
Antioxidant Activities in Kenaf (Hibiscus cannabinus) Shoots during Growth Stages and Destination of Chlorogenic Acid and Kaempferol Glycosides

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Solvents
2.2. Plant Materials
2.3. Drying Process
2.4. Antioxidant Metabolites Measurement in Kenaf Organs at Different Growth Stages
2.4.1. Determination of AA, Total Phenolic Content (TPC), Total Flavonoid Content (TFC), and Total Polysaccharide Content (TPSC)
2.4.2. Phytochemical Analysis
HPLC Analysis of Phenolic Compounds
LC-MS/MS Analysis for Phenolics Identification
Preparation of Standard Solutions and Quantification of Phenolic Compounds
2.5. Statistical Analysis
3. Results and Discussion
3.1. Variation of Fresh Weight, Dry Weight, and Water Content in Kenaf Organs
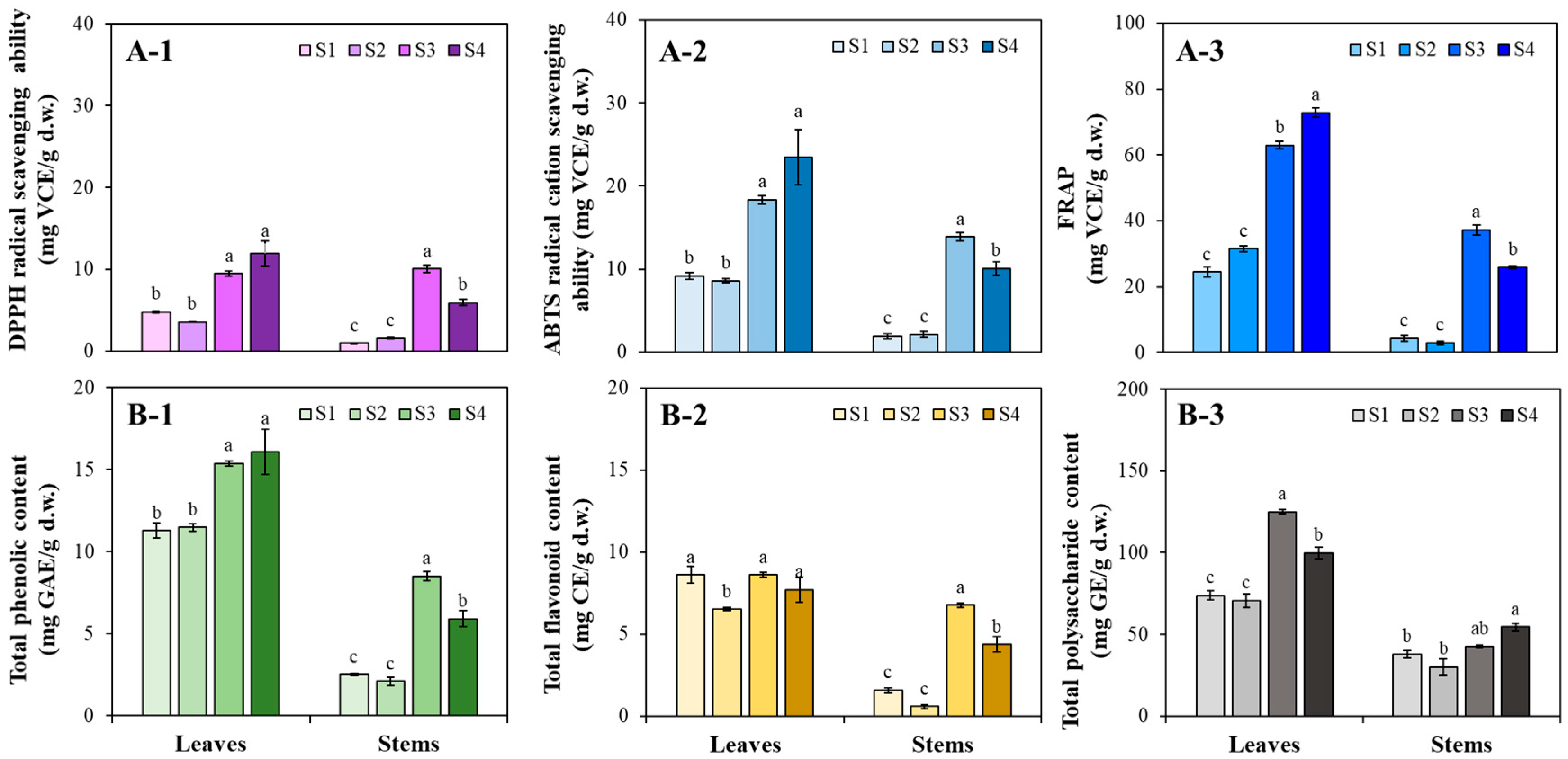
3.2. AA, TPC, TFC, and TPSC Changes in Kenaf Organs
3.3. Phenolics Variations in Kenaf Shoots
3.3.1. Phenolics Variations in Kenaf Leaves
3.3.2. Phenolics Variations in Kenaf Stems
3.4. Antioxidant Activity of Individual Phenolics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2015, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Phenolic profiles and antioxidant activities in selected drought-tolerant leafy vegetable amaranth. Sci. Rep. 2020, 10, 18287. [Google Scholar] [CrossRef] [PubMed]
- Boivin, D.; Lamy, S.; Lord-Dufour, S.; Jackson, J.; Beaulieu, E.; Côté, M.; Moghrabi, A.; Barrette, S.; Gingras, D.; Béliveau, R. Antiproliferative and antioxidant activities of common vegetables: A comparative study. Food Chem. 2009, 112, 374–380. [Google Scholar] [CrossRef]
- Chu, Y.F.; Sun, J.I.E.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common vegetables. J. Agric. Food Chem. 2002, 50, 6910–6916. [Google Scholar] [CrossRef] [PubMed]
- Kujoana, T.C.; Weeks, W.J.; Van der Westhuizen, M.M.; Mabelebele, M.; Sebola, N.A. Potential significance of kenaf (Hibiscus cannabinus L.) to global food and feed industries. Cogent Food Agric. 2023, 9, 2184014. [Google Scholar] [CrossRef]
- Kho, K.; Sim, Y.Y.; Nyam, K.L. Antioxidant activities of tea prepared from kenaf (Hibiscus cannabinus L. KR9) leaves at different maturity stages. J. Food Meas. Charact. 2019, 13, 2009–2016. [Google Scholar] [CrossRef]
- Jin, C.W.; Ghimeray, A.K.; Wang, L.; Xu, M.L.; Piao, J.P.; Cho, D.H. Far infrared assisted kenaf leaf tea preparation and its effect on phenolic compounds, antioxidant and ACE inhibitory activity. J. Med. Plants Res. 2013, 7, 1121–1128. [Google Scholar]
