

Inhibition of α-Amylase, α-Glucosidase, Pancreatic Lipase, 15-Lipooxygenase and Acetylcholinesterase Modulated by Polyphenolic Compounds, Organic Acids, and Carbohydrates of Prunus domestica Fruit

Abstract
:1. Introduction
2. Methods and Materials
2.1. Plant Material
2.2. Sample Preparation
2.3. Identification and Quantification of Polyphenolic Compounds
2.4. Analysis of Polymeric Procyanidins by Phloroglucinolysis
2.5. Organic Acid and Carbohydrate Content
2.6. Analysis of Biological Activity
2.6.1. Analysis of Antioxidant Activities of ABTSo+ and FRAP
2.6.2. Inhibition of α-Amylase, α-Glucosidase, and Pancreatic Lipase
2.6.3. Inhibition of 15-Lipoxygenase
2.6.4. Inhibition of Acetylcholinesterase (AChE) and Butyrylcholinesterase (BuChE)
2.7. Statistical Analysis
3. Results and Discussion
3.1. Content of Carbohydrates and Organic Acids
3.2. Identification and Quantification of Polyphenolic Compounds in P. domestica Fruits
3.3. Antioxidant Activity
3.3.1. Antioxidant Capacity
3.3.2. Antidiabetic and Antiobesity Properties and Inhibition of Lipoxygenase
3.3.3. Inhibition of AChE and BuChE Activity
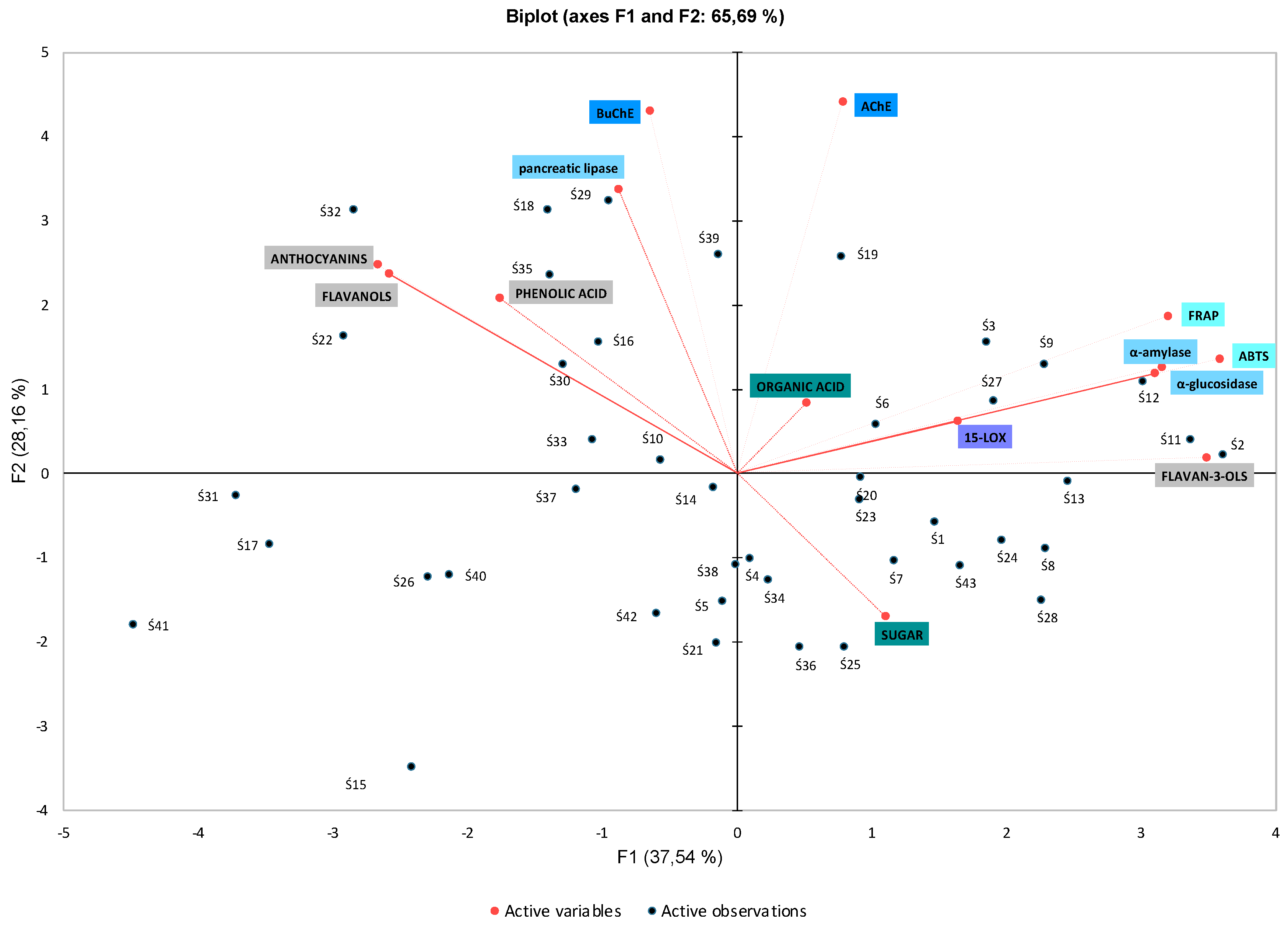
3.4. The Elements of Primary Component Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bahrin, A.A.; Moshawih, S.; Dhaliwal, J.S.; Kanakal, M.M.; Khan, A.; Lee, K.S.; Goh, B.H.; Goh, H.P.; Kifli, N.; Ming, L.C. Cancer Protective Effects of Plums: A Systematic Review. Biomed. Pharmacother. 2022, 146, 112568. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Nan, G.; Nisar, M.F.; Wan, C. Chemical Constituents and Health Benefits of Four Chinese Plum Species. J. Food Qual. 2020, 2020, 8842506. [Google Scholar] [CrossRef]
- Borowska, A. Zmiany Na Rynku Śliwek w Polsce w Latach 2010–2016 Ze Szczególnym Uwzględnieniem Śliwek Regionalnych z Oznaczeniami Geograficznymi ChOG. Zesz. Nauk. SGGW-Ekon. I Organ. Gospod. Żywnościowej 2019, 124, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Sottile, F.; Caltagirone, C.; Giacalone, G.; Peano, C.; Barone, E. Unlocking Plum Genetic Potential: Where Are We At? Horticulturae 2022, 8, 128. [Google Scholar] [CrossRef]
- Ropelewska, E. Diversity of Plum Stones Based on Image Texture Parameters and Machine Learning Algorithms. Agronomy 2022, 12, 762. [Google Scholar] [CrossRef]
- Silvan, J.M.; Ciechanowska, A.M.; Martinez-Rodriguez, A.J. Modulation of Antibacterial, Antioxidant, and Anti-Inflammatory Properties by Drying of Prunus domestica L. Plum Juice Extracts. Microorganisms 2020, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- Soares Mateus, A.R.; Pena, A.; Sendón, R.; Almeida, C.; Nieto, G.A.; Khwaldia, K.; Sanches Silva, A. By-Products of Dates, Cherries, Plums and Artichokes: A Source of Valuable Bioactive Compounds. Trends Food Sci. Technol. 2023, 131, 220–243. [Google Scholar] [CrossRef]
- Jabeen, Q.; Aslam, N. The Pharmacological Activities of Prunes: The Dried Plums. J. Med. Plants Res. 2011, 5, 1508–1511. [Google Scholar]
- Fang, J.; Zhou, Q.; Liu, L.Z.; Xia, C.; Hu, X.; Shi, X.; Jiang, B.H. Apigenin Inhibits Tumor Angiogenesis through Decreasing HIF-1α and VEGF Expression. Carcinogenesis 2007, 28, 858–864. [Google Scholar] [CrossRef] [Green Version]
- Igwe, E.O.; Charlton, K.E. A Systematic Review on the Health Effects of Plums (Prunus domestica and Prunus salicina). Phytother. Res. 2016, 30, 701–731. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Bąbelewski, P. Phenolic and Carotenoid Profile of New Goji Cultivars and Their Anti-Hyperglycemic, Anti-Aging and Antioxidant Properties. J. Funct. Foods 2018, 48, 632–642. [Google Scholar] [CrossRef]
- Wojdyło, A.; Carbonell-Barrachina, Á.A.; Legua, P.; Hernández, F. Phenolic Composition, Ascorbic Acid Content, and Antioxidant Capacity of Spanish Jujube (Ziziphus jujube Mill.) Fruits. Food Chem. 2016, 201, 307–314. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J. Antioxidant Activity Modulated by Polyphenol Contents in Apple and Leaves during Fruit Development and Ripening. Antioxidants 2020, 9, 567. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Turkiewicz, I.P.; Tkacz, K. Profiling of Polyphenols by LC-QTOF/ESI-MS, Characteristics of Nutritional Compounds and In Vitro Effect on Pancreatic Lipase, α-Glucosidase, α-Amylase, Cholinesterase and Cyclooxygenase Activities of Sweet (Prunus avium) and Sour (P. cerasus) Cherries Leaves and Fruits. Ind. Crops Prod. 2021, 174, 114214. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Turkiewicz, I.P.; Tkacz, K.; Hernandez, F. Comparison of Bioactive Compounds and Health Promoting Properties of Fruits and Leaves of Apple, Pear and Quince. Sci. Rep. 2021, 11, 20253. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Carbonell-Barrachina, Á.A.; Hernández, F. Phenolic Compounds, Antioxidant and Antidiabetic Activity of Different Cultivars of Ficus carica L. Fruits. J. Funct. Foods 2016, 25, 421–432. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Tkacz, K.; Turkiewicz, I.P. Sprouts vs. Microgreens as Novel Functional Foods: Variation of Nutritional and Phytochemical Profiles and Their In Vitro Bioactive Properties. Molecules 2020, 25, 4648. [Google Scholar] [CrossRef]
