
Methanethiol: A Scent Mark of Dysregulated Sulfur Metabolism in Cancer
 , , , and
, , , and 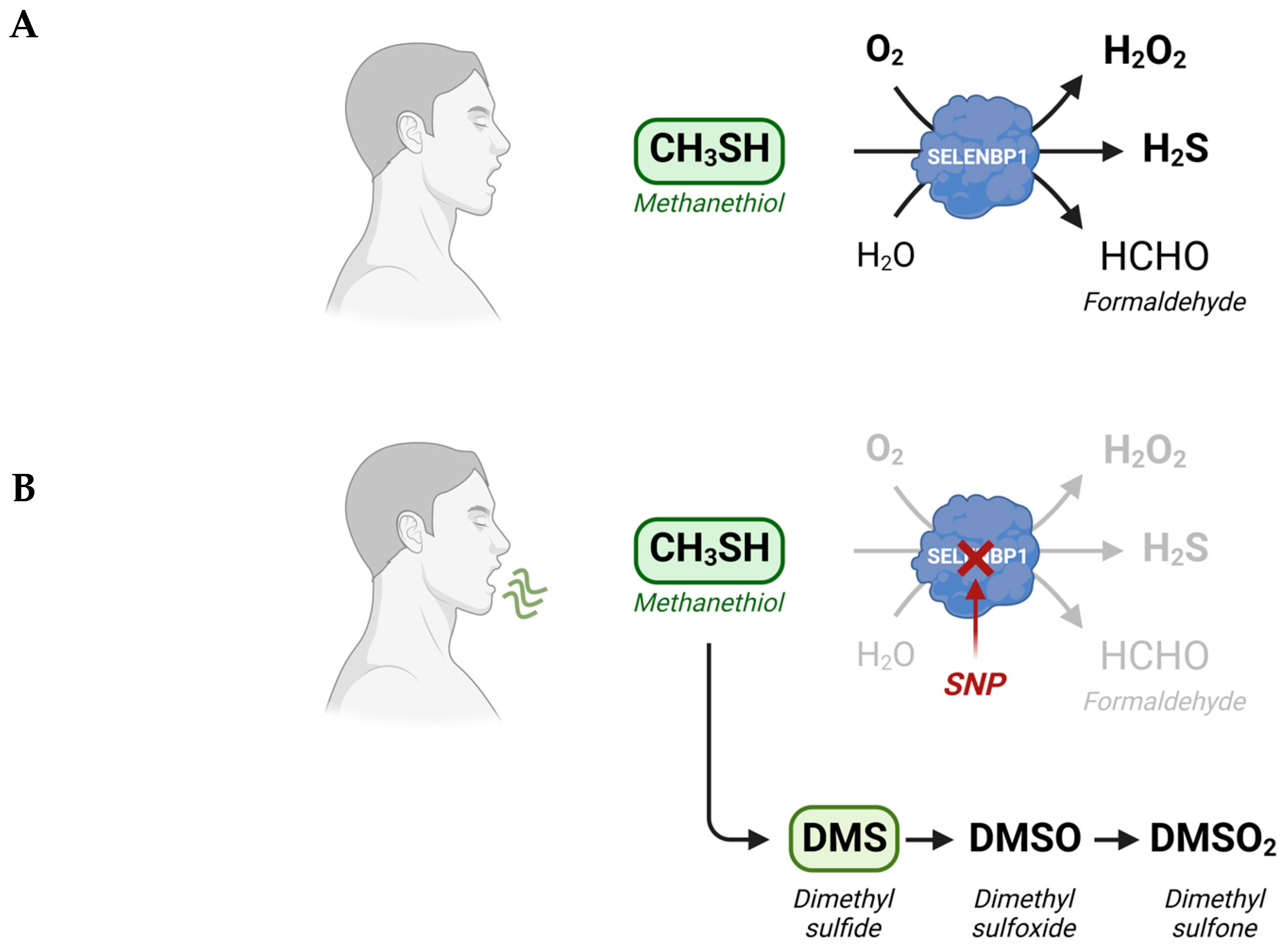{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
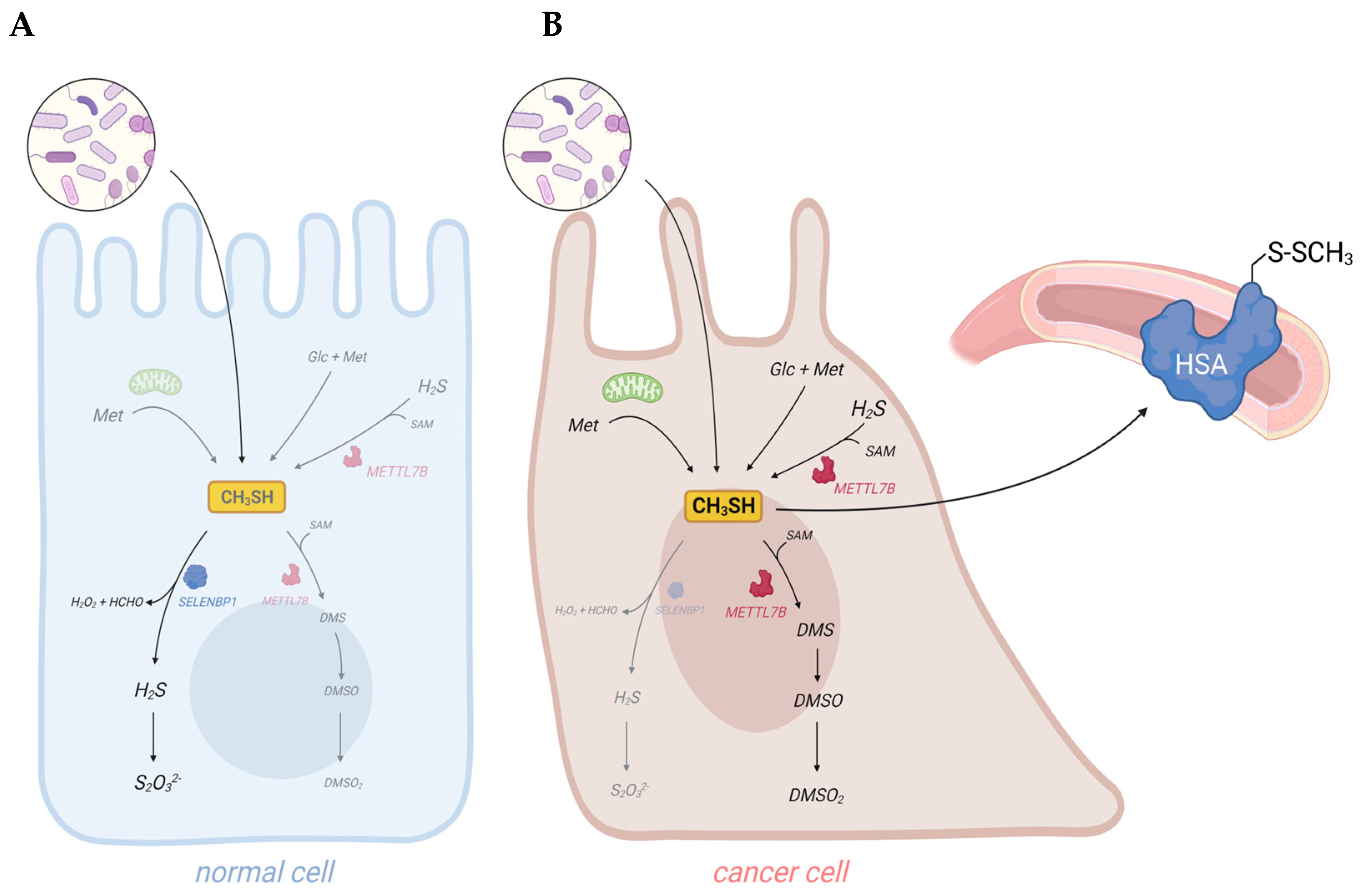
2. Exogenous and Endogenous Methanethiol Production in Humans
3. SELENBP1-Catalyzed Degradation of Methanethiol in Humans
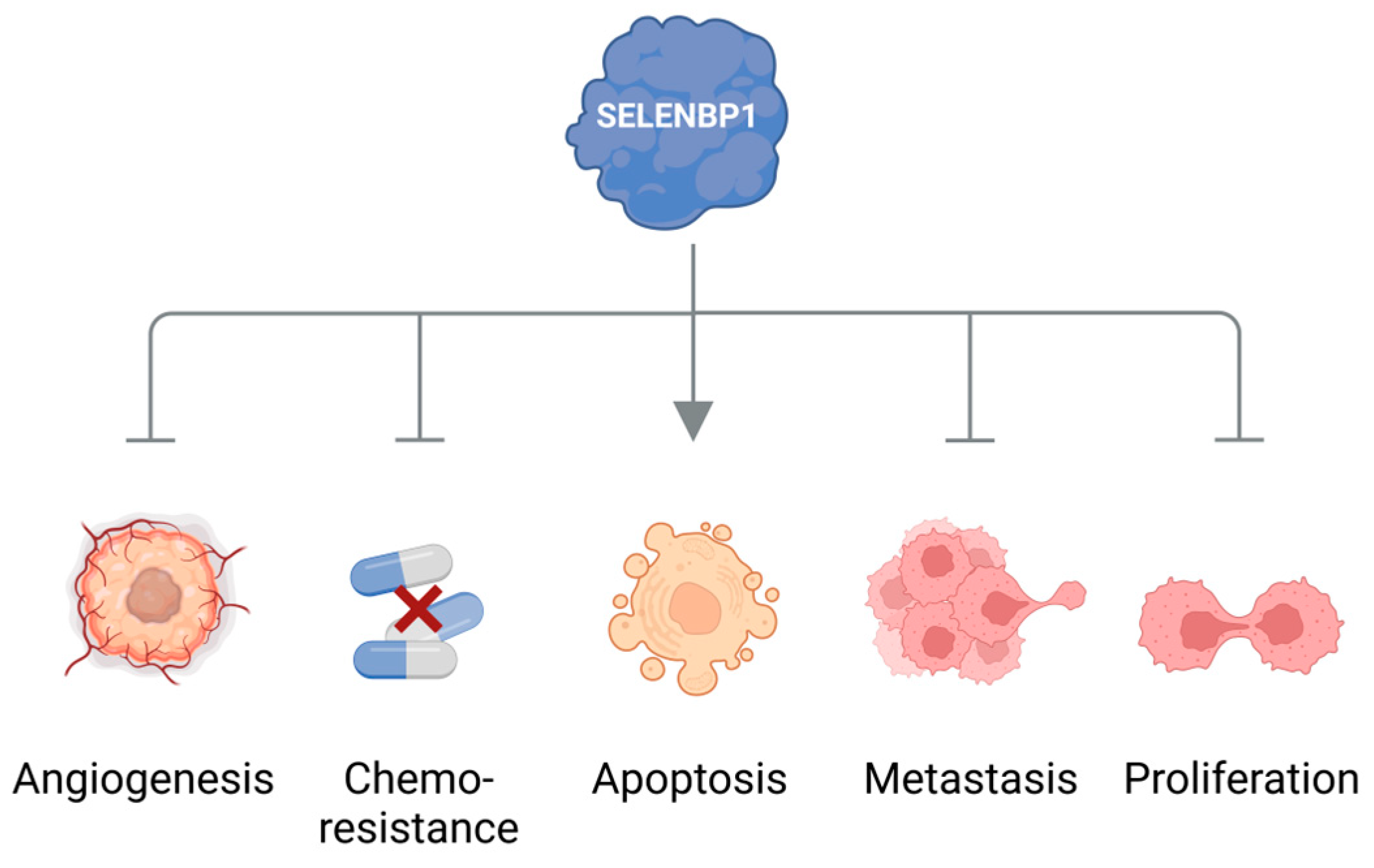
4. Elevated Levels of Methanethiol and Its Derivatives in Various Types of Cancer
5. Conclusions and Outlook: Methanethiol as a Promising Biomarker for Non-Invasive Cancer Diagnosis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Koppenol, W.H. The warburg effect—Discovered 100 years ago. Free Radic. Biol. Med. 2023, 204, 325. [Google Scholar] [CrossRef]
- Warburg, O.; Minami, S. Versuche an überlebendem carcinom-gewebe. Klin. Wochenschr. 1923, 2, 776–777. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Chandel, N.S. Fundamentals of cancer metabolism. Sci. Adv. 2016, 2, e1600200. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Reyes, I.; Chandel, N.S. Cancer metabolism: Looking forward. Nat. Rev. Cancer 2021, 21, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Sedillo, J.C.; Cryns, V.L. Targeting the methionine addiction of cancer. Am. J. Cancer Res. 2022, 12, 2249–2276. [Google Scholar]
- Bian, Y.; Li, W.; Kremer, D.M.; Sajjakulnukit, P.; Li, S.; Crespo, J.; Nwosu, Z.C.; Zhang, L.; Czerwonka, A.; Pawlowska, A.; et al. Cancer slc43a2 alters t cell methionine metabolism and histone methylation. Nature 2020, 585, 277–282. [Google Scholar] [CrossRef]
- Scheller, A.S.; Philipp, T.M.; Klotz, L.O.; Steinbrenner, H. Altered capacity for h(2)s production during the spontaneous differentiation of caco-2 cells to colonocytes due to reciprocal regulation of cbs and selenbp1. Antioxidants 2022, 11, 1957. [Google Scholar] [CrossRef]
- Shackelford, R.E.; Mohammad, I.Z.; Meram, A.T.; Kim, D.; Alotaibi, F.; Patel, S.; Ghali, G.E.; Kevil, C.G. Molecular functions of hydrogen sulfide in cancer. Pathophysiology 2021, 28, 437–456. [Google Scholar] [CrossRef]
- Amal, H.; Ding, L.; Liu, B.B.; Tisch, U.; Xu, Z.Q.; Shi, D.Y.; Zhao, Y.; Chen, J.; Sun, R.X.; Liu, H.; et al. The scent fingerprint of hepatocarcinoma: In-vitro metastasis prediction with volatile organic compounds (vocs). Int. J. Nanomed. 2012, 7, 4135–4146. [Google Scholar]
