelial cells. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef]
- Aoyama, K.; Nakaki, T. Glutathione in cellular redox homeostasis: Association with the excitatory amino acid carrier 1 (EAAC1). Molecules 2015, 20, 8742–8758. [Google Scholar] [CrossRef]
- Di Pietro, V.; Lazzarino, G.; Amorini, A.M.; Tavazzi, B.; D’Urso, S.; Longo, S.; Vagnozzi, R.; Signoretti, S.; Clementi, E.; Giardina, B.; et al. Neuroglobin expression and oxidant/antioxidant balance after graded traumatic brain injury in the rat. Free Radic. Biol. Med. 2014, 69, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, M.I.; Hasan, M.; Islam, M.S.; Sarwar, M.S.; Amin, M.N.; Uddin, S.M.N.; Rahaman, M.Z.; Banik, S.; Hussain, M.S.; Yokota, K.; et al. Elevated serum MDA and depleted non-enzymatic antioxidants, macro-minerals and trace elements are associated with bipolar disorder. J. Trace Elem. Med. Biol. 2017, 39, 162–168. [Google Scholar] [CrossRef]
- Pfeifer, S.; Goldberg, J.; Lobo, R.; Thomas, M.; Widra, E.; Licht, M.; Collins, J.; Cedars, M.; Vernon, M.; Davis, O.; et al. Definitions of infertility and recurrent pregnancy loss: A committee opinion. Practice Committee of American Society for Reproductive Medicine. Fertil. Steril. 2013, 99. [Google Scholar] [CrossRef]
- Winters, B.R.; Walsh, T.J. The epidemiology of male infertility. Urol. Clin. N. Am. 2014, 41, 195–204. [Google Scholar] [CrossRef]
- Ko, E.Y.; Sabanegh, E.S., Jr.; Agarwal, A. Male infertility testing: Reactive oxygen species and antioxidant capacity. Fertil. Steril. 2014, 102, 1518–1527. [Google Scholar] [CrossRef]
- Szkodziak, P.; Wozniak, S.; Czuczwar, P.; Wozniakowska, E.; Milart, P.; Mroczkowski, A.; Paszkowski, T. Infertility in the light of new scientific reports - focus on male factor. Ann. Agric. Environ. Med. 2016, 23, 227–230. [Google Scholar] [CrossRef]
- Cui, X.; Jing, X.; Wu, X.; Yan, M. Protective effect of resveratrol on spermatozoa function in male infertility induced by excess weight and obesity. Mol. Med. Rep. 2016, 14, 4659–4665. [Google Scholar] [CrossRef] [Green Version]
- Lanzafame, F.M.; La Vignera, S.; Vicari, E.; Calogero, A.E. Oxidative stress and medical antioxidant treatment in male infertility. Reprod. Biomed. Online 2009, 19, 638–659. [Google Scholar] [CrossRef]
- Agarwal, A.; Sharma, R.; Sharma, R.; Assidi, M.; Abuzenadah, A.; Alshahrani, S.; Durairajanayagam, D.; Sabanegh, E. Characterizing semen parameters and their association with reactive oxygen species in infertile men. Reprod. Biol. Endocrinol. 2014, 12. [Google Scholar] [CrossRef]