- Adnan, M.; Oh, K.K.; Azad, M.O.K.; Shin, M.H.; Wang, M.H.; Cho, D.H. Kenaf (Hibiscus cannabinus L.) leaves and seed as a potential source of the bioactive compounds: Effects of various extraction solvents on biological properties. Life 2020, 10, 223. [Google Scholar] [CrossRef] [PubMed]
- Shukri, S.Z.M.; Daud, N.M.A.N.; Bakar, A.R.A.; Arsad, S.S.; Zainudin, M.A.M. Profiling of bioactive compounds and bioactivity of the Kenaf (Hibiscus Cannabinus L.) leaf extract. In Proceedings of the International Conference on Biomass Utilization and Sustainable Energy, Virtual, 20–21 September 2022; Springer Nature: Singapore, 2022; pp. 415–427. [Google Scholar]
- Norhisham, D.A.; Saad, N.M.; Ahmad Usuldin, S.R.; Vayabari, D.A.; Ilham, Z.; Ibrahim, M.F.; Wan-Mohtar, W.A.A.Q.I. Bioactivities of kenaf biomass extracts: A review. Processes 2023, 11, 1178. [Google Scholar] [CrossRef]
- Ryu, J.; Kwon, S.-J.; Ahn, J.W.; Jo, Y.D.; Kim, S.H.; Jeong, S.W.; Lee, M.K.; Kim, J.-B.; Kang, S.-Y. Phytochemicals and antioxidant activity in the kenaf plant (Hibiscus cannabinus L.). J. Plant Biotechnol. 2017, 44, 191–202. [Google Scholar] [CrossRef]
- Birhanie, Z.M.; Xiao, A.; Yang, D.; Huang, S.; Zhang, C.; Zhao, L.; Liu, L.; Li, J.; Chen, A.; Tang, H.; et al. Polysaccharides, total phenolic, and flavonoid content from different kenaf (Hibiscus cannabinus L.) genotypes and their antioxidants and antibacterial properties. Plants 2021, 10, 1900. [Google Scholar] [CrossRef] [PubMed]
- Pascoal, A.; Quirantes-Piné, R.; Fernando, A.L.; Alexopoulou, E.; Segura-Carretero, A. Phenolic composition and antioxidant activity of kenaf leaves. Ind. Crops Prod. 2015, 78, 116–123. [Google Scholar] [CrossRef]
- Ryu, J.; Kwon, S.-J.; Kim, D.-G.; Lee, M.-K.; Kim, J.M.; Jo, Y.D.; Kim, S.H.; Jeong, S.W.; Kang, K.-Y.; Kim, S.W.; et al. Morphological characteristics, chemical and genetic diversity of kenaf (Hibiscus cannabinus L.) genotypes. J. Plant Biotechnol. 2017, 44, 416–430. [Google Scholar] [CrossRef]
- Park, C.H.; Kwon, S.-J.; Kim, N.S.; Baek, S.-A.; Yeo, H.J.; Park, Y.E.; Chung, Y.S.; Kim, J.K.; Park, S.U. Metabolic analysis of carotenoids and phenolic compounds found in green and purple kenaf. Nat. Prod. Commun. 2020, 15, 1934578X2097113. [Google Scholar] [CrossRef]
- Kim, J.H.; Lim, Y.J.; Duan, S.; Park, T.J.; Eom, S.H. Accumulation of antioxidative phenolics and carotenoids using thermal processing in different stages of Momordica charantia fruit. Molecules 2023, 28, 1500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Ren, X.; Zhang, X.; Wu, Z.; Liu, L. The positive correlation of antioxidant activity and prebiotic effect about oat phenolic compounds. Food Chem. 2023, 402, 134231. [Google Scholar] [CrossRef] [PubMed]
