
Effect of Purple Sweet Potato Using Different Cooking Methods on Cytoprotection against Ethanol-Induced Oxidative Damage through Nrf2 Activation in HepG2 Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Chemicals
2.2. Cooking Methods
2.3. Sample Preparation for Cell-Based Assay
2.4. LC-QTOF/MS Analysis
2.5. Cytotoxicity
2.6. Measurement of Intracellular ROS
2.7. Measurement of Glutathione and Antioxidant Enzyme Activities
2.8. Western Blotting Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. Cytoprotective Effects of Purple Sweet Potato with Different Cooking Methods against Alcohol Damage in Hepatocytes
3.2. Antioxidative Effects of Purple Sweet Potato with Different Cooking Methods against Alcohol-Induced Damage
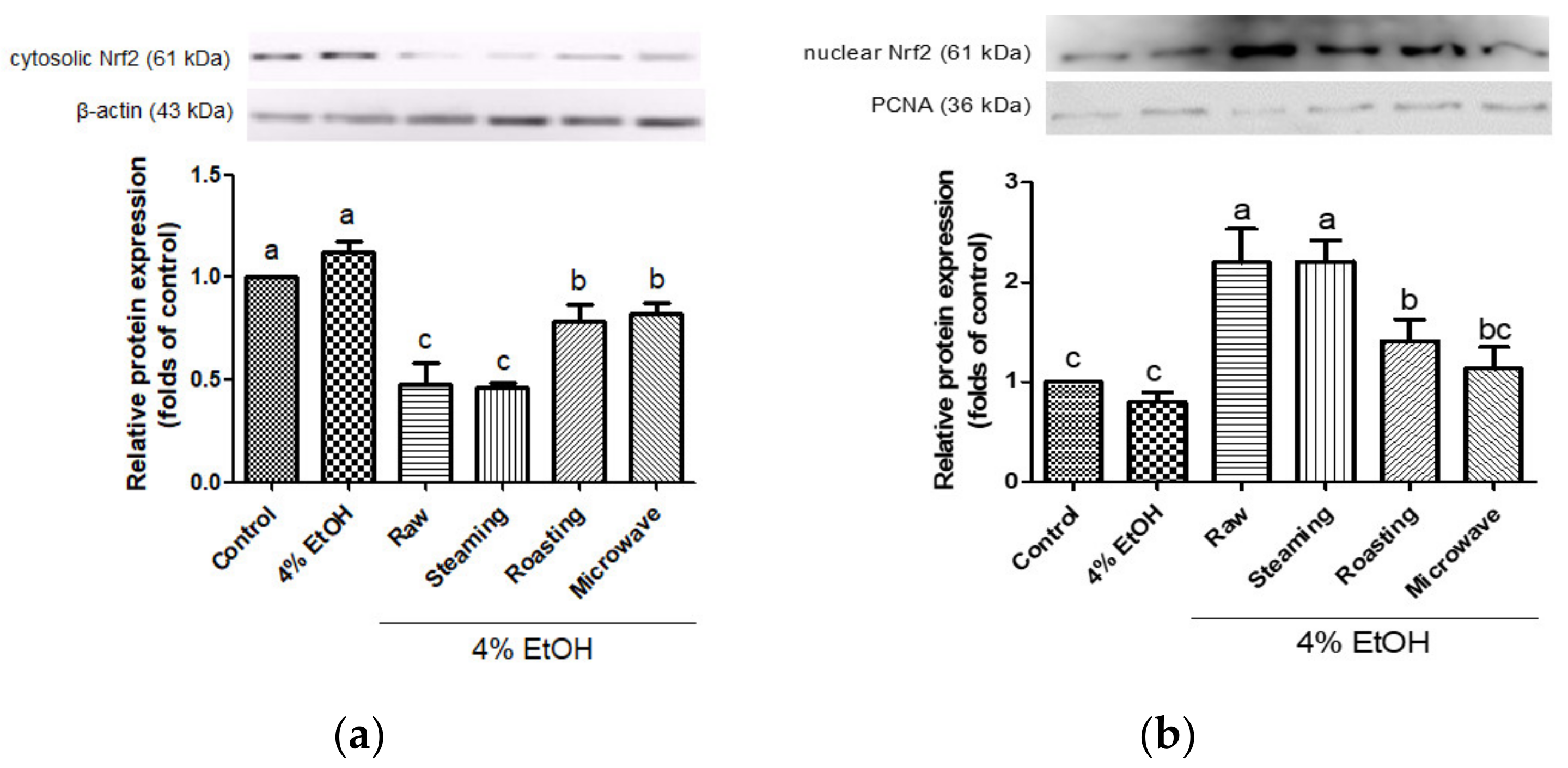
3.3. Effects of Purple Sweet Potato with Different Cooking Methods on the Expressions of Antioxidant Enzymes and the Activation of Nrf2
3.4. Identification of Bioactive Compounds in Purple Sweet Potato with Different Cooking Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, B.; Bataller, R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Beier, J.I.; McClain, C.J. Mechanisms and cell signaling in alcoholic liver disease. Biol. Chem. 2010, 391, 1249–1264. [Google Scholar] [CrossRef] [PubMed]
- Abdelmegeed, M.A.; Banerjee, A.; Jang, S.; Yoo, S.H.; Yun, J.W.; Gonzalez, F.J.; Keshavarzian, A.; Song, B.J. CYP2E1 potentiates binge alcohol-induced gut leakiness, steatohepatitis, and apoptosis. Free Radic. Biol. Med. 2013, 65, 1238–1245. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Cederbaum, A.I. Overexpression of CYP2E1 in mitochondria sensitizes HepG2 cells to the toxicity caused by depletion of glutathione. J. Biol. Chem. 2006, 281, 5128–5136. [Google Scholar] [CrossRef] [PubMed]
- Dai, N.; Zou, Y.; Zhu, L.; Wang, H.F.; Dai, M.G. Antioxidant properties of proanthocyanidins attenuate carbon tetrachloride (CCl4)–induced steatosis and liver injury in rats via CYP2E1 regulation. J. Med. Food 2014, 17, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, B. Teaching the basics of redox biology to medical and graduate students: Oxidants, antioxidants and disease mechanisms. Redox Biol. 2013, 1, 244–257. [Google Scholar] [CrossRef]
- Liu, Z.; Ren, Z.; Zhang, J.; Chuang, C.C.; Kandaswamy, E.; Zhou, T.; Zuo, L. Role of ROS and nutritional antioxidants in human diseases. Front. Physiol. 2018, 9, 477. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Pullikotil, P.; Chen, H.; Muniyappa, R.; Greenberg, C.C.; Yang, S.; Reiter, C.E.; Lee, J.W.; Chung, J.H.; Quon, M.J. Epigallocatechin gallate induces expression of heme oxygenase-1 in endothelial cells via p38 MAPK and Nrf-2 that suppresses proinflammatory actions of TNF-alpha. J. Nutr. Biochem. 2012, 23, 1134–1145. [Google Scholar] [CrossRef]
