

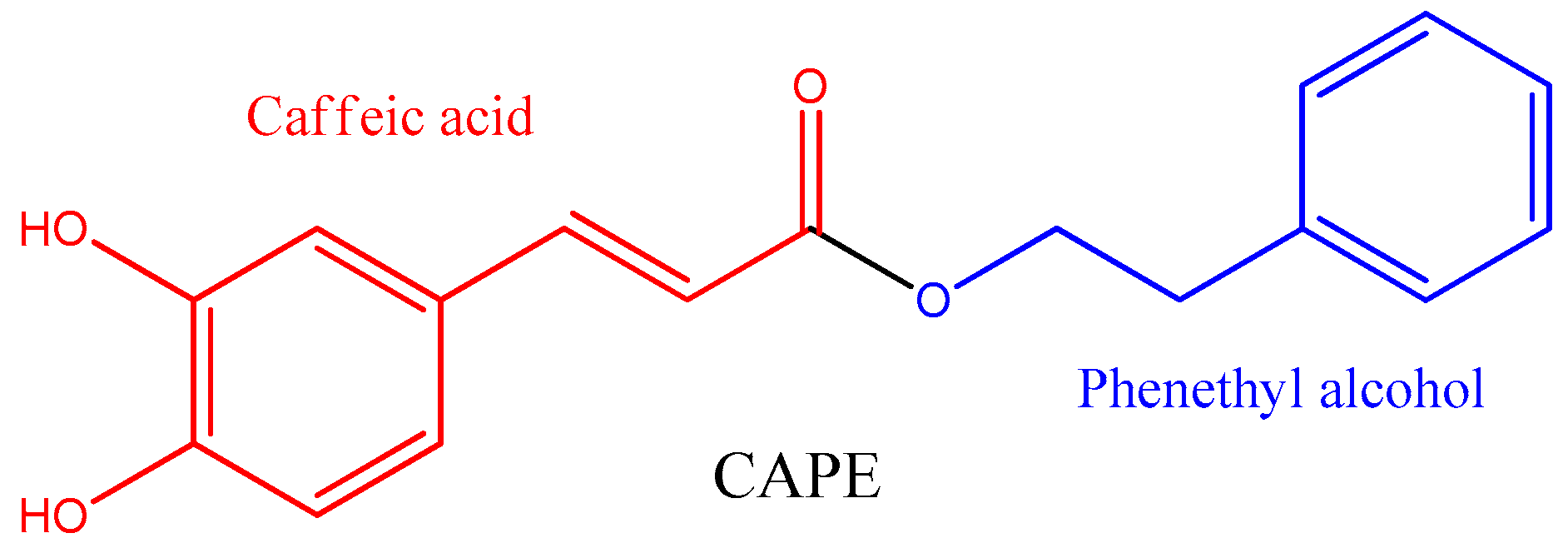
Caffeic Acid Phenethyl Ester (CAPE): Biosynthesis, Derivatives and Formulations with Neuroprotective Activities
 , ,
, , 
Abstract
:1. Introduction
2. Natural Sources of Caffeic Acid Phenethyl Ester
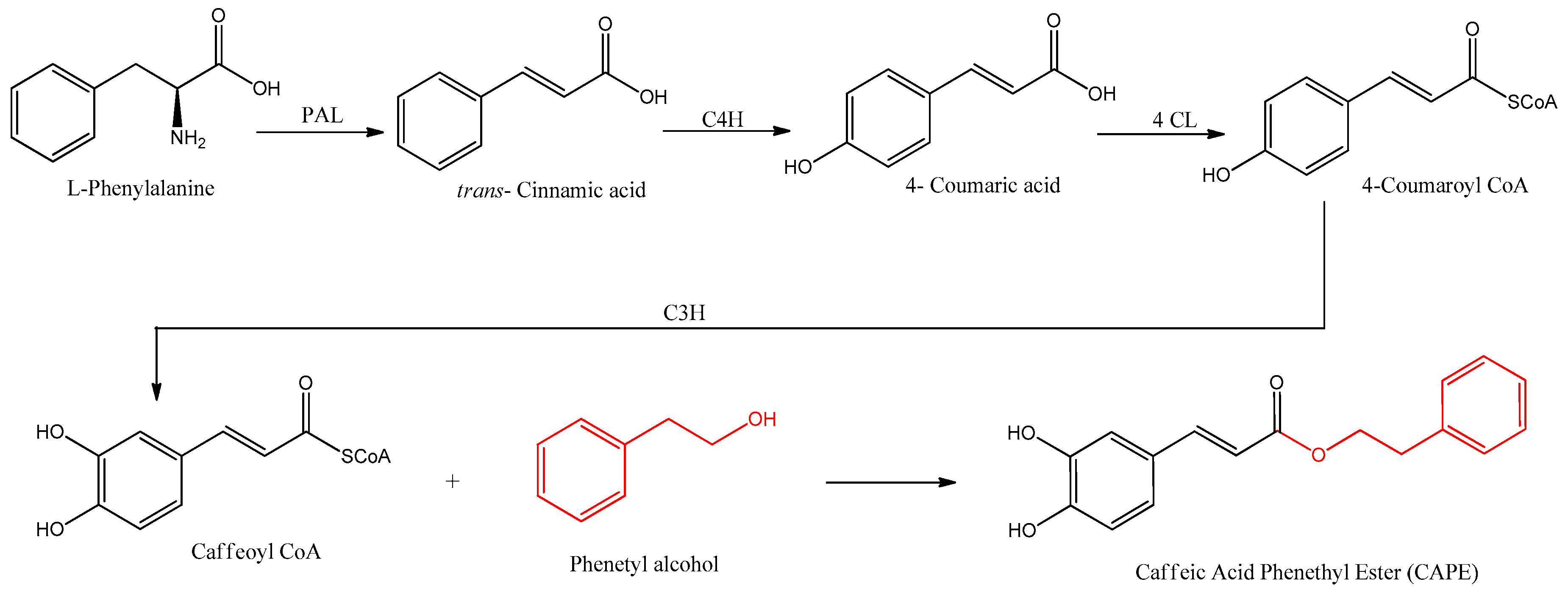
3. Biosynthesis of CAPE
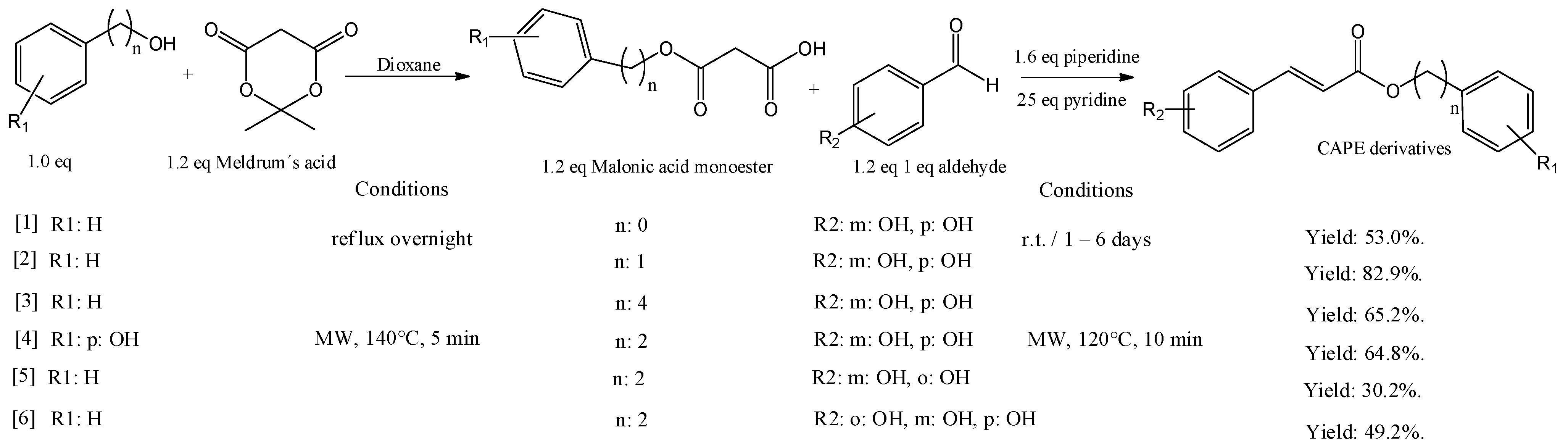
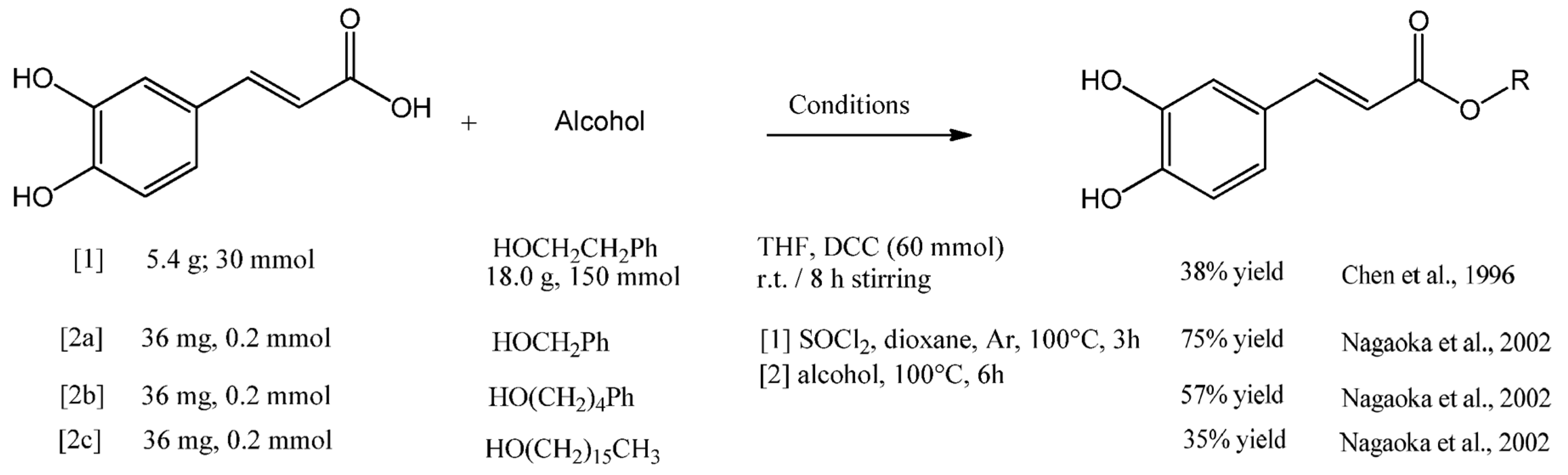
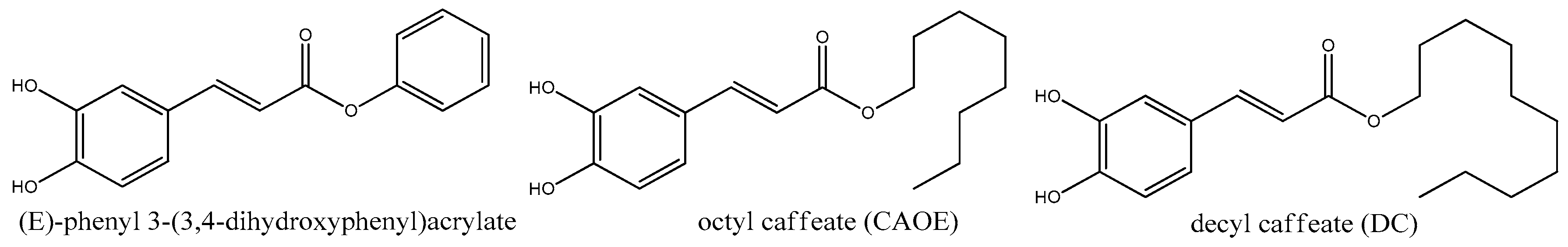
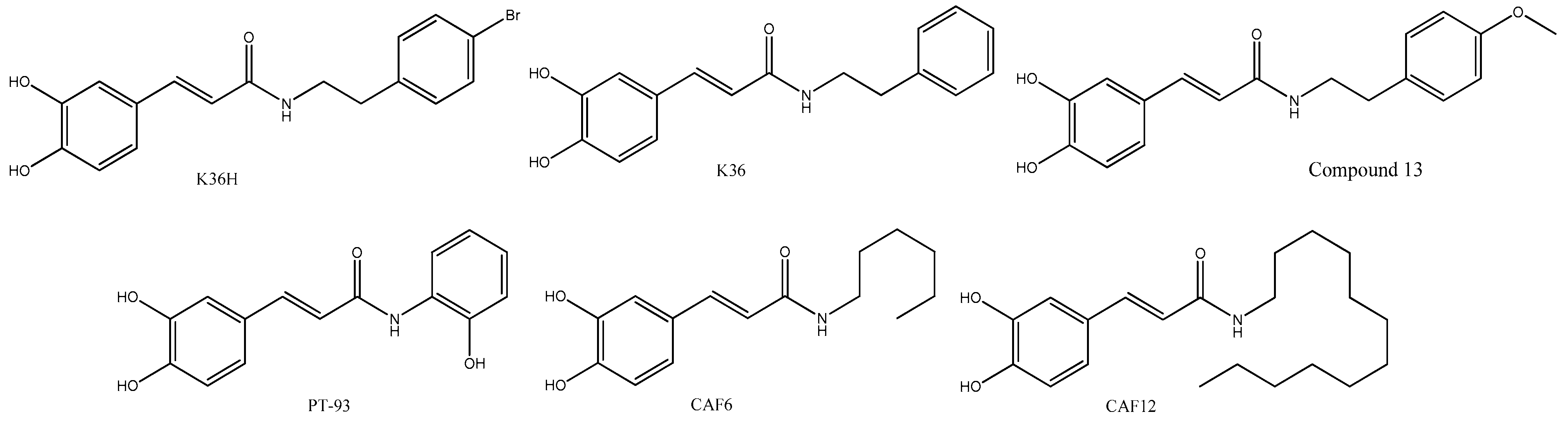
4. CAPE Derivatives as Novel Bioactive Compounds
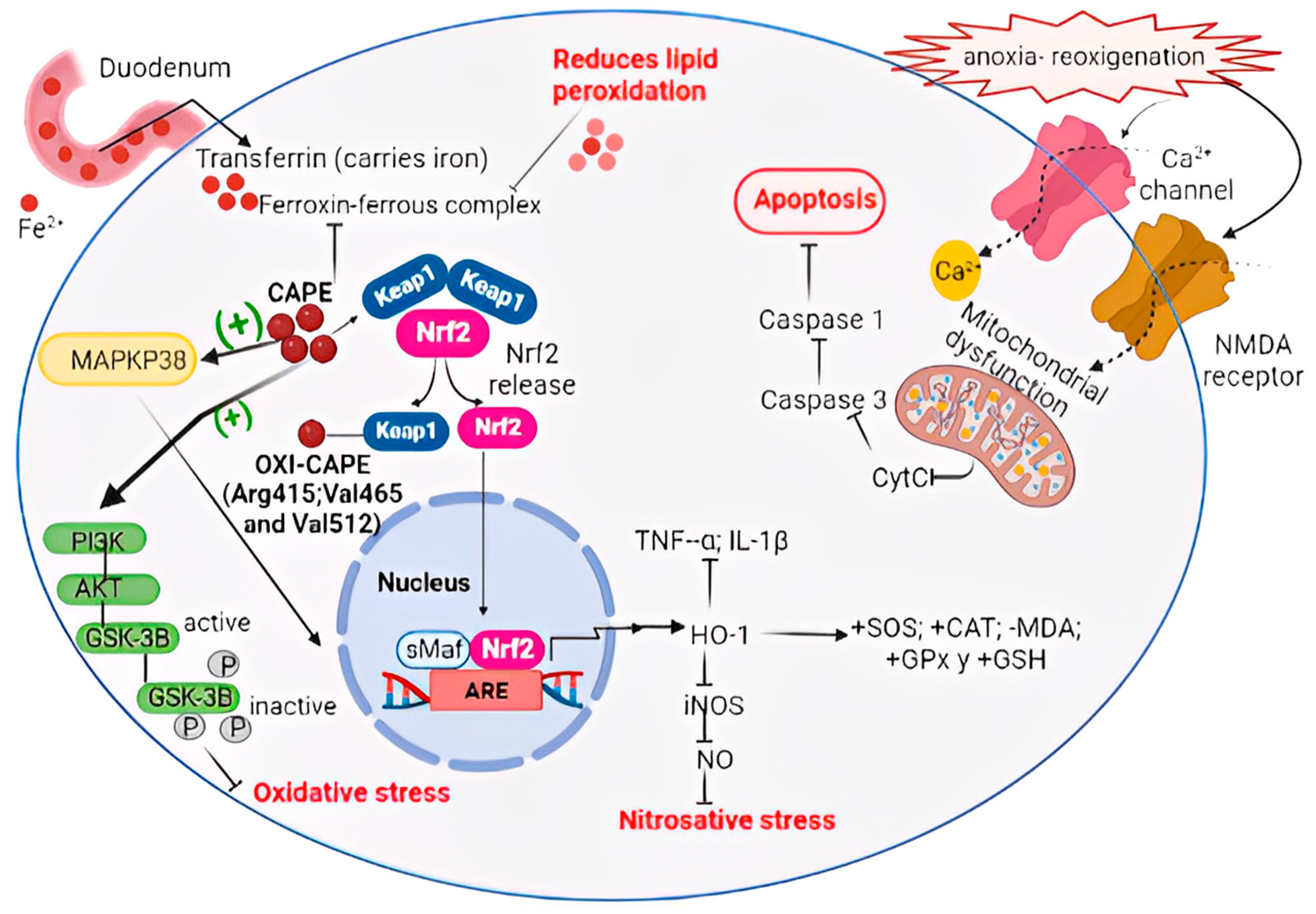
5. CAPE Inhibits Oxidative Stress by Modulation of the Nrf2 Pathway
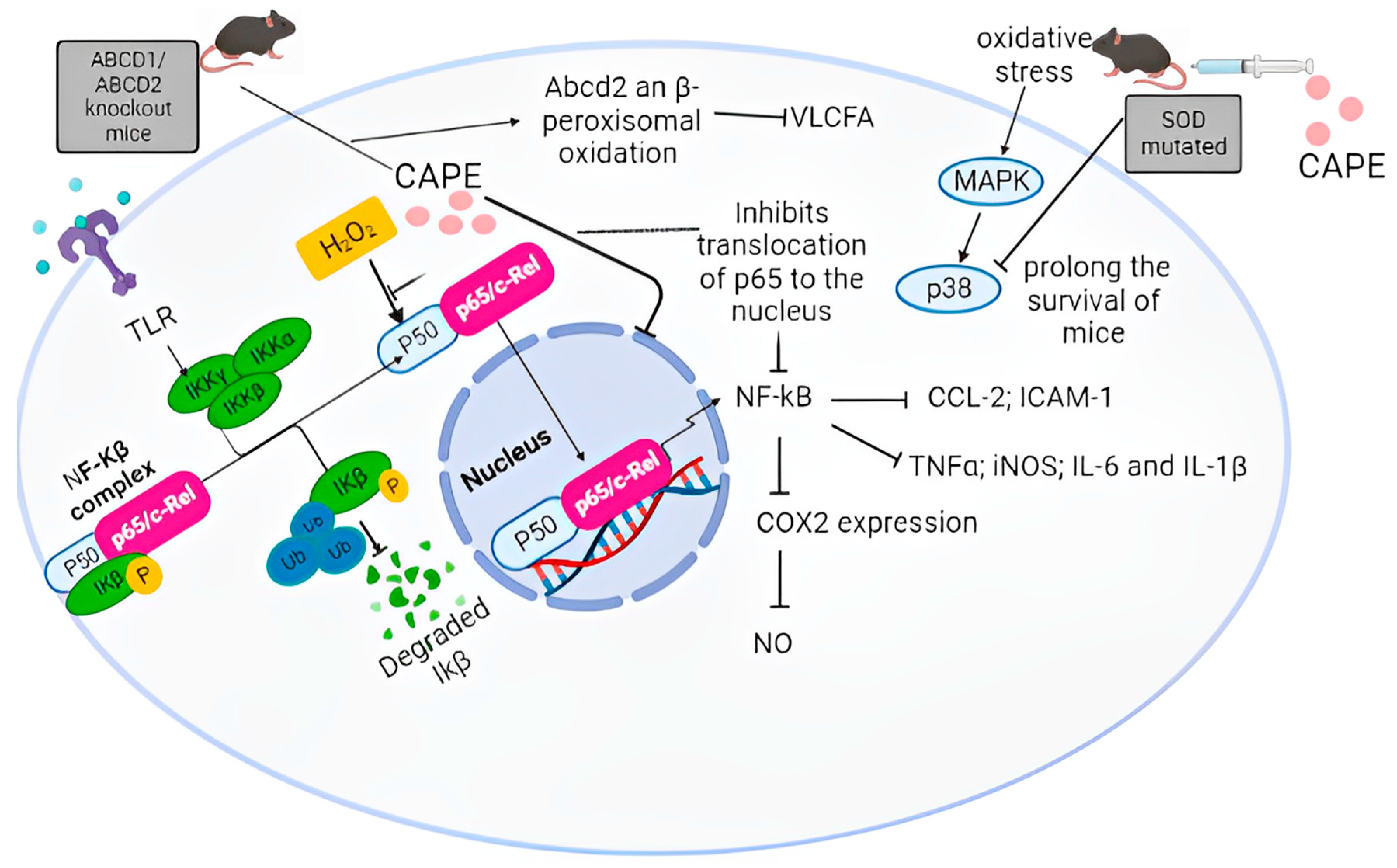
6. CAPE Inhibits Neuroinflammation by Modulation of the NF-κB Pathway
7. Biological Properties of CAPE
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dubois, B.; Villain, N.; Frisoni, G.B.; Rabinovici, G.D.; Sabbagh, M.; Cappa, S.; Bejanin, A.; Bombois, S.; Epelbaum, S.; Teichmann, M.; et al. Clinical Diagnosis of Alzheimer’s Disease: Recommendations of the International Working Group. Lancet Neurol. 2021, 20, 484–496. [Google Scholar] [CrossRef]
- Fukuhara, S.; Tanigaki, R.; Kimura, K.; Kataoka, T. International Immunopharmacology Kujigamberol Interferes with Pro-in Fl Ammatory Cytokine-Induced Expression of and N-Glycan Modi Fi Cations to Cell Adhesion Molecules at Di Ff Erent Stages in Human Umbilical Vein Endothelial Cells. Int. Immunopharmacol. 2018, 62, 313–325. [Google Scholar] [CrossRef]
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune Attack: The Role of Inflammation in Alzheimer Disease. Nat. Rev. Neurosci. 2015, 16, 358–372. [Google Scholar] [CrossRef]
- Ebadi, M.; Srinivasan, S.K.; Baxi, M.D. Oxidative Stress and Antioxidant Therapy in Parkinson’s Disease. Prog. Neurobiol. 1996, 48, 1–19. [Google Scholar] [CrossRef]
- Markesbery, W.R.; Carney, J.M. Oxidative Alterations in Alzheimer’s Disease. Brain Pathol. 1999, 9, 133–146. [Google Scholar] [CrossRef]
- Barber, S.C.; Shaw, P.J. Oxidative Stress in ALS: Key Role in Motor Neuron Injury and Therapeutic Target. Free Radic. Biol. Med. 2010, 48, 629–641. [Google Scholar] [CrossRef]
- Leiva, A.M.; Martínez-Sanguinetti, M.A.; Troncoso-Pantoja, C.; Nazar, G.; Petermann-Rocha, F.; Celis-Morales, C. Parkinson’s Disease in Chile: Highest Prevalence in Latin America. Rev. Med. Chil. 2019, 147, 535–536. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kang, M.-I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative Stress Sensor Keap1 Functions as an Adaptor for Cul3-Based E3 Ligase to Regulate Proteasomal Degradation of Nrf2. Mol. Cell. Biol. 2004, 24, 7130–7139. [Google Scholar] [CrossRef]
- Cullinan, S.B.; Gordan, J.D.; Jin, J.; Harper, J.W.; Diehl, J.A. The Keap1-BTB Protein Is an Adaptor That Bridges Nrf2 to a Cul3-Based E3 Ligase: Oxidative Stress Sensing by a Cul3-Keap1 Ligase. Mol. Cell. Biol. 2004, 24, 8477–8486. [Google Scholar] [CrossRef]
- Sykiotis, G.P.; Bohmann, D. Stress-Activated Cap’n’collar Transcription Factors in Aging and Human Disease. Sci. Signal. 2010, 3, re3. [Google Scholar] [CrossRef]
- Almowallad, S.; Alqahtani, L.S.; Mobashir, M. NF-ΚB in Signaling Patterns and Its Temporal Dynamics Encode/Decode Human Diseases. Life 2022, 12, 2012. [Google Scholar] [CrossRef]
- Borba, R.S.; Wilson, M.B.; Spivak, M. Hidden Benefits of Honeybee Propolis in Hives. In Beekeeping—From Science to Practice; Springer: Cham, Switzerland, 2017; pp. 17–38. [Google Scholar]
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef]
- Drescher, N.; Klein, A.-M.; Neumann, P.; Yañez, O.; Leonhardt, S.D. Inside Honeybee Hives: Impact of Natural Propolis on the Ectoparasitic Mite Varroa Destructor and Viruses. Insects 2017, 8, 15. [Google Scholar] [CrossRef]
- Machado, B.A.S.; Silva, R.P.D.; Barreto, G.d.A.; Costa, S.S.; Silva, D.F.d.; Brandão, H.N.; Rocha, J.L.C.d.; Dellagostin, O.A.; Henriques, J.A.P.; Umsza-Guez, M.A.; et al. Chemical Composition and Biological Activity of Extracts Obtained by Supercritical Extraction and Ethanolic Extraction of Brown, Green and Red Propolis Derived from Different Geographic Regions in Brazil. PLoS ONE 2016, 11, e0145954. [Google Scholar] [CrossRef]
- Freires, I.A.; Queiroz, V.C.P.P.; Furletti, V.F.; Ikegaki, M.; de Alencar, S.M.; Duarte, M.C.T.; Rosalen, P.L. Chemical Composition and Antifungal Potential of Brazilian Propolis against Candida spp. J. Mycol. Med. 2016, 26, 122–132. [Google Scholar] [CrossRef]
- Pazin, W.M.; Monaco, L.d.M.; Egea Soares, A.E.; Miguel, F.G.; Berretta, A.A.; Ito, A.S. Antioxidant Activities of Three Stingless Bee Propolis and Green Propolis Types. J. Apic. Res. 2017, 56, 40–49. [Google Scholar] [CrossRef]
- Tiveron, A.P.; Rosalen, P.L.; Franchin, M.; Lacerda, R.C.C.; Bueno-Silva, B.; Benso, B.; Denny, C.; Ikegaki, M.; de Alencar, S.M. Chemical Characterization and Antioxidant, Antimicrobial, and Anti-Inflammatory Activities of South Brazilian Organic Propolis. PLoS ONE 2016, 11, e0165588. [Google Scholar] [CrossRef]
- Metzner, J.; Bekemeier, H.; Paintz, M.; Schneidewind, E. On the antimicrobial activity of propolis and propolis constituents (author’s transl). Pharmazie 1979, 34, 97–102. [Google Scholar]
- Romero, F.; Palacios, J.; Jofré, I.; Paz, C.; Nwokocha, C.R.; Paredes, A.; Cifuentes, F. Aristoteline, an Indole-Alkaloid, Induces Relaxation by Activating Potassium Channels and Blocking Calcium Channels in Isolated Rat Aorta. Molecules 2019, 24, 2748. [Google Scholar] [CrossRef]
