Amelioration of the Oxidative Stress Generated by Simple or Combined Abiotic Stress through the K+ and Ca2+ Supplementation in Tomato Plants

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preliminary Experiments on K+ and Ca2+ Doses in the Nutrient Solution
2.2. Experimental Design, Plant Material and Growth Conditions
2.3. Plant Sampling
2.4. Ion Analysis and Quantification
2.5. Leaf Gas Exchange
2.6. H2O2 Quantification
2.7. Lipid Peroxidation
2.8. RNA Extraction and qRT-PCR Experiments
2.9. Enzymatic Activities
2.10. Statistical Analysis
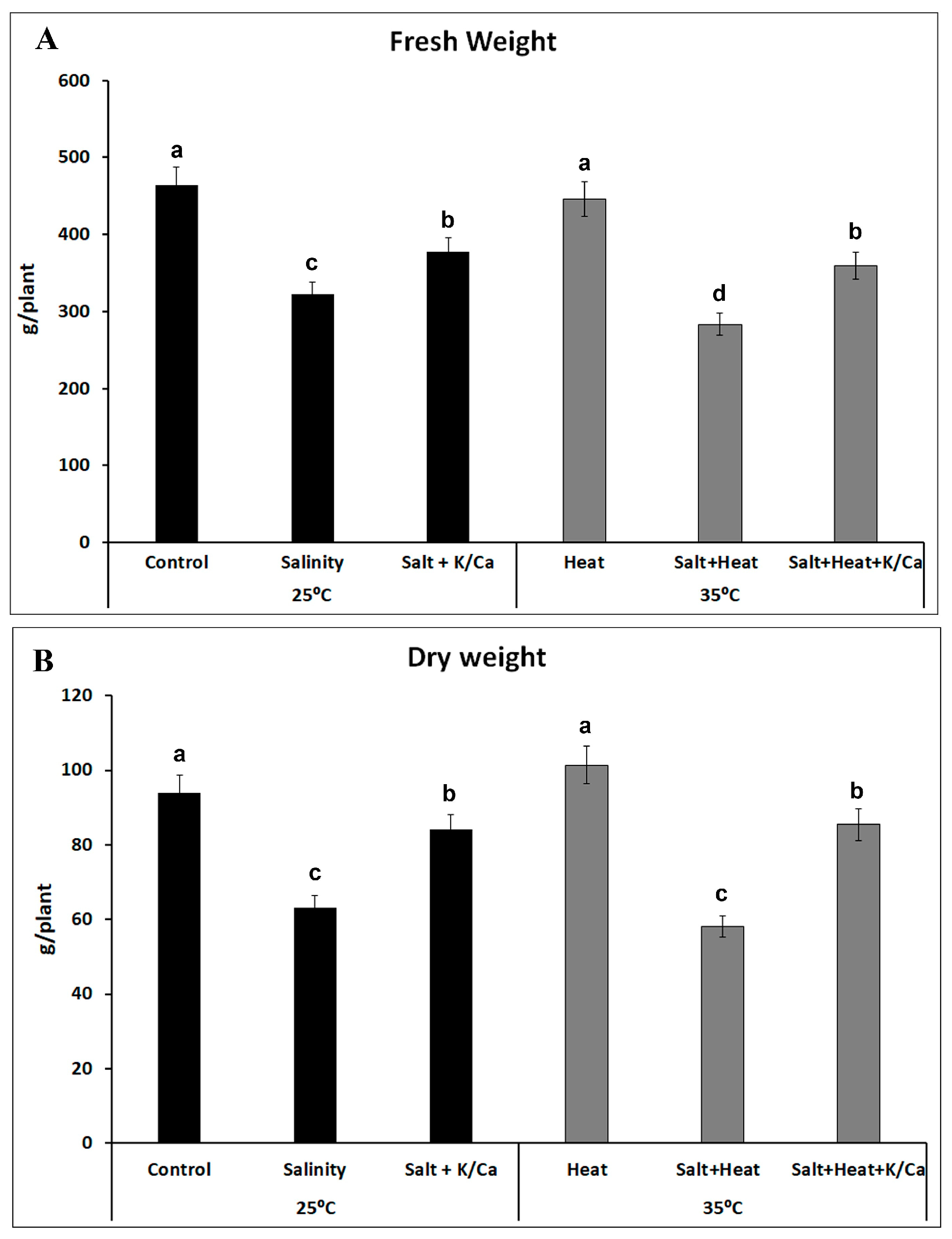
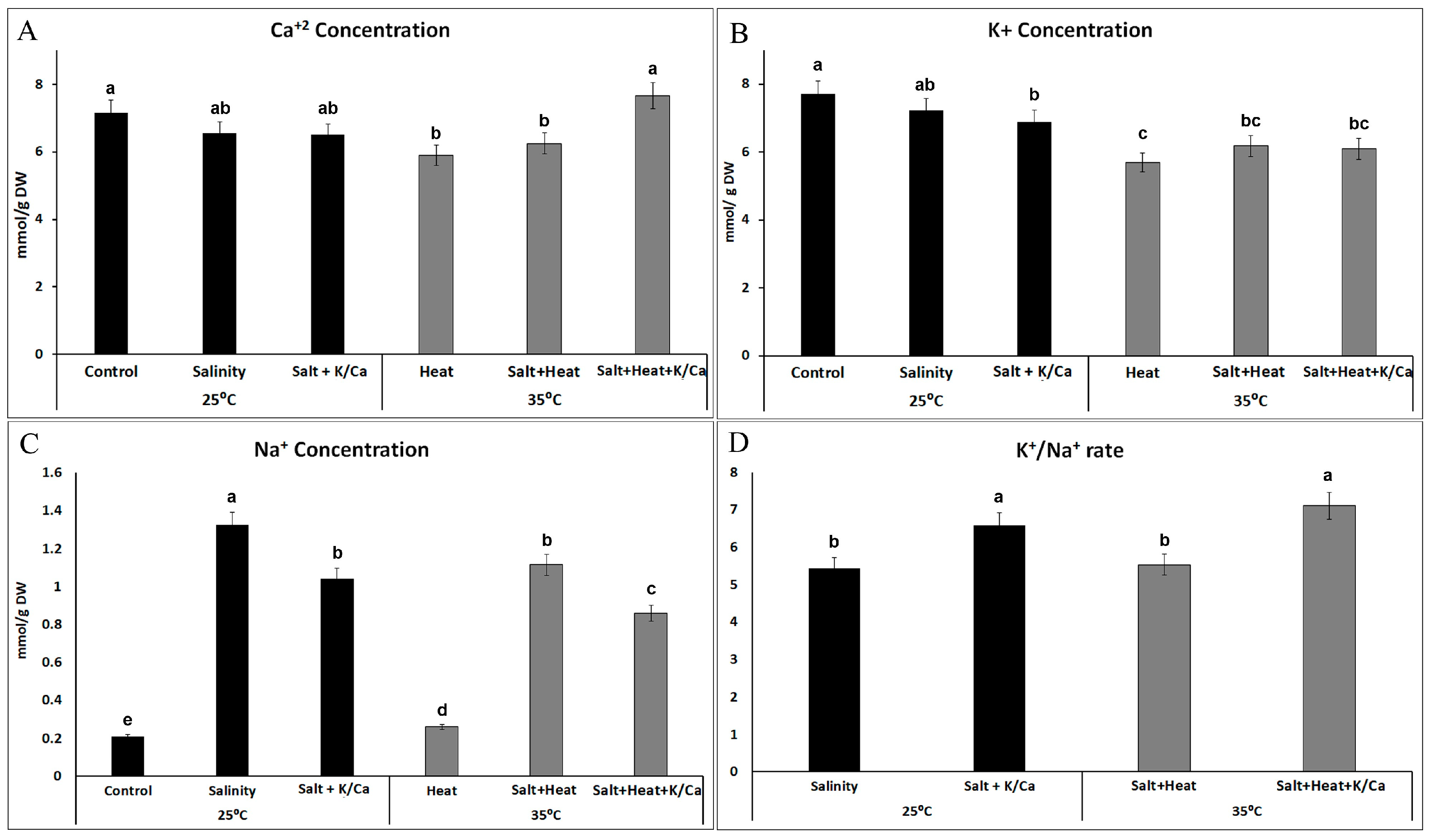
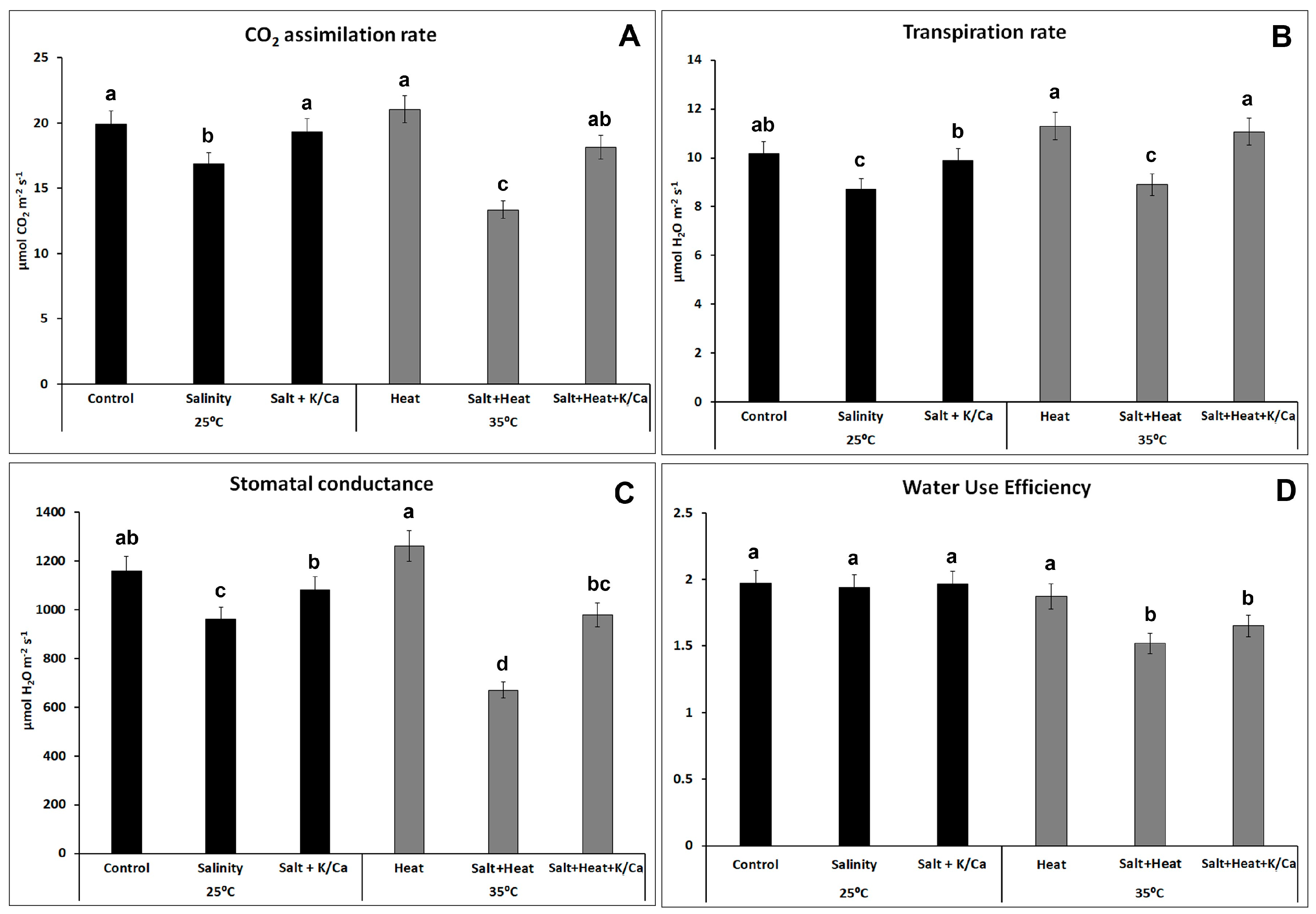
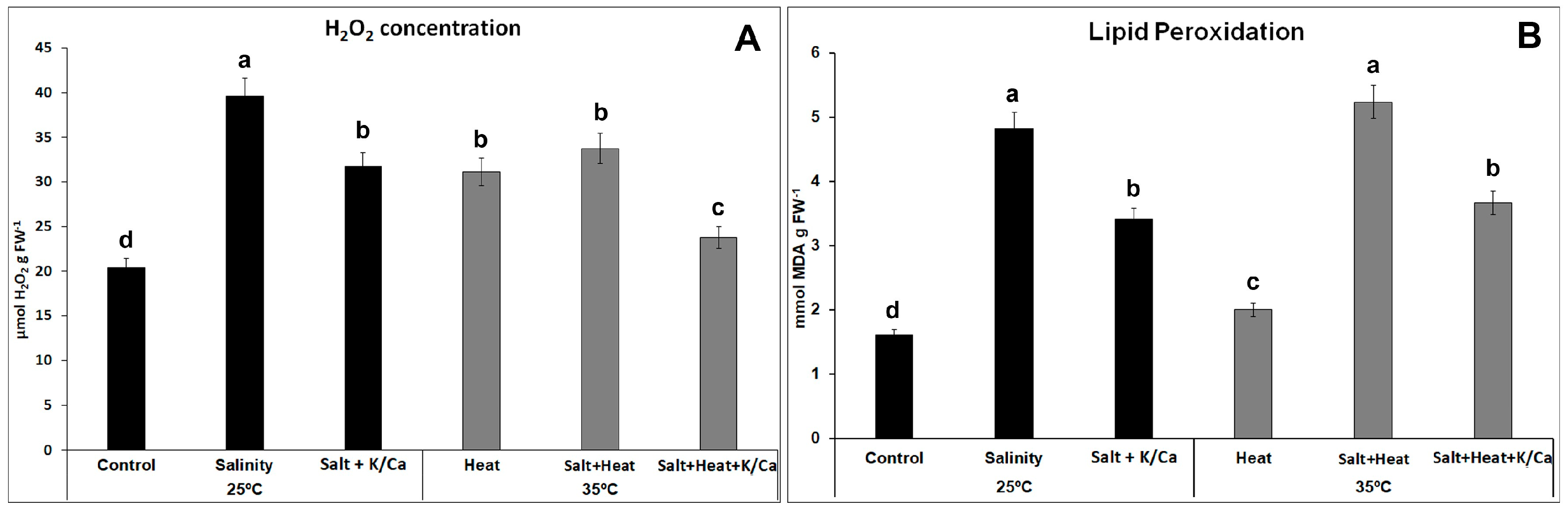
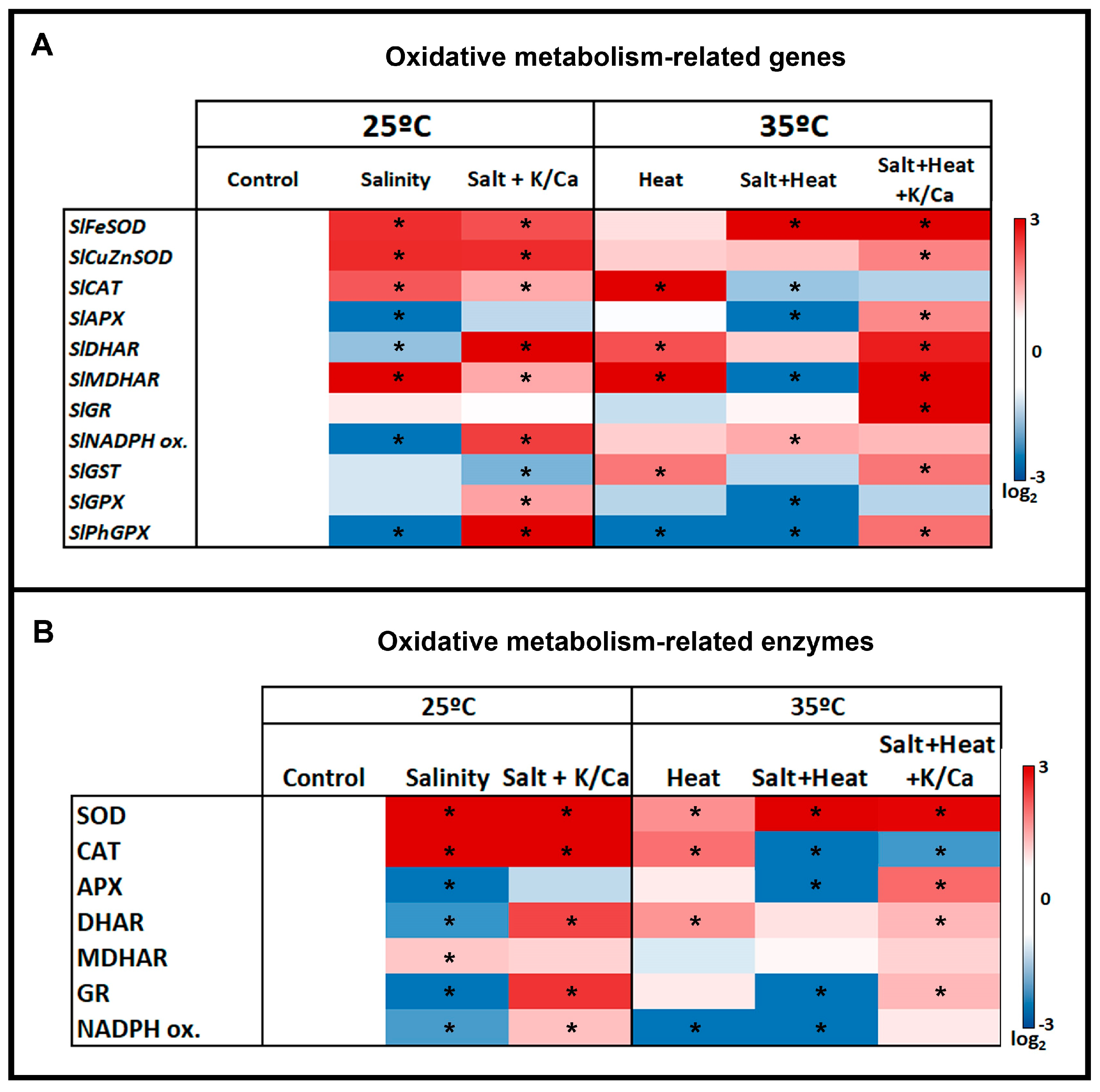
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2015: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2015. [Google Scholar]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Prasch, C.M.; Sonnewald, U. Signaling events in plants: Stress factors in combination change the picture. Environ. Exp. Bot. 2015, 114, 4–14. [Google Scholar] [CrossRef]
- Rivero, R.; Ruiz, J.; Romero, L. Oxidative metabolism in tomato plants subjected to heat stress. J. Hortic. Sci. Biotechnol. 2004, 79, 560–564. [Google Scholar] [CrossRef]
- Martinez, V.; Rubio, F.; Girones-Vilaplana, A.; Mittler, R.; Mestre, T.C.; Moreno, D.A.; Rivero, R.M. Accumulation of Flavonols over Hydroxycinnamic Acids Favors Oxidative Damage Protection under Abiotic Stress. Front. Plant Sci. 2016, 7, 838. [Google Scholar] [CrossRef]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; García-Sánchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to Stress Combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [PubMed]
