
Hepatoprotective Effects of Flavonoids against Benzo[a]Pyrene-Induced Oxidative Liver Damage along Its Metabolic Pathways

Abstract
:1. Introduction
2. B[a]P-Induced Oxidative Liver Damage
2.1. Oxidative Damage Induced by B[a]P in the Liver
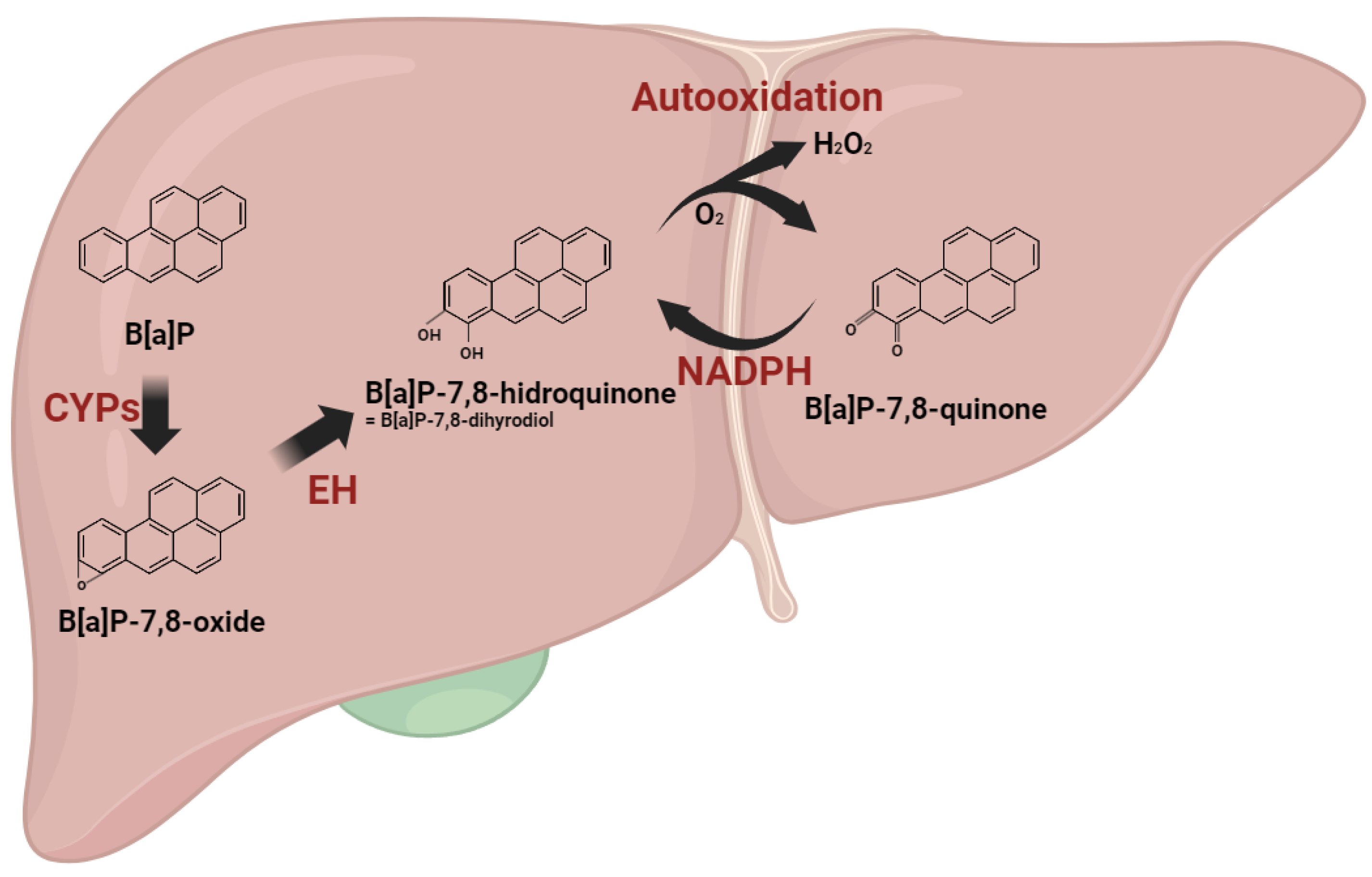
2.2. Metabolism and Excretion of B[a]P and Their Hepatic Consequences
3. Hepatoprotective Potential of Flavonoids against B[a]P-Induced Liver Damage
3.1. Classification of Flavonoids
3.2. Antioxidant Effects of Flavonoids and B[a]P in the Liver

{kind=link}
{kind=link}
{kind=link}
| Flavonoid | ROS | MDA | Antioxidant Enzyme | NRF2 Regulator | Refs. | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NRF2 | HO1 | NQO | SOD | CAT | GSH | GPX | ||||||
| Anthocyanidin | Pelargonidin | ↓ | ↓ | ↑ | ↑ | ↑ | [77] | |||||
| Cyanidin | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | [78] | |||
| ↓ | ↑ | ↑ | JNK↓ | [79] | ||||||||
| Delphinidin | ↓ | ↑ | ↑ | ↑ | Keap1↓ | [80] | ||||||
| ↓ | ↑ | PI3K/Akt↑ | [81] | |||||||||
| ↑ | ↑ | [82] | ||||||||||
| Chalcone | Phloretin | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | AMPK↑ Keap1↓ | [83] | |||
| ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | AMPK↑ | [84] | |||||
| ↓ | ↑ | ↑ | ↑ | ERK↑ | [85] | |||||||
| Xanthohumol | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | AKT↑ AMPK↑ | [86] | ||||
| ↓ | ↓ | ↑ | ↑ | ↑ | [87] | |||||||
| Flavonols | Quercetin | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | Keap1↓ | [88] | ||
| ↓ | ↓ | ↑ | NF-kB↓ | [89] | ||||||||
| ↓ | ↓ | ↑ | ↑ | ↑ | NF-kB↓ | [90] | ||||||
| Kaempferol | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | [91] | |||||
| ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [92] | |||||
| ↓ | ↓ | ↑ | ↑ | ↑ | AMPK↑ | [93] | ||||||
| ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | AKT↑ | [94] | ||||
| ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [95] | |||||
| Rutin | ↓ | ↑ | ↑ | [96] | ||||||||
| ↓ | ↑ | ↑ | ↑ | [97] | ||||||||
| ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | [98] | |||||
| Myricetin | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [99] | |||
| ↓ | ↓ | ↑ | ↑ | [100] | ||||||||
| ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | AMPK↑ | [101] | |||||
| Isorhamnetin | ↓ | ↑ | [102] | |||||||||
| Flavanone | Naringenin | ↓ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [103] | ||||
| ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | SIRT1↑ | [104] | |||||
| ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [105] | |||||
| Hesperidin | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | Keap1↓ | [106] | ||||
| ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [107] | ||||
| ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [108] | |||||
| ↓ | ↑ | ↑ | MAPK↑ | [109] | ||||||||
| Eriodictyol | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | [110] | ||||
| Flavanonols | Taxifolin | ↓ | ↓ | ↑ | ↑ | NF-kB↓ | [111] | |||||
| Flavones | Apigenin | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [112] | |
| ↓ | ↓ | ↑ | ↑ | NF-kB↓ | [113] | |||||||
| Wogonin | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [114] | ||||
| Luteolin | ↓ | ↑ | ↑ | ↑ | ↑ | NF-kB↓ | [115] | |||||
| ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | KEAP↓ | [116] | ||
| ↓ | ↑ | [117] | ||||||||||
| Tangeretin | ↓ | ↑ | ↑ | ↑ | ↑ | MAPK↑ | [118] | |||||
| Chrysin | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | [119] | |||||
| Isoflavonoids | Genistein | ↓ | ↑ | ↑ | [120] | |||||||
| Daidzein | ↓ | ↑ | [121] | |||||||||
| ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | [122] | ||||||
3.3. Regulation of B[a]P Metabolism by Flavonoids in the Liver
| Flavonoid | Phase I Enzyme | Phase II Enzyme | Phase III Enzyme | B[a]P | Refs. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AhR | CYP1A1 | CYP1B1 | Other CYPs | NRF2 | PXR | GST | UGT | Metabolism | Detoxification | ||||
| Anthocyanidin | Pelargonidin | ↑ | ↑ | CYP1A2↑ | [132] | ||||||||
| Chalcone | Phloretin | CYP1A2↓ CYP3A4↓ | ↑ | ↑ | [133] | ||||||||
| ↑ | [134] | ||||||||||||
| Flavonols | Quercetin | ↑ | [135] | ||||||||||
| ↑ | CYP2↑ CYP3↑ | [136] | |||||||||||
| ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | ABCC1↑ | ↑ | ↑ | [51] | ||||
| Kaempferol | ↓ | ↓ | CYP1A2↓ | [137] | |||||||||
| CYP2E1↓ | [138] | ||||||||||||
| ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | [126] | |||||||
| Rutin | ↑ | ↑ | [139] | ||||||||||
| ↑ | CYP1A2↑ | [140] | |||||||||||
| CYP2E1↑ | ↑ | [141] | |||||||||||
| Myricetin | CYP2C8↓ | [142] | |||||||||||
| ↓ | ↑ | ↓ | ↑ | [108] | |||||||||
| Isorhamnetin | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | ABCC1↑ | ↑ | ↑ | [51] | |||
| Flavanone | Eriodictyol | CYP2E1↓ CYP3A11↓ | ↑ | ↑ | [143] | ||||||||
| Sakuranetin | CYP1A2↓ CYP2C9↓ | ↑ | ↓ | [144] | |||||||||
| Flavanonols | Taxifolin | CYP2E1↓ CYP1A2↓ CYP3A4↓ | ↑ | [145] | |||||||||
| Flavones | Apigenin | CYP4F2↓ | [146] | ||||||||||
| ↓ | ↓ | ↓ | Total contents↓ | ↑ | ↓ | ↑ | [127] | ||||||
| Wogonin | ↓ | ↓ | [129] | ||||||||||
| ↓ | CYP2E1↓ | n.s. | ↓ | n.s. | [128] | ||||||||
| Luteolin | CYP1A2↓ CYP3A4↓ | ↓ | [147] | ||||||||||
| ABCA1↑ | [148] | ||||||||||||
| ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | [126] | |||||||
| Tangeretin | ↓ | ↓ | CYPs↓ | ↑ | [149] | ||||||||
| Chrysin | ↑ | [150] | |||||||||||
| CYP2E1↑ | ↑ | [151] | |||||||||||
| ↑ | n.s. | ↑ | ↑ | ↑ | [130] | ||||||||
| Isoflavonoids | Genistein | ↑ | ↑ | [152] | |||||||||
| ↓ | ↓ | [153] | |||||||||||
| ↑ | ↑ | [154] | |||||||||||
| ↑ | [155] | ||||||||||||
| Daidzein | ↑ | ↑ | [154] | ||||||||||
| ↑ | ↑ | [156] | |||||||||||
3.4. B[a]P-Induced Liver Disease and Flavonoid
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bukowska, B.; Mokra, K.; Michalowicz, J. Benzo[a]pyrene-Environmental Occurrence, Human Exposure, and Mechanisms of Toxicity. Int. J. Mol. Sci. 2022, 23, 6348. [Google Scholar] [CrossRef]
