3.1. Yield Extraction of Lipids from ACBP
All ACBP-FGE contained high amounts of PLs (
Table 1), as the majority of the lipids were found to be polar (>90%), which is in accordance with the literature in extracts from fruit-based by-products (apple pomace, grape pomace and citrus fruits’ by-products) that were however obtained using conventional extraction methods [
5,
6,
7,
8,
9,
22]. The high yield of PLs extracted with food-grade solvents from all apple pomaces further emphasizes their potential as good sources of bio-functional dietary PLs from apple pomace.
It is noteworthy to also discuss the greater portion of PLs in the food-grade extractions, in comparison to extracts from the same samples obtained by the combination of the Bligh and Dyer and Galanos and Kapoulas extraction and separation methods [
7,
8,
22]. More specifically, the food-grade extracted samples exhibited much greater amounts of lipids, which may be associated with the higher polarity of the food-grade solvents chosen, in comparison to the less polar conventional solvents used in the extraction of Bligh and Dyer and Kapoulas and Galanos methodology, such as chloroform and petroleum ether [
22]. This increase in the ratio may indicate a more efficient extraction method as PLs were previously described as the most bioactive portion of the lipids, the NLs with very low bioactivities and the TL (PL and NL) displaying an intermediate effect [
22]. The food-grade extraction utilized in the present study effectively extracted the more polar lipid and amphiphilic bioactives from apple pomaces of the three different apple varieties (Jonagold, Dabinett and Aston Bitter) with a higher yield in comparison to that observed by applying conventional methods [
22] (
Table 1), which are rich in bioactive PLs (more than 95% of TLs are PLs). Such ACBP-FGE-PLs extracts are bio-functional extracts rich in amphiphilic bioactives that are safe to be consumed as ingredients of food supplements and nutraceuticals, but are also safe to be infused in several food products in the food industry for human consumption.
3.2. Anti-Inflammatory and Anti-Platelet Properties of Food-Grade Extracts from ACBP
The biological activities of all extracts were evaluated by acquiring their putative anti-inflammatory and anti-platelet potency against human platelet activation and aggregation induced by the inflammatory and thrombotic mediator PAF, as well as by the classic platelet agonist ADP, were assessed as previously described [
7,
22]. In order to facilitate comparisons between the PL bioactives of apple pomace that were extracted either by conventional methods or by food-grade methods,
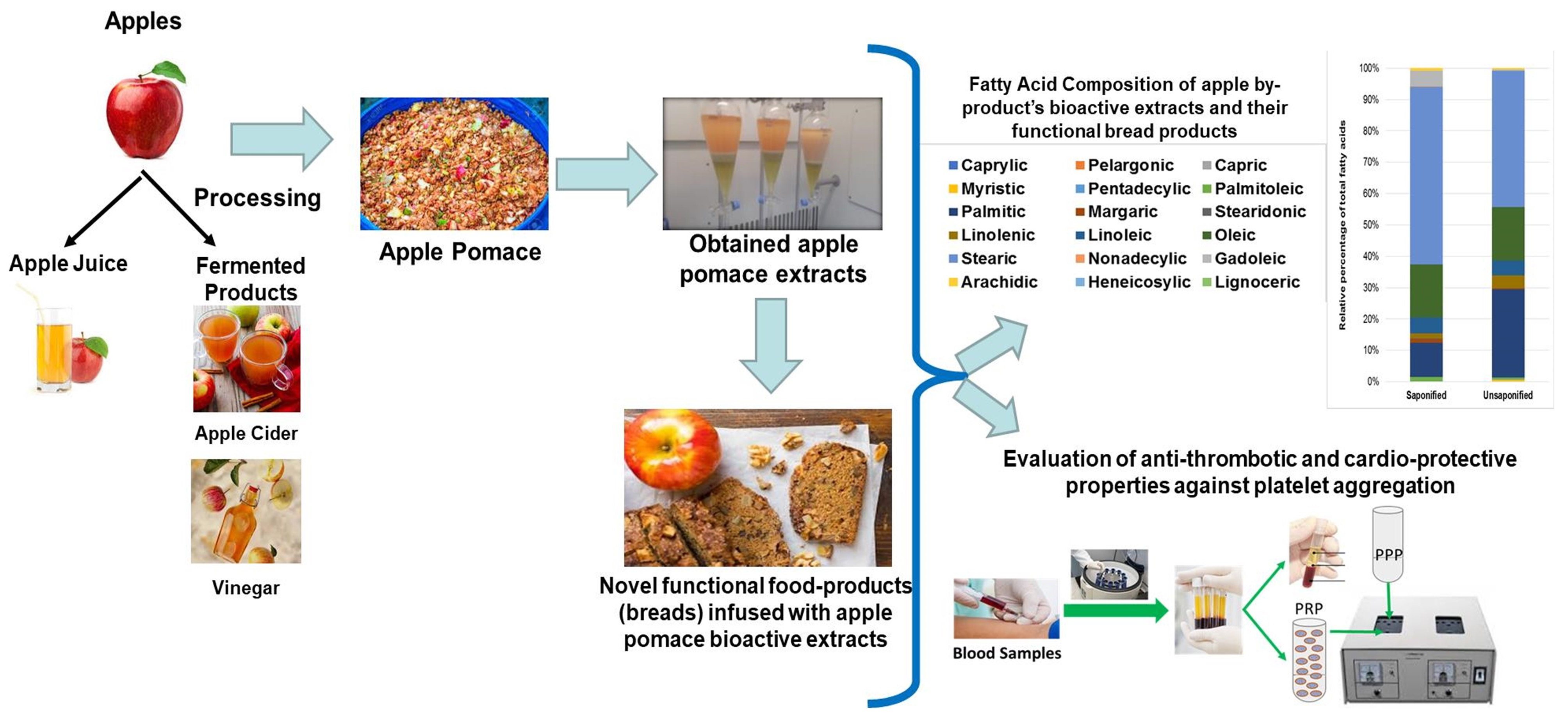
Figure 1 shows an overview of the anti-inflammatory and anti-platelet potency (IC50 value) of the ACBP-FGE-PLs of from apple pomace of low (ACBPA)-, intermediate (ACBPB)- and high-in-content tannins (ACBPC), respectively, against human platelet aggregation induced via the inflammatory and thrombotic mediator PAF or via a classic platelet agonist (ADP) (green and red bars depict the anti-PAF and anti-ADP activities of ACBP-FGE-PL, respectively). In the same figure, the previously reported anti-PAF and anti-ADP effects of bioactive PLs extracted from these ACBPs with conventional extraction methods (ACBP-CE-PLs) [
22] are also depicted (blue and yellow bars depict the anti-PAF and anti-ADP activities of ACBP-CE-PL, respectively), in order to facilitate comparisons. The results are expressed as means of the IC50 values in µg of PLs in the aggregometer cuvette that causes the 50% inhibition of PAF/ADP-induced platelet aggregation. It should be mentioned that the lower the IC50 value for an extract, the higher its inhibitory effect against the specific agonist of platelet aggregation.
