1. Introduction
Reactive oxygen species (ROS) are messenger molecules that play a prominent role in cell metabolism-regulating cell proliferation, differentiation and death [
1]. Among them, hydrogen peroxide (H
2O
2) is one of the most intriguing ROS. It reacts slowly with most molecules, however, possesses a high affinity to cysteine residues surrounded by particular sequences of amino acids. This explains the selectivity of H
2O
2 for thiolates moieties in active sites of various enzymes [
2]. H
2O
2 is produced by spontaneous and enzyme-mediated dismutation of superoxide (O
2•
−) and is mostly formed in the sites of O
2•
− production. Two well-known sources of O
2•
− in the living cell are the mitochondria electron transport chain (ETC) and the nicotinamide adenine dinucleotide 2′-phosphate (NADPH) oxidases. O
2•
− represents a side-product of ETC activity where it forms due to electron “escape” and reduction of dioxygen (O
2) present at high concentration in the mitochondrial membrane [
3]. O
2•
− formation in mitochondria is a hallmark of mitochondrial activity [
4]. In contrast with ETC, NADPH oxidases produce ROS as their main product. Seven members of NADPH oxidase family are actually described: NOX1-5, Duox1 and Duox2. All seven isoforms are flavocytochromes anchored to the membrane through six transmembrane helices binding two haem cofactors (N-terminal domain). The C-terminal dehydrogenase domain binds flavin adenine dinucleotide (FAD) and NADPH and allows electron transfer to the haem and further across the membrane to molecular oxygen [
5]. Interestingly, while NOX1, NOX2 and NOX3 are known to produce O
2•
−, the other three members of the NADPH oxidase family produce H
2O
2 as the main product. For instance, NOX4 produces 90% of H
2O
2 and 10% of O
2•
− by the currently accepted ‘two sequential single-electron reductions’ mechanism [
5].
NADPH oxidases have been extensively studied due to their involvement in a number of human pathologies, one of which is psoriasis vulgaris. Psoriasis is a dermatologic condition characterized by hyper-proliferation and altered keratinocytes differentiation that leads to the formation of psoriatic plaques—red scaly patches on the skin surface. In the sites of psoriatic lesions, the skin architecture is altered and characterized by the formation of pronounced epidermal protrusions into dermis greatly increasing the area of contact between epidermis and dermis. These morphological changes are associated with the notable redox imbalance. Our previous studies showed an increased intracellular ROS production and an enhanced NADPH oxidase activity in fibroblasts obtained from lesional skin of psoriatic patients [
6]. This data is in line with the findings of other authors who showed O
2•
− production, mitochondrial superoxide dismutase activity (Mn-SOD) and protein carbonylation level [
7,
8] to be significantly increased in fibroblasts obtained from involved and uninvolved skin of psoriatic patients in comparison to healthy skin fibroblasts.
In the present study, an in-depth investigation on redox status in fibroblasts obtained from lesional (“LES”, the area of psoriatic plaque) and non-lesional (“nLES”, visibly unaffected) skin of psoriatic patients with respect to primary control (CTR) fibroblasts obtained from healthy donors was performed. In particular, oxidative stress markers and NADPH oxidase activity in nLES and LES fibroblasts were investigated. Furthermore, in order to identify possible ROS sources in psoriatic fibroblasts, three members of the NADPH oxidase family were studied. Co-culture experiments of fibroblasts with healthy primary human keratinocytes were also performed in order to assess the possible effects of fibroblast-derived ROS on the proliferation rate and redox balance of keratinocytes. Finally, the involvement of stress-sensitive molecular pathways was investigated in psoriatic versus CTR fibroblasts and in keratinocytes co-cultured with the fibroblasts.
2. Materials and Methods
2.1. Reagents and Antibodies
The information of all reagents and antibodies used in the study is summarized in
Table 1 and
Table 2.
