
A Critical Review of Kaempferol in Intestinal Health and Diseases

Abstract
:1. Introduction
2. An Overview of Kaempferol
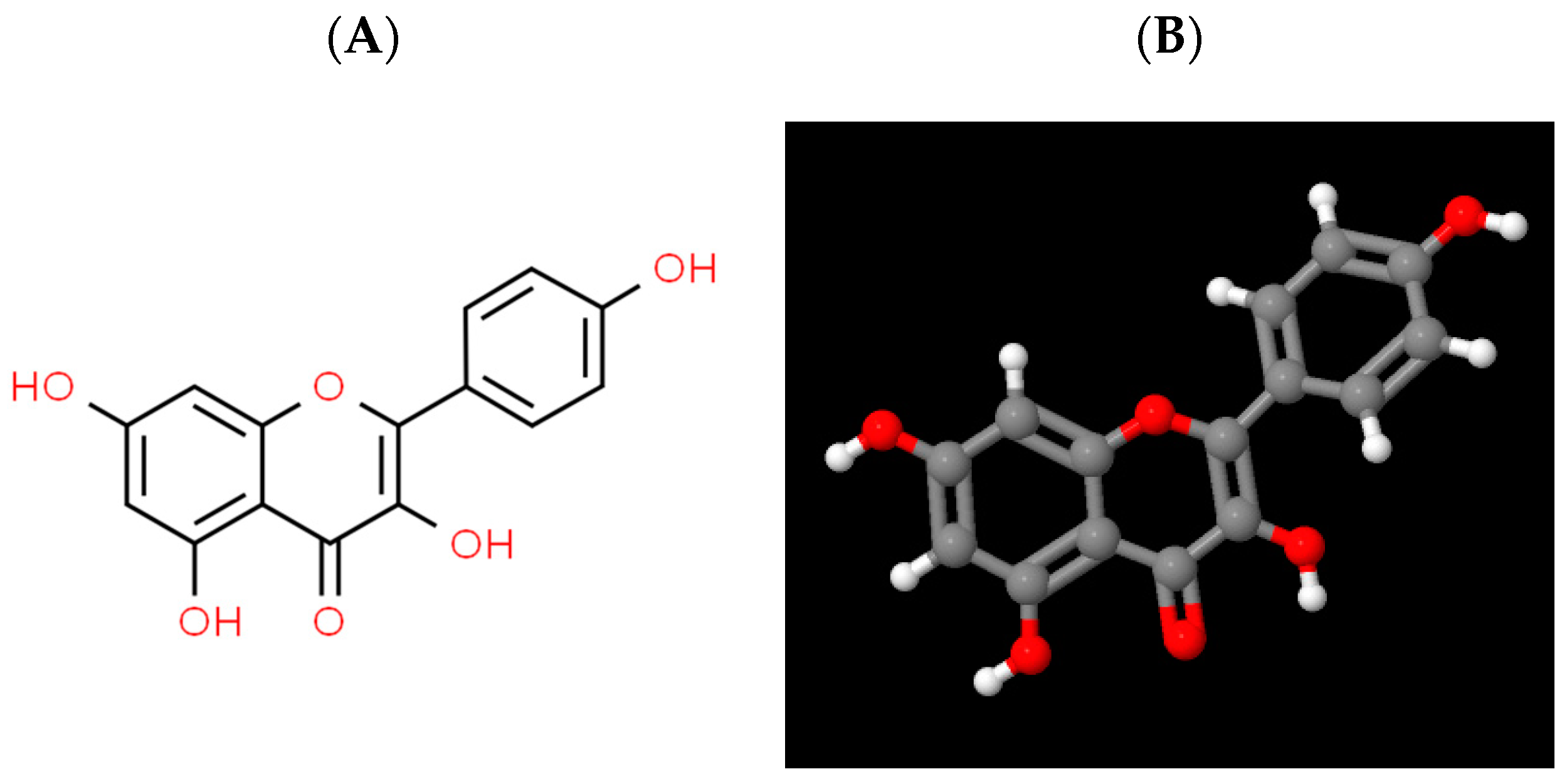
2.1. Physico-Chemical Properties of Kaempferol
2.2. Sources of Kaempferol
2.3. Absorption and Metabolism of Kaempferol
2.4. Biological Functions of Kaempferol
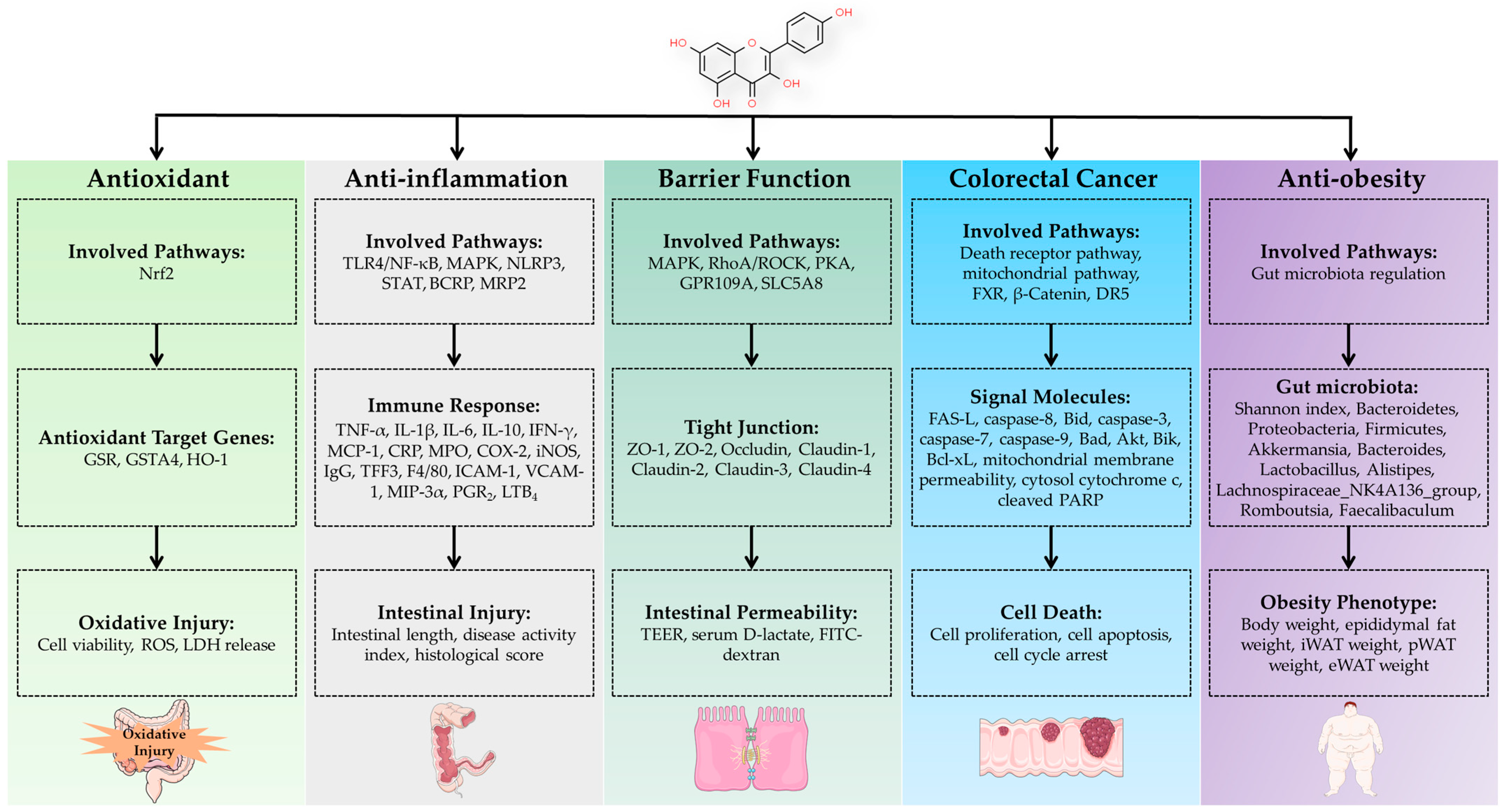
3. Regulation of Intestinal Health by Kaempferol
3.1. Regulation of Intestinal Antioxidant Function
3.2. Regulation of Intestinal Inflammation
3.3. Regulation of Intestinal Barrier function
3.4. Treatment of Colorectal Cancer
3.5. Anti-Obesity via Regulation of Gut Microbiota
3.6. Regulation of Other Physiological Functions
4. Future Research Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, Z.; Wu, C. Reciprocal interactions between regulatory T cells and intestinal epithelial cells. Front. Immunol. 2022, 13, 951339. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Rivera, L.R.; Cho, H.-J.; Bravo, D.M.; Callaghan, B. The gut as a sensory organ. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Calleja-Conde, J.; Echeverry-Alzate, V.; Bühler, K.-M.; Durán-González, P.; Morales-García, J.Á.; Segovia-Rodríguez, L.; Rodríguez de Fonseca, F.; Giné, E.; López-Moreno, J.A. The immune system through the lens of alcohol intake and gut microbiota. Int. J. Mol. Sci. 2021, 22, 7485. [Google Scholar] [CrossRef]
- Adolph, T.E.; Meyer, M.; Schwärzler, J.; Mayr, L.; Grabherr, F.; Tilg, H. The metabolic nature of inflammatory bowel diseases. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Wang, Z.; Zhang, T.; Chen, X.; Xu, H.; Wang, F.; Guo, L.; Chen, M.; Liu, K.; Wu, B. Deficiency of intestinal Bmal1 prevents obesity induced by high-fat feeding. Nat. Commun. 2021, 12, 5323. [Google Scholar] [CrossRef]
- Kang, Y.; Kang, X.; Yang, H.; Liu, H.; Yang, X.; Liu, Q.; Tian, H.; Xue, Y.; Ren, P.; Kuang, X.; et al. Lactobacillus acidophilus ameliorates obesity in mice through modulation of gut microbiota dysbiosis and intestinal permeability. Pharmacol. Res. 2022, 175, 106020. [Google Scholar] [CrossRef]
- Aa, L.-X.; Fei, F.; Qi, Q.; Sun, R.-B.; Gu, S.-H.; Di, Z.-Z.; Aa, J.-Y.; Wang, G.-J.; Liu, C.-X. Rebalancing of the gut flora and microbial metabolism is responsible for the anti-arthritis effect of kaempferol. Acta Pharmacol. Sin. 2020, 41, 73–81. [Google Scholar] [CrossRef]
- Nejabati, H.R.; Nikzad, S.; Roshangar, L. Kaempferol: A dietary flavonol in alleviating obesity. Curr. Pharm. Des. 2023, 29, 1547–1556. [Google Scholar] [CrossRef]