- Kurek-Górecka, A.; Rzepecka-Stojko, A.; Górecki, M.; Stojko, J.; Sosada, M.; Swierczek-Zieba, G. Structure and Antioxidant Activity of Polyphenols Derived from Propolis. Molecules 2013, 19, 78–101. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-Antioxidant Activity Relationships of Flavonoids and Phenolic Acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Ozdal, T.; Ceylan, F.D.; Eroglu, N.; Kaplan, M.; Olgun, E.O.; Capanoglu, E. Investigation of Antioxidant Capacity, Bioaccessibility and LC-MS/MS Phenolic Profile of Turkish Propolis. Food Res. Int. 2019, 122, 528–536. [Google Scholar] [CrossRef]
- Gómez-Caravaca, A.M.; Gómez-Romero, M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Advances in the Analysis of Phenolic Compounds in Products Derived from Bees. J. Pharm. Biomed. Anal. 2006, 41, 1220–1234. [Google Scholar] [CrossRef]
- Zhu, Y.; Shen, T.; Lin, Y.; Chen, B.; Ruan, Y.; Cao, Y.; Qiao, Y.; Man, Y.; Wang, S.; Li, J. Astragalus Polysaccharides Suppress ICAM-1 and VCAM-1 Expression in TNF-α-Treated Human Vascular Endothelial Cells by Blocking NF-ΚB Activation. Nat. Publ. Gr. 2013, 34, 1036–1042. [Google Scholar] [CrossRef]
- Hernandez, J.; Goycoolea, F.M.; Quintero, J.; Acosta, A.; Castañeda, M.; Dominguez, Z.; Robles, R.; Vazquez-Moreno, L.; Velazquez, E.F.; Astiazaran, H.; et al. Sonoran Propolis: Chemical Composition and Antiproliferative Activity on Cancer Cell Lines. Planta Med. 2007, 73, 1469–1474. [Google Scholar] [CrossRef]
- Borrás, M.J. Universitat Politècnica De València Perfil Fenólico De Propóleos De Diferentes Orígenes Geográficos. Master’s Thesis, Universitat Politècnica de València, València, Spain, 2018. [Google Scholar]
- Li, N.; Ng, T.B.; Wong, J.H.; Qiao, J.X.; Zhang, Y.N.; Zhou, R.; Chen, R.R.; Liu, F. Separation and Purification of the Antioxidant Compounds, Caffeic Acid Phenethyl Ester and Caffeic Acid from Mushrooms by Molecularly Imprinted Polymer. Food Chem. 2013, 139, 1161–1167. [Google Scholar] [CrossRef]
- Chen, Y.J.; Shiao, M.S.; Hsu, M.L.; Tsai, T.H.; Wang, S.Y. Effect of Caffeic Acid Phenethyl Ester, an Antioxidant from Propolis, on Inducing Apoptosis in Human Leukemic HL-60 Cells. J. Agric. Food Chem. 2001, 49, 5615–5619. [Google Scholar] [CrossRef]
- Watabe, M.; Hishikawa, K.; Takayanagi, A.; Shimizu, N.; Nakaki, T. Caffeic Acid Phenethyl Ester Induces Apoptosis by Inhibition of NFkappaB and Activation of Fas in Human Breast Cancer MCF-7 Cells. J. Biol. Chem. 2004, 279, 6017–6026. [Google Scholar] [CrossRef]
- Xiang, D.; Wang, D.; He, Y.; Xie, J.; Zhong, Z.; Li, Z.; Xie, J. Caffeic Acid Phenethyl Ester Induces Growth Arrest and Apoptosis of Colon Cancer Cells via the Beta-Catenin/T-Cell Factor Signaling. Anticancer. Drugs 2006, 17, 753–762. [Google Scholar] [CrossRef]
- Chen, J.H.; Shao, Y.; Huang, M.T.; Chin, C.K.; Ho, C.T. Inhibitory Effect of Caffeic Acid Phenethyl Ester on Human Leukemia HL-60 Cells. Cancer Lett. 1996, 108, 211–214. [Google Scholar] [CrossRef]
- Murtaza, G.; Sajjad, A.; Mehmood, Z.; Shah, S.H.; Siddiqi, A.R. Possible Molecular Targets for Therapeutic Applications of Caffeic Acid Phenethyl Ester in Inflammation and Cancer. J. Food Drug Anal. 2015, 23, 11–18. [Google Scholar] [CrossRef]
- Jiang, M.; Wu, N.; Xu, B.; Chu, Y.; Li, X.; Su, S.; Chen, D.; Li, W.; Shi, Y.; Gao, X.; et al. Fatty Acid-Induced CD36 Expression via O-GlcNAcylation Drives Gastric Cancer Metastasis. Theranostics 2019, 9, 5359–5373. [Google Scholar] [CrossRef]
- Omar, M.H.; Mullen, W.; Stalmach, A.; Auger, C.; Rouanet, J.-M.; Teissedre, P.-L.; Caldwell, S.T.; Hartley, R.C.; Crozier, A. Absorption, Disposition, Metabolism, and Excretion of [3-(14)C]Caffeic Acid in Rats. J. Agric. Food Chem. 2012, 60, 5205–5214. [Google Scholar] [CrossRef]
- Celli, N.; Dragani, L.K.; Murzilli, S.; Pagliani, T.; Poggi, A. In Vitro and in Vivo Stability of Caffeic Acid Phenethyl Ester, a Bioactive Compound of Propolis. J. Agric. Food Chem. 2007, 55, 3398–3407. [Google Scholar] [CrossRef]
- de Oliveira, D.M.; Sampaio, G.R.; Pinto, C.B.; Catharino, R.R.; Bastos, D.H.M. Bioavailability of Chlorogenic Acids in Rats after Acute Ingestion of Maté Tea (Ilex Paraguariensis) or 5-Caffeoylquinic Acid. Eur. J. Nutr. 2017, 56, 2541–2556. [Google Scholar] [CrossRef]
- Choi, W.; Villegas, V.; Istre, H.; Heppler, B.; Gonzalez, N.; Brusman, N.; Snider, L.; Hogle, E.; Tucker, J.; Oñate, A.; et al. Synthesis and Characterization of CAPE Derivatives as Xanthine Oxidase Inhibitors with Radical Scavenging Properties. Bioorg. Chem. 2019, 86, 686–695. [Google Scholar] [CrossRef]
- Nagaoka, T.; Banskota, A.H.; Tezuka, Y.; Saiki, I.; Kadota, S. Selective Antiproliferative Activity of Caffeic Acid Phenethyl Ester Analogues on Highly Liver-Metastatic Murine Colon 26-L5 Carcinoma Cell Line. Bioorg. Med. Chem. 2002, 10, 3351–3359. [Google Scholar] [CrossRef]
- Shen, H.; Yamashita, A.; Nakakoshi, M.; Yokoe, H.; Sudo, M.; Kasai, H.; Tanaka, T.; Fujimoto, Y.; Ikeda, M.; Kato, N.; et al. Inhibitory Effects of Caffeic Acid Phenethyl Ester Derivatives on Replication of Hepatitis C Virus. PLoS ONE 2013, 8, e82299. [Google Scholar] [CrossRef]
