
H2O2-Induced Oxidative Stress Responses in Eriocheir sinensis: Antioxidant Defense and Immune Gene Expression Dynamics

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Crabs and Rearing Conditions
2.3. H2O2 Stress Treatment
2.4. Biochemical Analysis
2.5. Quantitative Real-Time Fluorescent PCR (qPCR) Analysis
2.6. Data Analysis
3. Results
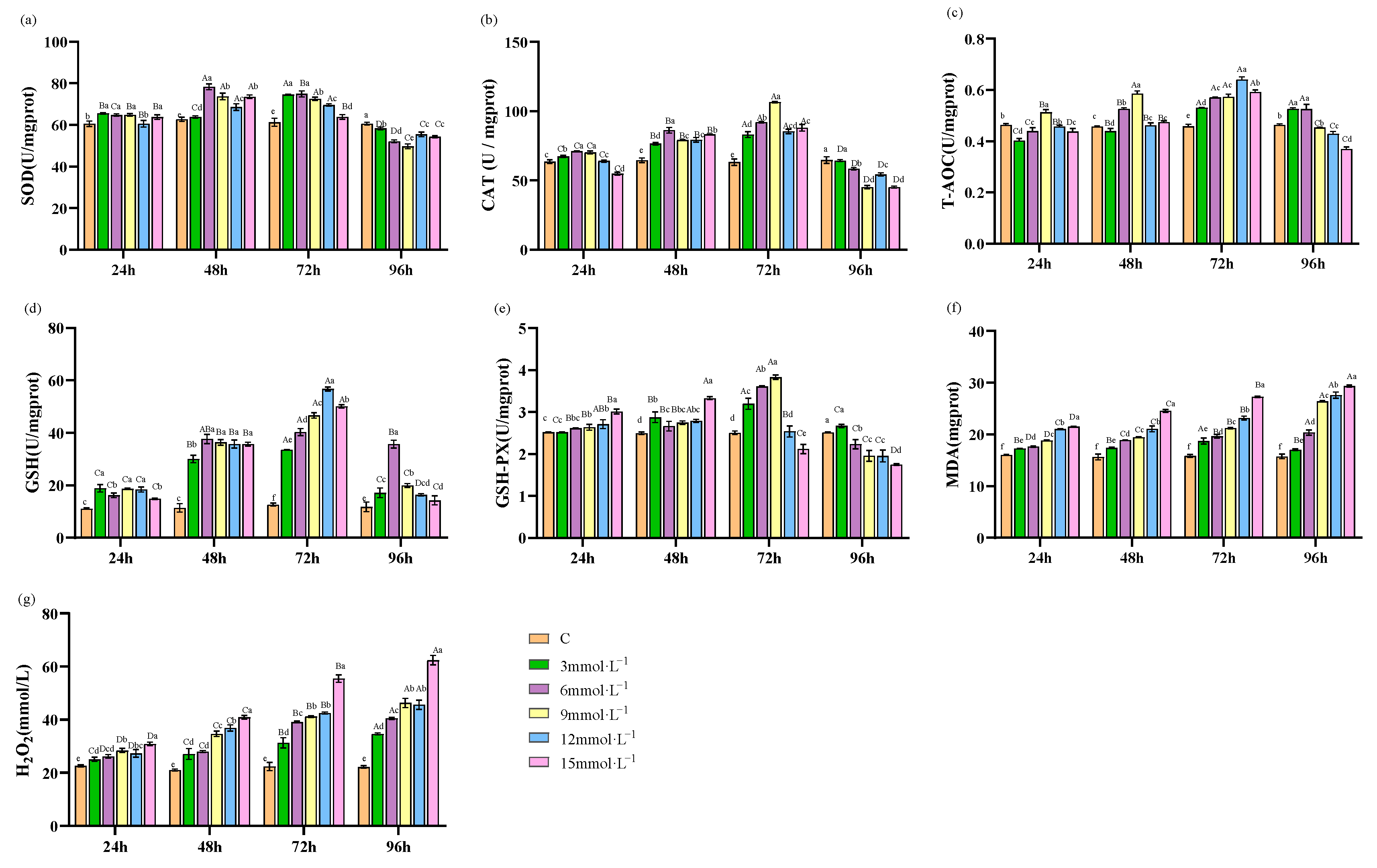
3.1. Effect of H2O2 Stress on Antioxidant Response in Hepatopancreas
3.2. Effect of H2O2 Stress on Antioxidant Response in Hemolymph
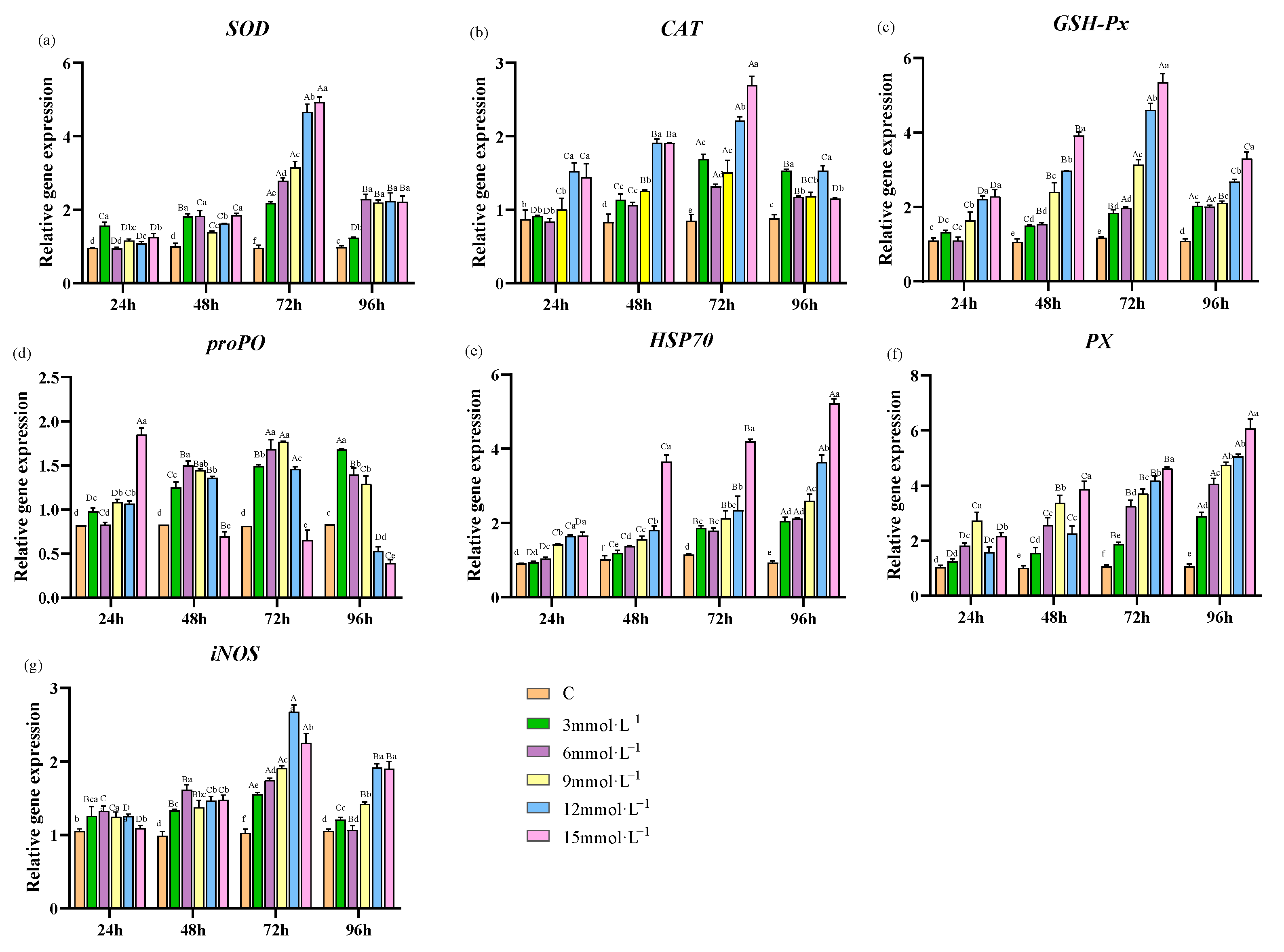
3.3. Effect of H2O2 Stress on Genes Expression in Hepatopancreas
4. Discussion
4.1. Effect of H2O2 Stress on Antioxidative Enzyme Activities in Hepatopancreas of E. sinensis
4.2. Effects of H2O2 Stress on the Antioxidant Enzyme Activities of Hemolymph in E. sinensis
4.3. Effects of H2O2 Stress on the Expression of Antioxidant- and Immune-Related Genes in Hepatopancreas
4.4. Effects of Stressors on the Antioxidant Capacity of Crustaceans
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Q.; Wu, X.; Long, X.; Zhu, W.; Ma, T.; Cheng, Y. Nutritional quality of different grades of adult male chinese mitten crab, Eriocheir sinensis. J. Food Sci. Technol. 2018, 55, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wang, J.; Yue, W.; Chen, J.; Gaughan, S.; Lu, W.; Lu, G.; Wang, C. Transcriptomic variation of hepatopancreas reveals the energy metabolism and biological processes associated with molting in Chinese mitten crab, Eriocheir sinensis. Sci. Rep. 2015, 5, 14015. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, W.; Xiong, Y.; Cheng, D.; Wang, J.; Jin, S.; Gong, Y.; Wu, Y.; Qiao, H.; Fu, H. Hepatopancreas transcriptome analyses provide new insights into the molecular regulatory mechanism of fast ovary maturation in Macrobrachium nipponense. BMC Genom. 2022, 23, 625. [Google Scholar] [CrossRef] [PubMed]
- Qiu, R.Q.R.; Cheng, Y.C.Y.; Huang, X.H.X.; Wu, X.W.X.; Yang, X.Y.X.; Tong, R.T.R. Effect of hypoxia on immunological, physiological response, and hepatopancreatic metabolism of juvenile Chinese mitten crab Eriocheir sinensis. Aquac. Int. 2011, 19, 283–299. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, Z.; Luo, L.; Wang, S.; Zhang, R.; Guo, K.; Yang, Y. Immune and intestinal microbiota responses to heat stress in Chinese mitten crab (Eriocheir sinensis). Aquaculture 2023, 563, 738965. [Google Scholar] [CrossRef]
- Bu, X.; Song, Y.; Pan, J.; Wang, X.; Qin, C.; Jia, Y.; Du, Z.; Qin, J.; Chen, L. Toxicity of chronic copper exposure on Chinese mitten crab (Eriocheir sinensis) and mitigation of its adverse impact by myo-inositol. Aquaculture 2022, 547, 737511. [Google Scholar] [CrossRef]