2.2. Patients
Five patients affected by plaque psoriasis and four healthy controls matched by age, and body mass index (BMI) were enrolled in this study approved by the Ethics Committee of Azienda Ospedaliero-Universitaria Careggi (N10709_bio from 27.03.2017) and conducted according to the Helsinki declaration. The demographic and clinical information for each patient and healthy subject is summarized in
Table 3. The patients did not comply with the pruritus at the site of the lesions chosen for the biopsy. No subjects involved in the study followed any systemic therapy before the study or had a history of any disease, e.g., diabetes mellitus and atherosclerosis, which might affect the redox status.
2.3. Primary Fibroblasts and Keratinocytes Isolation and Setting Up of Cell Cultures
Fibroblasts isolation was performed as previously reported [
6,
9,
10,
11]. Briefly, 5 mm punch-biopsies were obtained from LES and nLES skin of psoriatic patients and from the skin of healthy controls (CTR). With nLES skin, we intend to use the perilesional symptom-free area of the skin. Immediately after, biopsies were subjected to the procedure of epidermal-dermal separation in 0.5% dispase II solution in DMEM containing antibiotics and antimycotics overnight at 4 °C. As follows, epidermis and dermis were easily detached with the forceps and washed with ice-cold PBS. In order to establish the primary fibroblasts, dermis was cut in small pieces, dried for 5–10 min on-air and covered by the cover-glass performing a moderate pressure. A complete DMEM (20% FBS, 1 g/L glucose, 100 units/mL of penicillin, 100 µg/mL of streptomycin and 0,25 µg/mL of Amphotericin B) was added to the dish and changed every two days. After 10–12 days the semi-confluent layer of fibroblasts was observed in the dish. At this point, the dermal pieces were transferred to a new dish where the process of fibroblasts migration continued. The semi-confluent fibroblasts were trypsinized and cultivated in DMEM (10% FBS, 1 g/L glucose, 100 units/mL of penicillin, 100 µg/mL of streptomycin and 0,25 µg/mL of Amphotericin B) at 37 °C. The cells were frozen or used in experiments at their passages 2 to 5.
In order to establish the primary keratinocyte culture from control skin, the epidermis was chopped with a sterile scalpel into fine segments immediately after the separation from the dermis. Afterward, the epidermal slices were incubated in 0.25% Trypsin-EDTA solution (porcine trypsin) at 37 °C with frequent pipetting until the epidermal slice started to disaggregate into single cells. At this point, the pre-heated PBS was added to the dish and the cells were filtered through a cell strainer filter with 70 μm pores to obtain a single-cell suspension, which was further centrifuged for 5 min at 1000 rpm, resuspended in PBS and centrifuged again. The pellet was resuspended in KGM medium completed with all SingleQuotsTM Supplements and Growth Factors and 1% antibiotic-antimycotic solution. The anti-vimentin and anti-pan-cytokeratin immunocytochemical staining were further performed for both fibroblasts and keratinocytes cell cultures in order to verify the cell type purity.
Prior to studies, the cell cultures were tested for Mycoplasma infection by DAPI staining and confocal microscope imaging. Moreover, when the primary cultures were used in laboratory, the surfaces of the cell incubator (every week) and cell culture hood (every day) were disinfected with anti-Mycoplasma-EX Spray Surface Disinfectant (PromoCell GmbH, Heidelberg, Germany). All cell cultures used in the present study were negative for Mycoplasma infection.
2.4. Contactless Keratinocytes-Fibroblasts Co-Incubation Protocol
In order to study the effect of ROS-hyper-producing fibroblasts on redox homeostasis of primary keratinocytes (KER) obtained from healthy subjects, KER were co-cultured with CTR, nLES and LES following the contactless protocol (
Figure 1). Briefly, keratinocytes were plated on cover glasses, allowed to grow for 24 h and further put in Petri dish with semi-confluent fibroblasts seeded 24 h before. In this condition, continuous conditioning of KER by the fibroblasts factors is allowed although the two cell types are physically separated. After 24 h of co-incubation, the medium was changed. After 48 h of co-incubation, the KER-cover glasses were extracted from the Petri dishes and the keratinocytes were trypsinized and prepared for further studies.