All ACBP-FGE-PLs showed strong anti-inflammatory and anti-platelet bioactivities against human platelet aggregation induced by the inflammatory and thrombotic mediator PAF and the classic platelet agonist ADP. From all apple pomaces, the apple pomace of the low-in-tannins apple variety Jonagold (ACBPA) was found to contain food-grade extracted PL bioactives (ACBPA-FGE-PLs) with the strongest anti-PAF and anti-ADP potency (lower IC-50 values), with a statistical significant difference when compared with the ACBPB-FGE-PL and ACBPC-FGE-PL from the other two intermediate- and high-in-tannins ACBPs, respectively, in which both the anti-PAF and anti-ADP effects of their ACBPB-FGE-PLs and ACBPC-FGE-PLs were of lower potencies (higher IC50 values). (p < 0.05 for all these comparisons).
Thus, in contrast to the ACBP-CE-PLs, and as it is shown
Figure 1, the anti-inflammatory and anti-platelet potency of the low-in-tannins ACBPA-FGE-PLs against both the PAF pathway of inflammation and thrombosis and the ADP pathway of platelet aggregation was found to be statistically significantly stronger when compared to the relative bioactivities of the higher-in-tannins ACBPB-FGE-PLs and ACBPC -FGE-PLs. These results show that more bioactive PLs are present or were able to be extracted from the low-in-tannins apple pomace ACBPA rather than from the higher-in-tannins apple pomaces ACBPB and ACBPC. For this reason, the low-in-tannins ACBPA-FGE-PLs were chosen to be further valorized by incorporating them as functional ingredients of whole-grain functional bread products.
In comparison to the cider products, the by-products, although they display lower activities against PAF and ADP in all samples, show similar and comparable results to that of apple pomace PLs extracted with conventional methods [
22], while PL bioactives were extracted in higher quantities (one-order-of-magnitude-higher quantities exist in apple pomace than in apple juice and apple cider), suggesting that apple pomace is a better source for such anti-inflammatory and anti-platelet PL bioactives.
The anti-PAF activities of each ABPC-FGE-PL were of similar potencies to their anti-ADP activities from the same ACBP (
p > 0.05 when the anti-PAF activities of the ACBPA-FGE-PLs, ACBPB-FGE-PLs and ACBPC-FGE-PLs were compared to their relative anti-ADP effects within the same ACBP). This was not the case for the ACBP-CE-PLs in each ACBP where the anti-PAF effects were much stronger than their anti-ADP effects within the same ACBP (
p < 0.05 in all these comparisons) [
22], similarly to the conventionally extracted lipids found in apple juice [
7,
8].
Even though in each ACBP the ACBP-FGE-PLs showed stronger anti-ADP effects when compared to those of the ACBP-CE-PLs within the same ACBP (p < 0.05 for all these comparisons), only the ACBPA-FGE-PLs showed similar anti-PAF potency (IC50 value) to that of the ACBP-CE-PLs of the same low-in-tannins ACBPA (p > 0.05 for this comparison). This was not the case for the other two intermediate- and high-in-tannins ACBPB and ACBPC, respectively, in which the anti-PAF effects of their ACBPB-FGE-PLs and ACBPC-FGE-PLs were of lower potency (higher IC50 values) compared to the ACBP-CE-PL for these two ACBPs (p < 0.05). Based on the above, the ACBPA-FGE-PLs from the low-in-tannins-content apple cider by-product (ACBPA) seems to be the most promising ACBP-FGE-PL to be used either for producing novel food supplements with strong anti-inflammatory and anti-platelet potency and promising cardio-protective properties and preventative potential against inflammation-related chronic disorders or for the fortification of other foods in order to produce a novel functional food product with strong anti-inflammatory and anti-platelet potency and promising cardio-protective properties too. For this reason, several concentrations of the ACBPA-FGE-PLs were utilized for the fortification and production of novel whole-grain breads with functional anti-inflammatory and anti-platelet potency and promising cardio-protective properties.