- Yang, X.; Song, Y.; Zhang, C.; Pang, Y.; Song, X.; Wu, M.; Cheng, Y. Effects of the glyphosate-based herbicide roundup on the survival, immune response, digestive activities and gut microbiota of the Chinese mitten crab, Eriocheir sinensis. Aquat. Toxicol. 2019, 214, 105243. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yang, C.; Zhang, S.; Rong, L.; Yang, X.; Wu, Z.; Sun, W. Metabolic changes and stress damage induced by ammonia exposure in juvenile Eriocheir sinensis. Ecotoxicol. Environ. Saf. 2021, 223, 112608. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Wang, N.; Wang, Z.; Abdallah, G.; Zhang, B.; Wang, S.; Yao, Q.; Chen, Y.; Wang, Q.; Zhang, D. Effect of infection with Aeromonas hydrophila on antioxidant capacity, inflammation response, and apoptosis proteins in Chinese mitten crab (Eriocheir sinensis). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 252, 109220. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yao, Q.; Zhang, D.; Lei, X.; Wang, S.; Wan, J.; Liu, H.; Chen, Y.; Zhao, Y.; Wang, G. Effects of acute salinity stress on osmoregulation, antioxidant capacity and physiological metabolism of female Chinese mitten crabs (Eriocheir sinensis). Aquaculture 2022, 552, 737989. [Google Scholar] [CrossRef]
- Yang, X.; Shi, A.; Song, Y.; Niu, C.; Yu, X.; Shi, X.; Pang, Y.; Ma, X.; Cheng, Y. The effects of ammonia-N stress on immune parameters, antioxidant capacity, digestive function, and intestinal microflora of Chinese mitten crab, Eriocheir sinensis, and the protective effect of dietary supplement of melatonin. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 250, 109127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhao, Z.; Li, M.; Luo, L.; Wang, S.; Guo, K.; Xu, W. Effects of saline-alkali stress on the tissue structure, antioxidation, immunocompetence and metabolomics of Eriocheir sinensis. Sci. Total Environ. 2023, 871, 162109. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Y. Preliminary Studies on the Etiology and Pathogenic Mechanism of Hepatopancreatic Necrosis Syndrome in Chinese Mitten Crab Eriocheir Sinensis; Shanghai Ocean University: Shanghai, China, 2018; (In Chinese with English abstract). [Google Scholar]
- Chang, C.; Mayer, M.; Rivera-Ingraham, G.; Blondeau-Bidet, E.; Wu, W.; Lorin-Nebel, C.; Lee, T. Effects of temperature and salinity on antioxidant responses in livers of temperate (Dicentrarchus labrax) and tropical (Chanos chanos) marine euryhaline fish. J. Therm. Biol. 2021, 99, 103016. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Dong, S.; Liu, R.; Huang, M.; Dong, K.; Ge, J.; Gao, Q.; Zhou, Y. Effects of temperature, dissolved oxygen, and their interaction on the growth performance and condition of rainbow trout (Oncorhynchus mykiss). J. Therm. Biol. 2021, 98, 102928. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Wu, D.; Li, J.; Li, C.; Zheng, X.; Wang, L. Phosphorus Nutrition in Songpu Mirror Carp (Cyprinus carpio Songpu) During Chronic Carbonate Alkalinity Stress: Effects on Growth, Intestinal Immunity, Physical Barrier Function, and Intestinal Microflora. Front. Immunol. 2022, 13, 900793. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Ahmad, T.; Sharma, J.; Chakrabarti, R. Effect of temperature on food consumption, immune system, antioxidant enzymes, and heat shock protein 70 of Channa punctata (Bloch, 1793). Fish Physiol. Biochem. 2021, 47, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cai, C.; Cao, X.; Zhu, J.; He, J.; Wu, P.; Ye, Y. Supplementation of dietary astaxanthin alleviated oxidative damage induced by chronic high pH stress, and enhanced carapace astaxanthin concentration of Chinese mitten crab Eriocheir sinensis. Aquaculture 2018, 483, 230–237. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Moniruzzaman, M.; Kumar, S.; Das, D.; Sarbajna, A.; Chakraborty, S.B. Enzymatic, non enzymatic antioxidants and glucose metabolism enzymes response differently against metal stress in muscles of three fish species depending on different feeding niche. Ecotoxicol. Environ. Saf. 2020, 202, 110954. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Zhang, D.; Wang, F.; Dong, S. Immune responses of Litopenaeus vannamei to non-ionic ammonia stress: A comparative study on shrimps in freshwater and seawater conditions. Aquac. Res. 2017, 48, 177–188. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, G.; Feng, Y.; Kong, C.; Ayisi, C.L.; Huang, X.; Hua, X. Dietary soybean antigen impairs growth and health through stress-induced non-specific immune responses in Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 84, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Li, X.; Xing, Y.; Feng, C.; Jiang, H. Respiratory metabolism and antioxidant response in Chinese mitten crab Eriocheir sinensis during air exposure and subsequent reimmersion. Front. Physiol. 2019, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Parrilla-Taylor, D.P.; Zenteno-Savín, T. Antioxidant enzyme activities in Pacific white shrimp (Litopenaeus vannamei) in response to environmental hypoxia and reoxygenation. Aquaculture 2011, 318, 379–383. [Google Scholar] [CrossRef]