2.5. Indirect Immunocytochemistry (ICC)
Cells were fixed with 3% paraformaldehyde in PBS for 20 min at room temperature and treated with 0.5% Triton X-100 for 10 min on ice. The cells were thoroughly washed with PBS and incubated with primary antibodies (
Table 2) diluted in PBS for 1 h at room temperature in a humid chamber in the dark. After washing with PBS, the cells were incubated with secondary antibodies (
Table 2) diluted in PBS for 45 min at room temperature, dehydrated in alcohol gradient and embedded in Moviol containing DABCO (1,4-diazobicyclo[2,2,2]octane) (Sigma-Aldrich Italy S.r.l., Milan, Italy). The preparations were studied with Leica TCS SP5 Confocal Microscope (Mannheim, Germany) using 63×/NA 1.25 Plan Neofluar objectives (0.5 μm optical slice thickness, z-sections collected at 0.3 μm intervals). The excitation and emission wavelengths were 488 nm and 495–550 nm, respectively. If not stated alternatively the ICC protocol was the same for all experiments. The ICC staining was performed at the passages 0 and 5 for all psoriatic and control fibroblasts and for control keratinocytes.
2.6. Cell Cycle Progression Analysis
Cells were trypsinized, washed with PBS, resuspended in 0.5 mL of PBS containing 20 mg/mL propidium Iodide and left in dark at +4 °C for 1 h. Cell cycle distribution was analyzed by flow cytometry (FACSCanto, Becton-Dickinson, San Jose, CA, USA) and by FlowJo software (free software, Becton-Dickinson, San Jose, CA, USA). The percentage of cells in senescence/Gap 1 (G0/G1), synthesis (S) and Gap 2/mitotic (G2/M) phases were determined for the control and experimental conditions and averaged across triplicate experiments.
2.7. Assessment of Oxidative Stress Markers in Living Fibroblasts by Confocal Microscopy
CTR, nLES and LES fibroblasts were grown in glass-bottom Petri dishes (Nunc, Roskilde, Denmark) for confocal studies. At semi-confluence the DMEM was replaced with pre-heated serum-free and phenol red-free RPMI medium containing fluorescent probes H2DCF-DA (2.5 µM), DAF-FM (2.5 µM), Fluo-3 (2.5 µM), or MitoSOX (3 µM) dissolved in 0.1% DMSO and Pluronic acid F-127 (0.01% w/v) in order to visualize the intracellular ROS, nitric oxide (•NO), Ca2+ or mitochondrial O2•− production levels, respectively. After 30 min incubation at 37 °C and three wash cycles with pre-heated PBS the RPMI medium was added and the cells were incubated at 37 °C for another 30 min prior to the confocal imaging. The fluorescence was detected in living cells at excitation and emission wavelengths of 488 and 520 nm, respectively, using a Leica TCS SP5 Confocal Microscope (Mannheim, Germany) equipped with an argon laser source. The observations were performed by collecting the emitted fluorescence with a Leica 40× oil immersion objective.
2.8. Assessment of Intracellular Oxidative Stress Markers and Intracellular Ca2+ Levels in Living Fibroblasts and Keratinocytes by FACS Analysis
To determine intracellular ROS, •NO, Ca2+ and mitochondrial O2•− production, fibroblasts/keratinocytes were trypsinized, washed with PBS and resuspended in serum- and phenol red-free RPMI medium containing 1 μM of the following fluorescent probes: H2DCF-DA, DAF- FM DA, Fluo-3 and MitoSOX, respectively. After 30 min of incubation in the dark at 37 °C, cells were washed three times, resuspended in PBS and fluorescence was analyzed by FACSCanto flow cytometer (Becton-Dickinson, San Jose, CA, USA).
2.9. Whole Cell Homogenates Preparation
After trypsinization, fibroblasts (1 × 10
6) were resuspended on ice in 150 µL of lysis buffer (20 mM Tris-HCl pH8, 1% Triton X-100, 10% (
v/v) glycerol, 137 mM NaCl, 2 mM EDTA and 6 M urea supplemented with 0.2 mM PMSF, 10 mg/mL leupeptin + aprotinin). Samples were then subjected to three freeze–thaw cycles and sonicated three times (5 s each) on ice, centrifuged at 14,000×
g for 10 min at 4 °C, and the supernatant was collected. Protein concentration was determined according to the Bradford method [
12]. Samples were stored at –80 °C before use.