- Bhatt, A.; Parsi, M.A.; Stevens, T.; Gabbard, S.; Kumaravel, A.; Jang, S.; Grove, D.; Lopez, R.; Murthy, S.; Vargo, J.J.; et al. Volatile organic compounds in plasma for the diagnosis of esophageal adenocarcinoma: A pilot study. Gastrointest. Endosc. 2016, 84, 597–603. [Google Scholar] [CrossRef]
- Ishibe, A.; Ota, M.; Takeshita, A.; Tsuboi, H.; Kizuka, S.; Oka, H.; Suwa, Y.; Suzuki, S.; Nakagawa, K.; Suwa, H.; et al. Detection of gas components as a novel diagnostic method for colorectal cancer. Ann. Gastroenterol. Surg. 2018, 2, 147–153. [Google Scholar] [CrossRef]
- Xie, X.; Yu, W.; Chen, Z.; Wang, L.; Yang, J.; Liu, S.; Li, L.; Li, Y.; Huang, Y. Early-stage oral cancer diagnosis by artificial intelligence-based sers using ag nws@zif core-shell nanochains. Nanoscale 2023, 15, 13466–13472. [Google Scholar] [CrossRef]
- Ascencao, K.; Szabo, C. Emerging roles of cystathionine beta-synthase in various forms of cancer. Redox Biol. 2022, 53, 102331. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Yu, Y.; Zhu, L.; Lai, N.; Zhang, L.; Guo, Y.; Lin, X.; Yang, D.; Ren, N.; Zhu, Z.; et al. Implications of hydrogen sulfide in colorectal cancer: Mechanistic insights and diagnostic and therapeutic strategies. Redox Biol. 2023, 59, 102601. [Google Scholar] [CrossRef] [PubMed]
- Kabil, O.; Banerjee, R. Enzymology of h2s biogenesis, decay and signaling. Antioxid. Redox Signal. 2014, 20, 770–782. [Google Scholar] [CrossRef]
- Pol, A.; Renkema, G.H.; Tangerman, A.; Winkel, E.G.; Engelke, U.F.; de Brouwer, A.P.M.; Lloyd, K.C.; Araiza, R.S.; van den Heuvel, L.; Omran, H.; et al. Mutations in selenbp1, encoding a novel human methanethiol oxidase, cause extraoral halitosis. Nat. Genet. 2018, 50, 120–129. [Google Scholar] [CrossRef]
- Elhodaky, M.; Diamond, A.M. Selenium-binding protein 1 in human health and disease. Int. J. Mol. Sci. 2018, 19, 3437. [Google Scholar] [CrossRef]
- Schafer, H.; Eyice, O. Microbial cycling of methanethiol. Curr. Issues Mol. Biol. 2019, 33, 173–182. [Google Scholar] [CrossRef]
- Tansy, M.F.; Kendall, F.M.; Fantasia, J.; Landin, W.E.; Oberly, R.; Sherman, W. Acute and subchronic toxicity studies of rats exposed to vapors of methyl mercaptan and other reduced-sulfur compounds. J. Toxicol. Environ. Health 1981, 8, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Vahlkamp, T.; Meijer, A.J.; Wilms, J.; Chamuleau, R.A. Inhibition of mitochondrial electron transfer in rats by ethanethiol and methanethiol. Clin. Sci. 1979, 56, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Suarez, F.L.; Springfield, J.; Levitt, M.D. Identification of gases responsible for the odour of human flatus and evaluation of a device purported to reduce this odour. Gut 1998, 43, 100–104. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Slupsky, C.M. Metabolic fingerprint of dimethyl sulfone (dmso2) in microbial-mammalian co-metabolism. J. Proteome Res. 2014, 13, 5281–5292. [Google Scholar] [CrossRef] [PubMed]