- Xu, Z.; Regenstein, J.M.; Xie, D.; Lu, W.; Ren, X.; Yuan, J.; Mao, L. The oxidative stress and antioxidant responses of Litopenaeus vannamei to low temperature and air exposure. Fish Shellfish Immunol. 2018, 72, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Sohal, R.S.; Allen, R.G. Oxidative stress as a causal factor in differentiation and aging: A unifying hypothesis. Exp. Gerontol. 1990, 25, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y.; Kizaki, M.; Nakagiri, R.; Kamiya, T.; Sumi, H.; Osawa, T. Dietary glutathione protects rats from diabetic nephropathy and neuropathy. J. Nutr. 2002, 132, 897–900. [Google Scholar] [CrossRef]
- Ye, J.; Han, Y.; Zhao, J.; Lu, T.; Liu, H.; Yang, Y. Effects of dietary olaquindox on antioxidant enzymes system in hepatopancreas of Cyprinus carpio. Shuichan Xuebao 2004, 28, 231–235. [Google Scholar]
- Wang, Y.; Wang, B.; Liu, M.; Jiang, K.; Wang, M.; Wang, L. Aflatoxin B1 (AFB1) induced dysregulation of intestinal microbiota and damage of antioxidant system in pacific white shrimp (Litopenaeus vannamei). Aquaculture 2018, 495, 940–947. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, J.; Dong, H.; Wang, Y.; Liu, Q.; Li, H. Oxidative stress response of the black tiger shrimp Penaeus monodon to Vibrio parahaemolyticus challenge. Fish Shellfish Immunol. 2015, 46, 354–365. [Google Scholar] [CrossRef]
- Wang, J.; Sun, L.; Li, X.; Tao, S.; Wang, F.; Shi, Y.; Guan, H.; Yang, Y.; Zhao, Z. Alkali exposure induces autophagy through activation of the MAPKpathway by ROS and inhibition of mTOR in Eriocheir sinensis. Aquat. Toxicol. 2023, 258, 106481. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Wang, J.; Liu, L.; Li, Y.; Xun, A.; Zeng, W.; Jia, C.; Wei, X.; Feng, J.; Zhao, L. Anti-oxidant effects of resveratrol on mice with DSS-induced ulcerative colitis. Arch. Med. Res. 2010, 41, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Huang, Y.; Yan, G.; Huang, Z. Effects of deltamethrin on the antioxidant defense and heat shock protein expression in Chinese mitten crab, Eriocheir sinensis. Environ. Toxicol. Pharmacol. 2019, 66, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pilger, A.; Rüdiger, H.W. 8-Hydroxy-2′-deoxyguanosine as a marker of oxidative DNA damage related to occupational and environmental exposures. Int. Arch. Occup. Environ. Health 2006, 80, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Huang, J.; Dahms, H.; Zhen, J.; Ying, X. Cell damage and apoptosis in the hepatopancreas of Eriocheir sinensis induced by cadmium. Aquat. Toxicol. 2017, 190, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.H.; Zhang, Y.Q.; Yan, B.L.; Xu, J.T.; Tang, Y.; Du, D.D. Immunological and histological responses to sulfide in the crab Charybdis japonica. Aquat. Toxicol. 2014, 150, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Huang, Y.; Dong, Y.; Xu, D.; Huang, Q.; Huang, Z. Cytotoxicity induced by abamectin in hepatopancreas cells of Chinese mitten crab, Eriocheir sinensis: An in vitro assay. Ecotoxicol. Environ. Saf. 2023, 262, 115198. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Huang, Y.; Yan, G.; Yin, H.; Huang, Z. DNA damage, immunotoxicity, and neurotoxicity induced by deltamethrin on the freshwater crayfish, Procambarus clarkii. Environ. Toxicol. 2021, 36, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Wang, L.; Jiang, S.; Sun, M.; Wang, M.; Yi, Q.; Song, L. Functional characterization of hemocytes from Chinese mitten crab Eriocheir sinensis by flow cytometry. Fish Shellfish Immunol. 2017, 69, 15–25. [Google Scholar] [CrossRef]
- Singaram, G.; Harikrishnan, T.; Chen, F.; Bo, J.; Giesy, J.P. Modulation of immune-associated parameters and antioxidant responses in the crab (Scylla serrata) exposed to mercury. Chemosphere 2013, 90, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yuan, C.; Cui, Q. Acute toxic effects of thiamethoxam on Chinese mitten crab Eriocheir sinensis. Environ. Sci. Pollut. Res. Int. 2022, 29, 63512–63519. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Yang, X.; Huang, Y.; Yan, G.; Cheng, Y. Oxidative stress and genotoxic effect of deltamethrin exposure on the Chinese mitten crab, Eriocheir sinensis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 212, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Söderhäll, K. Early events in crustacean innate immunity. Fish Shellfish Immunol. 2002, 12, 421–437. [Google Scholar] [PubMed]
- Czapski, G.; Goldstein, S. The uniqueness both of superoxide toxicity and of the protective role of superoxide dismutase. In Oxygen Radicals in Biology and Medicine; Springer: Boston, MA, USA, 1988; pp. 43–46. [Google Scholar]
- Ren, X.; Lv, J.; Gao, B.; Li, J.; Liu, P. Immune response and antioxidant status of Portunus trituberculatus inoculated with pathogens. Fish Shellfish Immunol. 2017, 63, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Mao, Y.; Su, Y.; Wang, J. Effects of nitrite stress on mRNA expression of antioxidant enzymes, immune-related genes and apoptosis-related proteins in Marsupenaeus japonicus. Fish Shellfish Immunol. 2016, 58, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Gai, Y.; Zhao, J.; Song, L.; Li, C.; Zheng, P.; Qiu, L.; Ni, D. A prophenoloxidase from the Chinese mitten crab Eriocheir sinensis: Gene cloning, expression and activity analysis. Fish Shellfish Immunol. 2008, 24, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Goudru, H.G.; Kumar, S.; Jayalakshmi, S.K.; Ballal, C.R.; Sharma, H.C.; Sreeramulu, K. Purification and characterization of prophenoloxidase from cotton bollworm, Helicoverpa armigera. Entomol. Res. 2013, 43, 55–62. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, X.; Hou, L.; Jiang, X.; Li, C.; Zhang, J.; Pei, C.; Zhao, X.; Li, L.; Kong, X. The related immunity responses of red swamp crayfish (Procambarus clarkii) following infection with Aeromonas veronii. Aquac. Rep. 2021, 21, 100849. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, Z.; Tang, H.; Zhou, L.; Ao, S.; Zhou, Y.; Zhu, X.; Gao, X.; Jiang, Q.; Tu, C. Aeromonas hydrophila associated with red spot disease in Macrobrachium nipponense and host immune-related gene expression profiles. J. Invertebr. Pathol. 2021, 182, 107584. [Google Scholar] [CrossRef] [PubMed]
- Sivakamavalli, J.; Selvaraj, C.; Singh, S.K.; Vaseeharan, B. In vitro and in silico studies on cell adhesion protein peroxinectin from Fenneropenaeus indicus and screening of heme blockers against activity. J. Mol. Recognit. 2016, 29, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, C.U.P.; Wagner, H.; Miethke, T.C. Heat shock protein-mediated activation of innate immune cells. In Heat Shock Proteins and Inflammation; Springer: Basel, Switzerland, 2003; pp. 43–54. [Google Scholar]
- Mu, C.; Ge, Q.; Li, J. Exposure to Prorocentrum minimum induces oxidative stress and apoptosis in the ridgetail white prawn, Exopalaemon carinicauda. J. Ocean Univ. China 2019, 18, 727–734. [Google Scholar] [CrossRef]
- Park, K.; Kwak, I. Salinity and bisphenol A alter cellular homeostasis and immune defense by heat shock proteins in the intertidal crab Macrophthalmus japonicus. Estuar. Coast. Shelf Sci. 2019, 229, 106381. [Google Scholar] [CrossRef]
- Cheng, C.; Ma, H.; Deng, Y.; Feng, J.; Jie, Y.; Guo, Z. Immune and physiological responses of mud crab (Scylla paramamosain) under air exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 233, 108767. [Google Scholar] [CrossRef] [PubMed]
- Dorts, J.; Silvestre, F.; Tu, H.T.; Tyberghein, A.; Phuong, N.T.; Kestemont, P. Oxidative stress, protein carbonylation and heat shock proteins in the black tiger shrimp, Penaeus monodon, following exposure to endosulfan and deltamethrin. Environ. Toxicol. Pharmacol. 2009, 28, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Huang, Y.; Yan, G.; Pan, C.; Zhang, J. Antioxidative status, immunological responses, and heat shock protein expression in hepatopancreas of Chinese mitten crab, Eriocheir sinensis under the exposure of glyphosate. Fish Shellfish Immunol. 2019, 86, 840–845. [Google Scholar] [CrossRef] [PubMed]