2.10. Thiobarbituric Acid Reactive Substance (TBARS) Evaluation
Malondialdehyde (MDA) is the final product of fatty acid peroxidation. MDA levels in CTR, nLES and LES fibroblasts homogenates were quantified using TBARS assay kit (Oxitek-ZeptoMetrix Corporation Buffalo, NY, USA) following the manufacturer protocol. An amount of 5 µL of whole-cell homogenates was used per sample. The fluorescence emission of the recovered supernatant was measured with an excitation wavelength of 530 nm and an emission wavelength of 550 nm, using a Perkin-Elmer LS55 spectrofluorimeter (Waltham, MA, USA).
2.11. Oxygen Radical Antioxidant Capacity (ORAC) Assay
ORAC assay was performed on homogenates of CTR, nLES and LES fibroblasts. The reaction was carried out in 96-well black microplates (Nunc, Roskilde, Denmark) and Trolox (10–200 μM) was used as standard. The amount of sample per well was calculated in consideration of 4 μg of protein/well. Final assay mixture of the total volume (200 μL) contained: 70 μL of sample diluted in 75 mM phosphate buffer (pH 7) and 100 μL of reagent fluorescein at 6 nM final concentration. After 10 min of incubation in the dark at 37 °C, 30 μL of pre-heated at 37 °C AAPH solution (final concentration of AAPH—200 mM) were added to each well using multiwall channel pipette. The fluorescence was recorded using a fluorometric microplate reader Fluoroskan Ascent (Thermo Electron Corp., Vantaa, Finland) at 5 min intervals for 2 h at excitation and emission wavelengths of 485 and 537 nm, respectively. All assays were conducted in triplicates and at least two independent tests were done for each sample. The area under curve (AUC) was calculated for each sample by integrating the relative fluorescence curve. Regression equations obtained from the net value of Trolox was used to calculate the ORAC value for each assay. Final ORAC values were expressed as μmol of Trolox equivalent (TE) per mg of protein (μmol TE/mg).