3.3. Fatty Acid Composition of Food-Grade Extracted Bioactive PLs from ACBP
The fatty acid composition of the ACBP-FGE-PLs extracts from all ACBP apple pomaces was elucidated through LC–MS analysis, and the results are displayed in
Table 2. All PL extracts were found to be rich in polyunsaturated fatty acids (PUFAs), followed by lower amounts of saturated fatty acids (SFA) and the less abundant monounsaturated fatty acids (MUFA). More specifically, all bioactive ACBP-derived PL extracts contained high amounts of the most abundant essential omega-6 (n-6) PUFAs, linoleic acid (LA) (C18:2 c9, 12 n-6), followed by the essential omega-3 (n-3) PUFAs, alpha-linolenic acid (ALA) (C18:3 c9,12,15 n-3).
The results also display a variety of fatty acids that are similar to those previously reported from PL-extracts from the same orchard and breed; however, the composition of each sample is different. For example, the n-6 PUFA LA was found to be more abundant and the n-3 PUFA ALA much less abundant in the food-grade extracts, and subsequently the n-6/n-3 PUFA ratios of the food-grade extracted PL bioactives were much higher in comparison to the relative levels for these PUFA and n-6/n-3 PUFA ratios previously reported in conventionally extracted PL-bioactives from these three apple pomaces [
22]. In addition, long-chain n-3 PUFA like EPA, DPA and DHA were not detected in the food-grade PL extracts from any of the apple pomaces assessed in comparison to the very low but considerable amounts of these LC n-3 PUFA that were previously detected in conventionally extracted PL bioactives from these three apple pomaces [
22].
These alterations in the fatty acid composition and structure activity relationships between the food-grade extracted and the conventionally extracted PL-bioactives seem also to be associated with the lower efficacy of the food-grade extraction method to extract PLs containing more hydrophobic fatty acids with longer chains like the n-3 LC-PUFA (EPA, DPA and DHA), in comparison to the more hydrophobic organic solvents (chloroform and petroleum ether) used in the conventional extraction methodology, in which low but considerable amounts of such PL containing n-3 LC-PUFA like EPA, DPA and DHA were detected [
22]. Since such long-chain PUFAs are usually trapped within the pectin of apple pomace [
31], it seems that the milder and more polar food-grade solvents cannot break these complexes to extract these long-chain PUFA bioactives, in contrast to the more intense chloroform-based non-food-grade conventional extraction methods. These differences were expected with the change in lipid extraction procedures as it has been recorded in the literature that different extraction methods can produce different fatty acid compositions [
32].
Nevertheless, this result did not affect the bioactivities of the FGE-ACBP-PL, suggesting that other fatty acids bound in the food-grade extracted PL seems to participate in their anti-inflammatory properties like the ALA and bioactive monounsaturated fatty acids (MUFA) like oleic acid (OA, C18:1c9), which was the most abundant MUFA in all PL extracts of the three ACBPs. The MUFAs, however, were found to be in considerably lower amounts than the SFA and PUFA contents of all PLs from ACBPs, with exadecenoic acid (C16:0) being the most abundant SFA.
These results are in accordance with the previously reported fatty acid content of the bioactive PL extracts of apple products (apple juice and cider) from the same apple varieties (Jonagold, Dabinett, Aston Bitter) and of the PC bioactives of the PL extract of apple juices from the low-in-tannins Jonagold apple variety [
7,
8], and in apples and apple pomaces in general [
33,
34], while they again further suggest that such PL bioactives rich in n-3 PUFAs seem to migrate equally to their apple juice/cider products and the relevant ACBP apple pomace remnants/wastes during processing. Interestingly, the PUFA composition of the ACBP-FGE-PLs had comparable n-3 PUFA content and n-6/n-3 PUFA ratio with those observed in conventionally extracted lipids from the apple pomaces of the Jonathan and Golden Delicious apple varieties, but a much higher content of n-3 PUFA and thus a much lower n-6/n-3 PUFA ratio than those observed in conventionally extracted lipids from the apple skin of these two varieties, Jonathan skin and Golden Delicious skin, respectively [
34].
In addition, the presence of such essential an n-3 PUFA like ALA bound in the bioactive PL extracts of the ACBP further supports their anti-inflammatory potency and provides a rationale for their strong anti-PAF effects. Such dietary PLs rich in n-3 PUFA from fruits’ by-products have been found to inhibit platelet aggregation induced via the inflammatory and thrombotic mediators PAF and thrombin, but also via classic well-established platelet agonists such as collagen and ADP [
5,
6,
9,
22], as was also observed in the present study for the rich in n-3 PUFA bioactive ACBP-FGE-PLs.
Nevertheless, apart from the bioactivities observed on the PL bioactives, the n-3 PUFA content of these PLs has on its own several beneficial bio-functionalities, especially when released from these PLs in cells via specific cytoplasmic phospholipases’ A2 (PLA
2) enzymatic activities [
5]. For example, the PLA
2-based release of n-3 PUFA from such bioactive PLs in cell membranes and/or lipoproteins facilitates the production of anti-inflammatory eicosanoids that act antagonistically to other inflammatory and thrombotic eicosanoids (prostaglandins, leukotrienes, and thromboxanes) usually produced via n-6 PUFAs like arachidonic acid [
5]. The latter further supports the health benefits derived from the aforementioned n-3 PUFA ALA, while healthy dietary patterns based on these n-3 PUFA have shown strong preventative benefits against several chronic disorders, such as in a Mediterranean diet enriched in ALA for the secondary prevention of coronary heart disease [
35,
36,
37,
38].