- Tsai, T.-H.; Yu, C.-H.; Chang, Y.-P.; Lin, Y.-T.; Huang, C.-J.; Kuo, Y.-H.; Tsai, P.-J. Protective Effect of Caffeic Acid Derivatives on Tert-Butyl Hydroperoxide-Induced Oxidative Hepato-Toxicity and Mitochondrial Dysfunction in HepG2 Cells. Molecules 2017, 22, 702. [Google Scholar] [CrossRef]
- Chen, C.; Kuo, Y.-H.; Lin, C.-C.; Chao, C.-Y.; Pai, M.-H.; Chiang, E.-P.I.; Tang, F.-Y. Decyl Caffeic Acid Inhibits the Proliferation of Colorectal Cancer Cells in an Autophagy-Dependent Manner in Vitro and in Vivo. PLoS ONE 2020, 15, e0232832. [Google Scholar] [CrossRef]
- Kuo, Y.-H.; Chen, C.-W.; Chu, Y.; Lin, P.; Chiang, H.-M. In Vitro and In Vivo Studies on Protective Action of N-Phenethyl Caffeamide against Photodamage of Skin. PLoS ONE 2015, 10, e0136777. [Google Scholar] [CrossRef]
- Chu, Y.; Wu, P.-Y.; Chen, C.-W.; Lyu, J.-L.; Liu, Y.-J.; Wen, K.-C.; Lin, C.-Y.; Kuo, Y.-H.; Chiang, H.-M. Protective Effects and Mechanisms of N-Phenethyl Caffeamide from UVA-Induced Skin Damage in Human Epidermal Keratinocytes through Nrf2/HO-1 Regulation. Int. J. Mol. Sci. 2019, 20, 164. [Google Scholar] [CrossRef]
- Kuo, Y.-H.; Chiang, H.-L.; Wu, P.-Y.; Chu, Y.; Chang, Q.-X.; Wen, K.-C.; Lin, C.-Y.; Chiang, H.-M. Protection against Ultraviolet A-Induced Skin Apoptosis and Carcinogenesis through the Oxidative Stress Reduction Effects of N-(4-Bromophenethyl) Caffeamide, A Propolis Derivative. Antioxidants 2020, 9, 335. [Google Scholar] [CrossRef]
- Fathalipour, M.; Eghtedari, M.; Borges, F.; Silva, T.; Moosavi, F.; Firuzi, O.; Mirkhani, H. Caffeic Acid Alkyl Amide Derivatives Ameliorate Oxidative Stress and Modulate ERK1/2 and AKT Signaling Pathways in a Rat Model of Diabetic Retinopathy. Chem. Biodivers. 2019, 16, e1900405. [Google Scholar] [CrossRef]
- Moosavi, F.; Hosseini, R.; Rajaian, H.; Silva, T.; Magalhães e Silva, D.; Saso, L.; Edraki, N.; Miri, R.; Borges, F.; Firuzi, O. Derivatives of Caffeic Acid, a Natural Antioxidant, as the Basis for the Discovery of Novel Nonpeptidic Neurotrophic Agents. Bioorg. Med. Chem. 2017, 25, 3235–3246. [Google Scholar] [CrossRef]
- Cardiology, M.; Zhang, M.; Zhao, G.; Yin, K.; Xia, X.; Gong, D.; Zhao, Z.; Chen, L.; Zheng, X.; Tang, X.; et al. Apolipoprotein A-1 Binding Protein Inhibits Inflammatory Signaling Pathways by Binding to Apolipoprotein A-1 in THP-1 Macrophages. Circ. J. 2018, 82, 1396–1404. [Google Scholar] [CrossRef]
- Chai, T.; Zhao, X.-B.; Wang, W.-F.; Qiang, Y.; Zhang, X.-Y.; Yang, J.-L. Design, Synthesis of N-Phenethyl Cinnamide Derivatives and Their Biological Activities for the Treatment of Alzheimer’s Disease: Antioxidant, Beta-Amyloid Disaggregating and Rescue Effects on Memory Loss. Molecules 2018, 23, 2663. [Google Scholar] [CrossRef]
- Physiology, C. Tanshinone II a Attenuates TNF-α-Induced Expression of VCAM-1 and ICAM-1 in Endothelial Progenitor Cells by Blocking Activation of NF-ΚB. Cell. Physiol. Biochem. 2016, 40, 195–206, Erratum in Cell. Physiol. Biochem. 2017, 41, 2132. [Google Scholar] [CrossRef]
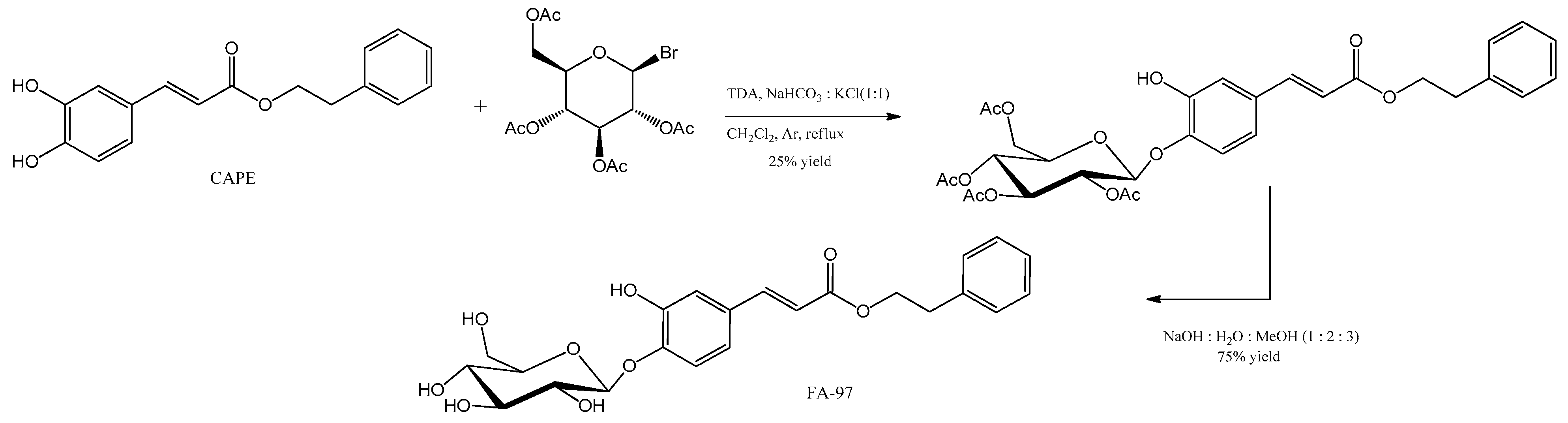
- Wan, T.; Wang, Z.; Luo, Y.; Zhang, Y.; He, W.; Mei, Y.; Xue, J.; Li, M.; Pan, H.; Li, W.; et al. FA-97, a New Synthetic Caffeic Acid Phenethyl Ester Derivative, Protects against Oxidative Stress-Mediated Neuronal Cell Apoptosis and Scopolamine-Induced Cognitive Impairment by Activating Nrf2/HO-1 Signaling. Oxid. Med. Cell. Longev. 2019, 2019, 8239642. [Google Scholar] [CrossRef]
- Sies, H. Biochemistry of Oxidative Stress. Angew. Chem. Int. Ed. Engl. 1986, 25, 1058–1071. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015; ISBN 0198717474. [Google Scholar]
- Dawson, T.M.; Dawson, V.L. Nitric Oxide Synthase: Role as a Transmitter/Mediator in the Brain and Endocrine System. Annu. Rev. Med. 1996, 47, 219–227. [Google Scholar] [CrossRef]
- Halliwell, B. Role of Free Radicals in the Neurodegenerative Diseases: Therapeutic Implications for Antioxidant Treatment. Drugs Aging 2001, 18, 685–716. [Google Scholar] [CrossRef]
- Hammer, B.; Parker, W.D.J.; Bennett, J.P.J. NMDA Receptors Increase OH Radicals in Vivo by Using Nitric Oxide Synthase and Protein Kinase C. Neuroreport 1993, 5, 72–74. [Google Scholar] [CrossRef]
- Sapolsky, R.M. Why Stress Is Bad for Your Brain. Science 1996, 273, 749–750. [Google Scholar] [CrossRef]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF Val66met Polymorphism Affects Activity-Dependent Secretion of BDNF and Human Memory and Hippocampal Function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef]
- Ceccatelli, S.; Tamm, C.; Zhang, Q.; Chen, M. Mechanisms and Modulation of Neural Cell Damage Induced by Oxidative Stress. Physiol. Behav. 2007, 92, 87–92. [Google Scholar] [CrossRef]
- Lee, C.; Fletcher, T.D.; Sun, G. Nitrogen Removal in Constructed Wetland Systems. Eng. Life Sci. 2009, 9, 11–22. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of Curcumin: Problems and Promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef]
- Tonda-Turo, C.; Origlia, N.; Mattu, C.; Accorroni, A.; Chiono, V. Current Limitations in the Treatment of Parkinson’s and Alzheimer’s Diseases: State-of-the-Art and Future Perspective of Polymeric Carriers. Curr. Med. Chem. 2018, 25, 5755–5771. [Google Scholar] [CrossRef]
- Hayashi, M. Oxidative Stress in Developmental Brain Disorders. Neuropathology 2009, 29, 1–8. [Google Scholar] [CrossRef]