- Rivero, R.M.; Mestre, T.C.; Mittler, R.; Rubio, F.; Garcia-Sanchez, F.; Martinez, V. The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ. 2014, 37, 1059–1073. [Google Scholar] [CrossRef]
- Kaur, H.; Sirhindi, G.; Bhardwaj, R.; Alyemeni, M.N.; Siddique, K.H.M.; Ahmad, P. 28-homobrassinolide regulates antioxidant enzyme activities and gene expression in response to salt- and temperature-induced oxidative stress in Brassica juncea. Sci. Rep. 2018, 8, 8735. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.L.; Yin, X.H.; Ma, H.C. Effects of hydrogen peroxide on seed germination, seedling growth and physiological characteristcs of bombax ceiba after heat shock Pakistan. J. Bot. 2018, 50, 1327–1333. [Google Scholar]
- Ahmad, P.; Alyemeni, M.N.; Ahanger, M.A.; Wijaya, L.; Alam, P.; Kumar, A.; Ashraf, M. Upregulation of antioxidant and glyoxalase systems mitigates NaCl stress in Brassica juncea by supplementation of zinc and calcium. J. Interact. 2018, 13, 151–162. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; López-Delacalle, M.; Ródenas, R.; Martínez, V.; Rubio, F.; Rivero, R.M. Critical responses to nutrient deprivation: A comprehensive review on the role of ROS and RNS. Environ. Exp. Bot. 2018. [Google Scholar] [CrossRef]
- Qu, C.; Liu, C.; Gong, X.; Li, C.; Hong, M.; Wang, L.; Hong, F. Impairment of maize seedling photosynthesis caused by a combination of potassium deficiency and salt stress. Environ. Exp. Bot. 2012, 75, 134–141. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.L.; Chao, L.; Zhou, M.; Hong, M.M.; Luo, L.Y.; Wang, L.; Ying, W.; Cai, J.W.; Gong, S.J.; Hong, F.S. Oxidative damages of maize seedlings caused by exposure to a combination of potassium deficiency and salt stress. Plant Soil 2011, 340, 443–452. [Google Scholar] [CrossRef]
- Martinez, V.; Lauchli, A. Effects of ca2+ on the salt-stress response of barley roots as observed by in-vivo 31p-nuclear magnetic resonance and in-vitro analysis. Planta 1993, 190, 519–524. [Google Scholar] [CrossRef]
- Reid, R.J.; Smith, F.A. The limits of sodium/calcium interactions in plant growth. Funct. Boil. 2000, 27, 709. [Google Scholar] [CrossRef]
- Bacha, H.; Rodenas, R.; Lopez-Gomez, E.; Francisco Garcia-Legaz, M.; Nieves-Cordones, M.; Rivero, R.M.; Martinez, V.; Angeles Botella, M.; Rubio, F. High ca2+ reverts the repression of high-affinity k+ uptake produced by Na+ in Solanum lycopersycum L. (var. Microtom) plants. J. Plant Physiol. 2015, 180, 72–79. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Shin, R.; Berg, R.H. Reactive Oxygen Species and Root Hairs in Arabidopsis Root Response to Nitrogen, Phosphorus and Potassium Deficiency. Cell Physiol. 2005, 46, 1350–1357. [Google Scholar]
- Hernández, M.; Fernandez-García, N.; Garcia-Garma, J.; Rubio-Asensio, J.; Rubio, F.; Olmos, E. Potassium starvation induces oxidative stress in Solanum lycopersicum L. roots. J. Physiol. 2012, 169, 1366–1374. [Google Scholar] [CrossRef]