- Moura, F.A.; de Andrade, K.Q.; dos Santos, J.C.F.; Araújo, O.R.P.; Goulart, M.O.F. Antioxidant therapy for treatment of inflammatory bowel disease: Does it work? Redox Biol. 2015, 6, 617–639. [Google Scholar] [CrossRef]
- Romier, B.; Schneider, Y.-J.; Larondelle, Y.; During, A. Dietary polyphenols can modulate the intestinal inflammatory response. Nutr. Rev. 2009, 67, 363–378. [Google Scholar] [CrossRef]
- Chen, A.Y.; Chen, Y.C. A review of the dietary flavonoid, kaempferol on human health and cancer chemoprevention. Food Chem. 2013, 138, 2099–2107. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, Z.; Cao, X.; Zou, T.; You, J.; Guan, W. Plant-derived polyphenols in sow nutrition: An update. Anim. Nutr. 2023, 12, 96–107. [Google Scholar] [CrossRef]
- Alam, W.; Khan, H.; Shah, M.A.; Cauli, O.; Saso, L. Kaempferol as a dietary anti-inflammatory agent: Current therapeutic standing. Molecules 2020, 25, 4073. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Shah, Z.A.; Saeed, F.; Imran, A.; Arshad, M.U.; Ahmad, B.; Bawazeer, S.; Atif, M.; Peters, D.G. Chemo-preventive and therapeutic effect of the dietary flavonoid kaempferol: A comprehensive review. Phytother. Res. 2019, 33, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Gao, Y.; Bajpai, V.K.; El-Kammar, H.A.; Simal-Gandara, J.; Cao, H.; Cheng, K.-W.; Wang, M.; Arroo, R.R.J.; Zou, L.; et al. Advance toward isolation, extraction, metabolism and health benefits of kaempferol, a major dietary flavonoid with future perspectives. Crit. Rev. Food Sci. Nutr. 2023, 63, 2773–2789. [Google Scholar] [CrossRef]
- Ren, J.; Lu, Y.; Qian, Y.; Chen, B.; Wu, T.; Ji, G. Recent progress regarding kaempferol for the treatment of various diseases. Exp. Ther. Med. 2019, 18, 2759–2776. [Google Scholar] [CrossRef]
- Calderon-Montano, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A review on the dietary flavonoid kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Cannataro, R.; Fazio, A.; La Torre, C.; Caroleo, M.C.; Cione, E. Polyphenols in the mediterranean diet: From dietary sources to microRNA modulation. Antioxidants 2021, 10, 328. [Google Scholar] [CrossRef]
- Kim, Y.A.; Tarahovsky, Y.S.; Gaidin, S.G.; Yagolnik, E.A.; Muzafarov, E.N. Flavonoids determine the rate of fibrillogenesis and structure of collagen type I fibrils in vitro. Int. J. Biol. Macromol. 2017, 104, 631–637. [Google Scholar] [CrossRef]