- Marlatt, M.W.; Lucassen, P.J.; Perry, G.; Smith, M.A.; Zhu, X. Alzheimer’s Disease: Cerebrovascular Dysfunction, Oxidative Stress, and Advanced Clinical Therapies. J. Alzheimers. Dis. 2008, 15, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Kamat, C.D.; Gadal, S.; Mhatre, M.; Williamson, K.S.; Pye, Q.N.; Hensley, K. Antioxidants in Central Nervous System Diseases: Preclinical Promise and Translational Challenges. J. Alzheimers. Dis. 2008, 15, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Tabrizi, S.J.; Cleeter, M.W.; Xuereb, J.; Taanman, J.W.; Cooper, J.M.; Schapira, A.H. Biochemical Abnormalities and Excitotoxicity in Huntington’s Disease Brain. Ann. Neurol. 1999, 45, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Sonnen, J.A.; Breitner, J.C.; Lovell, M.A.; Markesbery, W.R.; Quinn, J.F.; Montine, T.J. Free Radical-Mediated Damage to Brain in Alzheimer’s Disease and Its Transgenic Mouse Models. Free Radic. Biol. Med. 2008, 45, 219–230. [Google Scholar] [CrossRef]
- Borrás, C.; Sastre, J.; García-Sala, D.; Lloret, A.; Pallardó, F.V.; Viña, J. Mitochondria from Females Exhibit Higher Antioxidant Gene Expression and Lower Oxidative Damage than Males. Free Radic. Biol. Med. 2003, 34, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Huang, Y.; Przedborski, S. Oxidative Stress in Parkinson’s Disease: A Mechanism of Pathogenic and Therapeutic Significance. Ann. N. Y. Acad. Sci. 2008, 1147, 93–104. [Google Scholar] [CrossRef]
- Lee, J.-M.; Johnson, J.A. An Important Role of Nrf2-ARE Pathway in the Cellular Defense Mechanism. J. Biochem. Mol. Biol. 2004, 37, 139–143. [Google Scholar] [CrossRef]
- Itoh, K.; Tong, K.I.; Yamamoto, M. Molecular Mechanism Activating Nrf2-Keap1 Pathway in Regulation of Adaptive Response to Electrophiles. Free Radic. Biol. Med. 2004, 36, 1208–1213. [Google Scholar] [CrossRef]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 System in Development, Oxidative Stress Response and Diseases: An Evolutionarily Conserved Mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef]
- Wang, T.; Chen, L.; Wu, W.; Long, Y.; Wang, R. Potential Cytoprotection: Antioxidant Defence by Caffeic Acid Phenethyl Ester against Free Radical-Induced Damage of Lipids, DNA, and Proteins. Can. J. Physiol. Pharmacol. 2008, 86, 279–287. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Huang, A.-C.; Chang, H.-H.; Liao, H.-F.; Jiang, C.-M.; Lai, L.-Y.; Chan, J.-T.; Chen, Y.-Y.; Chiang, J. Caffeic Acid Phenethyl Ester, an Antioxidant from Propolis, Protects Peripheral Blood Mononuclear Cells of Competitive Cyclists against Hyperthermal Stress. J. Food Sci. 2009, 74, H162–H167. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Stavchansky, S.; Kerwin, S.M.; Bowman, P.D. Structure-Activity Relationships in the Cytoprotective Effect of Caffeic Acid Phenethyl Ester (CAPE) and Fluorinated Derivatives: Effects on Heme Oxygenase-1 Induction and Antioxidant Activities. Eur. J. Pharmacol. 2010, 635, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tran, J.-T.A.; Anderson, R.E.; Mandal, M.N.A. Caffeic Acid Phenethyl Ester Protects 661W Cells from H2O2-Mediated Cell Death and Enhances Electroretinography Response in Dim-Reared Albino Rats. Mol. Vis. 2012, 18, 1325–1338. [Google Scholar] [PubMed]
- Eşrefoğlu, M.; Iraz, M.; Ateş, B.; Gül, M. Not Only Melatonin but Also Caffeic Acid Phenethyl Ester Protects Kidneys against Aging-Related Oxidative Damage in Sprague Dawley Rats. Ultrastruct. Pathol. 2012, 36, 244–251. [Google Scholar] [CrossRef]
- Göçer, H.; Gülçin, I. Caffeic Acid Phenethyl Ester (CAPE): Correlation of Structure and Antioxidant Properties. Int. J. Food Sci. Nutr. 2011, 62, 821–825. [Google Scholar] [CrossRef]
- Kim, H.; Kim, W.; Yum, S.; Hong, S.; Oh, J.-E.; Lee, J.-W.; Kwak, M.-K.; Park, E.J.; Na, D.H.; Jung, Y. Caffeic Acid Phenethyl Ester Activation of Nrf2 Pathway Is Enhanced under Oxidative State: Structural Analysis and Potential as a Pathologically Targeted Therapeutic Agent in Treatment of Colonic Inflammation. Free Radic. Biol. Med. 2013, 65, 552–562. [Google Scholar] [CrossRef]
- Sun, W.; Xie, W.; Huang, D.; Cui, Y.; Yue, J.; He, Q.; Jiang, L.; Xiong, J.; Sun, W.; Yi, Q. Caffeic Acid Phenethyl Ester Attenuates Osteoarthritis Progression by Activating NRF2/HO-1 and Inhibiting the NF-κB Signaling Pathway. Int. J. Mol. Med. 2022, 50, 134. [Google Scholar] [CrossRef]
- Kim, J.-K.; Jang, H.-D. Nrf2-Mediated HO-1 Induction Coupled with the ERK Signaling Pathway Contributes to Indirect Antioxidant Capacity of Caffeic Acid Phenethyl Ester in HepG2 Cells. Int. J. Mol. Sci. 2014, 15, 12149–12165. [Google Scholar] [CrossRef]
- Khan, M.; Elango, C.; Ansari, M.A.; Singh, I.; Singh, A.K. Caffeic Acid Phenethyl Ester Reduces Neurovascular Inflammation and Protects Rat Brain Following Transient Focal Cerebral Ischemia. J. Neurochem. 2007, 102, 365–377. [Google Scholar] [CrossRef]
- Fontanilla, C.V.; Ma, Z.; Wei, X.; Klotsche, J.; Zhao, L.; Wisniowski, P.; Dodel, R.C.; Farlow, M.R.; Oertel, W.H.; Du, Y. Caffeic Acid Phenethyl Ester Prevents 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Neurodegeneration. Neuroscience 2011, 188, 135–141. [Google Scholar] [CrossRef]
- Kurauchi, Y.; Hisatsune, A.; Isohama, Y.; Mishima, S.; Katsuki, H. Caffeic Acid Phenethyl Ester Protects Nigral Dopaminergic Neurons via Dual Mechanisms Involving Haem Oxygenase-1 and Brain-Derived Neurotrophic Factor. Br. J. Pharmacol. 2012, 166, 1151–1168. [Google Scholar] [CrossRef] [PubMed]