2.12. NADPH Oxidase Activity by Luminometric Assay
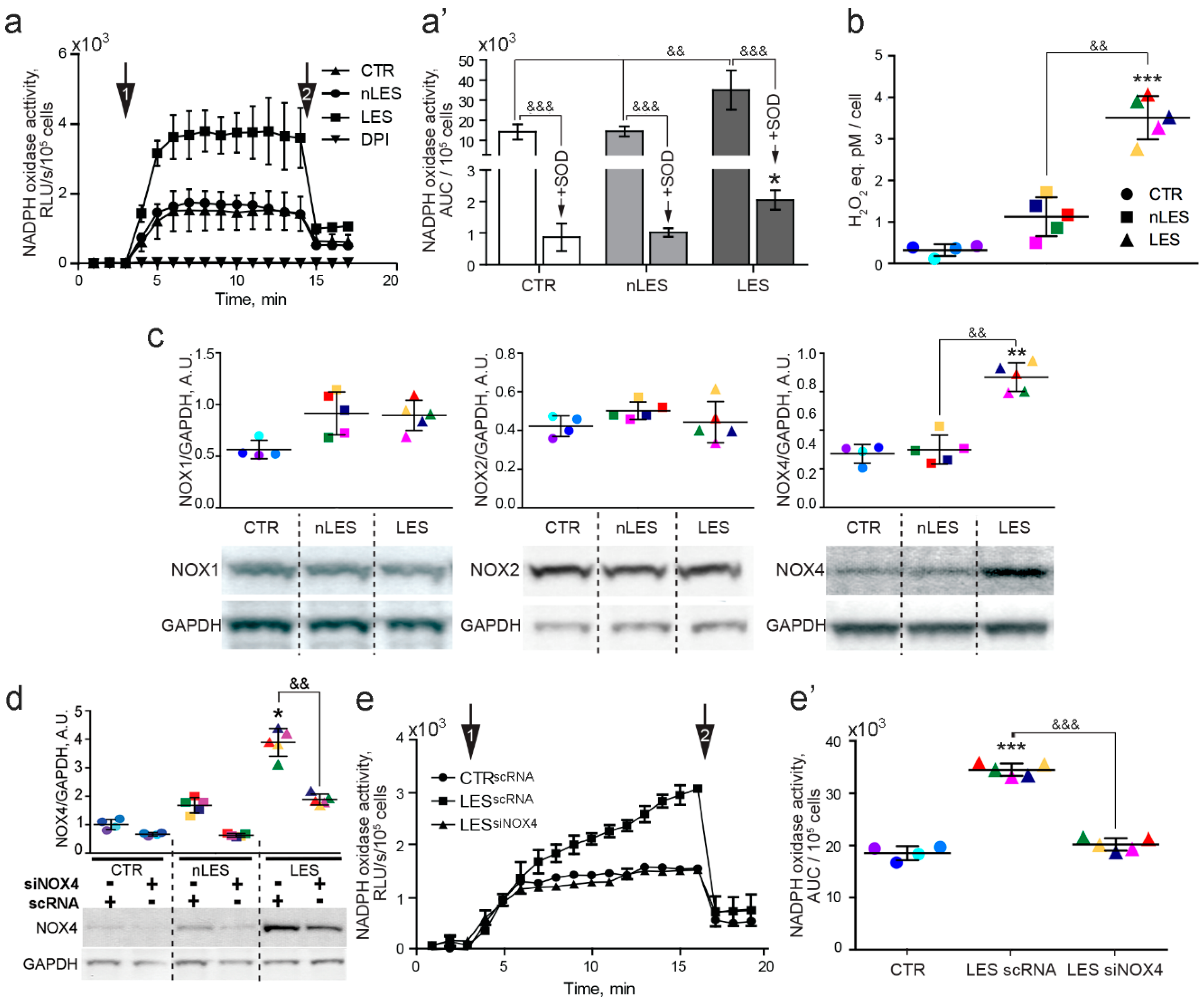
In order to measure the extracellular ROS production by CTR, nLES and LES fibroblasts, the cells were trypsinized, washed with PBS and 3 × 105 of cells per sample were resuspended in 125 μL of Krebs-HEPES buffer (99 mM NaCl, 4.7 mM KCl, 1.2 mM MgSO4, 1 mM KH2PO4, 1.9 mM CaCl2, 25 mM NaHCO3, 20 mM HEPES, and 11.1 mM glucose pH 7.44) and placed in tubes for luminometric assay. After 5 min of incubation at 37 °C, lucigenin (25 μM) was added to the sample. As soon as luminescence level stabilized (in 10 min), the blank value of luminescence was registered by Lumat LB 9507 single-tube luminometer (Berthold Technologies GmbH & Co. KG, Bad Wildbad, Germany). NADPH was then added to the sample at the final concentration of 500 μM and luminescence was registered with 1 min interval for 20 min. Between readings, the cells were maintained at 37 °C. NADPH-stimulated extracellular ROS production was completely abated in fibroblasts pre-incubated for 30 min with the flavoenzyme inhibitor diphenyleneiodonium chloride (DPI, 20 μM) confirming that NADPH oxidase is the source of extracellular ROS production in living fibroblasts. When the curve of NADPH oxidase activity reached the plateau, the SOD (final concentration 450 U/mL) was added to the sample to monitor the O2•− to H2O2 extracellular production ratio. NADPH oxidase activity was represented as RLU/s/cell versus Time (min) and AUC value was calculated. At least ten independent experiments were performed for each condition.