In the ACBP-FGE-PLs, each sample displayed considerable levels of the ALA n3PUFA and of the OA MUFA that can be considered beneficial against thrombo-inflammatory mediators and associated diseases [
35,
36,
37,
38,
39]. More specifically, such PL bioactives from fruits’ by-products that are rich in the n3 PUFA ALA and the MUFA OA have been associated with anti-PAF and anti-ADP properties [
5,
6,
9,
22].
The differences observed between the fatty acid content of the ACBP-FGE-PL with that of the previously assessed fatty acid content of the ACBP-CE-PLs [
22], seem to also be associated with the lower anti-inflammatory and anti-platelet potency observed in the intermediate-in-tannins ACBPB-FGE-PL and the high-in-tannins ACBPC-FGE-PL against PAF in comparison to the higher bioactivities observed in the relevant ACBP-CE-PL bioactives for these two apple pomaces. In contrast, in these apple pomaces, the food-grade extracted PL bioactives exhibited higher anti-ADP activities than those of their conventionally extracted PL-bioactives.
In contrast, such differences were not observed in the low-in-tannins ACBPA, in which the ACBPA-FGE-PLs showed similar anti-inflammatory potency against PAF and much stronger anti-platelet efficacy against ADP in comparison to the ACBPA-CE-PL bioactives. These results further suggest that it is not only the fatty acid content that provides bio-functionality in the food-grade extracted PL bioactives from apple pomace, rather than their overall structures, while it seems that the more polar food-grade extraction methods can extract more polar lipid bioactives, such as glycolipids and amphiphilic phenolic compounds, which further contribute to the overall anti-inflammatory potency for these PL-extracts against both the inflammatory and thrombotic pathways of PAF, as well as against the platelet activation pathways of ADP. Nevertheless, more targeted studies on structural elucidation are needed in order to fully evaluate the structure-activity relationships of the food-grade extracted apple pomace PL-bioactives, in comparison to the conventionally extracted PL.