- Tomić, J.; Štampar, F.; Glišić, I.; Jakopič, J. Phytochemical Assessment of Plum (Prunus domestica L.) Cultivars Selected in Serbia. Food Chem. 2019, 299, 125113. [Google Scholar] [CrossRef]
- García-Gómez, B.E.; Salazar, J.A.; Nicolás-Almansa, M.; Razi, M.; Rubio, M.; Ruiz, D.; Martínez-Gómez, P. Molecular Bases of Fruit Quality in Prunus Species: An Integrated Genomic, Transcriptomic, and Metabolic Review with a Breeding Perspective. Int. J. Mol. Sci. 2020, 22, 333. [Google Scholar] [CrossRef]
- Wu, H.; Xu, Y.; Wang, H.; Miao, Y.; Li, C.; Zhao, R.; Shi, X.; Wang, B. Physicochemical Characteristics, Antioxidant Activities, and Aroma Compound Analysis of Seven Peach Cultivars (Prunus persica L. Batsch) in Shihezi, Xinjiang. Foods 2022, 11, 2944. [Google Scholar] [CrossRef]
- Aprea, E.; Charles, M.; Endrizzi, I.; Laura Corollaro, M.; Betta, E.; Biasioli, F.; Gasperi, F. Sweet Taste in Apple: The Role of Sorbitol, Individual Sugars, Organic Acids and Volatile Compounds. Sci. Rep. 2017, 7, 44950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowicka, P.; Wojdyło, A.; Laskowski, P. Principal Component Analysis (PCA) of Physicochemical Compounds’ Content in Different Cultivars of Peach Fruits, Including Qualification and Quantification of Sugars and Organic Acids by HPLC. Eur. Food Res. Technol. 2019, 245, 929–938. [Google Scholar] [CrossRef] [Green Version]
- Colaric, M.; Veberic, R.; Stampar, F.; Hudina, M. Evaluation of Peach and Nectarine Fruit Quality and Correlations between Sensory and Chemical Attributes. J. Sci. Food Agric. 2005, 85, 2611–2616. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A.; Pogon, P. Charakterystyka Owoców Pigwowca Japońskiego i Derenia Jadalnego. Żywność Nauka Technol. Jakość 2010, 6, 100–108. [Google Scholar]
- Montenegro-Landívar, M.F.; Tapia-Quirós, P.; Vecino, X.; Reig, M.; Valderrama, C.; Granados, M.; Cortina, J.L.; Saurina, J. Polyphenols and Their Potential Role to Fight Viral Diseases: An Overview. Sci. Total Environ. 2021, 801, 149719. [Google Scholar] [CrossRef] [PubMed]
- Liaudanskas, M.; Okulevičiūtė, R.; Lanauskas, J.; Kviklys, D.; Zymonė, K.; Rendyuk, T.; Žvikas, V.; Uselis, N.; Janulis, V. Variability in the Content of Phenolic Compounds in Plum Fruit. Plants 2020, 9, 1611. [Google Scholar] [CrossRef]
- Nowicka, P.; Wojdyło, A.; Tkacz, K.; Turkiewicz, I.P. Quantitative and Qualitative Determination of Carotenoids and Polyphenolics Compounds in Selected Cultivars of Prunus persica L. and Their Ability to In Vitro Inhibit Lipoxygenase, Cholinoesterase, α-Amylase, α-Glucosidase and Pancreatic Lipase. Food Chem. X 2023, 17, 100619. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, L.; Yang, C.; Li, Z.; Rong, S. Procyanidins and Alzheimer’s Disease. Mol. Neurobiol. 2019, 56, 5556–5567. [Google Scholar] [CrossRef]
- Valencia-Hernandez, L.J.; Wong-Paz, J.E.; Ascacio-Valdés, J.A.; Chávez-González, M.L.; Contreras-Esquivel, J.C.; Aguilar, C.N. Procyanidins: From Agro-Industrial Waste to Food as Bioactive Molecules. Foods 2021, 10, 3152. [Google Scholar] [CrossRef]
- Navarro-Orcajada, S.; Matencio, A.; Vicente-Herrero, C.; García-Carmona, F.; López-Nicolás, J.M. Study of the Fluorescence and Interaction between Cyclodextrins and Neochlorogenic Acid, in Comparison with Chlorogenic Acid. Sci. Rep. 2021, 11, 3275. [Google Scholar] [CrossRef]
- Popović, B.M.; Blagojević, B.; Ždero Pavlović, R.; Mićić, N.; Bijelić, S.; Bogdanović, B.; Mišan, A.; Duarte, C.M.M.; Serra, A.T. Comparison between Polyphenol Profile and Bioactive Response in Blackthorn (Prunus spinosa L.) Genotypes from North Serbia-from Raw Data to PCA Analysis. Food Chem. 2020, 302, 125373. [Google Scholar] [CrossRef]
- Fang, N.; Yu, S.; Prior, R.L. LC/MS/MS Characterization of Phenolic Constituents in Dried Plums. J. Agric. Food Chem. 2002, 50, 3579–3585. [Google Scholar] [CrossRef]
- Yang, D.; Wang, T.; Long, M.; Li, P. Quercetin: Its Main Pharmacological Activity and Potential Application in Clinical Medicine. Oxidative Med. Cell. Longev. 2020, 8825387. [Google Scholar] [CrossRef]
- Patel, R.V.; Mistry, B.M.; Shinde, S.K.; Syed, R.; Singh, V.; Shin, H.S. Therapeutic Potential of Quercetin as a Cardiovascular Agent. Eur. J. Med. Chem. 2018, 155, 889–904. [Google Scholar] [CrossRef]
- Lupo, G.; Cambria, M.T.; Olivieri, M.; Rocco, C.; Caporarello, N.; Longo, A.; Zanghì, G.; Salmeri, M.; Foti, M.C.; Anfuso, C.D. Anti-angiogenic Effect of Quercetin and Its 8-methyl Pentamethyl Ether Derivative in Human Microvascular Endothelial Cells. J. Cell Mol. Med. 2019, 23, 6565–6577. [Google Scholar] [CrossRef] [Green Version]