- Zhou, R.; Kong, L.; Wu, Z.; Rosenqvist, E.; Wang, Y.; Zhao, L.; Zhao, T.; Ottosen, C.-O. Physiological response of tomatoes at drought, heat and their combination followed by recovery. Physiol. Plant. 2018, 165, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S.I.; Mittler, R.; Balfagon, D.; Arbona, V.; Gomez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 2018, 162, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Aleman, F.; Caballero, F.; Rodenas, R.; Rivero, R.M.; Martinez, V.; Rubio, F. The f130s point mutation in the arabidopsis high-affinity k transporter athak5 increases K+ over Na+ and Cs+ selectivity and confers na+ and cs+ tolerance to yeast under heterologous expression. Front. Plant Sci. 2014, 5, 11. [Google Scholar] [CrossRef]
- Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Martinez, V.; Rivero, R.M. Glutathione homeostasis as an important and novel factor controlling blossom-end rot development in calcium-deficient tomato fruits. J. Physiol. 2012, 169, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Ortega, W.; Martinez, V.; Rivero, R.; Cámara-Zapata, J.-M.; Mestre, T.; Garcia-Sanchez, F. Use of a smart irrigation system to study the effects of irrigation management on the agronomic and physiological responses of tomato plants grown under different temperatures regimes. Agric. Manag. 2017, 183, 158–168. [Google Scholar] [CrossRef]
- Macnevin, W.M.; Urone, P.F. Separation of hydrogen peroxide from organic hydroperoxides-aplication to polarographic analysis of mixtures. Anal. Chem. 1953, 25, 1760–1761. [Google Scholar] [CrossRef]
- Brennan, T.; Frenkel, C. Involvement of hydrogen-peroxide in regulation of senescence in pear. Plant Physiol. 1977, 59, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sakakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc. Natl. Acad. Sci. USA 2007, 104, 19631–19636. [Google Scholar] [CrossRef]