2.13. Hydrogen Peroxide by Fluorometric Detection
The concentration of H2O2 produced in the extracellular medium was determined in living CTR, nLES and LES fibroblasts using a commercially available hydrogen peroxide fluorometric detection kit (ADI-907-028, Enzo life sciences AG, Lausen, Switzerland) following manufacturer’s protocol. The Hydrogen Peroxide Fluorometric Detection Kit utilizes a non-fluorescent substrate, 10-Acetyl-3,7- dihydroxyphenoxazine (ADHP), to detect H2O2. H2O2 oxidizes ADHP in a one to one ratio to produce a fluorescent product, Resorufin. This oxidation is catalyzed by peroxidase in a homogeneous no-wash assay system. Briefly, 5 × 104 cells per well were plated in black 96-well plate, every sample triplicated. When the cells adhered well to the plate surface (normally, in 4 h), they were washed with pre-heated PBS and left in 50 μL of PBS for 30 min at 37 °C. After that, 50 μL of reaction cocktail containing Horseradish Peroxidase and ADHP were added to the wells. After 10 min incubation at the RT in the darkness, the fluorescence was measured with the fluorometric microplate reader Fluoroskan Ascent (Thermo Electron Corp., Vantaa, Finland). The standard curve was prepared with the H2O2 ranged in concentration from 0 to 1 μM. The amount of H2O2 excreted by the fibroblasts was expressed in pM H2O2 eq./cell.
2.14. Western Blot Analyses
To assess the levels of the proteins of interest, whole-cell lysates were diluted in Laemmli buffer (50 mM TrisHCl, pH 6.9, 10% glycerol, 1.4% 2 mercaptoethanol (ME), 6 M urea, 2% SDS, 0.01% bromphenol blue) and boiled at 96 °C for 5 min. Equal amounts of homogenates (50 µg of protein per line) were separated on 4–12% SDS-PAGE gels (Criterion XT, Bio-Rad Laboratories, Milan, Italy) and transferred to PVDF Hybond membrane (Millipore Corp., Billerica, MA, USA). The membrane was then incubated overnight at 4 °C with a primary antibody (
Table 2) diluted in TBS-T buffer (Tris buffered saline (TBS) with Tween-20 (0.1%
w/v)) with 1% BSA. After washing, the membranes were incubated with peroxidase-conjugated secondary antibodies (
Table 2) diluted in TBS-T buffer for 1 h at room temperature. The immune-labelled bands were then detected using a Super-Signal West Dura (Pierce, Rockford, IL, USA) and quantified using ImageJ free software. Results were expressed as the ratio between the densitometries of the protein of interest and loading control GAPDH revealed at the same blot. For this, the blots were stripped with the stripping buffer (100 mM 2-mercaptoethanol, 2% SDS, 62.5 mM Tris, pH 6.8) for 45 min at 50 °C with gentle shaking, washed with TBS-T buffer, re-blocked in TBS-T with 1% BSA for at least 1 h and re-probed with different antibody. The same re-probing protocol was applied also for MAPK molecular pathways activation: p38 and pp38, ERK1/2 and pERK1/2, JNK and pJNK pairs were revealed at the same blots.
2.15. Nox4 RNA Interference (RNAi) Assay
CTR, nLES and LES fibroblasts at semi-confluence were washed in pre-heated PBS and left in serum- and antibiotics-free DMEM for 30 min at 37 °C. Further, 50 nM of siNOX4 and Lipofectamin2000 separately diluted in Opti-MEM were mixed at a ratio 2.5/1, respectively, and incubated in a dark at RT for 20 min. Subsequently, the formed si-NOX4-Lipofectamin2000 complexes were added to the cells. In 4–5 h after transfection, the medium was changed with completed DMEM. In 24 h after the transfection (day 1) the cells were used for co-culture experiments. FITC-scRNA was used as a positive control for transfection and was controlled every 24 h following transfection. Transfection level was confirmed to be stable for at least 72 h after transfection and represented at least 80% of transfected cells. NOX4 protein expression was controlled by Western blot in 24 h and in 72 h (not shown) following transfection with siNOX4 RNA. Transfection with siNOX4 RNA suppressed NOX4 by 65–70% (24 h after the transfection) and was effective and didn’t change significantly for at least 72 h after transfection.