- Ben Salem, I.; Prola, A.; Boussabbeh, M.; Guilbert, A.; Bacha, H.; Lemaire, C.; Abid-Essefi, S. Activation of ER Stress and Apoptosis by α- and β-Zearalenol in HCT116 Cells, Protective Role of Quercetin. Neurotoxicology 2016, 53, 334–342. [Google Scholar] [CrossRef]
- Sharma, S.; Dahiya, A.; Kumar, S.; Verma, Y.K.; Dutta, A. Quercetin 3-O-Rutinoside Prevents Radiation Induced Oxidative Damage and Inflammation by Coordinated Regulation of Nrf2/NF-ΚB/NLRP3- Inflammasome Signaling in Gastrointestine. Phytomed. Plus 2023, 3, 100385. [Google Scholar] [CrossRef]
- Fitzpatrick, L.R.; Woldemariam, T. Small-Molecule Drugs for the Treatment of Inflammatory Bowel Disease. In Comprehensive Medicinal Chemistry III; Elsevier: Amsterdam, The Netherlands, 2017; pp. 495–510. [Google Scholar] [CrossRef]
- Kaur, S.; Tiwari, V.; Kumari, A.; Chaudhary, E.; Sharma, A.; Ali, U.; Garg, M. Protective and Defensive Role of Anthocyanins under Plant Abiotic and Biotic Stresses: An Emerging Application in Sustainable Agriculture. J. Biotechnol. 2023, 361, 12–29. [Google Scholar] [CrossRef]
- Michalska, A.; Łysiak, G. Przydatność do Suszenia Owoców Śliw Uprawianych w Polsce w Aspekcie Przemian Związków Bioaktywnych i Tworzących się Produktów Reakcji Maillarda. Żywność. Nauka. Technol. Jakość Food. Sci. Technol. Qual. 2014, 21, 29–38. [Google Scholar] [CrossRef]
- Liu, H.K.; Kang, Y.F.; Zhao, X.Y.; Liu, Y.P.; Zhang, X.W.; Zhang, S.J. Effects of Elicitation on Bioactive Compounds and Biological Activities of Sprouts. J. Funct. Foods 2019, 53, 136–145. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Laskowski, P.; Oszmiański, J. Evaluation of Sour Cherry (Prunus cerasus L.) Fruits for Their Polyphenol Content, Antioxidant Properties, and Nutritional Components. J. Agric. Food Chem. 2014, 62, 12332–12345. [Google Scholar] [CrossRef] [PubMed]
- Teleszko, M.; Wojdyło, A. Comparison of Phenolic Compounds and Antioxidant Potential between Selected Edible Fruits and Their Leaves. J. Funct. Foods 2015, 14, 736–746. [Google Scholar] [CrossRef]
- Yusuf, E.; Wojdyło, A.; Oszmiański, J.; Nowicka, P. Nutritional, Phytochemical Characteristics and In Vitro Effect on α-Amylase, α-Glucosidase, Lipase, and Cholinesterase Activities of 12 Coloured Carrot Varieties. Foods 2021, 10, 808. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Kumar, V.; Nayak, S.K.; Wadhwa, P.; Kaur, P.; Sahu, S.K. Alpha-Amylase as Molecular Target for Treatment of Diabetes Mellitus: A Comprehensive Review. Chem. Biol. Drug Des. 2021, 98, 539–560. [Google Scholar] [CrossRef]
- De Sales, P.M.; de Souza, P.M.; Simeoni, L.A.; Magalhães, P.d.O.; Silveira, D. α-Amylase Inhibitors: A Review of Raw Material and Isolated Compounds from Plant Source. J. Pharm. Pharm. Sci. 2012, 15, 141–183. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Kwon, C.S.; Son, K.H. Inhibition of Alpha-Glucosidase and Amylase by Luteolin, a Flavonoid. Biosci. Biotechnol. Biochem. 2000, 64, 2458–2461. [Google Scholar] [CrossRef]
- Zhang, X.; Su, M.; Du, J.; Zhou, H.; Li, X.; Li, X.; Ye, Z. Comparison of Phytochemical Differences of the Pulp of Different Peach [Prunus persica (L.) Batsch] Cultivars with Alpha-Glucosidase Inhibitory Activity Variations in China Using UPLC-Q-TOF/MS. Molecules 2019, 24, 1968. [Google Scholar] [CrossRef] [Green Version]
- Wojdyło, A.; Turkiewicz, I.P.; Tkacz, K.; Nowicka, P.; Bobak, Ł. Nuts as Functional Foods: Variation of Nutritional and Phytochemical Profiles and Their in Vitro Bioactive Properties. Food Chem. X 2022, 15, 100418. [Google Scholar] [CrossRef]
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P.; Golis, T.; Bąbelewski, P. ABTS On-Line Antioxidant, α-Amylase, α-Glucosidase, Pancreatic Lipase, Acetyl- and Butyrylcholinesterase Inhibition Activity of Chaenomeles Fruits Determined by Polyphenols and Other Chemical Compounds. Antioxidants 2020, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Lončarić, M.; Strelec, I.; Moslavac, T.; Šubarić, D.; Pavić, V.; Molnar, M. Lipoxygenase Inhibition by Plant Extracts. Biomolecules 2021, 11, 152. [Google Scholar] [CrossRef]
- Wszelaki, N. Plants as a source of acetylcholinesterase and butyrylcholinesterase inhibitors. Postępy Fitoter. 2009, 24–38. [Google Scholar]
- Bhullar, K.S.; Rupasinghe, H.P.V. Polyphenols: Multipotent Therapeutic Agents in Neurodegenerative Diseases. Oxidative Med. Cell. Longev. 2013, 2013, 891748. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Liu, M.; Wang, F.; Cai, J.; Luo, Q.; Li, S.; Zhu, J.; Tang, Z.; Fang, Z.; Wang, C.; et al. The Inhibition Mechanism of Polyphenols from Phyllanthus Emblica Linn. Fruit on Acetylcholinesterase: A Interaction, Kinetic, Spectroscopic, and Molecular Simulation Study. Food Res. Int. 2022, 158, 111497. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sample | Carbohydrates † | Organic Acid † | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fructose | Sorbitol | Glucose | Saccharose | Total | Oxalic Acid | Citric Acid | Malic Acid | Quinic Acid | Succinic Acid | Formic Acid | Total | |