- Fu, J.; Huang, B. Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environ. Exp. Bot. 2001, 45, 105–114. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Aebi, H. Catalase invitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Mccord, J.M.; Fridovich, I. The utility of superoxide dismutase in studying free radical reactions. I. Radicals generated by the interaction of sulfite, dimethyl sulfoxide, and oxygen. J. Boil. Chem. 1969, 244, 6056–6063. [Google Scholar]
- Spitz, D.R.; Oberley, L.W. An assay for superoxide dismutase activity in mammalian tissue homogenates. Anal. Biochem. 1989, 179, 8–18. [Google Scholar] [CrossRef]
- Miyake, C.; Asada, K. Thylakoid-Bound Ascorbate Peroxidase in Spinach Chloroplasts and Photoreduction of Its Primary Oxidation Product Monodehydroascorbate Radicals in Thylakoids. Cell Physiol. 1992, 33, 541–553. [Google Scholar]
- Kaundal, A.; Rojas, C.; Mysore, K. Measurement of NADPH Oxidase Activity in Plants. Bio-Protocol 2012, 2, 278. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Involvement of plasma-membrane NADPH oxidase in abscisic acid- and water stress-induced antioxidant defense in leaves of maize seedlings. Planta 2002, 215, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Capula-Rodríguez, R.; Valdez-Aguilar, L.A.; Cartmill, D.L.; Cartmill, A.D.; Alia-Tejacal, I. Supplementary Calcium and Potassium Improve the Response of Tomato (Solanum lycopersicum L.) to Simultaneous Alkalinity, Salinity and Boron Stress. Commun. Soil Sci. Anal. 2016, 47, 505–511. [Google Scholar]