- De Groote, M.A.; Testerman, T.; Xu, Y.; Stauffer, G.; Fang, F.C. Homocysteine antagonism of nitric oxide-related cytostasis in Salmonella typhimurium. Science 1996, 272, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Yu, R.; Zhou, M. Studies on nitric oxide synthase activity in haemocytes of shrimps Fenneropenaeus chinensis and Marsupenaeus japonicus after white spot syndrome virus infection. Nitric Oxide 2006, 14, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Z.; Li, C.; Zhou, L.; Liu, W.; Li, Y.; Zhang, Y.; Zheng, H.; Wen, X. Molecular cloning and expression profiles of nitric oxide synthase (NOS) in mud crab Scylla paramamosain. Fish Shellfish Immunol. 2012, 32, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Jianan, X.; Yutao, M.; Bin, L.; Hui, G.; Anli, W. Apoptosis of tiger shrimp (Penaeus monodon) haemocytes induced by Escherichia coli lipopolysaccharide. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 164, 301–306. [Google Scholar]
- Chen, X.; Feng, W.; Yan, F.; Li, W.; Xu, P.; Tang, Y. Alteration of antioxidant status, glucose metabolism, and hypoxia signal pathway in Eirocheir sinensis after acute hypoxic stress and reoxygenation. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2023, 268, 109604. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wei, L.; Cao, J.; Qiu, L.; Jiang, X.; Li, P.; Song, Q.; Zhou, H.; Han, Q.; Diao, X. Oxidative stress, DNA damage and antioxidant enzyme activities in the pacific white shrimp (Litopenaeus vannamei) when exposed to hypoxia and reoxygenation. Chemosphere 2016, 144, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Dai, T.; Zhong, S.; Jin, M.; Sun, P.; Zhou, Q. Vibrio parahaemolyticus Infection Influenced Trace Element Homeostasis, Impaired Antioxidant Function, and Induced Inflammation Response in Litopenaeus vannamei. Biol. Trace Element Res. 2020, 199, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, Y.; Dong, H.; Zhang, J. Effect of desiccation on oxidative stress and antioxidant response of the black tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 2016, 58, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Song, Z.; Zhong, S.; Gan, J.; Liang, H.; Yu, Y.; Wu, G.; He, L. Acute hypoxia and reoxygenation induces oxidative stress, glycometabolism, and oxygen transport change in red swamp crayfish (Procambarus clarkii): Application of transcriptome profiling in assessment of hypoxia. Aquac. Rep. 2022, 23, 101029. [Google Scholar] [CrossRef]
- Zeng, Q.; Yang, Q.; Chai, Y.; Wei, W.; Luo, M.; Li, W. Polystyrene microplastics enhanced copper-induced acute immunotoxicity in red swamp crayfish (Procambarus clarkii). Ecotoxicol. Environ. Saf. 2023, 249. [Google Scholar] [CrossRef] [PubMed]
- Ruan, G.; Li, S.; He, N.; Fang, L.; Wang, Q. Short-term adaptability to non-hyperthermal stress: Antioxidant, immune and gut microbial responses in the red swamp crayfish, Procambarus clarkii. Aquaculture 2022, 560. [Google Scholar] [CrossRef]