- He, M.; Siow, R.C.; Sugden, D.; Gao, L.; Cheng, X.; Mann, G.E. Induction of HO-1 and redox signaling in endothelial cells by advanced glycation end products: A role for Nrf2 in vascular protection in diabetes. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 277–285. [Google Scholar] [CrossRef]
- Xu, D.; Xu, M.; Jeong, S.; Qian, Y.; Wu, H.; Xia, Q.; Kong, X. The role of Nrf2 in liver disease: Novel molecular mechanisms and therapeutic approaches. Front. Pharmacol. 2018, 9, 1428. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zheng, Q.; Chen, Z. The Nrf2 pathway in liver diseases. Front. Cell Dev. Biol. 2022, 10, 826204. [Google Scholar] [CrossRef] [PubMed]
- Medoua, G.N.; Oldewage-Theron, W.H. Effect of drying and cooking on nutritional value and antioxidant capacity of morogo (Amaranthus hybridus) a traditional leafy vegetable grown in South Africa. J. Food Sci. Technol. 2014, 51, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Medina-Remon, A.; Perez-Jimenez, J.; Neveu, V.; Knaze, V.; Slimani, N.; Scalbert, A. Effects of food processing on polyphenol contents: A systematic analysis using Phenol-Explorer data. Mol. Nutr. Food Res. 2015, 59, 160–170. [Google Scholar] [CrossRef] [PubMed]
- D’Amelia, V.; Sarais, G.; Fais, G.; Dessi, D.; Giannini, V.; Garramone, R.; Carputo, D.; Melito, S. Biochemical characterization and effects of cooking methods on main phytochemicals of red and purple potato tubers, a natural functional food. Foods 2022, 11, 384. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G. Effect of blanching on the antioxidant properties of some tropical green leafy vegetables. LWT—Food Sci. Technol. 2005, 38, 513–517. [Google Scholar] [CrossRef]
- Ayeleso, T.B.; Ramachela, K.; Mukwevho, E. A review of therapeutic potentials of sweet potato: Pharmacological activities and influence of the cultivar. Trop. J. Pharm. Res. 2017, 15, 2751–2761. [Google Scholar] [CrossRef]
- Im, Y.R.; Kim, I.; Lee, J. Phenolic composition and antioxidant activity of purple sweet potato (Ipomoea batatas (L.) Lam.): Varietal comparisons and physical distribution. Antioxidants 2021, 10, 462. [Google Scholar] [CrossRef]
- Wu, Q.; Qu, H.; Jia, J.; Kuang, C.; Wen, Y.; Yan, H.; Gui, Z. Characterization, antioxidant and antitumor activities of polysaccharides from purple sweet potato. Carbohydr. Polym. 2015, 132, 31–40. [Google Scholar] [CrossRef]
- Zhao, J.G.; Yan, Q.Q.; Lu, L.Z.; Zhang, Y.Q. In vivo antioxidant, hypoglycemic, and anti-tumor activities of anthocyanin extracts from purple sweet potato. Nutr. Res. Pract. 2013, 7, 359–365. [Google Scholar] [CrossRef]