- Navarro, J.M.; Garrido, C.; Martinez, V.; Carvajal, M. Water relations and xylem transport of nutrients in pepper plants grown under two different salts stress regimes. Plant Growth Regul. 2003, 41, 237–245. [Google Scholar] [CrossRef]
- Munns, R.; Shabala, S. Salinity stress: Physiological constraints and adaptive mechanisms. In Plant Stress Physiology; CABI Publishing: Wallingford, CT, USA, 2017; pp. 24–63. [Google Scholar]
- Chakraborty, K.; Basak, N.; Bhaduri, D.; Ray, S.; Vijayan, J.; Chattopadhyay, K.; Sarkar, R.K. Ionic Basis of Salt Tolerance in Plants: Nutrient Homeostasis and Oxidative Stress Tolerance. In Plant Nutrients and Abiotic Stress Tolerance; Springer Nature: Singapore, 2018; pp. 325–362. [Google Scholar]
- Abbasi, G.H.; Akhtar, J.; Ahmad, R.; Jamil, M.; Anwar-Ul-Haq, M.; Ali, S.; Ijaz, M. Potassium application mitigates salt stress differentially at different growth stages in tolerant and sensitive maize hybrids. Plant Growth Regul. 2015, 76, 111–125. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Boil. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Tomar, N.S.; Tittal, M.; Argal, S.; Agarwal, R.M. Plant growth under water/salt stress: ROS production; antioxidants and significance of added potassium under such conditions. Physiol. Mol. Boil. Plants 2017, 23, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Pinero, M.C.; Houdusse, F.; Garcia-Mina, J.M.; Garnica, M.; del Amor, F.M. Regulation of hormonal responses of sweet pepper as affected by salinity and elevated co2 concentration. Physiol. Plant. 2014, 151, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Bohra, J.S.; Doerffling, K. Potassium nutrition of rice (Oryza sativa L.) varieties under NaCl salinity. Plant Soil 1993, 152, 299–303. [Google Scholar] [CrossRef]