- Cardoso, J.P.; Cocuzza, M.; Elterman, D. Optimizing male fertility: Oxidative stress and the use of antioxidants. World J. Urol. 2019, 37. [Google Scholar] [CrossRef]
- Aitken, R.J.; Ryan, A.L.; Baker, M.A.; McLaughlin, E.A. Redox activity associated with the maturation and capacitation of mammalian spermatozoa. Free Radic. Biol. Med. 2004, 36, 994–1010. [Google Scholar] [CrossRef]
- Baker, M.A.; Aitken, R.J. The importance of redox regulated pathways in sperm cell biology. Mol. Cell Endocrinol. 2004, 216, 47–54. [Google Scholar] [CrossRef]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S. Effect of oxidative stress on male reproduction. World J. Men’s Health 2014, 32. [Google Scholar] [CrossRef]
- Du Plessis, S.S.; Agarwal, A.; Halabi, J.; Tvrda, E. Contemporary evidence on the physiological role of reactive oxygen species in human sperm function. J. Assist. Reprod. Genet. 2015, 32, 509–520. [Google Scholar] [CrossRef] [Green Version]
- De Lamirande, E.; Jiang, H.; Zini, A.; Kodama, H.; Gagnon, C. Reactive oxygen species and sperm physiology. Rev. Reprod. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Sahnoun, S.; Sellami, A.; Chakroun, N.; Mseddi, M.; Attia, H.; Rebai, T.; Lassoued, S. Human sperm Toll-like receptor 4 (TLR4) mediates acrosome reaction, oxidative stress markers, and sperm parameters in response to bacterial lipopolysaccharide in infertile men. J. Assist. Reprod. Genet. 2017, 34, 1067–1077. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, H.W.; Irvine, D.S. On the nature of semen quality and infertility. Hum. Reprod. 1995, 10, 248–249. [Google Scholar] [CrossRef]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 2016, 28. [Google Scholar] [CrossRef]
- O’Flaherty, C.; Matsushita-Fournier, D. Reactive oxygen species and protein modifications in spermatozoa. Biol. Reprod. 2017, 97, 577–585. [Google Scholar] [CrossRef]
- Bisht, S.; Dada, R. Oxidative stress: Major executioner in disease pathology, role in sperm DNA damage and preventive strategies. Front. Biosci. 2017, 9, 420–447. [Google Scholar]
- Ghyasvand, T.; Goodarzi, M.T.; Amiri, I.; Karimi, J.; Ghorbani, M. Serum levels of lycopene, beta-carotene, and retinol and their correlation with sperm DNA damage in normospermic and infertile men. Int. J. Reprod. Biomed. 2015, 13, 787–792. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010; p. 271. [Google Scholar]