- Sato, D.; Nozaki, T. Methionine gamma-lyase: The unique reaction mechanism, physiological roles, and therapeutic applications against infectious diseases and cancers. IUBMB Life 2009, 61, 1019–1028. [Google Scholar] [CrossRef]
- Maldonato, B.J.; Russell, D.A.; Totah, R.A. Human mettl7b is an alkyl thiol methyltransferase that metabolizes hydrogen sulfide and captopril. Sci. Rep. 2021, 11, 4857. [Google Scholar] [CrossRef]
- Scislowski, P.W.; Pickard, K. The regulation of transaminative flux of methionine in rat liver mitochondria. Arch. Biochem. Biophys. 1994, 314, 412–416. [Google Scholar] [CrossRef]
- Finkelstein, A.; Benevenga, N.J. The effect of methanethiol and methionine toxicity on the activities of cytochrome c oxidase and enzymes involved in protection from peroxidative damage. J. Nutr. 1986, 116, 204–215. [Google Scholar] [CrossRef]
- Furne, J.; Springfield, J.; Koenig, T.; DeMaster, E.; Levitt, M.D. Oxidation of hydrogen sulfide and methanethiol to thiosulfate by rat tissues: A specialized function of the colonic mucosa. Biochem. Pharmacol. 2001, 62, 255–259. [Google Scholar] [CrossRef]
- Levitt, M.D.; Furne, J.; Springfield, J.; Suarez, F.; DeMaster, E. Detoxification of hydrogen sulfide and methanethiol in the cecal mucosa. J. Clin. Investig. 1999, 104, 1107–1114. [Google Scholar] [CrossRef]
- Landry, A.P.; Ballou, D.P.; Banerjee, R. Hydrogen sulfide oxidation by sulfide quinone oxidoreductase. Chembiochem 2021, 22, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Philipp, T.M.; Gernoth, L.; Will, A.; Schwarz, M.; Ohse, V.A.; Kipp, A.P.; Steinbrenner, H.; Klotz, L.O. Selenium-binding protein 1 (selenbp1) is a copper-dependent thiol oxidase. Redox Biol. 2023, 65, 102807. [Google Scholar] [CrossRef] [PubMed]
- Philipp, T.M.; Gong, W.; Kohnlein, K.; Ohse, V.A.; Muller, F.I.; Priebs, J.; Steinbrenner, H.; Klotz, L.O. Semo-1, a novel methanethiol oxidase in caenorhabditis elegans, is a pro-aging factor conferring selective stress resistance. Biofactors 2022, 48, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Eyice, O.; Myronova, N.; Pol, A.; Carrion, O.; Todd, J.D.; Smith, T.J.; Gurman, S.J.; Cuthbertson, A.; Mazard, S.; Mennink-Kersten, M.A.; et