- Liao, M.; Zou, B.; Chen, J.; Yao, Z.; Huang, L.; Luo, Z.; Wang, Z. Effect of domestic cooking methods on the anthocyanins and antioxidant activity of deeply purple-fleshed sweet potato GZ9. Heliyon 2019, 5, e01515. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Park, J.S.; Choi, D.S.; Jung, M.Y. Characterization and quantitation of anthocyanins in purple-fleshed sweet potatoes cultivated in Korea by HPLC-DAD and HPLC-ESI-QTOF-MS/MS. J. Agric. Food Chem. 2013, 61, 3148–3158. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Kim, Y.; Sung, J.; Lee, H.; Heo, H.; Jeong, H.S.; Lee, J. Jujube (Ziziphus jujuba Mill.) Protects hepatocytes against alcohol-induced damage through Nrf2 activation. Evid. Based Complement. Alternat. Med. 2020, 2020, 6684331. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Choi, Y.; Ham, H.; Jeong, H.S.; Lee, J. Protective effects of oligomeric and polymeric procyanidin fractions from defatted grape seeds on tert-butyl hydroperoxide-induced oxidative damage in HepG2 cells. Food Chem. 2013, 137, 136–141. [Google Scholar] [CrossRef]
- Baker, M.A.; Cerniglia, G.J.; Zaman, A. Microtiter plate assay for the measurement of glutathione and glutathione disulfide in large numbers of biological samples. Anal. Biochem. 1990, 190, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Ukeda, H.; Kawana, D.; Maeda, S.; Sawamura, M. Spectrophotometric assay for superoxide dismutase based on the reduction of highly water-soluble tetrazolium salts by xanthine–xanthine oxidase. Biosci. Biotechnol. Biochem. 1999, 63, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Fossati, P.; Prencipe, L.; Berti, G. Use of 3,5-dichloro-2-hydroxybenzenesulfonic acid/4-aminophenazone chromogenic system in direct enzyme assay of uric acid in serum and urine. Clin. Biochem. 1980, 26, 227–231. [Google Scholar] [CrossRef]
- Smith, I.K.; Vierheller, T.L.; Thorne, C.A. Assay of glutathione reductase in crude tissue homogenates using 5,50-dithiobis(2-nitrobenzoic acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Flohe, L.; Gunzler, W.A. Assays of glutathione peroxidase. Meth. Enzymol. 1984, 105, 114–121. [Google Scholar] [CrossRef]
- Xiao, T.; Luo, Z.; Guo, Z.; Wang, X.; Ding, M.; Wang, W.; Shen, X.; Zhao, Y. Multiple roles of black raspberry anthocyanins protecting against alcoholic liver disease. Molecules 2021, 26, 2313. [Google Scholar] [CrossRef]
- Sun, H.; Mu, T.; Liu, X.; Zhang, M.; Chen, J. Purple sweet potato (Ipomoea batatas L.) anthocyanins: Preventive effect on acute and subacute alcoholic liver damage and dealcoholic effect. J. Agric. Food. Chem. 2014, 62, 2364–2373. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, S.Y.; Yang, J.W.; Lee, J.S.; Oh, S.D.; Oh, S.; Lee, S.M.; Lim, M.H.; Park, S.K.; Jang, J.S.; et al. Comparative analysis of phytochemicals and polar metabolites from colored sweet potato (Ipomoea batatas L.) tubers. Food Sci. Biotechnol. 2016, 25, 283–291. [Google Scholar] [CrossRef]