- Tavazzi, B.; Lazzarino, G.; Leone, P.; Amorini, A.M.; Bellia, F.; Janson, C.G.; Di Pietro, V.; Ceccarelli, L.; Donzelli, S.; Francis, J.S.; et al. Simultaneous high-performance liquid chromatographic separation of purines, pyrimidines, N-acetylated amino acids, and dicarboxylic acids for the chemical diagnosis of inborn errors of metabolism. Clin. Biochem. 2005, 38, 997–1008. [Google Scholar] [CrossRef]
- Lazzarino, G.; Longo, S.; Amorini, A.M.; Di Pietro, V.; D’Urso, S.; Lazzarino, G.; Belli, A.; Tavazzi, B. Single-step preparation of selected biological fluids for the high-performance liquid chromatographic analysis of fat-soluble vitamins and antioxidants. J. Chromatogr. A 2017, 1527, 43–52. [Google Scholar] [CrossRef]
- Castiglione, D.; Platania, A.; Conti, A.; Falla, M.; D’Urso, M.; Marranzano, M. Dietary micronutrient and mineral intake in the mediterranean healthy eating, ageing, and lifestyle (MEAL) study. Antioxidants 2018, 7, 79. [Google Scholar] [CrossRef]
- Bürzle, M.; Suzuki, Y.; Ackermann, D.; Miyazaki, H.; Maeda, N.; Clémençon, B.; Burrier, R.; Hediger, M.A. The sodium-dependent ascorbic acid transporter family SLC23. Mol. Asp. Med. 2013, 34, 436–454. [Google Scholar] [CrossRef]
- Savini, I.; Rossi, A.; Pierro, C.; Avigliano, L.; Catani, M.V. SVCT1 and SVCT2: Key proteins for vitamin C uptake. Amino Acids. 2008, 34, 347–355. [Google Scholar] [CrossRef]
- Parker, W.H.; Qu, Z.C.; May, J.M. Ascorbic acid transport in brain microvascular pericytes. Biochem. Biophys. Res. Commun. 2015, 458, 262–267. [Google Scholar] [CrossRef] [Green Version]
- Angulo, C.; Castro, M.A.; Rivas, C.I.; Segretain, D.; Maldonado, R.; Yañez, A.J.; Slebe, J.C.; Vera, J.C.; Concha, I.I. Molecular identification and functional characterization of the vitamin C transporters expressed by Sertoli cells. J. Cell Physiol. 2008, 217, 708–716. [Google Scholar] [CrossRef]
- Colagar, A.H.; Marzony, E.T.; Chaichi, M.J. Zinc levels in seminal plasma are associated with sperm quality in fertile and infertile men. Nutr. Res. 2009, 29, 82–88. [Google Scholar] [CrossRef]
- Jeyendran, R.S.; Van der ven, H.H. Chemical constituents of human seminal plasma: Relationship to fertility. Andrologia 1989, 21, 423–428. [Google Scholar] [CrossRef]
- Luo, P.; Dai, S.; Zeng, J.; Duan, J.; Shi, H.; Wang, J. Inward-facing conformation of l-ascorbate transporter suggests an elevator mechanism. Cell Discov. 2018, 17. [Google Scholar] [CrossRef]