- Ghafoor, K.; Al Juhaimi, F.; Özcan, M.M.; Uslu, N.; Babiker, E.E.; Ahmed, I.A.M. Total phenolics, total carotenoids, individual phenolics and antioxidant activity of ginger (Zingiber officinale) rhizome as affected by drying methods. LWT 2020, 126, 109354. [Google Scholar] [CrossRef]
- Othman, A.; Mukhtar, N.J.; Ismail, N.S.; Chang, S.K. Phenolics, flavonoids content and antioxidant activities of 4 Malaysian herbal plants. Int. Food Res. J. 2014, 21, 759. [Google Scholar]
- Ma, Y.; Gao, J.; Wei, Z.; Shahidi, F. Effect of in vitro digestion on phenolics and antioxidant activity of red and yellow colored pea hulls. Food Chem. 2021, 337, 127606. [Google Scholar] [CrossRef]
- Sim, Y.Y.; Tan, C.P.; Cheong, L.Z.; Nyam, K.L. Hibiscus cannabinus L. leaf and seed in cosmetic formulation: An integrated approach as antioxidant and melanogenesis inhibitor. SMT 2022, 33, e00457. [Google Scholar] [CrossRef]
- Fernandes, P.A.R.; Coimbra, M.A. The antioxidant activity of polysaccharides: A structure-function relationship overview. Carbohydr. Polym. 2023, 314, 120965. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.D.N.S.; Abeysinghe, D.C.; Dharmadasa, R.M. Determination of phenolic contents and antioxidant capacity of different parts of Withania somnifera (L.) Dunal. from three different growth stages. Ind. Crops Prod. 2013, 50, 537–539. [Google Scholar] [CrossRef]
- Berezina, E.V.; Brilkina, A.A.; Veselov, A.P. Content of phenolic compounds, ascorbic acid, and photosynthetic pigments in Vaccinium macrocarpon Ait. dependent on seasonal plant development stages and age (the example of introduction in Russia). Sci. Hortic. 2017, 218, 139–146. [Google Scholar] [CrossRef]
- Feduraev, P.; Chupakhina, G.; Maslennikov, P.; Tacenko, N.; Skrypnik, L. Variation in phenolic compounds content and antioxidant activity of different plant organs from Rumex crispus L. and Rumex obtusifolius L. at different growth stages. Antioxidants 2019, 8, 237. [Google Scholar] [CrossRef] [PubMed]
- Pandjaitan, N.; Howard, L.R.; Morelock, T.; Gil, M.I. Antioxidant capacity and phenolic content of spinach as affected by genetics and maturation. J. Agric. Food Chem. 2005, 53, 8618–8623. [Google Scholar] [CrossRef]
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The versatility of antioxidant assays in food scienceand safety-chemistry, applications, strengths, and limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stages | Leaves | Stems | ||||
|---|---|---|---|---|---|---|