- Cevallos-Casals, B.v.A.; Cisneros-Zevallos, L. Stability of anthocyanin-based aqueous extracts of Andean purple corn and red-fleshed sweet potato compared to synthetic and natural colorants. Food Chem. 2004, 86, 69–77. [Google Scholar] [CrossRef]
- Hwang, Y.P.; Choi, J.H.; Yun, H.J.; Han, E.H.; Kim, H.G.; Kim, J.Y.; Park, B.H.; Khanal, T.; Choi, J.M.; Chung, Y.C.; et al. Anthocyanins from purple sweet potato attenuate dimethylnitrosamine-induced liver injury in rats by inducing Nrf2-mediated antioxidant enzymes and reducing COX-2 and iNOS expression. Food Chem. Toxicol. 2011, 49, 93–99. [Google Scholar] [CrossRef]
- Olszowy-Tomczyk, M.; Wianowska, D. Antioxidant properties of selected flavonoids in binary mixtures—Considerations on myricetin, kaempferol and quercetin. Int. J. Mol. Sci. 2023, 24, 10070. [Google Scholar] [CrossRef]
- Rodgers, E.H.; Grant, M.H. The effect of the flavonoids, quercetin, myricetin and epicatechin on the growth and enzyme activities of MCF7 human breast cancer cells. Chem. Biol. Interact. 1998, 116, 213–228. [Google Scholar] [CrossRef]
- Galicia-Moreno, M.; Gutierrez-Reyes, G. The role of oxidative stress in the development of alcoholic liver disease. Rev. Gastroenterol. Mex. 2014, 79, 135–144. [Google Scholar] [CrossRef]
- Tang, Y.; Cai, W.; Xu, B. Profiles of phenolics, carotenoids and antioxidative capacities of thermal processed white, yellow, orange and purple sweet potatoes grown in Guilin, China. Food Sci. Hum. Wellness 2015, 4, 123–132. [Google Scholar] [CrossRef]
- Dolinsky, M.; Agostinho, C.; Ribeiro, D.; Rocha, G.D.S.; Barroso, S.G.; Ferreira, D.; Polinati, R.; Ciarelli, G.; Fialho, E. Effect of different cooking methods on the polyphenol concentration and antioxidant capacity of selected vegetables. J. Culin. Sci. Technol. 2015, 14, 1–12. [Google Scholar] [CrossRef]
- Lee, K.; Lee, H.; Choi, Y.; Kim, Y.; Jeong, H.S.; Lee, J. Effect of different cooking methods on the true retention of vitamins, minerals, and bioactive compounds in Shiitake mushrooms (Lentinula edodes). Food Sci. Technol. Res. 2019, 25, 115–122. [Google Scholar] [CrossRef]
- Hwang, Y.P.; Choi, J.H.; Han, E.H.; Kim, H.K.; Kang, S.K.; Chung, Y.C.; Jeong, H.G. Protective mechanisms of Aralia continentalis extract against tert-butyl hydroperoxide-induced hepatotoxicity: In vivo and in vitro studies. Food Chem. Toxicol. 2008, 46, 3512–3521. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guan, L.; Wang, X.; Wen, T.; Xing, J.; Zhao, J. Protection of chlorophyllin against oxidative damage by inducing HO-1 and NQO1 expression mediated by PI3K/Akt and Nrf2. Free Radic. Res. 2008, 42, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Norris, K.M.; Okie, W.; Yakaitis, C.L.; Pazdro, R. The anthocyanin cyanidin-3-O-beta-glucoside modulates murine glutathione homeostasis in a manner dependent on genetic background. Redox Biol. 2016, 9, 254–263. [Google Scholar] [CrossRef]