- de Vries, J.H.M.; Hollman, P.C.H.; Meyboom, S.; Buysman, M.N.C.P.; Zock, P.L.; van Staveren, W.A.; Katan, M.B. Plasma concentrations and urinary excretion of the antioxidant flavonols quercetin and kaempferol as biomarkers for dietary intake123. Am. J. Clin. Nutr. 1998, 68, 60–65. [Google Scholar] [CrossRef]
- Nielsen, S.E.; Kall, M.; Justesen, U.; Schou, A.; Dragsted, L.O. Human absorption and excretion of flavonoids after broccoli consumption. Cancer Lett. 1997, 114, 173–174. [Google Scholar] [CrossRef]
- DuPont, M.S.; Day, A.J.; Bennett, R.N.; Mellon, F.A.; Kroon, P.A. Absorption of kaempferol from endive, a source of kaempferol-3-glucuronide, in humans. Eur. J. Clin. Nutr. 2004, 58, 947–954. [Google Scholar] [CrossRef]
- Oliveira, E.J.; Watson, D.G.; Grant, M.H. Metabolism of quercetin and kaempferol by rat hepatocytes and the identification of flavonoid glycosides in human plasma. Xenobiotica 2002, 32, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, excretion and metabolite profiling of methyl-, glucuronyl-, glucosyl- and sulpho-conjugates of quercetin in human plasma and urine after ingestion of onions. Br. J. Nutr. 2006, 96, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Fernández-del-Río, L.; Soubeyrand, E.; Basset, G.J.; Clarke, C.F. Metabolism of the flavonol kaempferol in kidney cells liberates the B-ring to enter coenzyme Q biosynthesis. Molecules 2020, 25, 2955. [Google Scholar] [CrossRef]
- Bangar, S.P.; Chaudhary, V.; Sharma, N.; Bansal, V.; Ozogul, F.; Lorenzo, J.M. Kaempferol: A flavonoid with wider biological activities and its applications. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef]
- Ďuračková, Z. Some current insights into oxidative stress. Physiol. Res. 2010, 59, 459–469. [Google Scholar] [CrossRef]
- George, S.; Abrahamse, H. Redox potential of antioxidants in cancer progression and prevention. Antioxidants 2020, 9, 1156. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Liu, X.; Chang, Y.; Wang, R.; Lv, T.; Cui, C.; Liu, M. Investigation of the anti-inflammatory and antioxidant activities of luteolin, kaempferol, apigenin and quercetin. S. Afr. J. Bot. 2021, 137, 257–264. [Google Scholar] [CrossRef]