- Uno, S.; Dalton, T.P.; Shertzer, H.G.; Genter, M.B.; Warshawsky, D.; Talaska, G.; Nebert, D.W. Benzo[a]pyrene-induced toxicity: Paradoxical protection in Cyp1a1(−/−) knockout mice having increased hepatic BaP-DNA adduct levels. Biochem. Biophys. Res. Commun. 2001, 289, 1049–1056. [Google Scholar] [CrossRef]
- Cavalieri, E.; Rogan, E. Role of radical cations in aromatic hydrocarbon carcinogenesis. Environ. Health Perspect. 1985, 64, 69–84. [Google Scholar] [CrossRef]
- Francis, P.; Navarro, V.J. Drug-Induced Hepatotoxicity; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- Chen, S.Y.; Wang, L.Y.; Lunn, R.M.; Tsai, W.Y.; Lee, P.H.; Lee, C.S.; Ahsan, H.; Zhang, Y.J.; Chen, C.J.; Santella, R.M. Polycyclic aromatic hydrocarbon-DNA adducts in liver tissues of hepatocellular carcinoma patients and controls. Int. J. Cancer 2002, 99, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chen, W.; Xu, X.; Li, B.; He, W.; Padilla, M.T.; Jang, J.H.; Nyunoya, T.; Amin, S.; Wang, X.; et al. RIP1 potentiates BPDE-induced transformation in human bronchial epithelial cells through catalase-mediated suppression of excessive reactive oxygen species. Carcinogenesis 2013, 34, 2119–2128. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Perez de la Lastra, J.M.; Plou, F.J.; Perez-Lebena, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Li, Y.; Xi, H.; Niu, Z.; Chen, N.; Wang, R.; Yan, Y.; Gan, X.; Wang, M.; Zhang, W.; et al. Benzo(a)pyrene and cardiovascular diseases: An overview of pre-clinical studies focused on the underlying molecular mechanism. Front. Nutr. 2022, 9, 978475. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Huang, P.; Zhang, L.; Qiu, Y.; Qi, H.; Leng, A.; Shang, D. Hepatoprotective effect of plant polysaccharides from natural resources: A review of the mechanisms and structure-activity relationship. Int. J. Biol. Macromol. 2020, 161, 24–34. [Google Scholar] [CrossRef]
- Gajender; Mazumder, A.; Sharma, A.; Azad, M.A.K. A Comprehensive Review of the Pharmacological Importance of Dietary Flavonoids as Hepatoprotective Agents. Evid. Based Complement. Alternat Med. 2023, 2023, 4139117. [Google Scholar] [CrossRef]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef] [PubMed]
- Hrdina, A.I.H.; Kohale, I.N.; Kaushal, S.; Kelly, J.; Selin, N.E.; Engelward, B.P.; Kroll, J.H. The Parallel Transformations of Polycyclic Aromatic Hydrocarbons in the Body and in the Atmosphere. Environ. Health Perspect. 2022, 130, 25004. [Google Scholar] [CrossRef]
- McQueen, C.A. Comprehensive Toxicology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef]
- Chakraborty, R.; Ghosh, D. The effect of sequence on the ionization of guanine in DNA. Phys. Chem. Chem. Phys. 2016, 18, 6526–6533. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Gasparutto, D.; Ravanat, J.L. Oxidative damage to DNA: Formation, measurement and biochemical features. Mutat. Res. 2003, 531, 5–23. [Google Scholar] [CrossRef]
- Jee, S.C.; Kim, M.; Kim, K.S.; Kim, H.S.; Sung, J.S. Protective Effects of Myricetin on Benzo[a]pyrene-Induced 8-Hydroxy-2′-Deoxyguanosine and BPDE-DNA Adduct. Antioxidants 2020, 9, 446. [Google Scholar] [CrossRef]
- Hahm, J.Y.; Park, J.; Jang, E.S.; Chi, S.W. 8-Oxoguanine: From oxidative damage to epigenetic and epitranscriptional modification. Exp. Mol. Med. 2022, 54, 1626–1642. [Google Scholar] [CrossRef]
- Li, C.; Xue, Y.; Ba, X.; Wang, R. The Role of 8-oxoG Repair Systems in Tumorigenesis and Cancer Therapy. Cells 2022, 11, 3798. [Google Scholar] [CrossRef] [PubMed]
- Chiorcea-Paquim, A.M. 8-oxoguanine and 8-oxodeoxyguanosine Biomarkers of Oxidative DNA Damage: A Review on HPLC-ECD Determination. Molecules 2022, 27, 1620. [Google Scholar] [CrossRef] [PubMed]
- Qing, X.; Shi, D.; Lv, X.; Wang, B.; Chen, S.; Shao, Z. Prognostic significance of 8-hydroxy-2′-deoxyguanosine in solid tumors: A meta-analysis. BMC Cancer 2019, 19, 997. [Google Scholar] [CrossRef] [PubMed]
- Lagadu, S.; Lechevrel, M.; Sichel, F.; Breton, J.; Pottier, D.; Couderc, R.; Moussa, F.; Prevost, V. 8-oxo-7,8-dihydro-2′-deoxyguanosine as a biomarker of oxidative damage in oesophageal cancer patients: Lack of association with antioxidant vitamins and polymorphism of hOGG1 and GST. J. Exp. Clin. Cancer Res. 2010, 29, 157. [Google Scholar] [CrossRef] [PubMed]
- Iida, T.; Furuta, A.; Kawashima, M.; Nishida, J.; Nakabeppu, Y.; Iwaki, T. Accumulation of 8-oxo-2′-deoxyguanosine and increased expression of hMTH1 protein in brain tumors. Neuro Oncol. 2001, 3, 73–81. [Google Scholar] [CrossRef]
- Larosche, I.; Letteron, P.; Berson, A.; Fromenty, B.; Huang, T.T.; Moreau, R.; Pessayre, D.; Mansouri, A. Hepatic mitochondrial DNA depletion after an alcohol binge in mice: Probable role of peroxynitrite and modulation by manganese superoxide dismutase. J. Pharmacol. Exp. Ther. 2010, 332, 886–897. [Google Scholar] [CrossRef]
- Ji, X.; Li, Y.; He, J.; Shah, W.; Xue, X.; Feng, G.; Zhang, H.; Gao, M. Depletion of mitochondrial enzyme system in liver, lung, brain, stomach and kidney induced by benzo(a)pyrene. Environ. Toxicol. Pharmacol. 2016, 43, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. Mitochondrial dysfunction indirectly elevates ROS production by the endoplasmic reticulum. Cell Metab. 2013, 18, 145–146. [Google Scholar] [CrossRef] [PubMed]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial electron transport chain: Oxidative phosphorylation, oxidant production, and methods of measurement. Redox Biol. 2020, 37, 101674. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Dang, F.; Gao, J.; Zhao, H.; Qi, S.; Gao, M. Acute benzo[a]pyrene treatment causes different antioxidant response and DNA damage in liver, lung, brain, stomach and kidney. Heliyon 2018, 4, e00898. [Google Scholar] [CrossRef]
- Cichoz-Lach, H.; Michalak, A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroentero 2014, 20, 8082–8091. [Google Scholar] [CrossRef]