- Aladag, M.A.; Turkoz, Y.; Ozcan, C.; Sahna, E.; Parlakpinar, H.; Akpolat, N.; Cigremis, Y. Caffeic Acid Phenethyl Ester (CAPE) Attenuates Cerebral Vasospasm after Experimental Subarachnoidal Haemorrhage by Increasing Brain Nitric Oxide Levels. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2006, 24, 9–14. [Google Scholar] [CrossRef]
- Irmak, M.K.; Fadillioglu, E.; Sogut, S.; Erdogan, H.; Gulec, M.; Ozer, M.; Yagmurca, M.; Gozukara, M.E. Effects of Caffeic Acid Phenethyl Ester and Alpha-Tocopherol on Reperfusion Injury in Rat Brain. Cell Biochem. Funct. 2003, 21, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Lu, Y.-W.; Xu, P.-H.; Long, Y.; Wu, W.-M.; Li, W.; Wang, R. Caffeic Acid Phenethyl Ester and Its Related Compounds Limit the Functional Alterations of the Isolated Mouse Brain and Liver Mitochondria Submitted to in Vitro Anoxia-Reoxygenation: Relationship to Their Antioxidant Activities. Biochim. Biophys. Acta 2008, 1780, 659–672. [Google Scholar] [CrossRef]
- Wei, X.; Zhao, L.; Ma, Z.; Holtzman, D.M.; Yan, C.; Dodel, R.C.; Hampel, H.; Oertel, W.; Farlow, M.R.; Du, Y. Caffeic Acid Phenethyl Ester Prevents Neonatal Hypoxic-Ischaemic Brain Injury. Brain 2004, 127, 2629–2635. [Google Scholar] [CrossRef]
- Mincheva, S.; Garcera, A.; Gou-Fabregas, M.; Encinas, M.; Dolcet, X.; Soler, R.M. The Canonical Nuclear Factor-ΚB Pathway Regulates Cell Survival in a Developmental Model of Spinal Cord Motoneurons. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 6493–6503. [Google Scholar] [CrossRef]
- Demchenko, Y.N.; Glebov, O.K.; Zingone, A.; Keats, J.J.; Bergsagel, P.L.; Kuehl, W.M. Classical and/or Alternative NF-KappaB Pathway Activation in Multiple Myeloma. Blood 2010, 115, 3541–3552. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.-L.; Liu, Y.-H.; Liu, C.; Qi, M.-P.; Liu, R.-N.; Zhu, X.-F.; Zhou, Q.-G.; Chen, Y.-Y.; Guo, A.-Z.; Hu, C.-M. Indirubin Inhibits LPS-Induced Inflammation via TLR4 Abrogation Mediated by the NF-ΚB and MAPK Signaling Pathways. Inflammation 2017, 40, 1–12. [Google Scholar] [CrossRef]
- Karin, M.; Ben-Neriah, Y. Phosphorylation Meets Ubiquitination: The Control of NF-[Kappa]B Activity. Annu. Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-KappaB Family of Transcription Factors and Its Regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Natarajan, K.; Singh, S.; Burke, T.R.J.; Grunberger, D.; Aggarwal, B.B. Caffeic Acid Phenethyl Ester Is a Potent and Specific Inhibitor of Activation of Nuclear Transcription Factor NF-Kappa B. Proc. Natl. Acad. Sci. USA 1996, 93, 9090–9095. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Shin, D.-H.; Kim, J.-H.; Hong, S.; Choi, D.; Kim, Y.-J.; Kwak, M.-K.; Jung, Y. Caffeic Acid Phenethyl Ester-Mediated Nrf2 Activation and IkappaB Kinase Inhibition Are Involved in NFkappaB Inhibitory Effect: Structural Analysis for NFkappaB Inhibition. Eur. J. Pharmacol. 2010, 643, 21–28. [Google Scholar] [CrossRef]
- Choi, K.; Choi, C. Differential Regulation of C-Jun N-Terminal Kinase and NF-KappaB Pathway by Caffeic Acid Phenethyl Ester in Astroglial and Monocytic Cells. J. Neurochem. 2008, 105, 557–564. [Google Scholar] [CrossRef]
- Singh, J.; Khan, M.; Singh, I. Caffeic Acid Phenethyl Ester Induces Adrenoleukodystrophy (Abcd2) Gene in Human X-ALD Fibroblasts and Inhibits the Proinflammatory Response in Abcd1/2 Silenced Mouse Primary Astrocytes. Biochim. Biophys. Acta 2013, 1831, 747–758. [Google Scholar] [CrossRef]
- Fontanilla, C.V.; Wei, X.; Zhao, L.; Johnstone, B.; Pascuzzi, R.M.; Farlow, M.R.; Du, Y. Caffeic Acid Phenethyl Ester Extends Survival of a Mouse Model of Amyotrophic Lateral Sclerosis. Neuroscience 2012, 205, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ilhan, A.; Akyol, O.; Gurel, A.; Armutcu, F.; Iraz, M.; Oztas, E. Protective Effects of Caffeic Acid Phenethyl Ester against Experimental Allergic Encephalomyelitis-Induced Oxidative Stress in Rats. Free Radic. Biol. Med. 2004, 37, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Serarslan, G.; Altuğ, E.; Kontas, T.; Atik, E.; Avci, G. Caffeic Acid Phenethyl Ester Accelerates Cutaneous Wound Healing in a Rat Model and Decreases Oxidative Stress. Clin. Exp. Dermatol. 2007, 32, 709–715. [Google Scholar] [CrossRef]
- dos Santos, J.S.; Monte-Alto-Costa, A. Caffeic Acid Phenethyl Ester Improves Burn Healing in Rats through Anti-Inflammatory and Antioxidant Effects. J. Burn Care Res. Off. Publ. Am. Burn Assoc. 2013, 34, 682–688. [Google Scholar] [CrossRef]
- Hassan, N.A.; El-Bassossy, H.M.; Mahmoud, M.F.; Fahmy, A. Caffeic Acid Phenethyl Ester, a 5-Lipoxygenase Enzyme Inhibitor, Alleviates Diabetic Atherosclerotic Manifestations: Effect on Vascular Reactivity and Stiffness. Chem. Biol. Interact. 2014, 213, 28–36. [Google Scholar] [CrossRef]
- Pittalà, V.; Salerno, L.; Romeo, G.; Acquaviva, R.; Di Giacomo, C.; Sorrenti, V. Therapeutic Potential of Caffeic Acid Phenethyl Ester (CAPE) in Diabetes. Curr. Med. Chem. 2018, 25, 4827–4836. [Google Scholar] [CrossRef]