| Ś1 | 3.52 ± 0.35 g–j | 0.69 ± 0.11 g–j | 2.94 ± 0.12 f–l | 1.84 ± 0.23 n–r | 8.98 i–l | 0.15 ± 0.02 k–n | nd f | 1.24 ± 0.21 g–l | 0.43 ± 0.10 m–p | 0.09 ± 0.03 ef | 0.06 ± 0.01 d | 1.97 h–o |
| Ś2 | 2.72 ± 0.54 k–p | nd r | 3.85 ± 0.21 b–d | 2.23 ± 0.18 l–p | 8.80 i–m | 0.17 ± 0.02 i–l | 0.07 ± 0.01 a | 1.01 ± 0.28 l–q | 0.42 ± 0.11 m–p | 0.07 ± 0.02 gh | 0.04 ± 0.01 f | 1.80 j–p |
| Ś3 | 3.21 ± 0.29 h–m | 1.04 ± 0.10 cd | 3.79 ± 0.15 b–d | 3.25 ± 0.32 k | 11.28 e–h | 0.32 ± 0.06 ef | 0.01 ± 0.00 e | 1.25 ± 0.11 g–l | 0.73 ± 0.01 g–j | 0.09 ± 0.03 ef | 0.08 ± 0.01 b | 2.49 b–f |
| Ś4 | 2.05 ± 0.32 p–s | 0.60 ± 0.09 j–m | 5.87 ± 0.14 a | 3.09 ± 0.33 kl | 11.61 d–h | 0.33 ± 0.04 e | 0.02 ± 0.00 d | 0.87 ± 0.21 p–s | 1.31 ± 0.26 cd | 0.09 ± 0.03 ef | 0.13 ± 0.03 a | 2.76 bc |
| Ś5 | 2.51 ± 0.10 m–r | 0.74 ± 0.04 g–i | 4.35 ± 0.26 b | 2.55 ± 0.12 k–o | 10.15 g–k | 0.16 ± 0.02 j–m | 0.01 ± 0.00 e | 0.76 ± 0.09 r–u | 0.61 ± 0.09 i–k | 0.05 ± 0.01 ij | 0.02 ± 0.00 h | 1.62 m–q |
| Ś6 | 2.06 ± 0.48 p–s | 1.61 ± 0.06 a | 3.48 ± 0.32 c–f | 5.31 ± 0.15 ij | 12.46 c–f | 0.28 ± 0.11 fg | 0.01 ± 0.00 e | 1.34 ± 0.19 e–j | 0.54 ± 0.11 k–n | 0.07 ± 0.02 gh | nd j | 2.24 d–j |
| Ś7 | 2.00 ± 0.49 p–s | 0.51 ± 0.08 l–n | 3.78 ± 0.54 b–e | 5.71 ± 0.67 h–j | 12.00 c–g | 0.21 ± 0.06 hi | 0.02 ± 0.00 d | 1.02 ± 0.11 l–q | 1.22 ± 0.11 d | 0.06 ± 0.01 hi | 0.07 ± 0.03 c | 2.61 b–d |
| Ś8 | 2.04 ± 0.19 p–s | 0.03 ± 0.00 qr | 3.21 ± 0.19 e–i | 5.32 ± 0.48 ij | 10.60 e–i | 0.28 ± 0.01 fg | 0.01 ± 0.00 e | 1.15 ± 0.19 i–o | 0.96 ± 0.16 e | 0.06 ± 0.03 hi | 0.08 ± 0.03 b | 2.56 b–e |
| Ś9 | 4.06 ± 0.20 fg | nd r | 1.81 ± 0.47 q–u | 5.89 ± 0.59 g–i | 11.76 d–h | 0.06 ± 0.11 rs | 0.01 ± 0.00 e | 1.52 ± 0.23 c–f | 0.77 ± 0.06 f–h | 0.07 ± 0.04 gh | 0.04 ± 0.01 f | 2.47 b–g |
| Ś10 | 5.32 ± 0.48 cd | nd r | 0.99 ± 0.11 w | 6.38 ± 0.72 e–h | 12.69 b–e | 0.09 ± 0.01 p–s | 0.01± 0.00 e | 1.19 ± 0.16 i–m | 0.20 ± 0.02 t | 0.07 ± 0.05 gh | 0.01 ± 0.00 i | 1.57 o–q |
| Ś11 | 2.88 ± 0.29 j–o | nd r | 2.98 ± 0.16 f–k | 8.75 ± 0.39 a | 14.61 ab | 0.35 ± 0.11 e | 0.02 ± 0.00 d | 1.23 ± 0.31 h–l | 0.80 ± 0.01 fg | 0.06 ± 0.01 hi | 0.04 ± 0.01 f | 2.50 b–f |
| Ś12 | 1.79 ± 0.34 rs | nd r | 1.92 ± 0.16 p–u | 8.46 ± 0.54 a | 12.17 c–g | 0.10 ± 0.02 o–r | nd f | 0.92 ± 0.21 n–r | 0.21 ± 0.04 st | 0.09 ± 0.03 ef | 0.02 ± 0.00 h | 1.34 q |
| Ś13 | 3.09 ± 0.24 h–n | 1.68 ± 0.02 a | 6.19 ± 0.54 a | 1.41 ± 0.28 p–s | 12.38 c–f | 0.25 ± 0.04 gh | 0.05 ± 0.01 b | 1.52 ± 0.15 c–f | 0.76 ± 0.16 f–i | 0.08 ± 0.01 fg | 0.08 ± 0.03 b | 2.74 bc |
| Ś14 | 1.97 ± 0.38 q–s | nd r | 1.58 ± 0.21 t–v | 7.54 ± 0.19 b–d | 11.09 e–h | 0.11 ± 0.05 n–q | 0.02 ± 0.00 d | 1.19 ± 0.32 i–m | 0.25 ± 0.05 r–t | 0.07 ± 0.01 gh | 0.03 ± 0.00 g | 1.68 k–q |
| Ś15 | 3.25 ± 0.18 h–l | nd r | 1.14 ± 0.21 vw | 2.96 ± 0.32 kl | 7.35 l–p | 0.36 ± 0.02 e | 0.04 ± 0.00 c | 0.61 ± 0.16 tu | 0.44 ± 0.19 l–o | 0.09 ± 0.03 ef | 0.04 ± 0.01 f | 1.60 n–q |
| Ś16 | 3.26 ± 0.43 h–l | 0.87 ± 0.03 ef | 3.09 ± 0.12 f–j | 1.73 ± 0.22 o–r | 8.96 i–l | 0.25 ± 0.02 gh | 0.01 ± 0.00 e | 1.32 ± 0.10 f–k | 0.41 ± 0.12 n–q | 0.07 ± 0.01 gh | 0.02 ± 0.00 h | 2.09 f–l |
| Ś17 | 1.50 ± 0.27 s | 0.47 ± 0.7 n | 1.89 ± 0.17 p–u | 1.85 ± 0.28 n–r | 5.71 p | 0.18 ± 0.01 i–l | 0.01 ± 0.00 e | 1.20 ± 0.25 i–l | 0.55 ± 0.11 k–n | 0.11 ± 0.01 cd | 0.08 ± 0.03 b | 2.15 e–j |
| Ś18 | 3.19 ± 0.29 h–m | 0.93 ± 0.11 de | 2.31 ± 0.12 m–r | 2.28 ± 0.32 l–p | 8.70 i–n | 0.49 ± 0.02 c | 0.01 ± 0.00 e | 1.76 ± 0.21 bc | 0.55 ± 0.01 k–n | 0.09 ± 0.02 ef | nd j | 2.90 b |
| Ś19 | 3.71 ± 0.19 f–i | 0.80 ± 0.37 fg | 2.97 ± 0.32 f–k | 2.21 ± 0.37 l–p | 9.70 h–k | 0.33 ± 0.03 e | 0.01 ± 0.00 e | 1.51 ± 0.22 d–f | 0.49 ± 0.15 k–o | 0.09 ± 0.02 ef | 0.02 ± 0.00 h | 2.45 b–g |
| Ś20 | 2.88 ± 0.39 j–o | 0.46 ± 0.29 no | 2.38 ± 0.18 l–q | 4.83 ± 0.47 j | 10.54 f–i | 0.15 ± 0.10 k–n | nd f | 0.74 ± 0.17 r–u | 1.00 ± 0.17 e | 0.04 ± 0.01 j | 0.02 ± 0.00 h | 1.96 h–o |
| Ś21 | 2.96 ± 0.25 j–n | 0.49 ± 0.05 mn | 2.75 ± 0.19 h–m | 1.25 ± 0.38 q–s | 7.44 l–p | 0.12 ± 0.13 m–p | 0.01 ± 0.00 e | 1.61 ± 0.21 cd | 0.28 ± 0.07 p–t | 0.10 ± 0.03 de | 0.02 ± 0.00 h | 2.14 e–j |
| Ś22 | 4.30 ± 0.50 ef | 0.34 ± 0.01 o | 1.98 ± 0.21 p–u | 1.54 ± 0.28 p–r | 8.16 k–o | 0.07 ± 0.11 q–s | 0.01 ± 0.00 e | 1.48 ± 0.15 d–g | 0.64 ± 0.16 h–k | 0.11 ± 0.03 cd | 0.02 ± 0.00 h | 2.33 c–h |
| Ś23 | 3.55 ± 0.19 g–j | 0.14 ± 0.02 pq | 3.29 ± 0.34 d–h | 2.84 ± 0.48 k–m | 9.82 h–k | 0.60 ± 0.23 a | 0.01 ± 0.00 e | 1.96 ± 0.18 ab | 1.44 ± 0.21 c | 0.07 ± 0.01 gh | 0.04 ± 0.00 f | 4.12 a |
| Ś24 | 3.02 ± 0.39 i–n | 0.52 ± 0.11 l–n | 1.78 ± 0.11 r–u | 2.99 ± 0.27 kl | 8.32 j–n | 0.42 ± 0.05 d | 0.01 ± 0.00 e | 1.34 ± 0.32 e–j | 0.57 ± 0.13 k–m | 0.11 ± 0.01 cd | 0.07 ± 0.03 c | 2.52 b–f |