3.5. Anti-Inflammatory and Anti-Platelet Potency of Lipid Bioactives from the Novel Functional Whole-Grain Breads Infused with Several Concentrations of the Low-in-Tannins ACBPA-FGE-PLs Bioactives
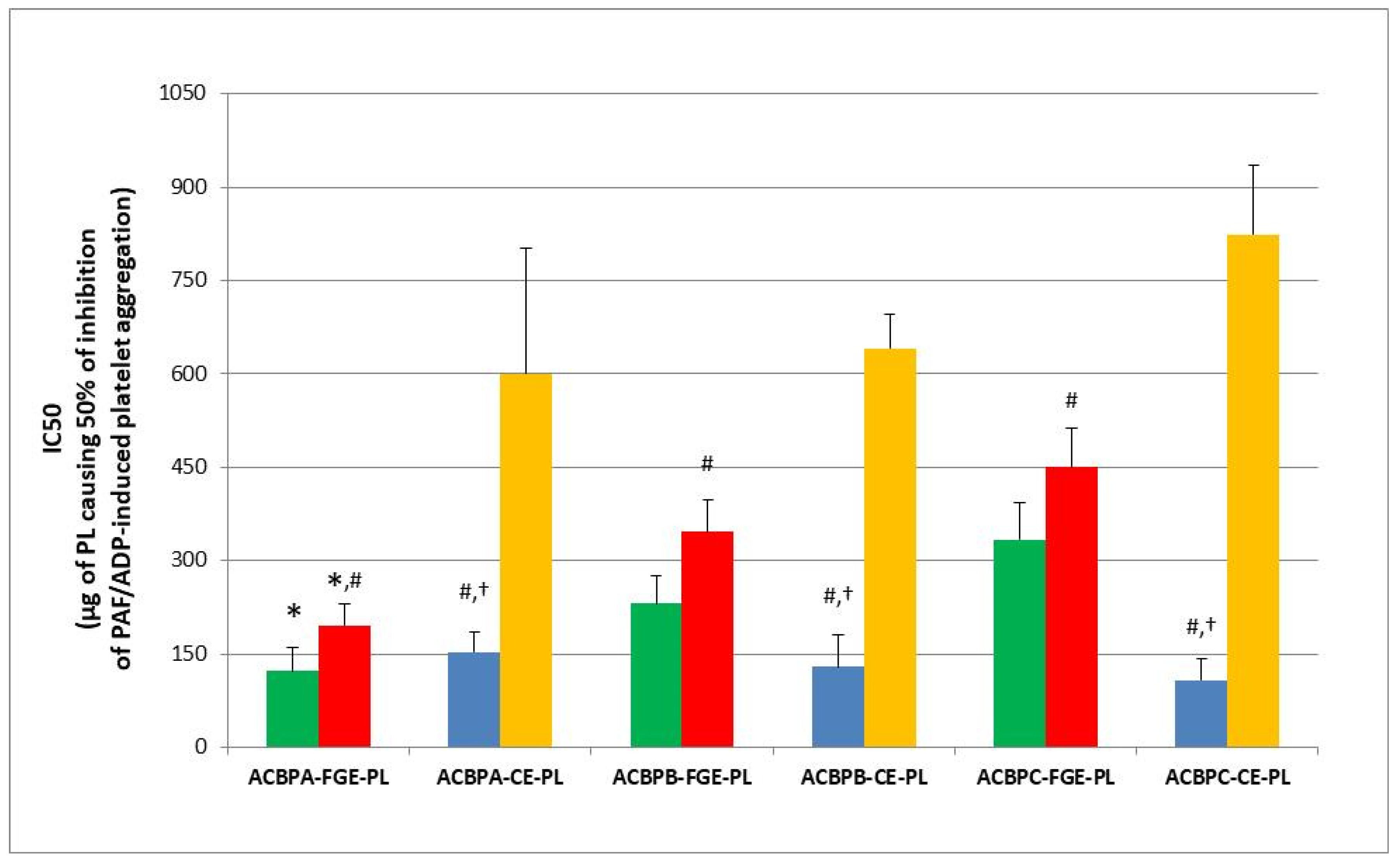
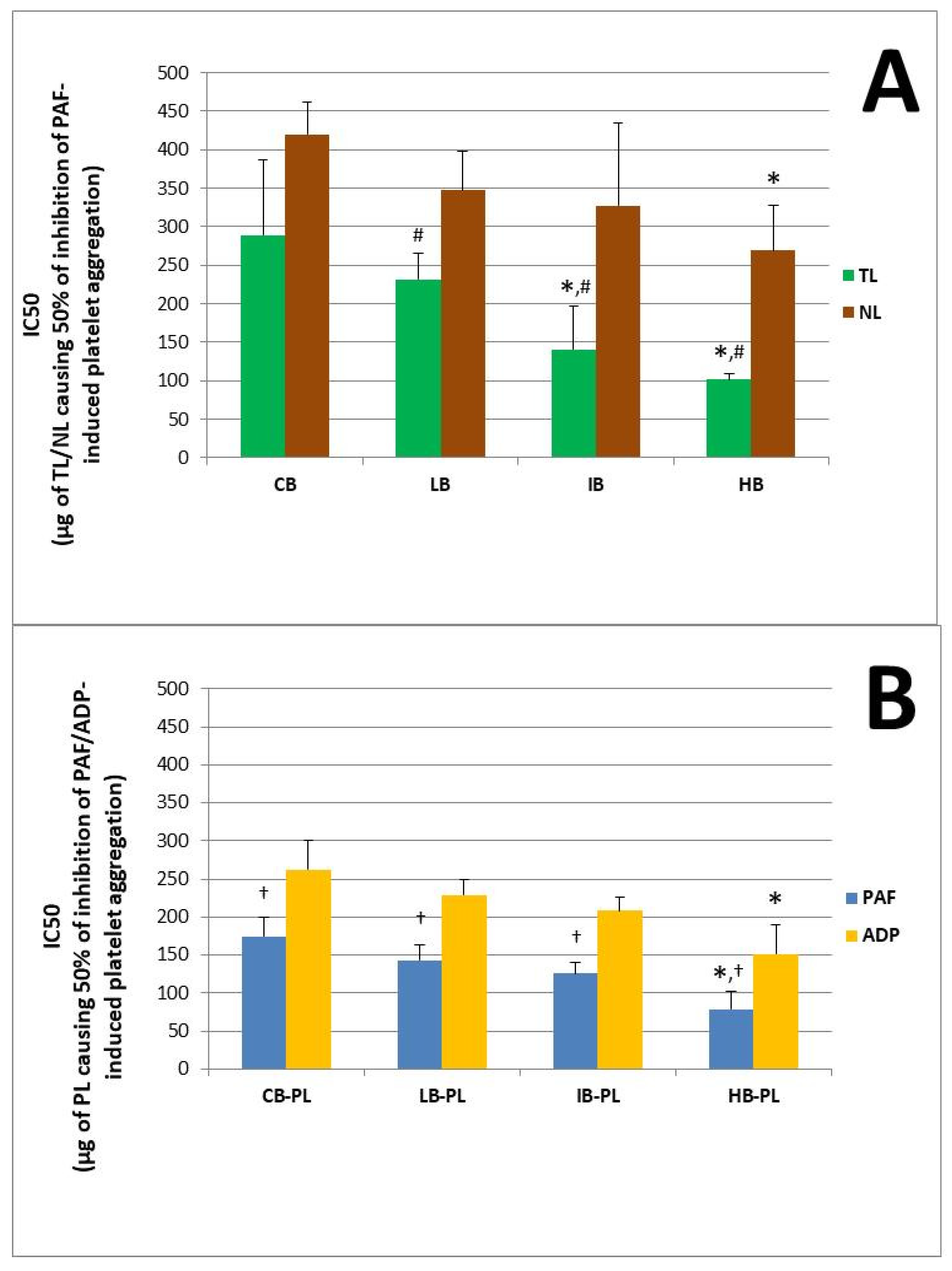
The anti-inflammatory and anti-platelet potency (IC50 value) of lipid bioactives from the novel functional whole-grain breads infused with several concentrations of the low-in-tannins ACBPA-FGE-PLs bioactives against human platelet aggregation induced via the inflammatory and thrombotic mediator PAF or via a classic platelet agonist (ADP) are depicted in
Figure 2A,B. More specifically,
Figure 2A depicts the anti-PAF potency (IC50 values) of the TL (green bars) and NL (brown bars) extracted and separated from novel functional whole-grain breads containing low (LB), intermediate (IB) and high (HB) contents (concentrations) of the infused ACBPA-FGE-PL bioactives, versus the control whole-grain bread (CB) that was not infused with ACBPA-FGE-PL bioactives. In addition,
Figure 2B depicts the anti-PAF (blue bars) and anti-ADP (yellow bars) potency (IC50 values) of the bio-functional PLs of the novel functional whole-grain LB, IB and HB breads, versus the CB control whole-grain bread. Again, results are expressed as means of the IC50 values in µg of bread-derived PLs in the aggregometer cuvette that causes the 50% inhibition of PAF/ADP-induced platelet aggregation. It should again be stressed out that the lower the IC50 value for a bread-derived PL extract, the higher its inhibitory effect against the specific agonist of platelet aggregation.