- Sorrenti, V.; Raffaele, M.; Vanella, L.; Acquaviva, R.; Salerno, L.; Pittalà, V.; Intagliata, S.; Di Giacomo, C. Protective Effects of Caffeic Acid Phenethyl Ester (CAPE) and Novel Cape Analogue as Inducers of Heme Oxygenase-1 in Streptozotocin-Induced Type 1 Diabetic Rats. Int. J. Mol. Sci. 2019, 20, 2441. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Choi, S.-S.; Kim, E.S.; Jedrychowski, M.P.; Yang, Y.R.; Jang, H.-J.; Suh, P.-G.; Banks, A.S.; Gygi, S.P.; Spiegelman, B.M. Thrap3 Docks on Phosphoserine 273 of PPARγ and Controls Diabetic Gene Programming. Genes Dev. 2014, 28, 2361–2369. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Roos, T.U.; Heiss, E.H.; Schwaiberger, A.V.; Schachner, D.; Sroka, I.M.; Oberan, T.; Vollmar, A.M.; Dirsch, V.M. Caffeic Acid Phenethyl Ester Inhibits PDGF-Induced Proliferation of Vascular Smooth Muscle Cells via Activation of P38 MAPK, HIF-1α, and Heme Oxygenase-1. J. Nat. Prod. 2011, 74, 352–356. [Google Scholar] [CrossRef]
- Chan, G.C.-F.; Cheung, K.-W.; Sze, D.M.-Y. The Immunomodulatory and Anticancer Properties of Propolis. Clin. Rev. Allergy Immunol. 2013, 44, 262–273. [Google Scholar] [CrossRef]
- Yasui, N.; Nishiyama, E.; Juman, S.; Negishi, H.; Miki, T.; Yamori, Y.; Ikeda, K. Caffeic Acid Phenethyl Ester Suppresses Oxidative Stress in 3T3-L1 Adipocytes. J. Asian Nat. Prod. Res. 2013, 15, 1189–1196. [Google Scholar] [CrossRef]
- Kazancioglu, H.O.; Bereket, M.C.; Ezirganli, S.; Aydin, M.S.; Aksakalli, S. Effects of Caffeic Acid Phenethyl Ester on Wound Healing in Calvarial Defects. Acta Odontol. Scand. 2015, 73, 21–27. [Google Scholar] [CrossRef]
- Kızıldağ, A.; Arabacı, T.; Albayrak, M.; Taşdemir, U.; Şenel, E.; Dalyanoglu, M.; Demirci, E. Therapeutic Effects of Caffeic Acid Phenethyl Ester on Alveolar Bone Loss in Rats with Endotoxin-Induced Periodontitis. J. Dent. Sci. 2019, 14, 339–345. [Google Scholar] [CrossRef]
- Krol, W.; Scheller, S.; Shani, J.; Pietsz, G.; Czuba, Z. Synergistic Effect of Ethanolic Extract of Propolis and Antibiotics on the Growth of Staphylococcus Aureus. Arzneimittelforschung 1993, 43, 607–609. [Google Scholar]
- Scheller, S.; Dworniczak, S.; Waldemar-Klimmek, K.; Rajca, M.; Tomczyk, A.; Shani, J. Synergism between Ethanolic Extract of Propolis (EEP) and Anti-Tuberculosis Drugs on Growth of Mycobacteria. Z. Naturforsch. C. 1999, 54, 549–553. [Google Scholar] [CrossRef]
- Veloz, J.J.; Alvear, M.; Salazar, L.A. Antimicrobial and Antibiofilm Activity against Streptococcus Mutans of Individual and Mixtures of the Main Polyphenolic Compounds Found in Chilean Propolis. BioMed Res. Int. 2019, 2019, 7602343. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wang, K.; Zheng, S.; Wang, Y.; Ren, Q.; Li, H.; Ding, L.; Li, W.; Zhang, L. Antibacterial Effect of Caffeic Acid Phenethyl Ester on Cariogenic Bacteria and Streptococcus Mutans Biofilms. Antimicrob. Agents Chemother. 2020, 64, e00251-20. [Google Scholar] [CrossRef]
- Chen, T.-G.; Lee, J.-J.; Lin, K.-H.; Shen, C.-H.; Chou, D.-S.; Sheu, J.-R. Antiplatelet Activity of Caffeic Acid Phenethyl Ester Is Mediated through a Cyclic GMP-Dependent Pathway in Human Platelets. Chin. J. Physiol. 2007, 50, 121–126. [Google Scholar] [PubMed]
- Jung, B.; Kim, M.; Kim, H.-A.; Kim, D.; Yang, J.; Her, S.; Song, Y.S. Caffeic Acid Phenethyl Ester, a Component of Beehive Propolis, Is a Novel Selective Estrogen Receptor Modulator. Phytother. Res. 2010, 24, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Tomur, A.; Kanter, M.; Gurel, A.; Erboga, M. The Efficiency of CAPE on Retardation of Hepatic Fibrosis in Biliary Obstructed Rats. J. Mol. Histol. 2011, 42, 451–458. [Google Scholar] [CrossRef]
- Cheng, C.-C.; Chi, P.-L.; Shen, M.-C.; Shu, C.-W.; Wann, S.-R.; Liu, C.-P.; Tseng, C.-J.; Huang, W.-C. Caffeic Acid Phenethyl Ester Rescues Pulmonary Arterial Hypertension through the Inhibition of AKT/ERK-Dependent PDGF/HIF-1α In Vitro and In Vivo. Int. J. Mol. Sci. 2019, 20, 1468. [Google Scholar] [CrossRef]
- Ang, E.S.M.; Pavlos, N.J.; Chai, L.Y.; Qi, M.; Cheng, T.S.; Steer, J.H.; Joyce, D.A.; Zheng, M.H.; Xu, J. Caffeic Acid Phenethyl Ester, an Active Component of Honeybee Propolis Attenuates Osteoclastogenesis and Bone Resorption via the Suppression of RANKL-Induced NF-KappaB and NFAT Activity. J. Cell. Physiol. 2009, 221, 642–649. [Google Scholar] [CrossRef]
- Tolba, M.F.; El-Serafi, A.T.; Omar, H.A. Caffeic Acid Phenethyl Ester Protects against Glucocorticoid-Induced Osteoporosis in Vivo: Impact on Oxidative Stress and RANKL/OPG Signals. Toxicol. Appl. Pharmacol. 2017, 324, 26–35. [Google Scholar] [CrossRef]
- Ha, J.; Choi, H.-S.; Lee, Y.; Lee, Z.H.; Kim, H.-H. Caffeic Acid Phenethyl Ester Inhibits Osteoclastogenesis by Suppressing NF KappaB and Downregulating NFATc1 and C-Fos. Int. Immunopharmacol. 2009, 9, 774–780. [Google Scholar] [CrossRef]
- Morroni, F.; Sita, G.; Graziosi, A.; Turrini, E.; Fimognari, C.; Tarozzi, A.; Hrelia, P. Neuroprotective Effect of Caffeic Acid Phenethyl Ester in A Mouse Model of Alzheimer’s Disease Involves Nrf2/HO-1 Pathway. Aging Dis. 2018, 9, 605–622. [Google Scholar] [CrossRef]