- Fan, J.; Zhao, X.-H.; Zhao, J.-R.; Li, B.-R. Galangin and kaempferol alleviate the indomethacin-caused cytotoxicity and barrier loss in rat intestinal epithelial (IEC-6) cells via mediating JNK/Src activation. ACS Omega 2021, 6, 15046–15056. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhai, Z.; Jia, H.; Lai, J.; Si, X.; Wu, Z. Kaempferol attenuates diquat-induced oxidative damage and apoptosis in intestinal porcine epithelial cells. Food Funct. 2021, 12, 6889–6899. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, Z.; Cao, X.; Chen, X.; Zou, T.; You, J. Plant-derived polyphenols as Nrf2 activators to counteract oxidative stress and intestinal toxicity induced by deoxynivalenol in swine: An emerging research direction. Antioxidants 2022, 11, 2379. [Google Scholar] [CrossRef]
- Devi, K.P.; Malar, D.S.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Kaempferol and inflammation: From chemistry to medicine. Pharmacol. Res. 2015, 99, 1–10. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, Y.; Li, X.; Zhang, Z.; Sun, L.; Wang, Y.; He, Z.; Liu, Z.; Zhu, L.; Fu, L. Kaempferol suppression of acute colitis is regulated by the efflux transporters BCRP and MRP2. Eur. J. Pharm. Sci. 2022, 179, 106303. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Zhou, Q.; Liu, T.; Liu, P.; Li, H.; Bian, Y.; Liu, Z. Kaempferol relieves the DSS-induced chronic colitis in C57BL/6J mice, alleviates intestinal angiogenesis, and regulates colonic microflora structure. J. Funct. Foods 2023, 107, 105646. [Google Scholar] [CrossRef]
- Qu, Y.; Li, X.; Xu, F.; Zhao, S.; Wu, X.; Wang, Y.; Xie, J. Kaempferol alleviates murine experimental colitis by restoring gut microbiota and inhibiting the LPS-TLR4-NF-κB axis. Front. Immunol. 2021, 12, 679897. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-Y.; Ji, G.E.; Sung, M.-K. Dietary kaempferol suppresses inflammation of dextran sulfate sodium-induced colitis in mice. Dig. Dis. Sci. 2012, 57, 355–363. [Google Scholar] [CrossRef]
- Bian, Y.; Lei, J.; Zhong, J.; Wang, B.; Wan, Y.; Li, J.; Liao, C.; He, Y.; Liu, Z.; Ito, K.; et al. Kaempferol reduces obesity, prevents intestinal inflammation, and modulates gut microbiota in high-fat diet mice. J. Nutr. Biochem. 2022, 99, 108840. [Google Scholar] [CrossRef]
- Bian, Y.; Liu, P.; Zhong, J.; Hu, Y.; Fan, Y.; Zhuang, S.; Liu, Z. Kaempferol inhibits multiple pathways involved in the secretion of inflammatory mediators from LPS-induced rat intestinal microvascular endothelial cells. Mol. Med. Rep. 2019, 19, 1958–1964. [Google Scholar] [CrossRef]