In the present study, it was found for the first time that in all breads assessed, their PLs were the most bioactive lipid extracts against PAF, followed by TL that had intermediate anti-PAF potency, and then NLs that showed the lowest bioactivities.
The TLs of all LB, IB and HB bread samples had statistically significantly higher anti-PAF potencies when compared to the relevant anti-PAF effects of the NL extracts of these breads (p < 0.05 for these comparisons). Thus, in these breads, their TLs had an anti-PAF potency with IC50 values closer to those of their PL bioactives. This was not the case for the TLs of the CB, in which the very low anti-PAF effect was of similar low potency to that of their NL extracts (p > 0.05 for these comparisons).
PL bioactives from the HB showed the highest anti-PAF potency (lowest IC50 values) in comparison to the bioactivities of the PL form all the other breads against the PAF-pathway (
p < 0.05 for all these comparisons). Also, the strong anti-inflammatory potency of the PL-bioactives extracted and separated from the novel functional HB bread is comparable to the relative ones previously observed in other fruit-derived functional foods, beverages and by-products [
5,
6,
7,
8,
9,
22,
27].
In addition, the TLs from both the HB and IB showed also statistically significantly higher anti-PAF activities (lower IC50 values) when compared to the TLs from the LB and CB (again, p < 0.05 for these comparisons). This further suggests that the higher the addition of the apple pomace lipids, the stronger the bioactivities observed in the infused breads against the inflammatory pathways of PAF-induced platelet aggregation.
Since the PLs were the most bioactive lipids of all infused breads against PAF, they were also chosen to be tested against ADP too, in a similar way to previous research for apple products and by-products [
7,
8,
22]. By this assessment, it was also found that the PL bioactives from the HB again exhibited the highest anti-ADP potency (lowest IC50 values) in comparison to the bioactivities of the PL from all the other breads against the ADP pathway (
p < 0.05 for these comparisons).
It should be noted that the anti-PAF activities of the PL bioactives in each bread were statistically significantly stronger (lower IC 50 values) when compared to their relative anti-ADP activities within the same bread sample (p < 0.05 for these comparisons), which indicates that the bread PL bioactives have higher specificity against the inflammatory and thrombotic PAF pathway rather than against the ADP pathway of platelet aggregation. Nevertheless, both bioactivities were within the same order of magnitude against both ADP- and PAF-associated pathways for the PL bioactives in each bread sample, indicating the strong cardio-protective potential for these functional breads against two distinct thrombo-inflammatory pathways.
Overall, the above observed findings further suggest that the infusion of apple pomace bioactive PL into whole-grain bread produced a novel functional bread (HB) with strong anti-inflammatory and anti-thrombotic potency against the PAF pathway and anti-platelet bioactivities against the ADP pathway of human platelet aggregation. Subsequently, such novel bio-functional breads that were developed in the present study by utilizing ACBP-FGE-PL bioactives as functional ingredients, with strong anti-inflammatory and anti-platelet properties, seem to provide more enhanced cardio-protection and preventative properties against inflammation-related chronic disorders than the classic whole-grain breads (CB) that do not contain these functional ingredients. Nevertheless, more studies are needed, mainly in vivo dietary interventions, in order to fully elucidate such an anti-platelet and anti-inflammatory potential for the novel bio-functional breads produced in the present study.
3.6. Fatty Acid Profile of the PL from Bread Infused with Apple Pomace PL
Table 4 displays the fatty acid profile of the PL extracts from all novel functional whole-grain LB, IB and HB breads that were infused with ACBPA-FGE-PL bioactives, versus the fatty acid content of the PL extracts from the control whole-grain CB bread. These samples showed many similarities in their fatty acid content; however, the addition of ACBP-FGE-PL extract caused some differences in their composition in specific fatty acids, as expected. Such differences in samples can be attributed solely to the addition of lipids, as there have been no other changes to the sample’s constituents/ingredients and the baking process. These results come in accordance with previously analyzed bread samples that were also enriched with functional lipids and fatty acids [
41].
The PL extracts from these breads displayed an abundance of unsaturated fatty acids (UFA) (>78%) with the remaining 18–22% being composed of SFA. The most abundant fatty acids in all the breads were the PUFAs with a range of 50–55%, followed by MUFA which ranged from 25–28%. Within the high levels of PUFA, the most abundant was again the omega 6 LA PUFA followed by the omega 3 ALA PUFA in all samples. Notably, the levels of the n6/n3 PUFA ratio for the PLs from all these bread samples were found to be approximately 4.5–6.5 and thus within the favorable range of healthy foods/diets and much lower than the unfavorable reported ones (>15) for western-type processed foods and diets [
35]. It has been reported that the lower the value for this ratio in a food, the better the health outcomes against inflammation-related chronic disorders [
35].
Nevertheless, these favorable low values of the n6/n3 PUFA ratio were also observed in the control CB bread too, suggesting that the added amounts of the ACBPA-FGE-PLs did not affect this ratio in the PLs of all bread samples. It seems that the same baking process applied to all breads affects similarly their PUFA content, which further suggests that higher amounts of ACBPA-GE-PLs may be needed to be infused in the bread in order to modulate the breads’ PUFA content independently of the baking process.