- Colpan, R.D.; Erdemir, A. Co-Delivery of Quercetin and Caffeic-Acid Phenethyl Ester by Polymeric Nanoparticles for Improved Antitumor Efficacy in Colon Cancer Cells. J. Microencapsul. 2021, 38, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, Y.; Aluani, D.; Tzankova, V.; Rangelov, S.; Odzhakov, F.; Apostolov, A.; Yoncheva, K. Safety Assessment of a Newly Synthesized Copolymer for Micellar Delivery of Hydrophobic Caffeic Acid Phenethyl Ester. Pharm. Dev. Technol. 2020, 25, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Fakhri, A.; Agarwal, S.; Ahmadi, E.; Nejad, P.A. Synthesis and Characterization of MnO(2)/NiO Nanocomposites for Photocatalysis of Tetracycline Antibiotic and Modification with Guanidine for Carriers of Caffeic Acid Phenethyl Ester-an Anticancer Drug. J. Photochem. Photobiol. B. 2017, 174, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-H.; Lee, D.-Y.; Kang, S.; Lee, M.-K.; Lee, J.-H.; Lee, S.-H.; Lee, H.-L.; Lee, H.-Y.; Jeong, Y.-I. Caffeic Acid Phenethyl Ester-Incorporated Radio-Sensitive Nanoparticles of Phenylboronic Acid Pinacol Ester-Conjugated Hyaluronic Acid for Application in Radioprotection. Int. J. Mol. Sci. 2021, 22, 6347. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-Y.; Jeong, Y.-I.; Kim, E.J.; Lee, K.D.; Choi, S.-H.; Kim, Y.J.; Kim, D.H.; Choi, K.-C. Preparation of Caffeic Acid Phenethyl Ester-Incorporated Nanoparticles and Their Biological Activity. J. Pharm. Sci. 2015, 104, 144–154. [Google Scholar] [CrossRef]
- Wang, J.; Bhargava, P.; Yu, Y.; Sari, A.N.; Zhang, H.; Ishii, N.; Yan, K.; Zhang, Z.; Ishida, Y.; Terao, K.; et al. Novel Caffeic Acid Phenethyl Ester-Mortalin Antibody Nanoparticles Offer Enhanced Selective Cytotoxicity to Cancer Cells. Cancers 2020, 12, 2370. [Google Scholar] [CrossRef]
- Kapare, H.S.; Lohidasan, S.; Sinnathambi, A.; Mahadik, K. Formulation Development of Folic Acid Conjugated PLGA Nanoparticles for Improved Cytotoxicity of Caffeic Acid Phenethyl Ester. Pharm. Nanotechnol. 2021, 9, 111–119. [Google Scholar] [CrossRef]
- Lu, H.; Li, S.; Dai, D.; Zhang, Q.; Min, Z.; Yang, C.; Sun, S.; Ye, L.; Teng, C.; Cao, X.; et al. Enhanced Treatment of Cerebral Ischemia-Reperfusion Injury by Intelligent Nanocarriers through the Regulation of Neurovascular Units. Acta Biomater. 2022, 147, 314–326. [Google Scholar] [CrossRef]
- Shahin, N.N.; Shamma, R.N.; Ahmed, I.S. A Nano-Liposomal Formulation of Caffeic Acid Phenethyl Ester Modulates Nrf2 and NF-Κβ Signaling and Alleviates Experimentally Induced Acute Pancreatitis in a Rat Model. Antioxidants 2022, 11, 1536. [Google Scholar] [CrossRef]
- Moriguchi, S.; Inagaki, R.; Saito, T.; Saido, T.C.; Fukunaga, K. Propolis Promotes Memantine-Dependent Rescue of Cognitive Deficits in APP-KI Mice. Mol. Neurobiol. 2022, 59, 4630–4646. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Concentration of CAPE (mg g−1) | Reference |
|---|---|---|
| Populus buds | 1.611 ± 0.272 | [25] |
| Propolis from Mexico (Ures) | 11.4 | [26] |
| Propolis from Spain (Viveros) | 10.1 | [27] |
| Propolis from Spain (Valencia) | 10.4 | [27] |
| Propolis from Chile | 5.8 | [27] |
| Propolis from China | 4.9 | [27] |
| Pharmacological Effect | Molecular Mechanism | Reference |
|---|---|---|
| Wound repair | CAPE promotes early inflammatory response (increased NOS2, TNF-α, and NF-κB) associated with a short-term event, leads to fast skin-wound healing and inhibition of inflammation. Significant increase in the glutathione (GSH) level, an endogenous antioxidant that plays a key role in cellular defense against oxidative stress. Considerable decrease in malondialdehyde (MDA) and superoxide dismutase activity. | [100,101] |
| Antidiabetic properties | CAPE is a heme oxygenase 1 (HO-1) inducer, combating the increase of reactive oxygen species (ROS) induced by hyperglycemia. It inhibits the 5-lipoxygenase, alleviating diabetic atherosclerotic symptoms (an important macrovascular complication of diabetes). It restores adipocyte function by increasing adiponectin and PPARγ (their activation leads to improved insulin sensitivity), leading to the reduction of proinflammatory factors. | [102,103,104,105] |
| Anticancer properties | Inhibition of DNA synthesis, disruption of growth signal transmission, induction of apoptosis through an internal apoptotic pathway, and promotion of anti-angiogenic effects. It enhances the anti-cancer effect of first-line chemotherapy drugs, an example being the drug paclitaxel in a rat model of DMBA-induced breast cancer, resulting in lower tumor weights compared to those with paclitaxel alone. CAPE protects normal cells against the effects of anticancer drugs by acting as a chemopreventive agent. The drug irinotecan protects normal blood, liver, and kidney cells without affecting the cytotoxicity of irinotecan in the in vivo model of Ehrlich ascites tumor cells. | [94,106,107,108,109] |
| Periodontitis Treatment | It improves bone healing, preventing RANKL-induced osteoclastogenesis (causes follicular bone destruction), suggesting its use as a regenerative agent in therapy of bone resorption. | [110,111] |
| Antibacterial activity | High activity against Mycobactrium tuberculosis, Mycobacterium avium, Streptococcus piogenes Klebsiella pneumoniae and Staphylococcus epidermidis. It presents synergistic activity with many antituberculosis drugs, including rifamycin, streptomycin, and isoniazid, and antibiotics such as gentamicin, tetracycline, chloramphenicol, vancomycin, clindamycin, netilmicin. CAPE inhibits biofilm formation and lactic acid and extracellular polysaccharide production in S. mutans. | [112,113,114,115] |
| Antiatherogenic effects | In human platelets, CAPE (15 and 25 µM) markedly inhibits platelet aggregation stimulated by collagen (2 µg/mL). | [116] |
| Estrogenic effects | CAPE is a selective agonist of ER-β estrogen receptors (present in the lungs, blood vessels, brain, and bones). | [117] |