- Kammermeier, H. The immediate environment of cardiomyocytes: Substantial concentration differences between the interstitial fluid and plasma water for substrates and transmitters. J. Mol. Cell. Cardiol. 1995, 27, 195–200. [Google Scholar] [CrossRef]
- Palnitkar, G.; Phillips, C.L.; Hoyos, C.M.; Marren, A.J.; Bowman, M.C.; Yee, B.J. Linking sleep disturbance to idiopathic male infertility. Sleep Med. Rev. 2018, 42, 149–159. [Google Scholar] [CrossRef]
- Adewoyin, M.; Ibrahim, M.; Roszaman, R.; Isa, M.L.M.; Alewi, N.A.M.; Rafa, A.A.A.; Anuar, M.N.N. Male Infertility: The effect of natural antioxidants and phytocompounds on seminal oxidative stress. Diseases 2017, 5, 9. [Google Scholar] [CrossRef]
- Ahmadi, S.; Bashiri, R.; Ghadiri-Anari, A.; Nadjarzadeh, A. Antioxidant supplements and semen parameters: An evidence-based review. Int. J. Reprod. Biomed. 2016, 14, 729–736. [Google Scholar] [CrossRef]
- Safarinejad, M.R.; Safarinejad, S.; Shafiei, N.; Safarinejad, S. Effects of the reduced form of coenzyme Q10 (ubiquinol) on semen parameters in men with idiopathic infertility: A double-blind, placebo controlled, randomized study. J. Urol. 2012, 188, 526–531. [Google Scholar] [CrossRef]
- Pahune, P.P.; Choudhari, A.R.; Muley, P.A. The total antioxidant power of semen and its correlation with the fertility potential of human male subjects. J. Clin. Diagn. Res. 2013, 7, 991–995. [Google Scholar] [CrossRef]
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Baskaran, S.; Dutta, S.; Sengupta, P.; Khorram Khorshid, H.R.; Esteves, S.; Gilany, K.; Hedayati, M.; et al. Reactive oxygen species-induced alterations in H19-Igf2 methylation patterns, seminal plasma metabolites, and semen quality. J. Assist. Reprod. Genet. 2018, 36. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Age (Years) | Sperm Concentration (Million/mL) | Total Sperm Motility (%) | Sperm Morphology (% of Normal Spermatozoa) |
|---|---|---|---|
| 39.12 ± 14.38 a (25–41) | 128.54 ± 56.39 (108–195) | 88.05 ± 10.44 (68–93) | 76.62 ± 10.65 (69–84) |
| Compounds | SP | Serum |
|---|---|---|
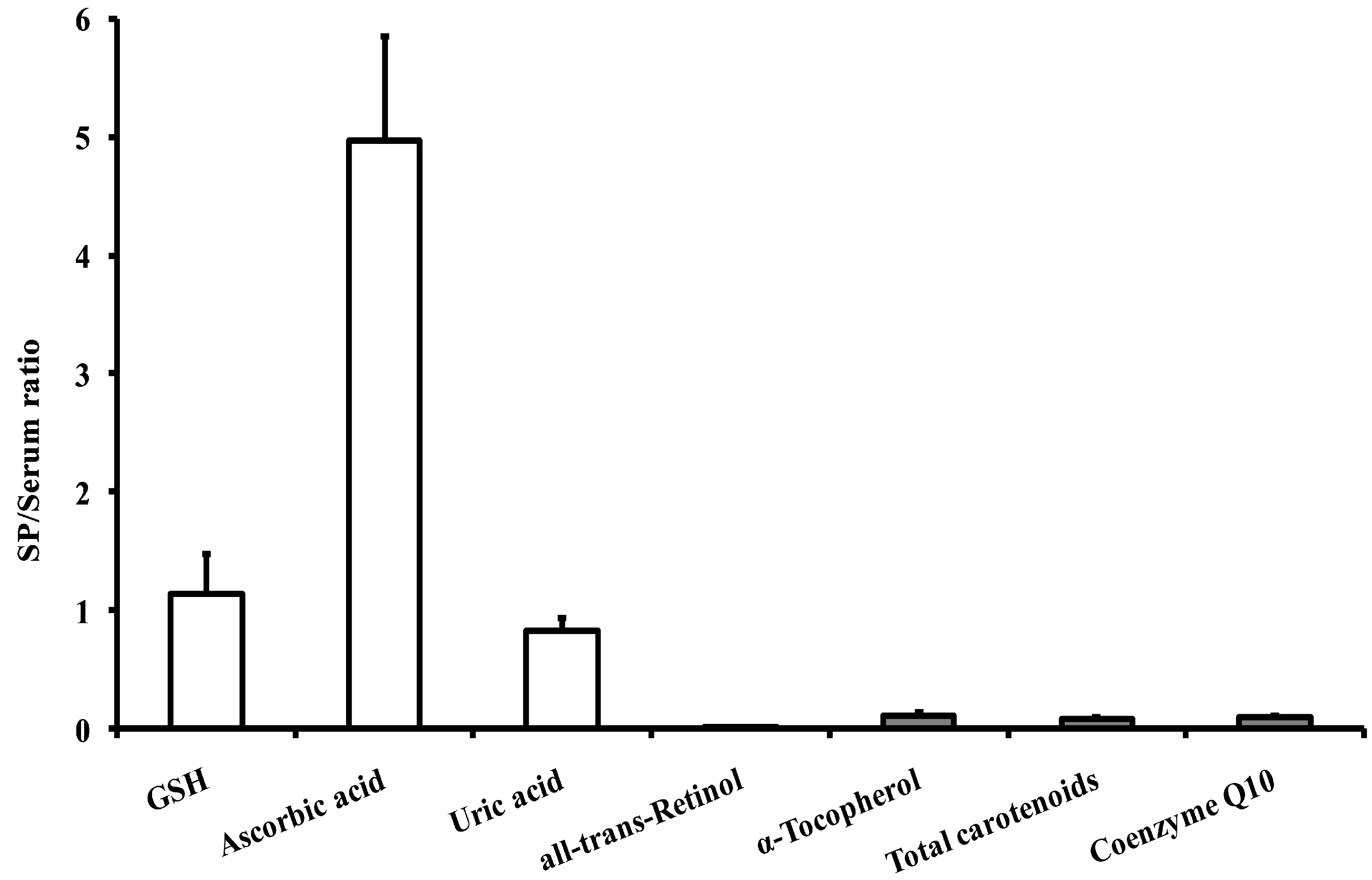
| Glutathione (GSH) | 17.64 ± 4.12 a (13–22) | 15.54 ± 2.66 (12–19) |
| Ascorbic acid | 286.01 ± 75.29 b (205–350) | 57.52 ± 14.81 (38–71) |
| Uric acid | 232.37 ± 44.13 (193–290) | 270.46 ± 57.90 (200–320) |