- Yu, R.; Zhong, J.; Zhou, Q.; Ren, W.; Liu, Z.; Bian, Y. Kaempferol prevents angiogenesis of rat intestinal microvascular endothelial cells induced by LPS and TNF-α via inhibiting VEGF/Akt/p38 signaling pathways and maintaining gut-vascular barrier integrity. Chem. Biol. Interact. 2022, 366, 110135. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-J.; Ji, G.-E.; Sung, M.-K. Quercetin and kaempferol suppress immunoglobulin E-mediated allergic inflammation in RBL-2H3 and Caco-2 cells. Inflamm. Res. 2010, 59, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, S.; Del Bo’, C.; Marino, M.; Gargari, G.; Cherubini, A.; Andrés-Lacueva, C.; Hidalgo-Liberona, N.; Peron, G.; González-Dominguez, R.; Kroon, P. Polyphenols and intestinal permeability: Rationale and future perspectives. J. Agric. Food Chem. 2019, 68, 1816–1829. [Google Scholar] [CrossRef] [PubMed]
- Salvo Romero, E.; Alonso Cotoner, C.; Pardo Camacho, C.; Casado Bedmar, M.; Vicario, M. The intestinal barrier function and its involvement in digestive disease. Rev. Esp. Enferm. Dig. 2015, 107, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Antoni, L.; Nuding, S.; Wehkamp, J.; Stange, E.F. Intestinal barrier in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 1165. [Google Scholar] [CrossRef]
- Chen, J.; Xuan, Y.-H.; Luo, M.-X.; Ni, X.-G.; Ling, L.-Q.; Hu, S.-J.; Chen, J.-Q.; Xu, J.-Y.; Jiang, L.-Y.; Si, W.-Z.; et al. Kaempferol alleviates acute alcoholic liver injury in mice by regulating intestinal tight junction proteins and butyrate receptors and transporters. Toxicology 2020, 429, 152338. [Google Scholar] [CrossRef]
- Suzuki, T.; Tanabe, S.; Hara, H. Kaempferol enhances intestinal barrier function through the cytoskeletal association and expression of tight junction proteins in Caco-2 cells. J. Nutr. 2011, 141, 87–94. [Google Scholar] [CrossRef]
- Fan, J.; Zhao, X.-H.; Li, T.-J. Heat treatment of galangin and kaempferol inhibits their benefits to improve barrier function in rat intestinal epithelial cells. J. Nutr. Biochem. 2021, 87, 108517. [Google Scholar] [CrossRef]
- Bian, Y.; Dong, Y.; Sun, J.; Sun, M.; Hou, Q.; Lai, Y.; Zhang, B. Protective effect of kaempferol on LPS-induced inflammation and barrier dysfunction in a coculture model of intestinal epithelial cells and intestinal microvascular endothelial cells. J. Agric. Food Chem. 2020, 68, 160–167. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Zhang, G. Impact of deoxynivalenol and kaempferol on expression of tight junction proteins at different stages of Caco-2 cell proliferation and differentiation. RSC Adv. 2019, 9, 34607–34616. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Zhang, G. A proteomic study on the protective effect of kaempferol pretreatment against deoxynivalenol-induced intestinal barrier dysfunction in a Caco-2 cell model. Food Funct. 2020, 11, 7266–7279. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Khan, I.; Huang, G.; Lu, Y.; Wang, L.; Liu, Y.; Lu, L.; Hsiao, W.L.W.; Liu, Z. Kaempferol acts on bile acid signaling and gut microbiota to attenuate the tumor burden in ApcMin/+ mice. Eur. J. Pharmacol. 2022, 918, 174773. [Google Scholar] [CrossRef]
- Cho, H.J.; Park, J.H. Kaempferol induces cell cycle arrest in HT-29 human colon cancer cells. J. Cancer Prev. 2013, 18, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Du, B.; Wang, T.; Wang, S.; Zhang, J. Kaempferol induces apoptosis in human HCT116 colon cancer cells via the ataxia-telangiectasia mutated-p53 pathway with the involvement of p53 upregulated modulator of apoptosis. Chem. Biol. Interact. 2009, 177, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Konishi, M.; Horinaka, M.; Yasuda, T.; Goda, A.E.; Taniguchi, H.; Yano, K.; Wakada, M.; Sakai, T. Kaempferol sensitizes colon cancer cells to TRAIL-induced apoptosis. Biochem. Biophys. Res. Commun. 2008, 375, 129–133. [Google Scholar] [CrossRef]
- Lee, H.S.; Cho, H.J.; Yu, R.; Lee, K.W.; Chun, H.S.; Park, J.H.Y. Mechanisms underlying apoptosis-inducing effects of Kaempferol in HT-29 human colon cancer cells. Int. J. Mol. Sci. 2014, 15, 2722–2737. [Google Scholar] [CrossRef]
- Duarte, L.; Gasaly, N.; Poblete-Aro, C.; Uribe, D.; Echeverria, F.; Gotteland, M.; Garcia-Diaz, D.F. Polyphenols and their anti-obesity role mediated by the gut microbiota: A comprehensive review. Rev. Endocr. Metab. Disord. 2021, 22, 367–388. [Google Scholar] [CrossRef]
- Osborn, L.J.; Schultz, K.; Massey, W.; DeLucia, B.; Choucair, I.; Varadharajan, V.; Banerjee, R.; Fung, K.; Horak III, A.J.; Orabi, D. A gut microbial metabolite of dietary polyphenols reverses obesity-driven hepatic steatosis. Proc. Natl. Acad. Sci. USA 2022, 119, e2202934119. [Google Scholar] [CrossRef]
- Wang, T.; Wu, Q.; Zhao, T. Preventive effects of kaempferol on high-fat diet-induced obesity complications in C57BL/6 mice. Biomed. Res. Int. 2020, 2020, 4532482. [Google Scholar] [CrossRef]
- Tilg, H.; Zmora, N.; Adolph, T.E.; Elinav, E. The intestinal microbiota fuelling metabolic inflammation. Nat. Rev. Immunol. 2020, 20, 40–54. [Google Scholar] [CrossRef]
- Serre, C.B.D.L.; Ellis, C.L.; Lee, J.; Hartman, A.L.; Rutledge, J.C.; Raybould, H.E. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G440–G448. [Google Scholar] [CrossRef] [PubMed]