With respect to the MUFA content of these samples, again the n9 OA was the most abundant, followed by small but considerable portions (>4%) of palmitoleic acid (16:1). Interestingly, the content of palmitic acid was increased in the PL samples of all the breads that were infused with the ACBPA-FGE-PL, in comparison to the CB, which may further imply a saturation effect of baking on the UFA content of PL. The SFA displayed the lowest percentage in comparison to MUFA and PUFA. However, this group consisted primarily of palmitic acid (91–92% of SFA).
Although there were small changes to the FA profiles of the breads that were infused with the lipids, it must be noted that there was an addition of 1% net weight added to the higher concentrated bread. The changes observed in the fatty acid composition can be attributed to a very low amount of added lipids. These lipids are also highly sensitive to oxidation which may have occurred during the proving and baking process due to the heat treatment and access to the air surrounding the bread. By such an oxidation of the long chain UFA, it is possible that the MUFA with a lower chain may be derived, such as the palmitoleic acid and/or an SFA like the palmitic acid, which may explain their increased levels in these breads.
Such few differences in the fatty acid composition in the PL extracts of these breads seem not to affect the bioactivities against platelet aggregation mediators (PAF and ADP). For example, the HB bread’s fatty acid profile seems to be quite similar in PUFA, MUFA and SFA content to that of the CB breads. However, the anti-inflammatory properties of the HP-derived PLs are at least twice stronger than the PL of the CB. This was the case for their activity against the ADP pathway too. These outcomes further indicate that other parameters in all the breads assessed are responsible for the improvement of their functional anti-inflammatory and anti-platelet cardio-protective properties, rather than just the fatty acid profile of their PLs.
Nevertheless, it should be stressed that if bioactive PLs are present in foods, beverages and/or food supplements, after their consumption, some amounts (5–20%) of these dietary bioactive PLs (with n-3 PUFA or MUFA at their sn-2 position) are not usually digested and due to their amphiphilic properties they are absorbed and diffused from the intestine to the blood stream where they preferably are incorporated into plasma lipoproteins and, from there, to several blood cells and tissues, including platelets, but also to tissues with accessibility issues, such as the brain [
1,
5]. Their amphiphilic nature facilitates this journey within the blood stream and their incorporation into lipoproteins, cell membranes and for surpassing difficult-to-access barriers like the blood–brain barrier. The incorporation of such PL bioactives in lipoproteins like LDL and HDL cholesterol reduces beneficially their oxidative and inflammation-inducing status, while they beneficially increase the levels of the “good” HDL cholesterol, as well as also inducing the anti-oxidant and anti-inflammatory enzymatic activities of HDL with anti-inflammatory health benefits [
1,
5,
42].
Moreover, the binding of such PL bioactives to the membranes of blood cells like leukocytes and platelets, but also to the membranes of endothelial cells, directly or indirectly, inhibits several thrombo-inflammatory cascades [
1,
2,
5]. For example, PL bioactives from several foods have been found to reduce PAF synthesis in several cells, and thus reduce PAF levels in blood with subsequent beneficial reduction in the inflammatory status of blood and blood vessels [
1,
2,
5]. In addition, such food-derived PL-bioactives interact directly through a strong inhibitory antagonistic or a weak agonistic effect or both effects (in different concentrations) against the PAF and thrombin pathways of activating cells, including platelets, reducing thus the activation of these cells, including the inhibition of platelet aggregation, because of their structural resemblance to the PAF molecule and thus due to antagonism against the binding of PAF on its receptor [
1,
2,
5].
Some of these PL bioactives can also inhibit beneficially platelet aggregation indirectly due to their bio-functional fatty acid composition. More specifically, when bound to the membranes of platelets, these phospholipids are substrates for the enzyme activity of PLA
2 with a subsequent release of their bioactive MUFAs and n-3 PUFAs from the
sn-2 position of their structures into the cytoplasm, whereby in binding to specific proteins they are able to affect several thrombotic and inflammatory intracellular signaling pathways and gene expression [
1,
2,
5].
For example, the released UFAs from such PL bioactives can additionally affect the formation of pro-inflammatory eicosanoids and thus inhibit their involvement in platelet aggregation [
1,
5,
35,
36,
37,
38,
39]. UFA released intracellularly from dietary PL can inhibit specific inflammatory cyclooxygenases (COX) that are the basic enzymes involved in synthesis of the pro-inflammatory eicosanoids, which are usually induced as an after-effect of the PAF- or thrombin-induced release of arachidonic acid. Subsequently, the indirect inhibition of the PAF-/thrombin-induced intracellular cascades for the aggregation of platelets can also be associated with the UFA content of dietary PL bioactives [
1,
5].
Several other mechanisms have also been proposed for the direct inhibitory effects of UFA like the OA and ALA against platelet aggregation [
35,
36,
37,
38,
39], suggesting that the release of such bio-functional UFA of the PL bioactives in the ACBP and in the novel bread products may be able also to contribute in a reduction in the risk of platelet activation and aggregation induced by several pathways and mediators, including the PAF and ADP pathways, with several anti-inflammatory and antithrombotic health benefits, especially against the risk for inflammation-related chronic disorders.
3.7. The Total Phenolic and Carotenoid Content and Antioxidant Activities of the Food-Grade Extracted Polar Lipids from the Apple Cider By-Products (Apple Pomace), as Well as of the Polar Lipids from the Novel Functional Breads Containing the Apple Pomace Extracts versus Control Breads
The total phenolic and carotenoid content of the ACBP-FGE-PL, as well as those of the PLs from all the novel functional breads infused with the ACBPA-FGE-PL versus the CB breads are shown in
Table 5.