al. Bacterial sbp56 identified as a cu-dependent methanethiol oxidase widely distributed in the biosphere. ISME J. 2018, 12, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Kohnlein, K.; Urban, N.; Guerrero-Gomez, D.; Steinbrenner, H.; Urbanek, P.; Priebs, J.; Koch, P.; Kaether, C.; Miranda-Vizuete, A.; Klotz, L.O. A caenorhabditis elegans ortholog of human selenium-binding protein 1 is a pro-aging factor protecting against selenite toxicity. Redox Biol. 2020, 28, 101323. [Google Scholar] [CrossRef]
- Braga, C.P.; Vieira, J.C.S.; Grove, R.A.; Boone, C.H.T.; Leite, A.L.; Buzalaf, M.A.R.; Fernandes, A.A.H.; Adamec, J.; Padilha, P.M. A proteomic approach to identify metalloproteins and metal-binding proteins in liver from diabetic rats. Int. J. Biol. Macromol. 2017, 96, 817–832. [Google Scholar] [CrossRef]
- Schild, F.; Kieffer-Jaquinod, S.; Palencia, A.; Cobessi, D.; Sarret, G.; Zubieta, C.; Jourdain, A.; Dumas, R.; Forge, V.; Testemale, D.; et al. Biochemical and biophysical characterization of the selenium-binding and reducing site in arabidopsis thaliana homologue to mammals selenium-binding protein 1. J. Biol. Chem. 2014, 289, 31765–31776. [Google Scholar] [CrossRef]
- Li, T.; Yang, W.; Li, M.; Byun, D.S.; Tong, C.; Nasser, S.; Zhuang, M.; Arango, D.; Mariadason, J.M.; Augenlicht, L.H. Expression of selenium-binding protein 1 characterizes intestinal cell maturation and predicts survival for patients with colorectal cancer. Mol. Nutr. Food Res. 2008, 52, 1289–1299. [Google Scholar] [CrossRef]
- Philipp, T.M.; Will, A.; Richter, H.; Winterhalter, P.R.; Pohnert, G.; Steinbrenner, H.; Klotz, L.O. A coupled enzyme assay for detection of selenium-binding protein 1 (selenbp1) methanethiol oxidase (mto) activity in mature enterocytes. Redox Biol. 2021, 43, 101972. [Google Scholar] [CrossRef]
- Funato, K.; Abe, T.; Kurita, R.; Watanabe, Y.; Nakamura, Y.; Miyata, S.; Furukawa, Y.; Satake, M. Identification of characteristic proteins at late-stage erythroid differentiation in vitro. Hum. Cell 2021, 34, 745–749. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Micoogullari, M.; Hoang, N.A.; Bergheim, I.; Klotz, L.O.; Sies, H. Selenium-binding protein 1 (selenbp1) is a marker of mature adipocytes. Redox Biol. 2019, 20, 489–495. [Google Scholar] [CrossRef]