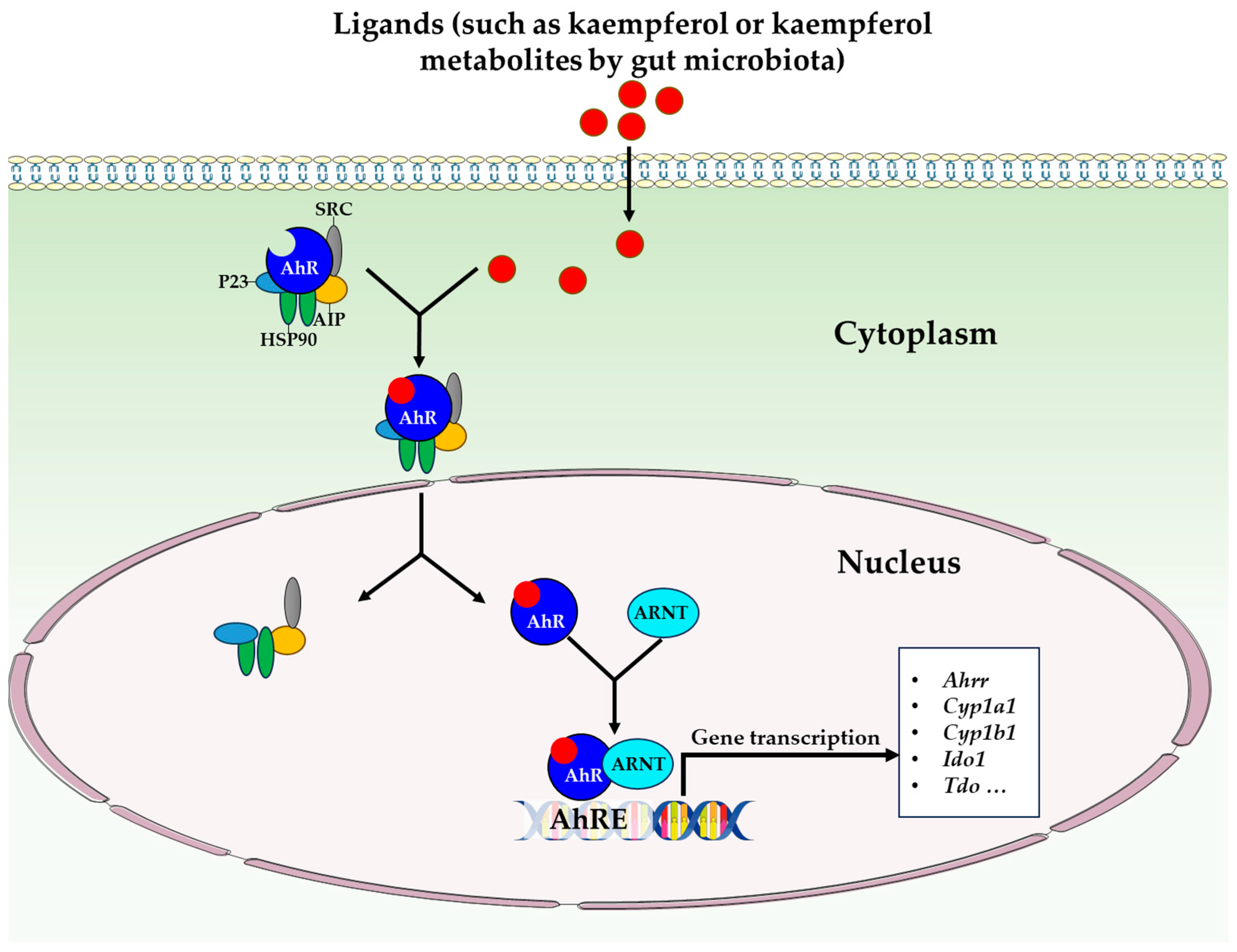
- Pinto, C.J.G.; Ávila-Gálvez, M.Á.; Lian, Y.; Moura-Alves, P.; Nunes dos Santos, C. Targeting the aryl hydrocarbon receptor by gut phenolic metabolites: A strategy towards gut inflammation. Redox Biol. 2023, 61, 102622. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.S.; Cait, A.; Li, Y.; Abolins-Thompson, H.; Gell, K.; Herst, P.M.; O’Sullivan, D.; Gasser, O. Practical approach to explore the effects of polyphenols on aryl hydrocarbon receptor regulated immune function. J. Agric. Food Chem. 2021, 69, 8625–8633. [Google Scholar] [CrossRef] [PubMed]
- Dever, D.P.; Adham, Z.O.; Thompson, B.; Genestine, M.; Cherry, J.; Olschowka, J.A.; DiCicco-Bloom, E.; Opanashuk, L.A. Aryl hydrocarbon receptor deletion in cerebellar granule neuron precursors impairs neurogenesis. Dev. Neurobiol. 2016, 76, 533–550. [Google Scholar] [CrossRef] [PubMed]
- Pernomian, L.; Duarte-Silva, M.; de Barros Cardoso, C.R. The aryl hydrocarbon receptor (AHR) as a potential target for the control of intestinal inflammation: Insights from an immune and bacteria sensor receptor. Clin. Rev. Allergy Immunol. 2020, 59, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Stockinger, B.; Shah, K.; Wincent, E. AHR in the intestinal microenvironment: Safeguarding barrier function. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 559–570. [Google Scholar] [CrossRef]
- Neavin, D.R.; Liu, D.; Ray, B.; Weinshilboum, R.M. The role of the aryl hydrocarbon receptor (AHR) in immune and inflammatory diseases. Int. J. Mol. Sci. 2018, 19, 3851. [Google Scholar] [CrossRef]
- Shivanna, B.; Chu, C.; Moorthy, B. The aryl hydrocarbon receptor (AHR): A novel therapeutic target for pulmonary diseases? Int. J. Mol. Sci. 2022, 23, 1516. [Google Scholar] [CrossRef]
- CIOLINO, H.P.; DASCHNER, P.J.; YEH, G.C. Dietary flavonols quercetin and kaempferol are ligands of the aryl hydrocarbon receptor that affect CYP1A1 transcription differentially. Biochem. J. 1999, 340, 715–722. [Google Scholar] [CrossRef]
- Mohammadi-Bardbori, A.; Bengtsson, J.; Rannug, U.; Rannug, A.; Wincent, E. Quercetin, resveratrol, and curcumin are indirect activators of the aryl hydrocarbon receptor (AHR). Chem. Res. Toxicol. 2012, 25, 1878–1884. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Subject | Stress Model | Kaempferol Dosage and Duration | Main Findings | Reference |