- Weisel, T.; Baum, M.; Eisenbrand, G.; Dietrich, H.; Will, F.; Stockis, J.P.; Kulling, S.; Rufer, C.; Johannes, C.; Janzowski, C. An anthocyanin/polyphenolic-rich fruit juice reduces oxidative DNA damage and increases glutathione level in healthy probands. Biotechnol. J. 2006, 1, 388–397. [Google Scholar] [CrossRef]
- Tian, B.; Zhao, J.; Xie, X.; Chen, T.; Yin, Y.; Zhai, R.; Wang, X.; An, W.; Li, J. Anthocyanins from the fruits of Lycium ruthenicum Murray improve high-fat diet-induced insulin resistance by ameliorating inflammation and oxidative stress in mice. Food Funct. 2021, 12, 3855–3871. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, P.; Unni, S.; Krishnappa, G.; Padmanabhan, B. The Keap1-Nrf2 pathway: Promising therapeutic target to counteract ROS-mediated damage in cancers and neurodegenerative diseases. Biophys. Rev. 2017, 9, 41–56. [Google Scholar] [CrossRef]
- Liu, S.; Hou, W.; Yao, P.; Li, N.; Zhang, B.; Hao, L.; Nussler, A.K.; Liu, L. Heme oxygenase-1 mediates the protective role of quercetin against ethanol-induced rat hepatocytes oxidative damage. Toxicol. In Vitro 2012, 26, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Sukprasansap, M.; Chanvorachote, P.; Tencomnao, T. Cyanidin-3-glucoside activates Nrf2-antioxidant response element and protects against glutamate-induced oxidative and endoplasmic reticulum stress in HT22 hippocampal neuronal cells. BMC Complement. Med. Ther. 2020, 20, 46. [Google Scholar] [CrossRef]
- Jin, M.; Feng, H.; Wang, Y.; Yan, S.; Shen, B.; Li, Z.; Qin, H.; Wang, Q.; Li, J.; Liu, G. Gentiopicroside ameliorates oxidative stress and lipid accumulation through nuclear factor erythroid 2-related factor 2 activation. Oxid. Med. Cell Longev. 2020, 2020, 2940746. [Google Scholar] [CrossRef]
- Jirattanarangsri, W. The effect of traditional thermal cooking processes on anthocyanin, total phenolic content, antioxidant activity and glycemic index in purple waxy corn. Food Appl. Biosci. J. 2018, 6, 154–166. [Google Scholar] [CrossRef]
- Jiménez, N.; Bassama, J.; Bohuon, P. Estimation of the kinetic parameters of anthocyanins degradation at different water activities during treatments at high temperature (100–140 °C) using an unsteady-state 3D model. J. Food Eng. 2020, 279, 109951. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | GR (1) | GPx (2) | CAT (3) | SOD (4) | |
|---|---|---|---|---|---|
| Control | 8.132 ± 1.027 ab | 25.150 ± 2.911 a | 53.718±1.135 c | 53.977±1.138 bc | |
| 4% Ethanol | 4.472 ± 0.422 d | 9.847 ± 0.769 d | 45.323 ± 2.491 d | 25.030 ± 3.758 d | |
| +4% Ethanol | Raw | 8.899 ± 0.745 a | 21.051 ± 1.818 b | 82.618 ± 3.072 a | 89.892 ± 14.354 a |
| Steaming | 7.861 ± 1.075 ab | 20.373 ± 0.211 bc | 77.386 ± 3.302 a | 85.988 ± 14.293 a | |