- Ringrose, J.H.; van Solinge, W.W.; Mohammed, S.; O’Flaherty, M.C.; van Wijk, R.; Heck, A.J.; Slijper, M. Highly efficient depletion strategy for the two most abundant erythrocyte soluble proteins improves proteome coverage dramatically. J. Proteome Res. 2008, 7, 3060–3063. [Google Scholar] [CrossRef] [PubMed]
- Randi, E.B.; Casili, G.; Jacquemai, S.; Szabo, C. Selenium-binding protein 1 (selenbp1) supports hydrogen sulfide biosynthesis and adipogenesis. Antioxidants 2021, 10, 361. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical biology of h(2)s signaling through persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Goubern, M.; Andriamihaja, M.; Nubel, T.; Blachier, F.; Bouillaud, F. Sulfide, the first inorganic substrate for human cells. FASEB J. 2007, 21, 1699–1706. [Google Scholar] [CrossRef]
- McClain, C.J.; Zieve, L.; Doizaki, W.M.; Gilberstadt, S.; Onstad, G.R. Blood methanethiol in alcoholic liver disease with and without hepatic encephalopathy. Gut 1980, 21, 318–323. [Google Scholar] [CrossRef]
- Stephen, A.S.; Dhadwal, N.; Nagala, V.; Gonzales-Marin, C.; Gillam, D.G.; Bradshaw, D.J.; Burnett, G.R.; Allaker, R.P. Interdental and subgingival microbiota may affect the tongue microbial ecology and oral malodour in health, gingivitis and periodontitis. J. Periodontal Res. 2021, 56, 1174–1184. [Google Scholar] [CrossRef]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Qin, H. Microbiota disbiosis is associated with colorectal cancer. Front. Microbiol. 2015, 6, 20. [Google Scholar] [CrossRef]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef]
- Yamagishi, K.; Onuma, K.; Chiba, Y.; Yagi, S.; Aoki, S.; Sato, T.; Sugawara, Y.; Hosoya, N.; Saeki, Y.; Takahashi, M.; et al. Generation of gaseous sulfur-containing compounds in tumour tissue and suppression of gas diffusion as an antitumour treatment. Gut 2012, 61, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Kwon, I.J.; Jung, T.Y.; Son, Y.; Kim, B.; Kim, S.M.; Lee, J.H. Detection of volatile sulfur compounds (vscs) in exhaled breath as a potential diagnostic method for oral squamous cell carcinoma. BMC Oral Health 2022, 22, 268. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Yang, Y.; Buettner, R.; Rosen, S.T. Targeting the methionine-methionine adenosyl transferase 2a- s -adenosyl methionine axis for cancer therapy. Curr. Opin. Oncol. 2022, 34, 546–551. [Google Scholar] [CrossRef]