- Baillie, T.A. Metabolism and toxicity of drugs. Two decades of progress in industrial drug metabolism. Chem. Res. Toxicol. 2008, 21, 129–137. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, Q.; Cui, Q.; Huang, G.; Pan, X.; Li, S. Flavonoids and Naphthoflavonoids: Wider Roles in the Modulation of Cytochrome P450 Family 1 Enzymes. ChemMedChem 2016, 11, 2102–2118. [Google Scholar] [CrossRef]
- Kozack, R.E.; Loechler, E.L. Molecular modeling of the major adduct of (+)-anti-B[a]PDE (N2-dG) in the eight conformations and the five DNA sequences most relevant to base substitution mutagenesis. Carcinogenesis 1999, 20, 85–94. [Google Scholar] [CrossRef]
- Guo, L.; Jiang, X.; Tian, H.Y.; Yao, S.J.; Li, B.Y.; Zhang, R.J.; Zhang, S.S.; Sun, X. Detection of BPDE-DNA adducts in human umbilical cord blood by LC-MS/MS analysis. J. Food Drug Anal. 2019, 27, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Menzies, G.E.; Prior, I.A.; Brancale, A.; Reed, S.H.; Lewis, P.D. Carcinogen-induced DNA structural distortion differences in the RAS gene isoforms; the importance of local sequence. BMC Chem. 2021, 15, 51. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Sayer, J.M.; Plosky, B.S.; Yagi, H.; Boudsocq, F.; Woodgate, R.; Jerina, D.M.; Yang, W. Crystal structure of a benzo[a]pyrene diol epoxide adduct in a ternary complex with a DNA polymerase. Proc. Natl. Acad. Sci. USA 2004, 101, 2265–2269. [Google Scholar] [CrossRef]
- Barnes, J.L.; Zubair, M.; John, K.; Poirier, M.C.; Martin, F.L. Carcinogens and DNA damage. Biochem. Soc. Trans. 2018, 46, 1213–1224. [Google Scholar] [CrossRef]
- Nebert, D.W.; Karp, C.L. Endogenous functions of the aryl hydrocarbon receptor (AHR): Intersection of cytochrome P450 1 (CYP1)-metabolized eicosanoids and AHR biology. J. Biol. Chem. 2008, 283, 36061–36065. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Zhang, L. The Role of the Aryl Hydrocarbon Receptor (AhR) and Its Ligands in Breast Cancer. Cancers 2022, 14, 5574. [Google Scholar] [CrossRef] [PubMed]
- Tew, K.D.; Townsend, D.M. Glutathione-s-transferases as determinants of cell survival and death. Antioxid. Redox Signal. 2012, 17, 1728–1737. [Google Scholar] [CrossRef]
- Srivastava, S.K.; Hu, X.; Xia, H.; Awasthi, S.; Amin, S.; Singh, S.V. Metabolic fate of glutathione conjugate of benzo[a]pyrene-(7R,8S)-diol (9S,10R)-epoxide in human liver. Arch. Biochem. Biophys. 1999, 371, 340–344. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, M.; Blair, I.A.; Penning, T.M. Interception of benzo[a]pyrene-7,8-dione by UDP glucuronosyltransferases (UGTs) in human lung cells. Chem. Res. Toxicol. 2013, 26, 1570–1578. [Google Scholar] [CrossRef]
- Jancova, P.; Anzenbacher, P.; Anzenbacherova, E. Phase II drug metabolizing enzymes. Biomed. Pap. Med. Fac. Univ. Palacky. Olomouc Czech Repub. 2010, 154, 103–116. [Google Scholar] [CrossRef]
- Garg, R.; Gupta, S.; Maru, G.B. Dietary curcumin modulates transcriptional regulators of phase I and phase II enzymes in benzo[a]pyrene-treated mice: Mechanism of its anti-initiating action. Carcinogenesis 2008, 29, 1022–1032. [Google Scholar] [CrossRef]
- Stoddard, E.G.; Killinger, B.J.; Nag, S.A.; Martin, J.; Corley, R.; Smith, J.N.; Wright, A.T. Benzo[a]pyrene Induction of Glutathione S-Transferases: An Activity-Based Protein Profiling Investigation. Chem. Res. Toxicol. 2019, 32, 1259–1267. [Google Scholar] [CrossRef]
- Konig, J.; Nies, A.T.; Cui, Y.; Leier, I.; Keppler, D. Conjugate export pumps of the multidrug resistance protein (MRP) family: Localization, substrate specificity, and MRP2-mediated drug resistance. Biochim. Biophys. Acta 1999, 1461, 377–394. [Google Scholar] [CrossRef]
- Lai, Y.; Chu, X.; Di, L.; Gao, W.; Guo, Y.; Liu, X.; Lu, C.; Mao, J.; Shen, H.; Tang, H.; et al. Recent advances in the translation of drug metabolism and pharmacokinetics science for drug discovery and development. Acta Pharm. Sin. B 2022, 12, 2751–2777. [Google Scholar] [CrossRef]
- Yuan, L.; Lv, B.; Zha, J.; Wang, W.; Wang, Z. Basal and benzo[a]pyrene-induced expression profile of phase I and II enzymes and ABC transporter mRNA in the early life stage of Chinese rare minnows (Gobiocypris rarus). Ecotoxicol. Environ. Saf. 2014, 106, 86–94. [Google Scholar] [CrossRef]
- Guo, B.Y.; Xu, Z.T.; Yan, X.J.; Buttino, I.; Li, J.J.; Zhou, C.; Qi, P.Z. Novel ABCB1 and ABCC Transporters Are Involved in the Detoxification of Benzo(alpha)pyrene in Thick Shell Mussel, Mytilus coruscus. Front. Mar. Sci. 2020, 7, 119. [Google Scholar] [CrossRef]
- Bak, Y.; Jang, H.J.; Seo, J.H.; No, S.H.; Chae, J.I.; Hong, J.; Yoon, D.Y. Benzo[a]pyrene Alters the Expression of Genes in A549 Lung Cancer Cells and Cancer Stem Cells. J. Microbiol. Biotechnol. 2018, 28, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Jee, S.C.; Kim, K.S.; Kim, H.S.; Yu, K.N.; Sung, J.S. Quercetin and Isorhamnetin Attenuate Benzo[a]pyrene-Induced Toxicity by Modulating Detoxification Enzymes through the AhR and NRF2 Signaling Pathways. Antioxidants 2021, 10, 787. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, M.R. Flavonoids a Bioactive Compound from Medicinal Plants and Its Therapeutic Applications. BioMed Res. Int. 2022, 2022, 5445291. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef] [PubMed]
- Kahkonen, M.P.; Heinonen, M. Antioxidant activity of anthocyanins and their aglycons. J. Agric. Food Chem. 2003, 51, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khan, J.; Dukhyil, A.A.B.; Alarousy, R.; Attah, E.I.; Sharma, T.; Khairnar, S.J.; Bendale, A.R. Chalcone Scaffolds, Bioprecursors of Flavonoids: Chemistry, Bioactivities, and Pharmacokinetics. Molecules 2021, 26, 7177. [Google Scholar] [CrossRef] [PubMed]
- Budziak-Wieczorek, I.; Kaminski, D.; Skrzypek, A.; Ciolek, A.; Skrzypek, T.; Janik-Zabrotowicz, E.; Arczewska, M. Naturally Occurring Chalcones with Aggregation-Induced Emission Enhancement Characteristics. Molecules 2023, 28, 3412. [Google Scholar] [CrossRef]