| Ś25 | 5.30 ± 0.79 cd | 0.46 ± 0.05 no | 3.87 ± 0.32 bc | 6.89 ± 0.33 d–f | 16.51 a | 0.14 ± 0.07 l–o | 0.01 ± 0.00 e | 0.79 ± 0.17 q–t | 0.37 ± 0.09 o–r | 0.05 ± 0.03 ij | 0.01 ± 0.00 i | 1.38 pq |
| Ś26 | 3.32 ± 0.55 h–k | 1.15 ± 0.11 bc | 2.93 ± 0.21 f–l | 7.24 ± 0.52 c–e | 14.64 ab | 0.09 ± 0.02 p–s | 0.01 ± 0.00 e | 1.09 ± 0.21 k–p | 0.36 ± 0.08 o–s | 0.06 ± 0.03 hi | 0.04 ± 0.00 f | 1.66 l–q |
| Ś27 | 2.21 ± 0.41 o–s | 0.18 ± 0.03 p | 1.11 ± 0.22 vw | 2.64 ± 0.57 k–n | 6.14 op | 0.33 ± 0.03 e | 0.02 ± 0.00 d | 1.86 ± 0.11 ab | 1.82 ± 0.11 b | 0.12 ± 0.03 c | 0.04 ± 0.00 f | 4.19 a |
| Ś28 | 6.27 ± 0.26 b | 0.56 ± 0.02 k–n | 2.85 ± 0.31 g–m | 0.12 ± 0.02 u | 9.80 h–k | 0.12 ± 0.01 m–p | 0.02 ± 0.00 d | 0.91 ± 0.32 o–r | 0.36 ± 0.02 o–s | 0.05 ± 0.01 ij | 0.07 ± 0.08 c | 1.55 o–q |
| Ś29 | 3.21 ± 0.12 h–m | 0.18 ± 0.11 p | 1.44 ± 0.28 u–w | 2.03 ± 0.26 m–q | 6.85 m–p | 0.10 ± 0.01 o–r | 0.02 ± 0.00 d | 1.37 ± 0.12 d–j | 0.26 ± 0.03 q–t | 0.04 ± 0.01 j | 0.03 ± 0.00 g | 1.83 i–p |
| Ś30 | 4.80 ± 0.32 de | nd r | 1.73 ± 0.43 s–u | 8.14 ± 0.33 a–c | 14.66 ab | 0.19 ± 0.02 i–k | 0.02 ± 0.00 d | 1.57 ± 0.25 c–e | 0.25 ± 0.01 r–t | 0.10 ± 0.01 de | 0.01 ± 0.00 i | 2.14 e-j |
| Ś31 | 7.00 ± 0.12 a | 0.09 ± 0.11 p–r | 3.14 ± 0.66 f–j | 0.03 ± 0.00 u | 10.27 g–j | 0.10 ± 0.01 o–r | 0.02 ± 0.00 d | 0.83 ± 0.11 q–t | 0.45 ± 0.08 l–o | 0.05 ± 0.01 ij | 0.02 ± 0.00 h | 1.48 pq |
| Ś32 | 2.44 ± 0.38 n–r | nd r | 1.43 ± 0.28 u–w | 1.62 ± 0.02 p–r | 5.49 p | 0.08 ± 0.02 p–s | 0.01 ± 0.00 e | 0.95 ± 0.10 m–r | 0.90 ± 0.09 ef | 0.10 ± 0.03 de | 0.03 ± 0.00 g | 2.07 f–m |
| Ś33 | 5.66 ± 0.63 bc | 0.77 ± 0.11 f–h | 3.09 ± 0.10 f–j | 0.15 ± 0.01 tu | 9.67 h–k | 0.25 ± 0.05 gh | 0.01 ± 0.00 e | 1.16 ± 0.25 i–n | 0.75 ± 0.10 f–i | 0.11 ± 0.03 cd | 0.07 ± 0.03 c | 2.37 c–h |
| Ś34 | 7.55 ± 0.38 a | nd r | 3.43 ± 0.29 c–f | 1.11 ± 0.11 rs | 12.09 c–g | 0.05 ± 0.01 s | 0.01 ± 0.00 e | 0.54 ± 0.03 u | 0.63 ± 0.09 h–k | 0.12 ± 0.01 c | 0.04 ± 0.00 f | 1.41 pq |
| Ś35 | 2.57 ± 0.28 l–q | 0.67 ± 0.14 h–k | 2.06 ± 0.21 o–t | 0.59 ± 0.02 s–u | 5.90 p | 0.11 ± 0.01 n–q | 0.01 ± 0.00 e | 2.09 ± 0.27 a | 0.26 ± 0.08 q–t | 0.07 ± 0.02 gh | 0.07 ± 0.01 c | 2.62 b–d |
| Ś36 | 2.42 ± 0.72 n–r | 0.76 ± 0.09 f–h | 1.91 ± 0.21 p–u | 7.40 ± 0.43 cd | 12.49 c–f | 0.14 ± 0.02 l–o | 0.01 ± 0.00 e | 0.64 ± 0.04 s–u | 0.75 ± 0.17 f–i | 0.06 ± 0.03 hi | 0.05 ± 0.01 e | 1.66 l–q |
| Ś37 | 2.56 ± 0.28 l–q | nd r | 2.17 ± 0.32 n–s | 3.33 ± 0.16 k | 8.06 k–o | 0.11 ± 0.03 n–q | 0.02 ± 0.00 d | 0.81 ± 0.03 q–t | 0.59 ± 0.08 j–l | 0.07 ± 0.02 gh | 0.05 ± 0.01 e | 1.66 l–q |
| Ś38 | 3.36 ± 0.32 g–k | 1.24 ± 0.23 b | 2.71 ± 0.25 i–n | 6.69 ± 0.32 d–g | 14.00 bc | 0.17 ± 0.06 i–l | 0.01 ± 0.00 e | 1.25 ± 0.11 g–l | 0.43 ± 0.08 m–p | 0.11 ± 0.01 cd | 0.04 ± 0.01 f | 2.03 g–n |
| Ś39 | 3.30 ± 0.11 h–k | 1.05 ± 0.11 cd | 2.41 ± 0.21 k–p | 1.40 ± 0.37 p–s | 8.16 k–o | 0.16 ± 0.02 j–m | 0.01 ± 0.00 e | 1.09 ± 0.10 k–p | 1.04 ±0.12 e | 0.09 ± 0.01 ef | 0.02 ± 0.00 h | 2.41 c–h |
| Ś40 | 5.19 ± 0.51 cd | nd r | 0.06 ± 0.00 x | 8.40 ± 0.65 ab | 13.65 b–d | 0.19± 0.03 i–k | 0.01 ± 0.00 e | 1.39 ± 0.16 d–i | 0.40 ± 0.11 n–r | 0.07 ± 0.03 gh | 0.05 ± 0.01 e | 2.13 e–k |
| Ś41 | 3.30 ± 0.37 h–k | 0.15 ± 0.03 pq | 2.58 ± 0.21 j–o | 6.05 ± 0.87 f–i | 12.08 c–g | 0.20 ± 0.02 ij | 0.02 ± 0.00 d | 1.24 ± 0.11 g–l | 0.99 ± 0.03 e | 0.14 ± 0.01 b | 0.05 ± 0.01 e | 2.63 b–d |
| Ś42 | 2.56 ± 0.09 l–q | 0.62 ± 0.02 i–l | 2.42 ± 0.17 k–p | 1.04 ± 0.25 r–t | 6.63 n–p | 0.41 ± 0.03 d | nd f | 1.45 ± 0.18 d–h | 2.22 ± 0.32 a | 0.18 ± 0.03 a | 0.04 ± 0.00 f | 4.33 a |
| Ś43 | 3.80 ± 0.27 f–h | 0.71 ± 0.10 g–j | 3.38 ± 0.27 c–g | 5.69 ± 0.33 h–j | 13.59 b–d | 0.55 ± 0.00 b | nd f | 1.14 ± 0.11 j–o | 0.44 ± 0.11 l–o | 0.10 ± 0.03 de | 0.04 ± 0.00 f | 2.28 d–i |
| tR (min) | Assigned Identity | Molecular Ion [M-H]− (m/z) | Main Ions MS/MS (m/z) |
|---|---|---|---|
| Flavan-3-ols | |||
| 2.30 | (+)-Catechin † | 289.0143 | 245.0367 |
| 3.30 | Procyanidin dimer (B1) † | 577.0538 | 289.0143 |
| 4.50 | Procyanidin dimer (B3) † | 577.0311 | 289.0152 |
| Phenolic acids | |||
| 3.71 | Neochlorogenic acid † | 353.0532 | 191.0232 |
| 3.92 | Cryptochlorogenic acid † | 353.0534 | 191.0241 |
| 4.02 | Chlorogenic acid † | 353.0531 | 191.0254 |
| 4.83 | 3-Caffeoylshikimic acid | 335.0713 | 191.0511 |
| 5.02 | 3-Feruloylquinic acid | 367.2312 | 193.2101/191.2302 |
| Flavonols | |||
| 3.36 | Quercetin-3-O-pentoside-hexoside | 595.0411 | 449.0311/301.0062 |
| 3.60 | Quercetin-3-O-glucoside † | 463.0523 | 301.0062 |
| 4.20 | Quercetin-3-O-arabinoside † | 433.0302 | 301.0666 |
| 6.62 | Quercetin-3-O-rutinoside † | 609.1018 | 301.0133 |
| 6.78 | Quercetin-3-O-galactoside † | 463.0523 | 301.0054 |
| 7.32 | Quercetin-3-O-rhamnoside † | 447.0902 | 301.0054 |
| 10.22 | Quercetin-O-pentoside-rhamnoside | 579.0332 | 301.1803 |
| Anthocyanins | |||
| 4.59 | Cyanidin-3-O-galactoside † | 449.0324 | 287.0180 |