- Malińska, D.; Mirandola, S.R.; Kunz, W.S. Mitochondrial potassium channels and reactive oxygen species. FEBS Lett. 2010, 584, 2043–2048. [Google Scholar] [CrossRef] [PubMed]
- Trono, D.; Laus, M.N.; Soccio, M.; Alfarano, M.; Pastore, D. Modulation of potassium channel activity in the balance of ros and atp production by durum wheat mitocondria—An amazing defense tool against hyperosmotic stress. Front. Plant Sci. 2015, 6, 1072. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S. Calcium-dependent signaling pathway in the heat-induced oxidative injury in Amaranthus lividus. Boil. Plant. 2008, 52, 137–140. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Modulation of Antioxidant Defense System Is Associated with Combined Drought and Heat Stress Tolerance in Citrus. Front. Sci. 2017, 8, 10. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Alamri, S.A.; Al-Khaishany, M.Y.; Al-Qutami, M.A.; Ali, H.M.; Khan, M.N. Sodium nitroprusside and indole acetic acid improve the tolerance of tomato plants to heat stress by protecting against DNA damage. J. Interact. 2017, 12, 177–186. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Chamber | Treatments | [NaCl] | [K+] | [Ca2+] |
|---|---|---|---|---|
| Chamber A (25 °C) | Control | 0 mM | 7 mM | 4 mM |
| Salinity | 60 mM | 7 mM | 4 mM | |
| Salinity + T1 | 60 mM | 8 mM | 4.7 mM | |
| Salinity + T2 | 60 mM | 9.8 mM | 5.6 mM | |
| Salinity + T3 | 60 mM | 11 mM | 6.5 mM | |
| Chamber B (35 °C) | Heat | 0 mM | 7 mM | 4 mM |