- Chagas, M.; Behrens, M.D.; Moragas-Tellis, C.J.; Penedo, G.X.M.; Silva, A.R.; Goncalves-de-Albuquerque, C.F. Flavonols and Flavones as Potential anti-Inflammatory, Antioxidant, and Antibacterial Compounds. Oxid. Med. Cell Longev. 2022, 2022, 9966750. [Google Scholar] [CrossRef]
- Rencoret, J.; Rosado, M.J.; Kim, H.; Timokhin, V.I.; Gutierrez, A.; Bausch, F.; Rosenau, T.; Potthast, A.; Ralph, J.; Del Rio, J.C. Flavonoids naringenin chalcone, naringenin, dihydrotricin, and tricin are lignin monomers in papyrus. Plant Physiol. 2022, 188, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Koosha, S.; Alshawsh, M.A.; Looi, C.Y.; Seyedan, A.; Mohamed, Z. An Association Map on the Effect of Flavonoids on the Signaling Pathways in Colorectal Cancer. Int. J. Med. Sci. 2016, 13, 374–385. [Google Scholar] [CrossRef]
- Yi, Y.S. Regulatory Roles of Flavonoids in Caspase-11 Non-Canonical Inflammasome-Mediated Inflammatory Responses and Diseases. Int. J. Mol. Sci. 2023, 24, 10402. [Google Scholar] [CrossRef]
- Jeong, S.H.; Kim, H.H.; Ha, S.E.; Park, M.Y.; Bhosale, P.B.; Abusaliya, A.; Park, K.I.; Heo, J.D.; Kim, H.W.; Kim, G.S. Flavones: Six Selected Flavones and Their Related Signaling Pathways That Induce Apoptosis in Cancer. Int. J. Mol. Sci. 2022, 23, 10965. [Google Scholar] [CrossRef]
- Lorent, K.; Gong, W.; Koo, K.A.; Waisbourd-Zinman, O.; Karjoo, S.; Zhao, X.; Sealy, I.; Kettleborough, R.N.; Stemple, D.L.; Windsor, P.A.; et al. Identification of a plant isoflavonoid that causes biliary atresia. Sci. Transl. Med. 2015, 7, 286ra267. [Google Scholar] [CrossRef]
- Zhou, J.; Zheng, Q.; Chen, Z. The Nrf2 Pathway in Liver Diseases. Front. Cell Dev. Biol. 2022, 10, 826204. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell Mol. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef] [PubMed]
- Robak, J.; Gryglewski, R.J. Flavonoids are scavengers of superoxide anions. Biochem. Pharmacol. 1988, 37, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Zheng, R.L.; Jia, Z.J.; Ju, Y. Flavonoids as superoxide scavengers and antioxidants. Free Radic. Biol. Med. 1990, 9, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Vomhof-Dekrey, E.E.; Picklo, M.J., Sr. The Nrf2-antioxidant response element pathway: A target for regulating energy metabolism. J. Nutr. Biochem. 2012, 23, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the KEAP1-NRF2 Pathway. Mol. Cell Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef]
- Moratilla-Rivera, I.; Sanchez, M.; Valdes-Gonzalez, J.A.; Gomez-Serranillos, M.P. Natural Products as Modulators of Nrf2 Signaling Pathway in Neuroprotection. Int. J. Mol. Sci. 2023, 24, 3748. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal 2018, 29, 1727–1745. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef]
- Liu, H.; Johnston, L.J.; Wang, F.; Ma, X. Triggers for the Nrf2/ARE Signaling Pathway and Its Nutritional Regulation: Potential Therapeutic Applications of Ulcerative Colitis. Int. J. Mol. Sci. 2021, 22, 11411. [Google Scholar] [CrossRef]
- Shi, Y.S.; Li, X.X.; Li, H.T.; Zhang, Y. Pelargonidin ameliorates CCl4-induced liver fibrosis by suppressing the ROS-NLRP3-IL-1beta axis via activating the Nrf2 pathway. Food Funct. 2020, 11, 5156–5165. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Cui, S.; Mao, B.; Zhang, Q.; Tian, F.; Zhao, J.; Tang, X.; Chen, W. Cyanidin Alleviated CCl4-Induced Acute Liver Injury by Regulating the Nrf2 and NF-kappaB Signaling Pathways. Antioxidants 2022, 11, 2383. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Jia, Q.; Wang, Y.; Zhang, Y.; Xia, M. The anthocyanin cyanidin-3-O-beta-glucoside, a flavonoid, increases hepatic glutathione synthesis and protects hepatocytes against reactive oxygen species during hyperglycemia: Involvement of a cAMP-PKA-dependent signaling pathway. Free Radic. Biol. Med. 2012, 52, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, Y.; Ren, G.; Yang, R.; Chen, J.; Xiang, X.; Qin, H.; Chen, J. Inhibitory Effect of Delphinidin on Oxidative Stress Induced by H2O2 in HepG2 Cells. Oxid. Med. Cell Longev. 2020, 2020, 4694760. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, H.Y.; Wang, D.; Guo, X.Z. Delphinidin protects beta2m-/Thy1+ bone marrow-derived hepatocyte stem cells against TGF-beta1-induced oxidative stress and apoptosis through the PI3K/Akt pathway in vitro. Chem. Biol. Interact. 2019, 297, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Maeda-Yamamoto, M.; Nesumi, A.; Murakami, A. Delphinidin-3-O-galactoside protects mouse hepatocytes from (-)-epigallocatechin-3-gallate-induced cytotoxicity via up-regulation of heme oxygenase-1 and heat shock protein 70. Nutr. Res. 2012, 32, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, Q.; Han, L.; Pan, C.; Lei, C.; Chen, H.; Lan, X. C2C12 Mouse Myoblasts Damage Induced by Oxidative Stress Is Alleviated by the Antioxidant Capacity of the Active Substance Phloretin. Front. Cell Dev. Biol. 2020, 8, 541260. [Google Scholar] [CrossRef]
- Yang, Q.; Han, L.; Li, J.; Xu, H.; Liu, X.; Wang, X.; Pan, C.; Lei, C.; Chen, H.; Lan, X. Activation of Nrf2 by Phloretin Attenuates Palmitic Acid-Induced Endothelial Cell Oxidative Stress via AMPK-Dependent Signaling. J. Agric. Food Chem. 2019, 67, 120–131. [Google Scholar] [CrossRef]
- Yang, Y.C.; Lii, C.K.; Lin, A.H.; Yeh, Y.W.; Yao, H.T.; Li, C.C.; Liu, K.L.; Chen, H.W. Induction of glutathione synthesis and heme oxygenase 1 by the flavonoids butein and phloretin is mediated through the ERK/Nrf2 pathway and protects against oxidative stress. Free Radic. Biol. Med. 2011, 51, 2073–2081. [Google Scholar] [CrossRef]
- Zhu, L.; Fan, X.; Cao, C.; Li, K.; Hou, W.; Ci, X. Xanthohumol protect against acetaminophen-induced hepatotoxicity via Nrf2 activation through the AMPK/Akt/GSK3beta pathway. Biomed. Pharmacother. 2023, 165, 115097. [Google Scholar] [CrossRef]