2.16. Figure Preparation
Images were assembled in panels using Adobe Photoshop CS (version 10.0) and Adobe Illustrator CS5.1 software provided by the department of Biomedical Experimental and Clinical Sciences, University of Florence (Florence, Italy). When the brightness/contrast were adjusted the exact same procedure was applied to all images of the corresponding panel in order to maintain the eventual differences and allow the images to be comparable to each other.
The protein bands from the Western blot images were quantified using Image J free software and GraphPad Prism 5 software provided by the department of Biomedical Experimental and Clinical Sciences, University of Florence (Florence, Italy).
2.17. Statistical Analysis
The data is expressed as mean ± SD. Comparisons between different groups were performed by one-way analysis of variance followed by Tukey Test with the GraphPad Prism 5 software (San Diego, CA, USA). A p-value of <0.05 was accepted as statistically significant.
4. Discussion
In the present study, we describe in detail the redox features of fibroblasts obtained from lesional (LES) and non-lesional (nLES) skin of psoriatic patients in comparison to control fibroblasts (CTR). nLES fibroblasts did not manifest any significant alteration in intracellular Ca2+, •NO and ROS which, instead, were significantly increased in LES fibroblasts compared to CTR. However, a significant increase in mitochondrial superoxide production was correlated with the increased lipid peroxidation and decreased total antioxidant capacity in nLES with respect to CTR fibroblasts. Although in-depth studies on lipid peroxidation should be conducted, our preliminary data indicate the presence of a redox shift towards higher oxidation in nLES vs. CTR in visually unaffected skin of psoriatic patients. This could explain the higher sensitivity of the patients’ skin to develop the psoriatic plaques under exposure to intrinsic and extrinsic factors.
The acute imbalance of intracellular ROS, Ca
2+ and •NO observed in LES fibroblasts confirms that diverse cellular signalling events are regulated by the concomitant increase in these transients [
13]. NOX4, which was found to be over-expressed in LES fibroblasts, was shown to be an upstream regulator of nitric oxide synthase (NOS) [
14]. Importantly, increased •NO synthesis can display a role in psoriasis pathogenesis as it may enhance the release and actions of calcitonin gene-related peptide and substance P, which can induce the production of adhesion molecules, keratinocyte hyper-proliferation, mast cell degranulation, vasodilatation, and chemotaxis of neutrophils. Some authors also reported that •NO stimulates epithelial cells to release chemokines and growth mediators which appear to be important for keratinocyte proliferation and angiogenesis [
15]. Our findings indicate that increased NOX4 expression parallels the significantly enhanced NADPH oxidase-dependent extracellular ROS production in LES fibroblasts. Indeed, NOX4 is the only member of NADPH oxidase family that is constitutively active and regulated on its expression level [
16]. The extracellular production of H
2O
2 by LES was confirmed in this study with two independent approaches. Importantly, enhanced mitochondrial activity in LES did not contribute to extracellular ROS over-production: ROS production was completely abolished by NADPH oxidase inhibitor diphenylene iodonium and was not different between CTR and nLES fibroblasts, where mitochondrial O
2•
− production was significantly increased. These findings demonstrate that fibroblasts obtained from the area of psoriatic plaque are characterized by altered redox balance and by a significantly enhanced NOX4 expression and activity that persists through several passages in vitro confirming the involvement of endogenous mechanisms. In our study, conducted on five psoriatic and on four control samples, we didn’t take into consideration the possible gender-associated differences in intracellular ROS production and in NADPH oxidases expression. Indeed, according to recent data obtained on animal models, although no uniform consensus reached, there may be sex-dependent differences in NOX isoforms expression [
17]. However, the present study was conducted on isolated cell cultures not exposed to gender-related blood factors, such as estrogen, and here we discuss the macro-effects exerted by fibroblasts on keratinocytes redox regulation. Thus, we speculate that for the scope of this study and in accordance with the obtained results, the impact of the patients’ sex can be neglected. Certainly, when passing to the animal model, gender will be taken into consideration.