| 6.29 | Cyanidin-3-O-glucoside † | 449.0646 | 299.0216/287.0180 |
| 6.55 | Cyanidin-3-O-rutinoside † | 595.1507 | 287.0536 |
| 6.72 | Delphinidin-3-O-glucoside † | 465.0243 | 303.0098 |
| 6.86 | Peonidin-3-O-glucoside † | 463.1201 | 301.0721 |
| Sample | Flavan-3-Ols † | Phenolic Acid † | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Procyanidin B1 | Procyanidin B3 | (+)-Catechin | PP | Other | Total | Neochlorogenic Acid | Cryptochlorogenic Acid | Chlorogenic Acid | 3-Caffeoylshikimic Acid | 3-Feruloylquinic Acid | Other | Total | |||
| Ś1 | 11.59 ± 1.11 n–p | 4.15 ± 0.35 j–l | 4.25 ± 0.23 kl | 529.3 ± 12.3 f–h | 31.55 ± 4.21 e | 580.89 g–k | 269.11 ± 1.32 h–k | 9.45 ± 0.32 l–o | 33.32 ± 1.32 i–l | 5.95 ± 0.32 ef | 10.47 ± 0.32 d–f | 13.47 ± 0.43 j–m | 341.77 i–l | ||
| Ś2 | 24.84 ± 2.11 ef | 9.77 ± 0.34 d | nd u | 1022.7 ± 24.5 a | 57.15 ± 3.65 b | 1114.47 a | 219.20 ± 11.23 j–n | 10.08 ± 0.12 k–n | 24.48 ± 2.13 l–p | 2.90 ± 0.23 n–s | 4.92 ± 0.14 mn | 2.74 ± 0.53 t | 264.32 n–r | ||
| Ś3 | 8.88 ± 1.76 p–s | 2.54 ± 0.62 o–r | 2.56 ± 0.38 n–s | 618.2 ± 23.1 d–f | 52.43 ± 3.87 b | 684.68 d–h | 216.32 ± 2.64 k–n | 10.41± 0.32 k–n | 17.85 ± 1.56 p–t | 3.77 ± 0.54 k–n | 6.71 ± 0.43 i–l | 17.30 ± 0.41 g–j | 272.34 l–r | ||
| Ś4 | 19.01 ± 1.87 h–j | 5.17 ± 0.34 h–j | 7.57 ± 0.25 de | 353.3 ± 17.5 kl | 40.66 ± 2.54 cd | 425.77 k–o | 357.06 ± 4.44 fg | nd t | 35.73 ± 2.11 h–k | 4.86 ± 0.54 g–i | 1.24 ± 0.32 s–u | 19.34 ± 0.67 f–h | 418.23 f–h | ||
| Ś5 | 11.03 ±1.63 o–q | 2.51 ± 0.12 o–r | 3.23 ± 0.23 m–o | 283.7 ± 12.3 lm | 10.82 ± 3.28 p–t | 311.35 n–r | 275.01 ± 3.32 h–j | 7.94 ± 1.03 n–q | 40.10 ± 1.65 g–i | 3.58 ± 0.34 l–o | 1.11 ± 0.11 tu | 12.41 ± 0.29 k–n | 340.16 i–m | ||
| Ś6 | 15.59 ± 1.66 j–l | 3.81 ± 0.13 k–n | 5.60 ± 0.45 h–j | 490.2 ± 17.3 g–i | 37.44 ± 4.29 d | 552.70 g–l | 301.18 ± 2.22 g–i | 7.22 ± 0.99 n–r | 24.23 ±2.43 m–q | 1.63 ± 0.12 u | 2.50 ± 0.24 o–t | 11.85 ± 0.76 k–o | 348.60 h–k | ||
| Ś7 | 24.79 ± 2.11 ef | 6.21 ± 0.47 f–h | 9.35 ± 0.11 c | 238.9 ± 11.1 m–p | 13.48 ± 1.32 n–s | 292.80 o–s | 52.29 ± 2.43 st | 4.02 ± 0.65 rs | 35.41 ± 1.44 h–k | 2.54 ± 0.32 q–t | 9.91 ± 0.15 d–f | 12.03 ± 0.49 k–o | 116.20 wx | ||
| Ś8 | 9.89 ± 1.04 o–s | 3.31 ± 0.23 k–p | 4.26 ± 0.15 kl | 540.0 ± 13.2 f–h | 4.24 ± 1.32 u–w | 561.74 g–l | 188.29 ± 3.22 m–p | 4.02 ± 1.43 rs | 15.08 ± 2.12 r–v | 1.76 ± 0.11 tu | 3.04 ± 0.17 o–r | 5.10 ± 0.65 r–t | 217.30 q–t | ||
| Ś9 | 23.17 ± 1.00 fg | 6.15 ± 0.47 f–h | 7.17 ± 0.48 e–g | 741.3 ± 10.0 c | 20.06 ± 2.76 h–l | 797.94 cd | 360.55 ± 4.23 ef | 21.73 ± 0.32 e | 30.57 ± 1.87 j–n | 4.74 ± 0.43 h–j | 5.41 ± 0.42 lm | 21.98 ± 0.89 f | 444.97 e–g | ||
| Ś10 | 15.37 ± 1.62 j–m | 1.58 ± 0.21 r–t | 2.88 ± 0.12 m–r | 380.2 ± 13.7 jk | nd w | 400.12 l–p | 308.43 ± 1.43 f–h | 16.33 ± 1.32 f–h | 28.07 ± 1.84 j–o | 3.46 ± 0.34 l–p | 9.05 ± 0.18 fg | 18.19 ± 0.77 f–i | 383.53 g–j | ||
| Ś11 | 11.65 ± 1.01 m–p | 4.41 ± 0.43 i–k | 5.36 ± 0.43 ij | 686.6 ± 12.2 cd | 70.17 ± 4.10 a | 778.27 c–f | 463.37 ± 3.12 bc | 33.83 ± 0.87 bc | 34.56 ± 1.99 i–k | 2.53 ± 0.24 q–u | 6.01 ± 0.61 k–m | 17.89 ± 0.99 f–i | 558.19 cd | ||
| Ś12 | 48.75 ± 0.23 a | 11.51 ± 0.76 c | 11.36 ± 0.65 a | 857.0 ± 10.4 b | 52.55 ± 5.10 b | 981.24 ab | 92.73 ± 1.44 r–t | 4.08 ± 0.54 rs | 19.62 ± 1.63 o–s | 2.61 ± 0.43 p–t | 3.61 ± 0.36 n–p | 3.10 ± 0.38 t | 125.74 v–x | ||
| Ś13 | 16.32 ± 1.11 j–l | 2.62 ± 0.43 o–r | 3.55 ± 0.34 l–n | 494.1 ± 10.3 g–i | 17.70 ± 2.10 i–n | 534.35 h–l | 266.54 ± 2.54 h–k | 11.36 ± 1.32 j–m | 26.90 ± 1.54 k–o | 3.01 ± 0.37 m–r | 3.71 ± 0.28 no | 10.98 ± 0.56 l–p | 322.50 j–m | ||
| Ś14 | 23.16 ± 0.54 fg | 15.86 ± 0.32 b | nd u | 429.7 ± 9.9 i–k | 15.58 ± 2.31 k–q | 484.40 j–m | 171.97 ± 2.11 n–p | 5.51 ± 0.55 qr | 45.83 ± 1.11 fg | 2.74 ± 0.12 o–s | 10.71 ± 0.37 de | 9.29 ± 0.47 m–r | 246.04 p–t | ||
| Ś15 | 4.41 ± 0.32 tu | 1.77 ± 0.43 q–t | 1.86 ± 0.11 st | 202.3 ± 10.7 m–q | 22.78 ± 2.54 g–i | 233.18 p–t | 237.00 ± 0.32 j–m | 9.01 ± 0.99 l–p | 22.38 ± 1.76 n–r | 4.13 ± 0.46 i–l | 2.18 ± 0.19 p–t | 10.41 ± 0.38 m–q | 285.11 k–q | ||
| Ś16 | 11.39 ± 0.43 n–p | 2.78 ± 0.25 n–q | 2.66 ± 0.23 n–s | 394.9 ± 11.4 jk | 5.88 ± 3.29 t–v | 417.66 k–o | 502.25 ± 1.54 b | 36.21 ± 1.11 b | 167.71 ± 2.54 a | 16.37 ± 1.01 a | 20.85 ± 0.57 a | 61.38 ± 0.88 a | 804.77 a | ||
| Ś17 | 15.51 ±0.26 j–l | 2.32 ± 0.52 o–s | 2.02 ± 0.15 q–s | 234.2 ± 14.3 m–p | 3.12 ± 0.54 vw | 257.25 o–t | 572.68 ± 2.15 a | 54.55 ± 3.22 a | 50.93 ± 1.02 ef | 6.66 ± 0.22 de | 3.15 ± 0.43 o–q | 40.23 ± 0.46 d | 728.20 b | ||
| Ś18 | 23.61 ± 0.54 e–g | 19.42 ± 0.51 a | nd u | 269.2 ± 10.5 l–n | 10.60 ± 1.11 q–t | 322.85 m–r | 462.49 ± 2.43 bc | 17.41 ± 1.05 fg | 64.27 ± 1.32 cd | 7.68 ± 0.54. bc | 8.25 ± 0.12 gh | 44.96 ± 0.29 c | 605.07 c | ||
| Ś19 | 16.23 ± 0.21 j–l | 3.43 ± 0.17 k–o | 3.05 ± 0.31 m–p | 513.1 ± 21.0 g–i | 20.61 ± 2.73 h–k | 556.50 g–l | 435.02 ± 0.33 c | 32.84 ± 0.76 c | 46.41± 1.72 fg | 3.74 ± 0.28 k–n | 14.54 ± 0.32 c | 36.36 ± 1.43 d | 568.91 cd | ||