| Salinity + Heat | 60 mM | 7 mM | 4 mM | |
| Salinity + Heat + T1 | 60 mM | 8 mM | 4.7 mM | |
| Salinity + Heat + T2 | 60 mM | 9.8 mM | 5.6 mM | |
| Salinity + Heat + T3 | 60 mM | 11 mM | 6.5 mM |
| Greenhouse | Treatments | [NaCl] | [K+] | [Ca2+] |
|---|---|---|---|---|
| Greenhouse A (25 °C) | Control | 0 mM | 7 mM | 4 mM |
| Salinity | 60 mM | 7 mM | 4 mM | |
| Salinity + K/Ca | 60 mM | 9.8 mM | 5.6 mM | |
| Greenhouse B (35 °C) | Heat | 0 mM | 7 mM | 4 mM |
| Salinity + Heat | 60 mM | 7 mM | 4 mM | |
| Salinity + Heat + K/Ca | 60 mM | 9.8 mM | 5.6 mM |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Martí, M.; Piñero, M.C.; García-Sanchez, F.; Mestre, T.C.; López-Delacalle, M.; Martínez, V.; Rivero, R.M. Amelioration of the Oxidative Stress Generated by Simple or Combined Abiotic Stress through the K+ and Ca2+ Supplementation in Tomato Plants. Antioxidants 2019, 8, 81. https://doi.org/10.3390/antiox8040081
García-Martí M, Piñero MC, García-Sanchez F, Mestre TC, López-Delacalle M, Martínez V, Rivero RM. Amelioration of the Oxidative Stress Generated by Simple or Combined Abiotic Stress through the K+ and Ca2+ Supplementation in Tomato Plants. Antioxidants. 2019; 8(4):81. https://doi.org/10.3390/antiox8040081
Chicago/Turabian StyleGarcía-Martí, María, María Carmen Piñero, Francisco García-Sanchez, Teresa C. Mestre, María López-Delacalle, Vicente Martínez, and Rosa M. Rivero. 2019. "Amelioration of the Oxidative Stress Generated by Simple or Combined Abiotic Stress through the K+ and Ca2+ Supplementation in Tomato Plants" Antioxidants 8, no. 4: 81. https://doi.org/10.3390/antiox8040081