- Ye, D.; Jiang, Y.; Sun, Y.; Li, Y.; Cai, Y.; Wang, Q.; Wang, O.; Chen, E.; Zhang, X. Mettl7b promotes migration and invasion in thyroid cancer through epithelial-mesenchymal transition. J. Mol. Endocrinol. 2019, 63, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Caswell, D.R.; Chuang, C.H.; Ma, R.K.; Winters, I.P.; Snyder, E.L.; Winslow, M.M. Tumor suppressor activity of selenbp1, a direct nkx2-1 target, in lung adenocarcinoma. Mol. Cancer Res. 2018, 16, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Ying, Q.; Ansong, E.; Diamond, A.M.; Lu, Z.; Yang, W.; Bie, X. Quantitative proteomic analysis reveals that anti-cancer effects of selenium-binding protein 1 in vivo are associated with metabolic pathways. PLoS ONE 2015, 10, e0126285. [Google Scholar] [CrossRef]
- Zeng, H.; Zhao, X.; Tang, C. Downregulation of selenbp1 enhances oral squamous cell carcinoma chemoresistance through keap1-nrf2 signaling. Cancer Chemother. Pharmacol. 2021, 88, 223–233. [Google Scholar] [CrossRef]
- Zhang, X.; Hong, R.; Bei, L.; Yang, J.; Zhao, X.; Hu, Z.; Chen, L.; Meng, H.; Zhang, Q.; Niu, G.; et al. Selenium binding protein 1 inhibits tumor angiogenesis in colorectal cancers by blocking the delta-like ligand 4/notch1 signaling pathway. Transl. Oncol. 2022, 18, 101365. [Google Scholar] [CrossRef]
- Zhu, Y.; Pu, Q.; Zhang, Q.; Liu, Y.; Ma, Y.; Yuan, Y.; Liu, L.; Zhu, W. Selenium-binding protein 1 inhibits malignant progression and induces apoptosis via distinct mechanisms in non-small-cell lung cancer. Cancer Med. 2023, 12, 17149–17170. [Google Scholar] [CrossRef]
- Buszewski, B.; Ulanowska, A.; Kowalkowski, T.; Cieslinski, K. Investigation of lung cancer biomarkers by hyphenated separation techniques and chemometrics. Clin. Chem. Lab. Med. 2011, 50, 573–581. [Google Scholar] [CrossRef]
- Miller-Atkins, G.; Acevedo-Moreno, L.A.; Grove, D.; Dweik, R.A.; Tonelli, A.R.; Brown, J.M.; Allende, D.S.; Aucejo, F.; Rotroff, D.M. Breath metabolomics provides an accurate and noninvasive approach for screening cirrhosis, primary, and secondary liver tumors. Hepatol. Commun. 2020, 4, 1041–1055. [Google Scholar] [CrossRef]
- Li, Z.; Shu, J.; Zhang, P.; Sun, W.; Yang, B.; Zhang, H. Real-time ultrasensitive vuv-pims detection of representative endogenous volatile markers in cancers. Cancer Biomark. 2016, 16, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.; Gallagher, M.; Ozdener, M.H.; Wysocki, C.J.; Goldsmith, B.R.; Isamah, A.; Faranda, A.; Fakharzadeh, S.S.; Herlyn, M.; Johnson, A.T.; et al. Volatile biomarkers from human melanoma cells. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2013, 931, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Kybert, N.; Prokop-Prigge, K.; Otto, C.M.; Ramirez, L.; Joffe, E.; Tanyi, J.; Piltz-Seymour, J.; Johnson, A.T.C.; Preti, G. Exploring ovarian cancer screening using a combined sensor approach: A pilot study. AIP Adv. 2020, 10, 035213. [Google Scholar] [CrossRef]