| Ś20 | 42.65 ± 0.32 b | 8.28 ± 0.51 e | 10.27 ± 0.56 bc | 229.0 ± 10.0 m–p | 15.31 ± 0.18 l–r | 305.56 n–r | 48.17 ± 2.65 t | nd t | 23.49 ± 1.99 m–r | nd v | 7.40 ± 0.37 h–k | 4.25 ± 0.12 st | 83.31 x | ||
| Ś21 | 10.89 ± 0.11 o–q | nd u | nd u | 885.1 ± 46.1 b | 13.96 ± 2.01 m–s | 910.04 bc | 157.66 ± 1.44 o–q | 8.24 ± 0.22 m–q | 8.81 ± 1.73 u–w | 2.09 ± 0.34 s–u | 3.29 ± 0.29 op | 7.46 ± 0.54 p–s | 187.54 s–w | ||
| Ś22 | 14.91 ± 0.42 k–n | nd u | 1.94 ± 0.13 rs | 83.5 ± 8.3 s | 21.05 ± 2.82 h–j | 121.45 st | 423.93 ± 0.65 cd | 14.34 ± 2.11 g–j | 28.99 ± 1.87 j–n | 5.23 ± 0.23 f–h | 11.30 ± 1.11 d | 20.74 ± 0.45 fg | 504.53 de | ||
| Ś23 | 27.28 ± 0.12 de | nd u | nd u | 267.4 ± 12.0 l–o | 12.93 ± 1.44 n–s | 307.62 n–r | 193.47 ± 0.99 l–p | 5.77 ± 0.32 qr | 24.59 ± 0.81 l–p | nd v | 3.23 ± 0.43 op | 7.78 ± 0.48 o–s | 234.84 p–t | ||
| Ś24 | 10.26 ± 0.74 o–r | 3.01 ± 0.43 l–p | 3.32 ± 0.25 l–n | 641.9 ± 10.1 de | 4.46 ± 0.54 u–w | 662.98 d–i | 219.18 ± 0.65 j–n | 13.81 ± 0.55 h–j | 15.58 ± 0.71 q–u | 3.13 ± 0.29 m–r | 2.68 ± 0.32 o–s | 11.68 ± 0.99 k–p | 266.07 m–r | ||
| Ś25 | 15.94 ± 0.12 j–l | nd u | 4.82 ± 0.11 jk | 458.8 ± 10.4 h–j | 12.42 ± 1.11 o–s | 491.99 i–m | 105.22 ± 0.56 q–t | 5.30 ± 0.99 qr | 59.55 ± 0.72 de | nd v | 9.77 ± 0.88 ef | 19.91± 0.94 f–h | 199.75 r–v | ||
| Ś26 | 10.76 ± 0.54 o–q | nd u | 1.81 ± 0.43 st | 185.2 ± 11.1 n–r | 14.29 ± 0.88 m–s | 212.16 q–t | 192.49 ± 1.32 l–p | 5.27 ± 0.54 qr | 43.59 ± 0.99 f–h | 2.29 ± 0.54 r–u | 6.83 ± 0.27 h–l | 8.18 ± 0.48 n–s | 258.64 o–s | ||
| Ś27 | 29.74 ± 0.62 cd | 6.60 ± 0.23 fg | 8.22 ± 0.56 d | 564.3 ± 20.1 e–g | 13.13 ± 3.11 n–s | 622.06 e–j | 231.54 ± 2.43 j–m | nd t | nd w | 3.56 ± 0.29 l–o | 1.63 ± 0.29 r–t | 17.14 ± 0.77 g–j | 253.86 o–t | ||
| Ś28 | 7.57 ± 1.12 q–t | 4.06 ± 0.43 j–m | 3.02 ± 0.54 m–q | 755.0 ± 20.5 c | 21.19 ± 1.11 h–j | 790.90 c–e | 107.59 ± 1.99 q–s | 5.80 ± 0.23 p–r | 6.50 ± 0.72 vw | 0.70 ± 0.54 v | 3.08 ± 0.11 o–r | 6.34 ± 0.56 q–t | 130.02 u–x | ||
| Ś29 | 13.37 ± 0.48 l–o | 5.40 ± 0.26 hi | 6.28 ± 0.12 g–i | 433.1 ± 16.4 i–k | 13.26 ± 1.43 n–s | 471.41 j–n | 426.42 ± 1.56 c | 13.73 ± 0.45 h–j | 32.08 ± 1.21 i–m | 6.89 ± 0.29 cd | 8.20 ± 0.99 g–i | 15.97± 0.77 h–k | 503.29 de | ||
| Ś30 | 13.07 ± 0.12 l–o | 1.13 ± 0.11 tu | 2.27 ± 0.13 o–s | 209.3 ± 10.3 m–q | 3.59 ± 1.03 vw | 229.36 p–t | 247.05 ± 1.99 i–l | 17.32 ± 0.76 fg | 12.99 ± 0.63 s–v | 3.91 ± 0.10 j–m | 6.28 ± 1.11 j–m | 10.41 ± 0.68 m–q | 297.95 k–p | ||
| Ś31 | 1.16 ± 0.15 u | nd u | 3.76 ± 0.43 lm | 235.0 ± 10.7 m–p | 43.40 ± 2.67 c | 283.37 o–t | 366.51 ± 2.54 d–f | 10.26 ± 0.34 k–n | 61.35 ± 0.71 cd | 8.57 ± 0.10 b | 7.45 ± 0.73 h–k | 27.16 ± 0.99 e | 481.29 ef | ||
| Ś32 | 10.09 ± 0.35 o–r | nd u | nd u | 175.0 ± 10.3 o–s | 16.77 ± 2.68 j–o | 201.88 q–t | 311.67 ± 2.14 f–h | 14.67 ± 0.99 g–i | 62.03 ± 0.29 cd | 2.39 ± 0.47 r–u | 5.96 ± 0.34 k–m | 20.52 ± 0.68 fg | 417.24 f–h | ||
| Ś33 | 22.93 ± 0.37 fg | 7.06 0.23 f | 7.21 ± 0.36 d–g | 503.6 ± 11.1 g–i | 25.07 ± 1.88 f–h | 565.91 g–l | 347.88 ± 4.21 fg | 12.21 ± 0.23 i–l | 68.69 ± 1.43 c | 3.30 ± 0.12 l–q | 16.13 ± 0.53 b | 11.41 ± 0.59 l–p | 459.62 ef | ||
| Ś34 | 8.35 ± 0.39 p–s | 2.94 ± 0.43 m–q | 3.09 ± 0.12 m–p | 568.6 ± 10.4 e–g | 28.32 ± 2.01 ef | 611.35 f–j | 212.83 ± 2.43 k–o | 5.68 ± 0.37 qr | 33.86 ± 1.72 i–k | 5.66 ± 0.15 fg | 4.88 ± 0.16 mn | 21.07 ± 0.47 fg | 283.97 k–q | ||
| Ś35 | 6.79 ± 0.66 r–t | nd u | 0.89 ± 0.32 tu | 105.6 ± 11.0 rs | 4.00 ± 0.36 vw | 117.35 t | 137.42 ± 1.99 p–r | 4.48 ± 0.29 rs | 23.36 ± 1.54 m–r | 2.49 ± 0.21 q–u | 2.84 ± 0.43 o–r | 11.80 ± 1.02 k–o | 182.39 t–w | ||
| Ś36 | 18.86 ± 0.10 ij | 6.97 ± 0.43 f | 6.51 ±0.23 f–h | 221.1 ± 15.3 m–p | 10.34 ± 1.01 r–t | 263.87 o–t | 162.35 ± 1.45 n–q | 6.31 ± 0.32 o–r | 11.82 ± 1.24 s–v | 1.78 ± 0.45 tu | 4.85 ± 0.54 mn | 17.10 ± 2.01 g–j | 204.20 r–u | ||
| Ś37 | 22.75 ± 1.04 f–h | 6.34 ± 0.46 f–h | 5.01 ± 0.43 jk | 211.0 ± 28 m–q | 27.74 ± 2.01 e–g | 272.85 o–t | 150.02 ± 2.11 p–r | 8.02 ± 0.58 n–q | 82.54 ± 1.14 b | nd v | 13.73 ± 1.11 c | 49.54 ± 0.12 b | 303.85 k–p | ||
| Ś38 | 31.11 ± 0.19 c | 9.36 ± 0.54 de | 11.18 ± 0.99 ab | 274.2 ± 35 l–n | 17.05 ± 2.07 j–o | 342.98 m–q | 362.40 ± 1.54 ef | 26.86 ± 0.99 d | 36.36 ± 1.63 h–j | 3.63 ± 0.52 l–o | 5.86 ± 1.88 lm | 20.49 ± 0.43 fg | 455.61 e–g | ||
| Ś39 | 20.90 ± 1.00 g–i | 5.22 ± 0.65 h–j | 7.51 ± 0.43 d–f | 427.6 ± 19 i–k | 15.84 ± 1.54 k–p | 477.12 j–n | 264.32 ± 1.78 h–k | 12.74 ± 0.79 i–k | 56.10 ± 1.71 de | 2.80 ± 0.32 o–s | 6.77 ± 0.75 h–l | 14.91 ± 0.19 j–l | 357.63 h–k | ||
| Ś40 | 10.95 ± 0.28 o–q | 1.21 ± 0.11 st | 2.17 ± 0.28 p–s | 154.2 ± 10.2 p–s | 30.93 ± 0.32 e | 199.51 q–t | 417.24 ± 2.66 c–e | 15.40 ± 0.39 f–i | 40.20 ± 1.92 g–i | 4.90 ± 0.54 g–i | 1.70 ± 0.12 q–t | 39.17 ± 1.01 d | 518.61 de | ||