From all ACBP-FGE-PL extracts assessed, the apple pomace from the intermediate-in-tannins apple variety (ACBPB) showed the highest hydrophilic, lipophilic and total phenolic content, as well as total carotenoid content, followed by that of the apple pomace from the high-in-tannins apple variety (ACBPC), with the apple pomace from low-in-tannins apple variety (ACBPA) showing the lowest hydrophilic, lipophilic and total phenolic content, as well as total carotenoid content. Notably, the total phenolic content of all ACBP-FGE-PLs were comparable to those previously observed in four other apple varieties (Rome Beauty, Idared, Cortland and Golden Delicious), in which the peel contained many more phenolics than the flesh [
43].
Moreover, the carotenoid content of the ACBPB-FGE-PL and ACBPC-FGE-PL extracts showed comparable carotenoid contents with those of other apple varieties [
44], while the ACBPA-FGE-PLs showed much lower carotenoid contents, which may explain why no carotenoids were detected in the PLs of all bread samples infused with this extract. This, however, was also observed in the control CB breads too, which were not infused with the ACBPA-FGE-PLs extract, suggesting that the carotenoids’ content in all breads was independent to the presence or not of the apple pomace bioactives, rather than the baking process that seems to have saturated all carotenoids.
Interestingly, in all the produced novel functional LB, IB and HB breads that were infused with several concentrations of the food-grade extracted polar lipids from this apple pomace (ACBPA-FGE-PL), the hydrophilic phenolic content was statistically higher than that of the control CB breads that did not contain any of the functional extracts from apple pomace. In all breads, no carotenoid content was detected.
The antioxidant activity of the ACBP-FGE-PL, as well as those of the PL from all the novel functional breads infused with the ACBBPA-FGE-PL versus the CB breads, are shown in
Table 6. For evaluating the anti-oxidant activity of these samples, the FRAP assay (ferric-reducing antioxidant power assay) and the ABTS assay, compared with a Trolox (water-soluble vitamin E analogue) standard, were applied as previously described [
28,
29]. The ABTS assay measures the free radical scavenging activity, meaning the relative ability of antioxidants to scavenge the ABTS generated in aqueous phase, while the FRAP assay measures the antioxidant potential in samples through the reduction of ferric iron (Fe
3+) to ferrous iron (Fe
2+) by antioxidants present in the samples, meaning a test measuring the ferric-reducing ability of plasma [
28,
29,
45].
The differences in the substrate used (an organic radical producer was used in one method, while the other method works with a metal ion for oxidation), as well as the differences in the reaction procedures in these two experimental approaches, seem to explain the big differences observed in the outcomes derived in each assay for the same sample, as shown in
Table 5. Nevertheless, despite these differences, the results of these in vitro assays give an idea of the protective efficacy of the apple pomace-derived supplement and the novel functional bread products, and we applied two different experimental procedures as it is strongly recommended to use at least two methods due to the differences between the test systems investigated [
46]
Interestingly, all the food-grade extracted PLs from each apple pomace showed strong hydrophilic, lipophilic and total antioxidant activities in both FRAP and ABTS assays, which were comparable to that of a standard phenolic compound, quercetin, with a known and well-established anti-oxidant capacity and subsequent health benefits. Thus, these results show that the food-grade PL extracts of apple pomace have strong anti-oxidant potency, which further supports their use for producing novel functional foods and/or food supplements/nutraceuticals with anti-oxidant cardio-protective health benefits too.
Most importantly, in all the produced novel functional breads that were infused with several concentrations of the food-grade extracted polar lipids from the apple pomace of the low-in-tannins apple variety (ACBPA-FGE-PL), the hydrophilic, lipophilic and total antioxidant activities in the FRAP were statistically significantly higher than that of the control breads that do not contain any of the functional extracts from apple pomace. This result indicates that the infusion of apple pomace food-grade extracted PL (ACBPA-FGE-PL) into the ingredients for making breads produced novel functional breads with strong anti-oxidant properties.
Within the present study, the co-extraction of phenolics into the apple pomace PL extract was achieved, which were also infused within the functional breads produced. The co-presence of phenolics in these ACBP-PL extracts provides not only anti-oxidant protection for their PL bioactives, but also several health benefits against oxidative stress and blood plasma oxidation with several health-promoting effects [
1,
2,
5,
14,
19]. Moreover, the infusion of whole-grain breads with apple pomace extracts rich in PLs and phenolics resulted in an increase in their phenolic content and antioxidant capacity too, in contrast to the control breads.
Taking into account that apart from the antioxidant benefits observed, several phenolics also possess strong anti-inflammatory potency against platelet aggregation and the activities of PAF, but also have an inhibitory effect against PAF-synthesis, which further reduces the levels of this inflammatory mediator and thus the inflammatory status and PAF associated manifestations, but also several other inflammatory pathways and related disorders [
1,
2,
5,
47], further support the strong anti-inflammatory health promoting potential for both the apple pomace extracts and the functional products, in which these extracts are infused, such as in the functional breads assessed in this study. Nevertheless, more tests are needed to fully evaluate the association of the health-promoting properties of these food-grade extracts of apple pomace and their functional food products with their amphiphilic phenolics that are co-extracted with PLs in these conditions applied, as well as on the synergism(s) of these two important molecular classes of apple pomace bioactive ingredients.



 ,
, 




{kind=link}
{kind=link}
{kind=link}