- Pinto, C.; Cestero, J.J.; Rodriguez-Galdon, B.; Macias, P. Xanthohumol, a prenylated flavonoid from hops (Humulus lupulus L.), protects rat tissues against oxidative damage after acute ethanol administration. Toxicol. Rep. 2014, 1, 726–733. [Google Scholar] [CrossRef]
- Wang, J.; Wang, K.; Ding, L.; Zhao, P.; Zhang, C.; Wang, H.; Yang, Z.; Liu, Z. Alleviating effect of quercetin on cadmium-induced oxidative damage and apoptosis by activating the Nrf2-keap1 pathway in BRL-3A cells. Front. Pharmacol. 2022, 13, 969892. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Dou, B.; Liang, J.; Hou, W.; Ma, C.; Zhang, Q. Quercetin Reduces Oxidative Stress and Apoptosis by Inhibiting HMGB1 and Its Translocation, Thereby Alleviating Liver Injury in ACLF Rats. Evid. Based Complement. Alternat Med. 2021, 2021, 2898995. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Tian, L.; Chai, G.; Wen, B.; Wang, B. Targeting heme oxygenase-1 by quercetin ameliorates alcohol-induced acute liver injury via inhibiting NLRP3 inflammasome activation. Food Funct. 2018, 9, 4184–4193. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Weng, Q.; Gong, S.; Zhang, W.; Wang, J.; Huang, Y.; Li, Y.; Guo, J.; Lan, T. Kaempferol prevents acetaminophen-induced liver injury by suppressing hepatocyte ferroptosis via Nrf2 pathway activation. Food Funct. 2023, 14, 1884–1896. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, A.S.; El-Kott, A.F.; El-Gerbed, M.S.A.; El-Kenawy, A.E.; Albadrani, G.M.; Khalifa, H.S. Kaempferol prevents cadmium chloride-induced liver damage by upregulating Nrf2 and suppressing NF-kappaB and keap1. Environ. Sci. Pollut. Res. Int. 2022, 29, 13917–13929. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Yoon, S.; Moon, J.O. Kaempferol Suppresses Carbon Tetrachloride-Induced Liver Damage in Rats via the MAPKs/NF-kappaB and AMPK/Nrf2 Signaling Pathways. Int. J. Mol. Sci. 2023, 24, 6900. [Google Scholar] [CrossRef]
- Rajendran, P.; Ammar, R.B.; Al-Saeedi, F.J.; Mohamed, M.E.; ElNaggar, M.A.; Al-Ramadan, S.Y.; Bekhet, G.M.; Soliman, A.M. Kaempferol Inhibits Zearalenone-Induced Oxidative Stress and Apoptosis via the PI3K/Akt-Mediated Nrf2 Signaling Pathway: In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2020, 22, 217. [Google Scholar] [CrossRef]
- Du, Y.C.; Lai, L.; Zhang, H.; Zhong, F.R.; Cheng, H.L.; Qian, B.L.; Tan, P.; Xia, X.M.; Fu, W.G. Kaempferol from Penthorum chinense Pursh suppresses HMGB1/TLR4/NF-kappaB signaling and NLRP3 inflammasome activation in acetaminophen-induced hepatotoxicity. Food Funct. 2020, 11, 7925–7934. [Google Scholar] [CrossRef]
- Saafan, S.M.; Mohamed, S.A.; Noreldin, A.E.; El Tedawy, F.A.; Elewa, Y.H.A.; Fadly, R.S.; Al Jaouni, S.K.; El-Far, A.H.; Alsenosy, A.A. Rutin attenuates D-galactose-induced oxidative stress in rats’ brain and liver: Molecular docking and experimental approaches. Food Funct. 2023, 14, 5728–5751. [Google Scholar] [CrossRef]
- Freitas, P.A.; Oliveira, K.A.; Magalhaes, L.A.; Neves, R.J.D.; Maia, C.S.C.; Silveira, L.R.; de Lima, T.T.; Vasconcelos, R.P.; Brito, L.C.; Torres-Leal, F.L.; et al. Improvement of 2,2′-Azobis(2-Methylpropionamidine) Dihydrochloride-Induced Hepatic Redox Imbalance in Swiss Mice and HepG2 Cells by Rutin. J. Med. Food 2022, 25, 630–635. [Google Scholar] [CrossRef]
- Singh, S.; Singh, D.K.; Meena, A.; Dubey, V.; Masood, N.; Luqman, S. Rutin protects t-butyl hydroperoxide-induced oxidative impairment via modulating the Nrf2 and iNOS activity. Phytomedicine 2019, 55, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Rostami, A.; Baluchnejadmojarad, T.; Roghani, M. Hepatoprotective Effect of Myricetin following Lipopolysaccharide/DGalactosamine: Involvement of Autophagy and Sirtuin 1. Curr. Mol. Pharmacol. 2023, 16, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, J.; Zhang, H.; Bai, W.; Dong, J.; Yang, Z.; Yang, P.; Gu, Z.; Li, Y.; Chen, X.; et al. Antioxidative myricetin-enriched nanoparticles towards acute liver injury. J. Mater. Chem. B 2022, 10, 7875–7883. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; An, B.; Yu, Q.; Cao, Y.; Liu, Y.; Li, S. The hepatoprotective effect of myricetin against lipopolysaccharide and D-galactosamine-induced fulminant hepatitis. Int. J. Biol. Macromol. 2020, 155, 1092–1104. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.Z.; Lee, J.H.; Ki, S.H.; Yang, J.H.; Cho, I.J.; Kang, S.H.; Zhao, R.J.; Kim, S.C.; Kim, Y.W. AMPK activation by isorhamnetin protects hepatocytes against oxidative stress and mitochondrial dysfunction. Eur. J. Pharmacol. 2014, 740, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Alanazi, A.Z.; Alhazzani, K.; Alrewily, S.Q.; Aljerian, K.; Algahtani, M.M.; Alqahtani, Q.H.; Haspula, D.; Alhamed, A.S.; Alqinyah, M.; Raish, M. The Potential Protective Role of Naringenin against Dasatinib-Induced Hepatotoxicity. Pharmaceuticals 2023, 16, 921. [Google Scholar] [CrossRef]
- Hua, Y.Q.; Zeng, Y.; Xu, J.; Xu, X.L. Naringenin alleviates nonalcoholic steatohepatitis in middle-aged Apoe(−/−)mice: Role of SIRT1. Phytomedicine 2021, 81, 153412. [Google Scholar] [CrossRef]
- Wali, A.F.; Rashid, S.; Rashid, S.M.; Ansari, M.A.; Khan, M.R.; Haq, N.; Alhareth, D.Y.; Ahmad, A.; Rehman, M.U. Naringenin Regulates Doxorubicin-Induced Liver Dysfunction: Impact on Oxidative Stress and Inflammation. Plants 2020, 9, 550. [Google Scholar] [CrossRef]
- Jia, Y.; Li, J.; Liu, P.; Si, M.; Jin, Y.; Wang, H.; Ma, D.; Chu, L. Based on Activation of p62-Keap1-Nrf2 Pathway, Hesperidin Protects Arsenic-Trioxide-Induced Cardiotoxicity in Mice. Front. Pharmacol. 2021, 12, 758670. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Mohammed, H.M.; Khadrawy, S.M.; Galaly, S.R. Hesperidin protects against chemically induced hepatocarcinogenesis via modulation of Nrf2/ARE/HO-1, PPARgamma and TGF-beta1/Smad3 signaling, and amelioration of oxidative stress and inflammation. Chem. Biol. Interact. 2017, 277, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, R.M.; Abdelazem, A.Z.; Hashem, K.S.; Attia, Y.A. Protective effects of hesperidin against MTX-induced hepatotoxicity in male albino rats. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 1405–1417. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Ye, Y.Y.; Ji, G.; Liu, J.W. Hesperidin upregulates heme oxygenase-1 to attenuate hydrogen peroxide-induced cell damage in hepatic L02 cells. J. Agric. Food Chem. 2010, 58, 3330–3335. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Meng, X.; Wang, F.; Bao, Y.; Huo, J. Eriodictyol attenuates arsenic trioxide-induced liver injury by activation of Nrf2. Oncotarget 2017, 8, 68668–68674. [Google Scholar] [CrossRef]