- Skorupa, A.; Ponski, M.; Ciszek, M.; Cichon, B.; Klimek, M.; Witek, A.; Pakulo, S.; Boguszewicz, L.; Sokol, M. Grading of endometrial cancer using (1)h hr-mas nmr-based metabolomics. Sci. Rep. 2021, 11, 18160. [Google Scholar] [CrossRef] [PubMed]
- Sever, A.; Abd Elkadir, A.; Matana, Y.; Gopas, J.; Zeiri, Y. Biomarkers for detection and monitoring of b16 melanoma in mouse urine and feces. J. Biomark. 2015, 2015, 841245. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, H.; Schiffman, C.; Gunter, M.J.; Naccarati, A.; Polidoro, S.; Dagnino, S.; Dudoit, S.; Vineis, P.; Rappaport, S.M. Cys34 adductomics links colorectal cancer with the gut microbiota and redox biology. Cancer Res. 2019, 79, 6024–6031. [Google Scholar] [CrossRef]
- Shirasu, M.; Nagai, S.; Hayashi, R.; Ochiai, A.; Touhara, K. Dimethyl trisulfide as a characteristic odor associated with fungating cancer wounds. Biosci. Biotechnol. Biochem. 2009, 73, 2117–2120. [Google Scholar] [CrossRef]
- Shirasu, M.; Touhara, K. The scent of disease: Volatile organic compounds of the human body related to disease and disorder. J. Biochem. 2011, 150, 257–266. [Google Scholar] [CrossRef]
- Haick, H.; Broza, Y.Y.; Mochalski, P.; Ruzsanyi, V.; Amann, A. Assessment, origin, and implementation of breath volatile cancer markers. Chem. Soc. Rev. 2014, 43, 1423–1449. [Google Scholar] [CrossRef]
- Janssens, E.; van Meerbeeck, J.P.; Lamote, K. Volatile organic compounds in human matrices as lung cancer biomarkers: A systematic review. Crit. Rev. Oncol. Hematol. 2020, 153, 103037. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Xiong, X.; Hao, T.; Zhang, C.; Gao, Z.; Zhong, L.; Zhao, Y. Volatile organic compounds as a potential screening tool for neoplasm of the digestive system: A meta-analysis. Sci. Rep. 2021, 11, 23716. [Google Scholar] [CrossRef]
- Jenkins, E.K.; DeChant, M.T.; Perry, E.B. When the nose doesn’t know: Canine olfactory function associated with health, management, and potential links to microbiota. Front. Vet. Sci. 2018, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Kokocinska-Kusiak, A.; Woszczylo, M.; Zybala, M.; Maciocha, J.; Barlowska, K.; Dzieciol, M. Canine olfaction: Physiology, behavior, and possibilities for practical applications. Animals 2021, 11, 2463. [Google Scholar] [CrossRef] [PubMed]