| Hepatoprotective effects | In the liver, CAPE elevates tissue catalase (CAT) activity and ameliorates ultrastructural changes associated with aging; it also gives protection against necrosis, lipid peroxidation, aberrant cell proliferation, and p65 activation (decreased number of preneoplastic nodules and reduced incidence of liver tumors). Reduced malondialdehyde (MDA) levels and increased activities of glutathione peroxidase (GPx), together with superoxide dismutase (SOD) in liver tissue. | [77,118] |
| Lung protection | Effects on pulmonary arterial hypertension and fibrosis by HIF-1α/platelet-derived growth factor (PDGF)-dependent Akt/ERK pathways. | [119] |
| Protection of the bone diseases | CAPE induces osteoclast apoptosis, antioxidant effects and the modulation of osteoprotegrin signaling pathways. By suppressing NF-κB activity, CAPE significantly inhibits osteoclastogenesis and osteoclast differentiation. | [120,121,122] |
| Alzheimer’s disease | CAPE significantly inhibits neuronal apoptosis and neuroinflammation, induces Nrf2 activation, inhibits glycogen synthase kinase 3β in the hippocampus, and improves learning and memory and cognition. | [123] |
| Formulation | Target | Results | Reference |
|---|---|---|---|
| Poly(lactic-co-glycolic acid) (PLGA) co-loaded nanoparticles (QuCaNP) quercetin and CAPE | Improving anticancer efficacy in HT-29 human colorectal carcinoma. | Increased caspase-3 (2.38-fold) and caspase-9 (2-fold) mRNA levels and expressions of key proteins in the intrinsic apoptosis pathway in HT-29 cells. | [124] |
| Copolymer: polyglycerol and poly(allyl glycidyl ether) (C12-PAGE-PG), loaded with CAPE | To evaluate the in vitro and in vivo safety of a CAPE-loaded micellar system as a drug delivery platform on HepG2 cells. | Empty micelles loaded with CAPE showed no cytotoxic effects and retained the cytotoxic activity of CAPE loaded in the micelles, making it a good strategy to use this hydrophobic compound and improve the effectiveness of the treatments. | [125] |
| NiO nanoparticles and MnO2/NiO nanocomposites with guanidine and CAPE as a carrier | Evaluate its capacity as an anchoring method for drug carriers. | The drug loading time was 100 min and drug release in 1–10 h with 20–80% drug release. | [126] |
| Hyaluronic acid (HA) conjugated with phenylboronic acid pinacol ester (PBPE) with radiosensitive delivery of CAPE | Manufacture radiosensitive delivery of CAPE for application in radioprotection. | Prevention of radiation-induced apoptosis and intracellular ROS accumulation, with increased survivability of mice against radiation-induced death. | [127] |
| Methoxy poly(ethylene glycol)-b -poly(ε-caprolactone) copolymer nanoparticles (CE) with CAPE. | Study antitumor activity against lung metastasis by CT26 colon carcinoma cells. | Superior anti-metastatic efficacy against the tumor than CAPE itself. | [128] |
| Caffeic acid phenethyl ester-morphthalin antibody nanoparticles | Generate a potent anticancer drug by recruiting an anti-mortalin antibody (hsp70 chaperone that is enriched on the cancer cell surface). | Enhanced growth arrest/apoptosis of cancer cells through down-regulation of cyclin D1-CDK4, phospho-Rb, PARP-1 and the anti-apoptotic protein Bcl2. Significantly increased expression of p53, p21 WAF1, and caspase cleavage. Significantly improved down-regulation of proteins involved in cell migration. | [129] |
| Folic acid-conjugated PLGA nanoparticles. | Improve cytotoxicity, solubility, and achieve sustained release of CAPE. | It showed enhanced cytotoxicity in vivo and in vitro, causing a decrease in cell proliferation by 46%. | [130] |
| Stimuli-responsive liposomal nanocarrier loaded with CAPE modified with ac peptide (RGDyK) | Attack ischemic lesions and remodel neurovascular units (NVU) to reduce the progression of brain injury. | Drugs release in response to pathological signaling stimuli, localization of cerebral ischemia-reperfusion injury, and remodeling of neurovascular units by reducing neuronal apoptosis, regulating microglia polarization and repair of vascular endothelial cells. | [131] |
| Nano-Liposomal Formulation of CAPE | New strategies for acute pancreatitis treatment, evaluated in rat model | CAPE-loaded-NL showed better antioxidant, anti-inflammatory, and anti-apoptotic effects than free CAPE | [132] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez, R.; Burgos, V.; Marín, V.; Camins, A.; Olloquequi, J.; González-Chavarría, I.; Ulrich, H.; Wyneken, U.; Luarte, A.; Ortiz, L.; et al. Caffeic Acid Phenethyl Ester (CAPE): Biosynthesis, Derivatives and Formulations with Neuroprotective Activities. Antioxidants 2023, 12, 1500. https://doi.org/10.3390/antiox12081500
Pérez R, Burgos V, Marín V, Camins A, Olloquequi J, González-Chavarría I, Ulrich H, Wyneken U, Luarte A, Ortiz L, et al. Caffeic Acid Phenethyl Ester (CAPE): Biosynthesis, Derivatives and Formulations with Neuroprotective Activities. Antioxidants. 2023; 12(8):1500. https://doi.org/10.3390/antiox12081500
Chicago/Turabian StylePérez, Rebeca, Viviana Burgos, Víctor Marín, Antoni Camins, Jordi Olloquequi, Iván González-Chavarría, Henning Ulrich, Ursula Wyneken, Alejandro Luarte, Leandro Ortiz, and et al. 2023. "Caffeic Acid Phenethyl Ester (CAPE): Biosynthesis, Derivatives and Formulations with Neuroprotective Activities" Antioxidants 12, no. 8: 1500. https://doi.org/10.3390/antiox12081500