| Compounds | SP | Serum |
|---|---|---|
| all-trans-retinoic acid | 0.001 ± 0.001 a,b (0–0.002) | 0.006 ± 0.003 (0.004–0.012) |
| all-trans-retinol | 0.068 ± 0.028 b (0.030–0.080) | 5.69 ± 1.89 (4.00–8.50) |
| α-tocopherol | 3.06 ± 0.85 b (2.20–4.10) | 28.51 ± 7.08 (15.50–34.00) |
| γ-tocopherol | 0.066 ± 0.024 b (0.041–0.086) | 1.68 ± 0.76 (0.65–2.20) |
| Astaxhantin | N.D. | 0.004 ± 0.002 (0.003–0.008) |
| Lutein | 0.067 ± 0.032 b (0.036–0.089) | 0.593 ± 0.211 (0.330–0.770) |
| Zeaxhantin | 0.001 ± 0.001 b (0–0.002) | 0.009 ± 0.003 (0.003–0.013) |
| trans-β-apo-8′-carotenal | 0.022 ± 0.005 b (0.017–0.028) | 0.152 ± 0.033 (0.080–0.185) |
| β-cryptoxanthin | 0.003 ± 0.001 b (0.001–0.004) | 0.019 ± 0.005 (0.012–0.029) |
| Lycopene | 0.003 ± 0.001 b (0.001–0.004) | 0.211 ± 0.096 (0.135–0.350) |
| α-carotene | 0.004 ± 0.002 b (0.002–0.007) | 0.035 ± 0.006 (0.028–0.048) |
| β-carotene | 0.008 ± 0.005 b (0.003–0.140) | 0.274 ± 0.099 (0.200–0.435) |
| Total carotenoids | 0.108 ± 0.035 b (0.050–0.190) | 1.30 ± 0.28 (0.98–2.00) |
| Coenzyme Q10 | 0.013 ± 0.006 b (0.008–0.020) | 0.152 ± 0.042 (0.100–0.320) |
| Compounds | SP | Serum |
|---|---|---|
| Malondialdehyde (MDA) | 0.005 ± 0.005 a,b (0.001–0.012) | 0.015 ± 0.008 (0.005–0.022) |
| Nitrites (–NO2−) | 2.76 ± 0.85 b (1.50–5.25) | 4.19 ± 1.61 (2.20–7.80) |
| Nitrates (–NO3−) | 22.63 ± 13.55 b (12.00–46.00) | 42.34 ± 12.88 (24.00–76.00) |
| 8-hydroxy-2′-deoxyguanosine (8-OHdG) | N.D. | N.D. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazzarino, G.; Listorti, I.; Bilotta, G.; Capozzolo, T.; Amorini, A.M.; Longo, S.; Caruso, G.; Lazzarino, G.; Tavazzi, B.; Bilotta, P. Water- and Fat-Soluble Antioxidants in Human Seminal Plasma and Serum of Fertile Males. Antioxidants 2019, 8, 96. https://doi.org/10.3390/antiox8040096
Lazzarino G, Listorti I, Bilotta G, Capozzolo T, Amorini AM, Longo S, Caruso G, Lazzarino G, Tavazzi B, Bilotta P. Water- and Fat-Soluble Antioxidants in Human Seminal Plasma and Serum of Fertile Males. Antioxidants. 2019; 8(4):96. https://doi.org/10.3390/antiox8040096
Chicago/Turabian StyleLazzarino, Giacomo, Ilaria Listorti, Gabriele Bilotta, Talia Capozzolo, Angela Maria Amorini, Salvatore Longo, Giuseppe Caruso, Giuseppe Lazzarino, Barbara Tavazzi, and Pasquale Bilotta. 2019. "Water- and Fat-Soluble Antioxidants in Human Seminal Plasma and Serum of Fertile Males" Antioxidants 8, no. 4: 96. https://doi.org/10.3390/antiox8040096