- Feil, C.; Staib, F.; Berger, M.R.; Stein, T.; Schmidtmann, I.; Forster, A.; Schimanski, C.C. Sniffer dogs can identify lung cancer patients from breath and urine samples. BMC Cancer 2021, 21, 917. [Google Scholar] [CrossRef]
- Kure, S.; Iida, S.; Yamada, M.; Takei, H.; Yamashita, N.; Sato, Y.; Miyashita, M. Breast cancer detection from a urine sample by dog sniffing: A preliminary study for the development of a new screening device, and a literature review. Biology 2021, 10, 517. [Google Scholar] [CrossRef]
- Murarka, M.; Vesley-Gross, Z.I.; Essler, J.L.; Smith, P.G.; Hooda, J.; Drapkin, R.; Otto, C.M. Testing ovarian cancer cell lines to train dogs to detect ovarian cancer from blood plasma: A pilot study. J. Vet. Behav. 2019, 32, 42–48. [Google Scholar] [CrossRef]
- Sonoda, H.; Kohnoe, S.; Yamazato, T.; Satoh, Y.; Morizono, G.; Shikata, K.; Morita, M.; Watanabe, A.; Morita, M.; Kakeji, Y.; et al. Colorectal cancer screening with odour material by canine scent detection. Gut 2011, 60, 814–819. [Google Scholar] [CrossRef]
- Taverna, G.; Tidu, L.; Grizzi, F.; Torri, V.; Mandressi, A.; Sardella, P.; La Torre, G.; Cocciolone, G.; Seveso, M.; Giusti, G.; et al. Olfactory system of highly trained dogs detects prostate cancer in urine samples. J. Urol. 2015, 193, 1382–1387. [Google Scholar] [CrossRef]
- Rudnicka, J.; Walczak, M.; Kowalkowski, T.; Jezierski, T.; Buszewski, B. Determination of volatile organic compounds as potential markers of lung cancer by gas chromatography-mass spectrometry versus trained dogs. Sens. Actuators B-Chem. 2014, 202, 615–621. [Google Scholar] [CrossRef]
- Schroeder, W. Volatile s-nitrosothiols and the typical smell of cancer. J. Breath Res. 2015, 9, 016010. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Philipp, T.M.; Scheller, A.S.; Krafczyk, N.; Klotz, L.-O.; Steinbrenner, H. Methanethiol: A Scent Mark of Dysregulated Sulfur Metabolism in Cancer. Antioxidants 2023, 12, 1780. https://doi.org/10.3390/antiox12091780
Philipp TM, Scheller AS, Krafczyk N, Klotz L-O, Steinbrenner H. Methanethiol: A Scent Mark of Dysregulated Sulfur Metabolism in Cancer. Antioxidants. 2023; 12(9):1780. https://doi.org/10.3390/antiox12091780
Chicago/Turabian StylePhilipp, Thilo Magnus, Anne Sophie Scheller, Niklas Krafczyk, Lars-Oliver Klotz, and Holger Steinbrenner. 2023. "Methanethiol: A Scent Mark of Dysregulated Sulfur Metabolism in Cancer" Antioxidants 12, no. 9: 1780. https://doi.org/10.3390/antiox12091780