| Fresh Weight (g/Plant) | Dry Weight (g/Plant) | Water Content (%) | Fresh Weight (g/Plant) | Dry Weight (g/Plant) | Water Content (%) | |
| S1 | 18 ± 1 c | 0.8 ± 0.2 c | 95 ± 0 a | 16.3 ± 0.2 c | 0.5 ± 0.0 d | 97 ± 0 a |
| S2 | 315 ± 34 b | 64 ± 10 b | 80 ± 3 b | 566 ± 16 b | 130 ± 12 c | 77 ± 3 b |
| S3 | 568 ± 81 a | 114 ± 17 a | 80 ± 5 b | 1314 ± 138 a | 263 ± 37 b | 80 ± 5 b |
| S4 | 417 ± 51 a | 102 ± 11 a | 76 ± 5 b | 1408 ± 117 a | 486 ± 55 a | 65 ± 4 c |
| Peak No. | Rt (min) | MW | m/z + | MS/MS b | λmax (nm) c | Identification |
|---|---|---|---|---|---|---|
| 1 | 14.92 | 354.3 | 216.5/325.5 | CGA d | ||
| 2 a | 25.20 | 594.2 | 593.2 | 238.9/160.9 | 264.8/345.8 | Kf-gly 1 |
| 3 | 26.12 | 594.3 | 595.3 | 433.2/287.1 | 264.8/347.0 | Kf-gly 2 |
| 4 | 26.99 | 564.2 | 565.2 | 433.1/287.1 | 264.8/345.8 | Kf-rham-xyl |
| 5 | 29.31 | 578.5 | 579.2 | 433.2/287.1 | 263.7/342.2 | Kfr |
| Organs | Stages | CGA | Kf-gly 1 | Kf-gly 2 | Kf-rham-xyl | Kfr |
|---|---|---|---|---|---|---|
| Leaf | S1 | 2.43 ± 0.04 b | 3.16 ± 0.03 b | 3.31 ± 0.03 b | 1.47 ± 0.01 b | 17.90 ± 0.23 b |
| S2 | 1.84 ± 0.02 c | 5.59 ± 0.06 a | 5.62 ± 0.08 ab | 2.35 ± 0.04 ab | 26.90 ± 0.42 ab | |
| S3 | 5.41 ± 0.07 a | 5.87 ± 0.12 a | 6.88 ± 0.16 a | 2.79 ± 0.06 a | 27.30 ± 0.62 ab | |
| S4 | 6.01 ± 0.82 a | 5.24 ± 0.95 ab | 6.95 ± 1.20 a | 2.49 ± 0.42 a | 28.13 ± 4.10 a | |
| Stem | S1 | 0.07 ± 0.00 c | N.D | N.D | N.D | 0.57 ± 0.04 a |
| S2 | 0.04 ± 0.00 c | N.D | N.D | N.D | 0.41 ± 0.01 b | |
| S3 | 0.38 ± 0.00 a | N.D | N.D | N.D | 0.19 ± 0.00 c | |
| S4 | 0.19 ± 0.02 b | N.D | N.D | N.D | 0.09 ± 0.01 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, S.; Kwon, S.-J.; Jeong, D.Y.; Kim, J.H.; Park, Y.R.; Kim, C.K.; Kim, J.-H.; Eom, S.H. Antioxidant Activities in Kenaf (Hibiscus cannabinus) Shoots during Growth Stages and Destination of Chlorogenic Acid and Kaempferol Glycosides. Antioxidants 2024, 13, 532. https://doi.org/10.3390/antiox13050532
Duan S, Kwon S-J, Jeong DY, Kim JH, Park YR, Kim CK, Kim J-H, Eom SH. Antioxidant Activities in Kenaf (Hibiscus cannabinus) Shoots during Growth Stages and Destination of Chlorogenic Acid and Kaempferol Glycosides. Antioxidants. 2024; 13(5):532. https://doi.org/10.3390/antiox13050532
Chicago/Turabian StyleDuan, Shucheng, Soon-Jae Kwon, Da Yun Jeong, Ji Hye Kim, You Rang Park, Chang Kyu Kim, Jae-Hee Kim, and Seok Hyun Eom. 2024. "Antioxidant Activities in Kenaf (Hibiscus cannabinus) Shoots during Growth Stages and Destination of Chlorogenic Acid and Kaempferol Glycosides" Antioxidants 13, no. 5: 532. https://doi.org/10.3390/antiox13050532