|---|---|---|---|---|
| C57BL/6J mice | Colitis induced by DSS |
|
| [36] |
| C57BL/6J mice | Obesity induced by high-fat diet |
|
| [39] |
| C57BL/6J mice | Obesity induced by high-fat diet |
|
| [59] |
| C57BL/6J mice | Colorectal cancer by impaired tumor suppress gene |
|
| [52] |
| C57BL/6 mice | Colitis induced by DSS |
|
| [35] |
| C57BL/6J mice | Colitis induced by DSS |
|
| [37] |
| ICR mice | Acute liver injury by alcohol |
|
| [46] |
| DBA/1J mice | Arthritis induced by collagen |
|
| [8] |
| C57BL/6J mice | Colitis induced by DSS |
|
| [38] |
| Subject | Stress Model | Kaempferol Dosage and Duration | Main Findings | References |
|---|---|---|---|---|
| Rat intestinal microvascular endothelial cells | Inflammation model induced by LPS and TNF-α |
|
| [41] |
| IPEC-1 cells (intestinal porcine epithelial cells) | Oxidative stress model induced by diquat |
|
| [32] |
| IEC-6 cells (Rat intestinal epithelial cells) | No stress treatment |
|
| [48] |
| IEC-6 cells | Cytotoxicity model induced by indomethacin |
|
| [31] |
| Caco-2 cells (human colon epithelial cancer cells) | Deoxynivalenol-challenge model |
|
| [51] |
| Caco-2 cells cocultured with rat intestinal microvascular endothelial cells | Inflammation model induced by LPS |
|
| [49] |
| Rat intestinal microvascular endothelial cells | Inflammation model induced by LPS |
|
| [40] |
| Caco-2 cells | Deoxynivalenol-challenge model |
|
| [50] |
| HT-29 cells (Human colon cancer cells) | Mutant p53 gene |
|
| [56] |
| HT-29 cells | Mutant p53 gene |
|
| [53] |
| Caco-2 cells | No stress treatment |
|
| [47] |
| Caco-2 cells | OVA-IgE complexes stimulation model |
|
| [42] |
| HCT116 cells (Human colon cancer cells) | Mutant p53 gene |
|
| [54] |
| SW480 cells (Human colon cancer cells) | No stress treatment |
|
| [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Zhong, H.; Huang, Z.; Chen, X.; You, J.; Zou, T. A Critical Review of Kaempferol in Intestinal Health and Diseases. Antioxidants 2023, 12, 1642. https://doi.org/10.3390/antiox12081642
Chen J, Zhong H, Huang Z, Chen X, You J, Zou T. A Critical Review of Kaempferol in Intestinal Health and Diseases. Antioxidants. 2023; 12(8):1642. https://doi.org/10.3390/antiox12081642
Chicago/Turabian StyleChen, Jun, Haopeng Zhong, Zhouyin Huang, Xingping Chen, Jinming You, and Tiande Zou. 2023. "A Critical Review of Kaempferol in Intestinal Health and Diseases" Antioxidants 12, no. 8: 1642. https://doi.org/10.3390/antiox12081642