The complex distribution of NOX4 in the human fibroblasts evidenced by the confocal microscopy and immunocytochemical staining confirms the role of NOX4 in redox signalling in physiological and pathological conditions. Indeed, other authors have shown on A549 and human heart tissue that the alternative splicing of NOX4 brings to the presence of several NOX4 isoforms that differ in intracellular localization [
18,
19]. The study of splicing isoforms in human skin fibroblasts and keratinocytes as well as the expression of other two members of NADPH oxidase family releasing H
2O
2 as the main product, DUOX1 and DUOX2, may be the purpose of further studies.
In order to understand if the increased ROS production by fibroblasts in the extracellular medium could affect keratinocytes in co-culture, we established an easy and low-cost model of contactless co-culture of keratinocytes with CTR, nLES or LES fibroblasts. Interestingly, all types of fibroblasts significantly affected keratinocytes. In particular, we observed in keratinocytes increased ERK1/2 phosphorylation, higher intracellular ROS and cell cycle acceleration. These findings suggest that fibroblasts-derived factor(s) are able to activate the mitogenic ERK pathway resulting in increased keratinocytes proliferation. Indeed, existing data highlights that the canonical Ras–Raf–MEK–ERK MAPK pathway can activate cyclin D transcription and, thus, accelerate cell cycle progression [
20]. Importantly, the ERK pathway can be directly activated by ROS [
21].
A significantly higher ERK activation, intracellular ROS accumulation and cell cycle progression rate were evident in keratinocytes co-incubated with LES fibroblasts. The significant increase in p38 phosphorylation only in keratinocytes co-cultured with LES suggests that p38 activation may be dependent on ROS concentration. Indeed, p38 and JNK pathways are stress-sensitive pathways activated by multiple stress stimuli including H
2O
2 [
22]. A different time-dependent activation of the JNK pathway by H
2O
2 was shown for distinct human cell types [
23]; this can explain the absence of pJNK activation in keratinocytes co-cultured with the fibroblasts in our conditions. Co-culture with LES fibroblasts significantly increased •NO and Ca
2+ levels in keratinocytes confirming again the tight interference between ROS and these signalling species.
It has been already proposed that fibroblasts could produce pro-proliferative factors for keratinocytes in psoriatic lesions. Among others, the attention was given to cytokines. Debets and co-authors [
24] have shown that psoriatic fibroblasts release increased amounts of IL-6 with respect to control fibroblasts. It was also demonstrated that IL-6 along with some other cytokines (for instance, IL-4 and IL-8) and growth factors mediate epidermal hyperplasia [
25]. At the same time, ROS can stimulate proliferation: Kim and co-authors have shown that the treatment of keratinocytes with H
2O
2 (100 μM) or superoxide anion (generated by 1 mM xanthine and 1 mU/mL xanthine oxidase) increased proliferation rate by approximately 50% [
26]. Impressively, according to our data, NOX4 silencing not only abolished the mitogenic effect of fibroblasts on keratinocytes but also resulted in higher accumulation of cells in G0/G1 phase in case of co-culture with LES fibroblasts. Thus, our data demonstrate that NOX4-mediated redox signalling has a central role in the fibroblast/keratinocyte cross-talk.
Taken together, the findings here reported indicate a marked redox imbalance in LES fibroblasts associated with increased intracellular oxidative stress markers and excessive extracellular NOX4-derived ROS production. Future studies have to elucidate the mechanism through which NOX4 remains hyper-activated in vitro for at least seven passages of LES fibroblasts. The search in PheGenI and EMBL-EBI databases didn’t reveal any NOX4 polymorphism associated with psoriasis. Thus, the dermis is able to generate and maintain the pathologic condition associated with the NOX4 hyper-activation in the absence of external stimuli highlighting the possible role of epigenetic mechanisms. Indeed, it was shown that the NOX4 mRNA expression is finely regulated by epigenetic mechanisms, involving microRNA-dependent posttranscriptional repression [
27]. Studies are in progress to reveal novel molecular mechanisms of NOX4 regulation and paracrine redox signalling.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}