- Okkay, U.; Ferah Okkay, I.; Cicek, B.; Aydin, I.C.; Ozkaraca, M. Hepatoprotective and neuroprotective effect of taxifolin on hepatic encephalopathy in rats. Metab. Brain Dis. 2022, 37, 1541–1556. [Google Scholar] [CrossRef]
- Al-Amarat, W.; Abukhalil, M.H.; Alruhaimi, R.S.; Alqhtani, H.A.; Aldawood, N.; Alfwuaires, M.A.; Althunibat, O.Y.; Aladaileh, S.H.; Algefare, A.I.; Alanezi, A.A.; et al. Upregulation of Nrf2/HO-1 Signaling and Attenuation of Oxidative Stress, Inflammation, and Cell Death Mediate the Protective Effect of Apigenin against Cyclophosphamide Hepatotoxicity. Metabolites 2022, 12, 648. [Google Scholar] [CrossRef]
- Pan, X.; Shao, Y.; Wang, F.; Cai, Z.; Liu, S.; Xi, J.; He, R.; Zhao, Y.; Zhuang, R. Protective effect of apigenin magnesium complex on H2O2-induced oxidative stress and inflammatory responses in rat hepatic stellate cells. Pharm. Biol. 2020, 58, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.M.; Guo, W.N.; Tan, Y.Z.; Niu, K.W.; Zhang, J.J.; Liu, C.L.; Yang, X.M.; Tao, K.S.; Chen, Z.N.; Dai, J.Y. Wogonin alleviates liver injury in sepsis through Nrf2-mediated NF-kappaB signalling suppression. J. Cell Mol. Med. 2021, 25, 5782–5798. [Google Scholar] [CrossRef]
- Yang, D.; Tan, X.; Lv, Z.; Liu, B.; Baiyun, R.; Lu, J.; Zhang, Z. Regulation of Sirt1/Nrf2/TNF-alpha signaling pathway by luteolin is critical to attenuate acute mercuric chloride exposure induced hepatotoxicity. Sci. Rep. 2016, 6, 37157. [Google Scholar] [CrossRef]
- Rajput, S.A.; Shaukat, A.; Wu, K.; Rajput, I.R.; Baloch, D.M.; Akhtar, R.W.; Raza, M.A.; Najda, A.; Rafal, P.; Albrakati, A.; et al. Luteolin Alleviates AflatoxinB(1)-Induced Apoptosis and Oxidative Stress in the Liver of Mice through Activation of Nrf2 Signaling Pathway. Antioxidants 2021, 10, 1268. [Google Scholar] [CrossRef]
- Janda, E.; Martino, C.; Riillo, C.; Parafati, M.; Lascala, A.; Mollace, V.; Boutin, J.A. Apigenin and Luteolin Regulate Autophagy by Targeting NRH-Quinone Oxidoreductase 2 in Liver Cells. Antioxidants 2021, 10, 776. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Fang, Y.; Cao, W.; Zhang, Z.; Pan, S.; Xu, X. Attenuation of tert-Butyl Hydroperoxide (t-BHP)-Induced Oxidative Damage in HepG2 Cells by Tangeretin: Relevance of the Nrf2-ARE and MAPK Signaling Pathways. J. Agric. Food Chem. 2018, 66, 6317–6325. [Google Scholar] [CrossRef] [PubMed]
- Tekeli, M.Y.; Cakir Bayram, L.; Eraslan, G.; Soyer Sarica, Z. The protective effect of chrysin against oxidative stress and organ toxicity in rats exposed to propetamphos. Drug Chem. Toxicol. 2022, 45, 2664–2677. [Google Scholar] [CrossRef] [PubMed]
- El-Far, Y.M.; Khodir, A.E.; Emarah, Z.A.; Ebrahim, M.A.; Al-Gayyar, M.M.H. Chemopreventive and hepatoprotective effects of genistein via inhibition of oxidative stress and the versican/PDGF/PKC signaling pathway in experimentally induced hepatocellular carcinoma in rats by thioacetamide. Redox Rep. 2022, 27, 9–20. [Google Scholar] [CrossRef]
- Kampkotter, A.; Chovolou, Y.; Kulawik, A.; Rohrdanz, E.; Weber, N.; Proksch, P.; Watjen, W. Isoflavone daidzein possesses no antioxidant activities in cell-free assays but induces the antioxidant enzyme catalase. Nutr. Res. 2008, 28, 620–628. [Google Scholar] [CrossRef]
- Mishra, P.; Kar, A.; Kale, R.K. Prevention of chemically induced mammary tumorigenesis by daidzein in pre-pubertal rats: The role of peroxidative damage and antioxidative enzymes. Mol. Cell Biochem. 2009, 325, 149–157. [Google Scholar] [CrossRef]
- Gu, X.; Manautou, J.E. Molecular mechanisms underlying chemical liver injury. Expert. Rev. Mol. Med. 2012, 14, e4. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Hu, M. Mutual interactions between flavonoids and enzymatic and transporter elements responsible for flavonoid disposition via phase II metabolic pathways. RSC Adv. 2012, 2, 7948–7963. [Google Scholar] [CrossRef] [PubMed]
- Arlt, V.M.; Stiborova, M.; Henderson, C.J.; Thiemann, M.; Frei, E.; Aimova, D.; Singh, R.; Gamboa da Costa, G.; Schmitz, O.J.; Farmer, P.B.; et al. Metabolic activation of benzo[a]pyrene in vitro by hepatic cytochrome P450 contrasts with detoxification in vivo: Experiments with hepatic cytochrome P450 reductase null mice. Carcinogenesis 2008, 29, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Kitakaze, T.; Makiyama, A.; Nakai, R.; Kimura, Y.; Ashida, H. Kaempferol modulates TCDD- and t-BHQ-induced drug-metabolizing enzymes and luteolin enhances this effect. Food Funct. 2020, 11, 3668–3680. [Google Scholar] [CrossRef]
- Khan, T.H.; Jahangir, T.; Prasad, L.; Sultana, S. Inhibitory effect of apigenin on benzo(a)pyrene-mediated genotoxicity in Swiss albino mice. J. Pharm. Pharmacol. 2006, 58, 1655–1660. [Google Scholar] [CrossRef]
- Ueng, Y.F.; Shyu, C.C.; Lin, Y.L.; Park, S.S.; Liao, J.F.; Chen, C.F. Effects of baicalein and wogonin on drug-metabolizing enzymes in C57BL/6J mice. Life Sci. 2000, 67, 2189–2200. [Google Scholar] [CrossRef]
- Ni, Z.; Ma, H.; Li, X.; Zou, L.; Liu, Z.; Wang, X.; Ma, H.; Yang, L. Wogonin alleviates BaP-induced DNA damage and oxidative stress in human airway epithelial cells by dual inhibiting CYP1A1 activity and expression. Environ. Toxicol. 2023, 38, 2717–2729. [Google Scholar] [CrossRef]