- Hossain, M.; Huang, H.; Yuan, Y.; Wan, T.; Jiang, C.; Dai, Z.; Xiong, S.; Cao, M.; Tu, S. Silicone stressed response of crayfish (Procambarus clarkii) in antioxidant enzyme activity and related gene expression. Environ. Pollut. 2020, 274, 115836. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.Q.; Wang, M.J.; Chen, H.G.; Chen, J.H.; Gao, H.; Huang, H.H. Immunological responses in haemolymph and histologic changes in the hepatopancreas of Charybdis japonica (A. Milne-Edwards, 1861) (Decapoda: Brachyura: Portunidae) exposed to bisphenol A. J. Crustac. Biol. 2018, 38, 489–496. [Google Scholar] [CrossRef]
- Meng, X.-L.; Liu, P.; Li, J.; Gao, B.-Q.; Chen, P. Physiological responses of swimming crab Portunus trituberculatus under cold acclimation: Antioxidant defense and heat shock proteins. Aquaculture 2014, 434, 11–17. [Google Scholar] [CrossRef]
- Wang, S.; Ji, C.; Li, F.; Wu, H. Toxicological responses of juvenile Chinese shrimp Fenneropenaeus chinensis and swimming crab Portunus trituberculatus exposed to cadmium. Ecotoxicol. Environ. Saf. 2022, 234, 113416. [Google Scholar] [CrossRef]
- Lu, Y.; Qiu, Q.; Li, C.; Cheng, L.; Liu, J. Antioxidant responses of Fenneropenaeus chinensis to white spot syndrome virus challenge. Aquac. Int. 2019, 28, 139–151. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Product Length (bp) | GenBank Accession Number |
|---|---|---|---|
| iNOS | TTGCCAGAGCCGTCAAGTT GCGCCTCGTGTTCTATGTTG | 201 | XM_050876720.1 |
| GSH-Px | ATCCTGTACCCTGCAACCAC CTCTGGGAACAGCTTCTTGG | 174 | FJ617305.1 |
| SOD | TGGACTGACGGAAGGGCTGC TGGCGTTAGGGGCGGAGTG | 128 | FJ617306.1 |
| CAT | CCTGCTCGCAGGAATCGGTG GTCCAAGGAGGTGGCGGTCA | 159 | MH178391.1 |
| HSP70 | GGCAAGGCAGCGAAGGTCATC CGGCATTGGTGACAGACTGACG | 127 | KC493625.1 |
| peroxinectin | CAGCAACGACTACAACCCGA TCCTTGCACCAGGGAATGAC | 91 | GU353176.1 |
| Prophenoloxidase | CCATGTCATCATTGCAGCGG TGTACTTGTGCCAGCGGTAG | 119 | EF493829.1 |
| β-actin | TGGGTATGGAATCCGTTGGC AGACAGAACGTTGTTGGCGA | 101 | KM244725.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Q.; Feng, W.; Chen, X.; Xu, Y.; Zhou, J.; Li, J.; Xu, P.; Tang, Y. H2O2-Induced Oxidative Stress Responses in Eriocheir sinensis: Antioxidant Defense and Immune Gene Expression Dynamics. Antioxidants 2024, 13, 524. https://doi.org/10.3390/antiox13050524
He Q, Feng W, Chen X, Xu Y, Zhou J, Li J, Xu P, Tang Y. H2O2-Induced Oxidative Stress Responses in Eriocheir sinensis: Antioxidant Defense and Immune Gene Expression Dynamics. Antioxidants. 2024; 13(5):524. https://doi.org/10.3390/antiox13050524
Chicago/Turabian StyleHe, Qinghong, Wenrong Feng, Xue Chen, Yuanfeng Xu, Jun Zhou, Jianlin Li, Pao Xu, and Yongkai Tang. 2024. "H2O2-Induced Oxidative Stress Responses in Eriocheir sinensis: Antioxidant Defense and Immune Gene Expression Dynamics" Antioxidants 13, no. 5: 524. https://doi.org/10.3390/antiox13050524