| Roasting | 6.603 ± 1.011 bc | 19.132 ± 1.400 bc | 64.606 ± 5.398 b | 64.257 ± 9.080 b | |
| Microwave | 5.469 ± 0.451 cd | 18.207 ± 0.740 c | 68.246 ± 2.732 b | 40.143 ± 1.188 cd | |
| Compound Class | Ion Mode | Rt (min) (1) | m/z Calculated (2) | m/z Observed (3) | Error (ppm) | Fragment Ions (m/z) | Molecular Formula | Compound Name | Observed Sample (4) |
|---|---|---|---|---|---|---|---|---|---|
| Anthocyanin | POS | 4.073 | 773.21 | 773.21 | −0.77 | 611/449/287 | C33H41O21 | cyanidin-3-sophoroside-5-glucoside | R, S, RO, M |
| 4.083 | 787.23 | 787.23 | −1.01 | 625/459/301 | C34H43O21 | peonidin-3-sophoroside-5-glucoside | R, S, RO, M | ||
| 14.20 | 893.24 | 893.24 | 0.11 | 731/449/287 | C40H45O23 | cyanidin 3-P-hydroxybenzoyl sophoroside-5-glucoside | R | ||
| 15.98 | 907.25 | 907.25 | −0.55 | 745/463/301 | C41H47O23 | peonidin 3-P-hydroxybenzoyl sophoroside-5-glucoside | R | ||
| 18.51 | 935.25 | 935.2463 | 1.18 | 773/449/287 | C42H47O24 | cyanidin 3-(6′′′-caffeoyl sophoroside)-5-glucoside | R | ||
| 18.67 | 1097.28 | 1097.28 | −0.78 | 935/449/287 | C51H53O27 | cyanidin 3-dicaffeoyl sophoroside-5-glucoside | R, S | ||
| 18.87 | 1055.27 | 1055.27 | 1.33 | 898/449/287 | C49H51O26 | cyanidin 3-dicaffeoyl sophoroside-p-hydroxybenzoyl sophoroside-5-glucoside | R | ||
| 19.99 | 949.26 | 949.26 | 1.07 | 787/463/301 | C43H49O24 | peonidin 3-(6′′′-caffeoyl sophoroside)-5-glucoside | R, S | ||
| 20.56 | 1111.29 | 1111.29 | −0.33 | 949/463/301 | C52H55O27 | peonidin 3-dicaffeoyl sophoroside-5-glucoside | R, S | ||
| 20.61 | 1069.28 | 1069.28 | 2.01 | 907/463/301 | C50H53O26 | peonidin 3-caffeoyl sophoroside-p-hydroxybenzoyl sophoroside-5-glucoside | R | ||
| Phenolic compound | POS | 5.24 | 162.03 | 162.03 | −0.58 | 133/117 | C9H6O3 | 7-hydroxycoumarin | S |
| NEG | 6.30 | 516.13 | 516.13 | −3.37 | 353 | C25H24O12 | 3,4-di-O-caffeoylquinic acid | R, S, RO, M | |
| 5.23 | 354.10 | 354.09 | −3.18 | 191 | C16H18O9 | 3-caffeoylquinic acid | R, S, RO, M |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Kim, Y.; Kim, Y. Effect of Purple Sweet Potato Using Different Cooking Methods on Cytoprotection against Ethanol-Induced Oxidative Damage through Nrf2 Activation in HepG2 Cells. Antioxidants 2023, 12, 1650. https://doi.org/10.3390/antiox12081650
Kim D, Kim Y, Kim Y. Effect of Purple Sweet Potato Using Different Cooking Methods on Cytoprotection against Ethanol-Induced Oxidative Damage through Nrf2 Activation in HepG2 Cells. Antioxidants. 2023; 12(8):1650. https://doi.org/10.3390/antiox12081650
Chicago/Turabian StyleKim, Dagyeong, Yoonjeong Kim, and Younghwa Kim. 2023. "Effect of Purple Sweet Potato Using Different Cooking Methods on Cytoprotection against Ethanol-Induced Oxidative Damage through Nrf2 Activation in HepG2 Cells" Antioxidants 12, no. 8: 1650. https://doi.org/10.3390/antiox12081650