- Uhl, M.; Ecker, S.; Kassie, F.; Lhoste, E.; Chakraborty, A.; Mohn, G.; Knasmuller, S. Effect of chrysin, a flavonoid compound, on the mutagenic activity of 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) and benzo(a)pyrene (B(a)P) in bacterial and human hepatoma (HepG2) cells. Arch. Toxicol. 2003, 77, 477–484. [Google Scholar] [CrossRef]
- Sulc, M.; Indra, R.; Moserova, M.; Schmeiser, H.H.; Frei, E.; Arlt, V.M.; Stiborova, M. The impact of individual cytochrome P450 enzymes on oxidative metabolism of benzo[a]pyrene in human livers. Environ. Mol. Mutagen. 2016, 57, 229–235. [Google Scholar] [CrossRef]
- Kamenickova, A.; Anzenbacherova, E.; Pavek, P.; Soshilov, A.A.; Denison, M.S.; Anzenbacher, P.; Dvorak, Z. Pelargonidin activates the AhR and induces CYP1A1 in primary human hepatocytes and human cancer cell lines HepG2 and LS174T. Toxicol. Lett. 2013, 218, 253–259. [Google Scholar] [CrossRef]
- Gao, S.S.; Chen, X.Y.; Zhu, R.Z.; Choi, B.M.; Kim, S.J.; Kim, B.R. Dual effects of phloretin on aflatoxin B1 metabolism: Activation and detoxification of aflatoxin B1. Biofactors 2012, 38, 34–43. [Google Scholar] [CrossRef]
- Singh, G.; Thaker, R.; Sharma, A.; Parmar, D. Therapeutic effects of biochanin A, phloretin, and epigallocatechin-3-gallate in reducing oxidative stress in arsenic-intoxicated mice. Environ. Sci. Pollut. Res. Int. 2021, 28, 20517–20536. [Google Scholar] [CrossRef] [PubMed]
- Dudka, J.; Jodynis-Liebert, J.; Korobowicz, E.; Burdan, F.; Korobowicz, A.; Szumilo, J.; Tokarska, E.; Klepacz, R.; Murias, M. Activity of NADPH-cytochrome P-450 reductase of the human heart, liver and lungs in the presence of (-)-epigallocatechin gallate, quercetin and resveratrol: An in vitro study. Basic. Clin. Pharmacol. Toxicol. 2005, 97, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Rahden-Staron, I.; Czeczot, H.; Szumilo, M. Induction of rat liver cytochrome P450 isoenzymes CYP 1A and CYP 2B by different fungicides, nitrofurans, and quercetin. Mutat. Res. 2001, 498, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.K.; Chen, J.; Yeung, E.Y. Effect of Ginkgo biloba extract on procarcinogen-bioactivating human CYP1 enzymes: Identification of isorhamnetin, kaempferol, and quercetin as potent inhibitors of CYP1B1. Toxicol. Appl. Pharmacol. 2006, 213, 18–26. [Google Scholar] [CrossRef]
- BinMowyna, M.N.; AlFaris, N.A. Kaempferol suppresses acetaminophen-induced liver damage by upregulation/activation of SIRT1. Pharm. Biol. 2021, 59, 146–156. [Google Scholar] [CrossRef]
- Karakurt, S. Modulatory effects of rutin on the expression of cytochrome P450s and antioxidant enzymes in human hepatoma cells. Acta Pharm. 2016, 66, 491–502. [Google Scholar] [CrossRef]
- Kravchenko, L.V.; Avren’eva, L.I.; Aksenov, I.V.; Balakina, A.S.; Guseva, G.V.; Trusov, N.V. Effects of rutin on protective capacity in rats. Vopr. Pitan. 2015, 84, 22–30. [Google Scholar]
- Khan, R.A.; Khan, M.R.; Sahreen, S. CCl4-induced hepatotoxicity: Protective effect of rutin on p53, CYP2E1 and the antioxidative status in rat. BMC Complement. Altern. Med. 2012, 12, 178. [Google Scholar] [CrossRef]
- Bhatt, S.; Manhas, D.; Kumar, V.; Gour, A.; Sharma, K.; Dogra, A.; Ojha, P.K.; Nandi, U. Effect of Myricetin on CYP2C8 Inhibition to Assess the Likelihood of Drug Interaction Using In Silico, In Vitro, and In Vivo Approaches. ACS Omega 2022, 7, 13260–13269. [Google Scholar] [CrossRef]
- Wang, Z.; Lan, Y.; Chen, M.; Wen, C.; Hu, Y.; Liu, Z.; Ye, L. Eriodictyol, Not Its Glucuronide Metabolites, Attenuates Acetaminophen-Induced Hepatotoxicity. Mol. Pharm. 2017, 14, 2937–2951. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Lee, J.; Kim, S.; Yeo, Y.Y.; So, H.; Wu, H.; Song, Y.S.; Jang, C.Y.; Kim, H.D.; Kim, M.J.; et al. Hepatic Metabolism of Sakuranetin and Its Modulating Effects on Cytochrome P450s and UDP-Glucuronosyltransferases. Molecules 2018, 23, 1542. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Ye, J.; Zhao, L.; Li, X.; Wang, Y.; Liu, X.; Pan, L.; You, L.; Chen, L.; Jia, Y.; et al. 5,7,3′,4′-flavan-on-ol (taxifolin) protects against acetaminophen-induced liver injury by regulating the glutathione pathway. Life Sci. 2019, 236, 116939. [Google Scholar] [CrossRef] [PubMed]
- Steuck, M.; Hellhake, S.; Schebb, N.H. Food Polyphenol Apigenin Inhibits the Cytochrome P450 Monoxygenase Branch of the Arachidonic Acid Cascade. J. Agric. Food Chem. 2016, 64, 8973–8976. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Kwara, A.; Greenblatt, D.J. Metabolic interactions between acetaminophen (paracetamol) and two flavonoids, luteolin and quercetin, through in-vitro inhibition studies. J. Pharm. Pharmacol. 2017, 69, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Francisco, V.; Figueirinha, A.; Costa, G.; Liberal, J.; Ferreira, I.; Lopes, M.C.; Garcia-Rodriguez, C.; Cruz, M.T.; Batista, M.T. The Flavone Luteolin Inhibits Liver X Receptor Activation. J. Nat. Prod. 2016, 79, 1423–1428. [Google Scholar] [CrossRef]
- Lakshmi, A.; Subramanian, S. Chemotherapeutic effect of tangeretin, a polymethoxylated flavone studied in 7, 12-dimethylbenz(a)anthracene induced mammary carcinoma in experimental rats. Biochimie 2014, 99, 96–109. [Google Scholar] [CrossRef]
- Chlouchi, A.; Girard, C.; Bonet, A.; Viollon-Abadie, C.; Heyd, B.; Mantion, G.; Martin, H.; Richert, L. Effect of chrysin and natural coumarins on UGT1A1 and 1A6 activities in rat and human hepatocytes in primary culture. Planta Med. 2007, 73, 742–747. [Google Scholar] [CrossRef]
- Soliman, M.M.; Aldhahrani, A.; Gaber, A.; Alsanie, W.F.; Mohamed, W.A.; Metwally, M.M.M.; Elbadawy, M.; Shukry, M. Ameliorative impacts of chrysin against gibberellic acid-induced liver and kidney damage through the regulation of antioxidants, oxidative stress, inflammatory cytokines, and apoptosis biomarkers. Toxicol. Res. 2022, 11, 235–244. [Google Scholar] [CrossRef]
- Steiner, C.; Peters, W.H.; Gallagher, E.P.; Magee, P.; Rowland, I.; Pool-Zobel, B.L. Genistein protects human mammary epithelial cells from benzo(a)pyrene-7,8-dihydrodiol-9,10-epoxide and 4-hydroxy-2-nonenal genotoxicity by modulating the glutathione/glutathione S-transferase system. Carcinogenesis 2007, 28, 738–748. [Google Scholar] [CrossRef]
- Froyen, E.B.; Steinberg, F.M. Genistein decreases basal hepatic cytochrome P450 1A1 protein expression and activity in Swiss Webster mice. Nutr. Res. 2016, 36, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Froyen, E.B.; Reeves, J.L.; Mitchell, A.E.; Steinberg, F.M. Regulation of phase II enzymes by genistein and daidzein in male and female Swiss Webster mice. J. Med. Food 2009, 12, 1227–1237. [Google Scholar] [CrossRef]
- Wiegand, H.; Wagner, A.E.; Boesch-Saadatmandi, C.; Kruse, H.P.; Kulling, S.; Rimbach, G. Effect of dietary genistein on Phase II and antioxidant enzymes in rat liver. Cancer Genom. Proteom. 2009, 6, 85–92. [Google Scholar]
- Atherton, K.M.; Mutch, E.; Ford, D. Metabolism of the soyabean isoflavone daidzein by CYP1A2 and the extra-hepatic CYPs 1A1 and 1B1 affects biological activity. Biochem. Pharmacol. 2006, 72, 624–631. [Google Scholar] [CrossRef]
- Wang, Y.; Pan, T.; Li, L.; Wang, H.; Zhang, D.; Yang, H. Benzo(a)pyrene promotes Hep-G2 cell migration and invasion by upregulating phosphorylated extracellular signal-regulated kinase expression. Oncol. Lett. 2018, 15, 8325–8332. [Google Scholar] [CrossRef]
- Ba, Q.; Li, J.; Huang, C.; Qiu, H.; Li, J.; Chu, R.; Zhang, W.; Xie, D.; Wu, Y.; Wang, H. Effects of benzo[a]pyrene exposure on human hepatocellular carcinoma cell angiogenesis, metastasis, and NF-kappaB signaling. Environ. Health Perspect. 2015, 123, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Asha, A.; Girija, D. Benzo[a]pyrene induced liver and kidney cancer in swiss albino mice. Res. Pharm. 2015, 1, 21. [Google Scholar]
- Ge, Y.; Gu, P.; Wang, W.; Cao, L.; Zhang, L.; Li, J.; Mu, W.; Wang, H. Benzo[a]pyrene stimulates miR-650 expression to promote the pathogenesis of fatty liver disease and hepatocellular carcinoma via SOCS3/JAK/STAT3 cascades. J. Mol. Cell Biol. 2021, 13, 556–564. [Google Scholar] [CrossRef]
- Bucher, S.; Tete, A.; Podechard, N.; Liamin, M.; Le Guillou, D.; Chevanne, M.; Coulouarn, C.; Imran, M.; Gallais, I.; Fernier, M.; et al. Co-exposure to benzo[a]pyrene and ethanol induces a pathological progression of liver steatosis in vitro and in vivo. Sci. Rep. 2018, 8, 5963. [Google Scholar] [CrossRef]
- Bucher, S.; Le Guillou, D.; Allard, J.; Pinon, G.; Begriche, K.; Tete, A.; Sergent, O.; Lagadic-Gossmann, D.; Fromenty, B. Possible Involvement of Mitochondrial Dysfunction and Oxidative Stress in a Cellular Model of NAFLD Progression Induced by Benzo[a]pyrene/Ethanol CoExposure. Oxid. Med. Cell Longev. 2018, 2018, 4396403. [Google Scholar] [CrossRef]
- Collin, A.; Hardonniere, K.; Chevanne, M.; Vuillemin, J.; Podechard, N.; Burel, A.; Dimanche-Boitrel, M.T.; Lagadic-Gossmann, D.; Sergent, O. Cooperative interaction of benzo[a]pyrene and ethanol on plasma membrane remodeling is responsible for enhanced oxidative stress and cell death in primary rat hepatocytes. Free Radic. Biol. Med. 2014, 72, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Michurina, S.V.; Borodin Iu, I.; Kolesnikov, S.I.; Ischenko, I.; Konenkov, V.I. Liver and Its Lymph Region at Benzo[a]pyrene Effects in an Experiment. Vestn. Ross. Akad. Med. Nauk. 2015, 70, 242–248. [Google Scholar] [CrossRef]
- Kolade, O.Y.; Oladiji, T.A. Protective Effects of Curcumin against Benzopyrene Induced Liver Toxicity in Albino Rats. Iop C Ser. Earth Env. 2018, 210, 012013. [Google Scholar] [CrossRef]
- Li, H.; Yuan, L.; Li, X.; Luo, Y.; Zhang, Z.; Li, J. Isoorientin Attenuated the Pyroptotic Hepatocyte Damage Induced by Benzo[a]pyrene via ROS/NF-kappaB/NLRP3/Caspase-1 Signaling Pathway. Antioxidants 2021, 10, 1275. [Google Scholar] [CrossRef]
- Li, X.Y.; He, S.Y.; Gao, C.X.; Deng, H.; Liu, Y.F.; Li, C.Q.; Yuan, L.; Luo, Y. Isoorientin attenuates benzo[a]pyrene-induced liver injury by inhibiting autophagy and pyroptosis in vitro and vivo. Food Agric. Immunol. 2019, 30, 841–861. [Google Scholar] [CrossRef]
- Li, H.; Yuan, L.; Wang, Z.L.; Shi, L.; Dong, R.; Hu, X.Z. Effects of dietary whole grain buckwheat and oat on benzo[a] pyrene-induced genotoxicity, oxidative and pyroptotic injury in liver of mice. J. Funct. Foods 2022, 93, 105082. [Google Scholar] [CrossRef]
- Hernandez-Aquino, E.; Muriel, P. Beneficial effects of naringenin in liver diseases: Molecular mechanisms. World J. Gastroenterol. 2018, 24, 1679–1707. [Google Scholar] [CrossRef] [PubMed]
- Plazar, J.; Zegura, B.; Lah, T.T.; Filipic, M. Protective effects of xanthohumol against the genotoxicity of benzo(a)pyrene (BaP), 2-amino-3-methylimidazo[4,5-f]quinoline (IQ) and tert-butyl hydroperoxide (t-BOOH) in HepG2 human hepatoma cells. Mutat. Res. 2007, 632, 1–8. [Google Scholar] [CrossRef]
- Wen, X.; Walle, U.K.; Walle, T. 5,7-Dimethoxyflavone downregulates CYP1A1 expression and benzo[a]pyrene-induced DNA binding in Hep G2 cells. Carcinogenesis 2005, 26, 803–809. [Google Scholar] [CrossRef]
- Sehgal, A.; Kumar, M.; Jain, M.; Dhawan, D.K. Piperine as an adjuvant increases the efficacy of curcumin in mitigating benzo(a)pyrene toxicity. Hum. Exp. Toxicol. 2012, 31, 473–482. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.; Jee, S.-C.; Sung, J.-S. Hepatoprotective Effects of Flavonoids against Benzo[a]Pyrene-Induced Oxidative Liver Damage along Its Metabolic Pathways. Antioxidants 2024, 13, 180. https://doi.org/10.3390/antiox13020180
Kim M, Jee S-C, Sung J-S. Hepatoprotective Effects of Flavonoids against Benzo[a]Pyrene-Induced Oxidative Liver Damage along Its Metabolic Pathways. Antioxidants. 2024; 13(2):180. https://doi.org/10.3390/antiox13020180
Chicago/Turabian StyleKim, Min, Seung-Cheol Jee, and Jung-Suk Sung. 2024. "Hepatoprotective Effects of Flavonoids against Benzo[a]Pyrene-Induced Oxidative Liver Damage along Its Metabolic Pathways" Antioxidants 13, no. 2: 180. https://doi.org/10.3390/antiox13020180