| Ś41 | 11.27 ± 0.32 n–q | 2.17 ± 0.12 p–t | 3.45 ± 0.47 l–n | 127.9 ± 10.3 q–s | 18.69 ± 1.15 i–m | 163.49 r–t | 340.10 ± 3.05 fg | 18.07 ± 1.07 f | 30.57 ± 1.44 j–n | 3.86 ± 0.73 j–m | nd u | 20.81± 1.44 fg | 413.41 f–i | ||
| Ś42 | 6.15 ± 0.11 st | 1.16 ± 0.09 s–u | 2.01 ± 0.18 q–s | 215.6 ± 11.0 m–q | 9.24 ± 1.05 s–u | 134.26 p–t | 63.88 ± 2.43 st | 1.56 ± 0.69 st | 9.67 ± 1.32 t–v | 0.64 ± 0.11 v | 3.21 ± 0.43 op | 4.25 ± 0.43 st | 83.20 x | ||
| Ś43 | 17.88 ± 0.38 i–k | 5.76 ± 0.32 gh | 7.00 ± 0.67 e–g | 662.2 ± 28.9 cd | 16.76 ± 1.33 j–o | 709.69 d–g | 335.25 ± 3.12 fg | 17.48 ± 1.37 fg | 19.42 ± 0.23 o–s | 4.57 ± 0.12 h–k | 7.77 ± 1.65 g–j | 8.85 ± 0.99 n–r | 336.43 j–n | ||
| Sample | Flavonols † | Anthocyanins † | |||||||||||||
| Q-pentoside-hexoside | Q-3-galactoside | Q-3-glucoside | Q-3-rutinoside | Q-arabinoside | Q-rhamnoside | Q-penthoside rhamnoside | Other | Total | C-3-O-galactoside | C-3-O-glucoside | C-3-O-rutinoside | p-3-O-glucoside | Other | Total | |
| Ś1 | 2.49 ± 0.23 f–h | 3.33 ± 0.11 o–t | 10.73 ± 0.66 g | 17.58 ± 2.10 e–h | 5.88 ± 0.38 f–h | 1.52 ± 0.43 jk | 4.03 ± 0.21 de | 7.76 ± 1.01 d | 53.31 f–i | 1.08 ± 0.23 o–u | 3.48 ± 0.12 g | 11.34 ± 0.32 e–h | 1.91 ± 0.32 f–h | 5.13 ± 0.54 cd | 22.94 j–m |
| Ś2 | 1.09 ± 0.19 n–p | 11.92 ± 1.11 hi | 3.75 ± 0.43 m–p | 8.59 ± 0.55 q–s | 1.99 ± 0.63 p–u | 0.84 ± 0.10 l–n | 0.21 ± 0.02 p–s | nd u | 28.38 n–r | 3.87 ± 0.32 hi | 1.22 ± 0.11 m–p | 5.54 ± 0.21 q–s | 0.65 ± 0.11 p–u | 0.69 ± 0.09 uv | 11.97 q–t |
| Ś3 | 1.22 ± 0.21 m–o | 5.77 ± 0.21 l–o | 4.78 ± 1.32 l–n | 11.39 ± 1.77 k–q | 3.56 ± 0.62 l–o | nd o | 1.21 ± 0.14 j–n | 0.87 ± 0.10 q–u | 28.80 n–r | 1.87 ± 0.44 l–o | 1.55 ± 0-.13 l–n | 7.35 ± 0.16 k–q | 1.16 ± 0.32 l–o | 1.07 ± 0.23 tu | 13.00 q–s |
| Ś4 | 1.09 ± 0.32 n–p | 3.72 ± 0.39 n–s | 1.26 ± 0.37 s–v | 10.60 ± 0.77 m–r | 1.17 ± 0.38 s–v | 0.64 ± 0.01 n | 0.52 ± 0.19 o–s | 2.41 ± 0.21 m–p | 21.42 q–t | 1.21 ± 0.12 n–t | 0.41 ± 0.02 s–u | 6.84 ± 0.43 m–r | 0.38 ± 0.14 s–v | 1.52 ± 0.32 p–t | 10.35 r–v |
| Ś5 | 1.01 ± 0.11 n–p | 4.62 ± 0.22 m–q | 1.47 ± 0.34 r–v | 12.48 ± 0.99 k–p | 5.65 ± 0.29 f–h | nd o | nd s | 1.51 ± 0.09 o–r | 26.74 o–s | 1.50 ± 0.11 m–q | 0.48 ± 0.11 r–u | 8.05 ± 0.37 k–p | 1.83 ± 0.32 f–h | 0.82 ± 0.23 t–v | 12.68 q–t |
| Ś6 | 2.50 ± 0.17 f–h | 7.01 ± 0.36 k–m | 2.90 ± 0.66 o–s | 20.59 ± 0.96 e | 5.99 ± 0.76 fg | 1.27 ± 0.22 j–l | 2.05 ± 0.11 g | 2.67 ± 0.47 l–n | 44.97 i–l | 2.28 ± 0.11 k–m | 0.94 ± 0.11 o–s | 13.29 ± 0.54 e | 1.94 ± 0.11 fg | 2.76 ± 0.12 j–m | 21.20 k–n |
| Ś7 | 1.81 ± 0.11 j–l | 9.13 ± 0.64 i–k | 3.16 ± 0.28 n–q | 7.71 ± 1.21 q–t | 2.67 ± 0.99 n–r | 1.70 ± 0.28 ij | 0.69 ± 0.32 n–r | 1.88 ± 0.54 n–q | 28.75 n–r | 2.96 ± 0.43 i–k | 1.03 ± 0.23 n–q | 4.97 ± 0.76 r–t | 0.87 ± 0.21 n–r | 1.97 ±0.43 n–q | 11.80 q–t |
| Ś8 | 1.11 ± 0.09 n–p | 2.64 ± 0.72 p–t | 1.19 ± 0.46 t–v | 8.99 ± 0.77 p–s | 2.12 ± 0.10 p–u | 0.73 ± 0.19 mn | 1.00 ± 0.11 j–o | nd u | 17.78 s–u | 0.86 ± 0.15 p–u | 0.39 ± 0.02 tu | 5.80 ± 0.49 p–s | 0.69 ± 0.13 p–u | 0.92 ± 0.18 t–v | 8.66 s–w |
| Ś9 | 2.54 ± 0.11 fg | 1.05 ± 0.11 st | 0.71 ± 0.02 v | 13.87 ± 0.48 i–m | 1.59 ± 0.24 r–u | nd o | 3.41 ± 0.41 ef | 3.37 ± 0.36 j–m | 26.53 o–s | 0.34 ± 0.43 tu | 0.23 ± 0.12 u | 8.95 ± 0.54 i–m | 0.51 ± 0.04 r–u | 3.03 ± 0.54 i–l | 13.06 q–s |
| Ś10 | 0.91 ± 0.12 o–q | 12.10 ± 0.43 h | 3.91 ± 0.34 m–p | 19.88 ± 0.54 ef | 2.46 ± 0.19 o–s | 3.62 ± 0.46 d | 2.93 ± 0.38 f | 1.42 ± 0.43 o–s | 47.25 h–k | 3.93 ± 0.16 h | 1.27 ± 0.02 m–p | 12.83 ± 0.28 ef | 0.80 ± 0.06 o–s | 2.88 ± 0.23 j–l | 21.71 k–n |
| Ś11 | 1.24 ± 0.11 m–o | 11.04 ± 0.77 h–j | 4.41 ± 0.54 m–o | 12.94 ± 0.48 j–o | 3.80 ± 0.55 j–n | 1.13 ± 0.21 k–m | 0.85 ± 0.22 k–o | 0.37 ± 0.19 tu | 35.78 l–o | 3.58 ± 0.32 h–j | 1.43 ± 0.32 m–o | 8.35 ± 0.87 j–o | 1.23 ± 0.16 j–n | 1.16 ± 0.33 r–u | 15.76 o–q |
| Ś12 | 0.77 ± 0.21 p–r | 20.09 ± 1.32 e–f | 8.83 ± 0.84 ij | 8.50 ± 0.77 q–s | 3.09 ± 0.75 m–q | 3.24 ± 0.32 de | 0.60 ± 0.10 n–s | 1.14 ± 0.63 q–t | 46.28 h–l | 6.52 ± 0.13 ef | 2.87 ± 0.12 ij | 5.49 ± 0.76 q–s | 1.00 ± 0.11 m–q | 1.87 ± 0.19 n–r | 17.75 n–p |
| Ś13 | nd t | 1.57 ± 0.33 r–t | 1.42 ± 0.39 r–v | 13.63 ± 0.59 i–n | 4.61 ± 0.39 h–l | 1.46 ± 0.15 jk | 4.12 ± 0.32 d | 3.28 ± 0.49 j–m | 30.09 n–r | 0.51 ± 0.33 r–u | 0.46 ± 0.02 r–u | 8.79 i± 0.99 -n | 1.50 ± 0.18 h–l | 2.88 ± 0.12 j–l | 14.14 p–r |
| Ś14 | nd t | 23.72 ± 0.54 d | 10.56 ± 0.44 gh | 14.86 ± 0.74 g–k | 3.19 ± 0.76 m–p | 2.15 ± 0.29 hi | 0.73 ± 0.11 m–r | 3.07 ± 0.29 k–m | 58.28 e–g | 7.70 ± 0.12 d | 3.43 ± 0.23 gh | 9.59 ± 0.27 g–k | 1.03 ± 0.43 m–p | 1.93 ± 0.32 n–q | 23.68 j–l |
| Ś15 | 3.55 ± 0.56 d | 1.12 ± 0.10 st | 3.05 ± 0.93 o–r | 7.58 ± 0.82 r–t | 2.00 ± 0.39 p–u | 0.86 ± 0.11 l–n | 1.98 ± 0.29 gh | 7.72 ± 0.73 d | 27.85 o–s | 0.36 ± 0.10 s–u | 0.99 ± 0.11 o–r | 4.89 0.56 r–t | 0.65 ± 0.4 p–u | 4.58 ± 0.43 d–f | 11.47 q–u |
| Ś16 | 2.06 ± 0.29 h–k | 6.53 ± 0.43 k–n | 4.17 0.37 m–o | 32.46 ± 0.73 cd | 9.18 ± 3.66 cd | 2.91 ± 0.29 ef | 4.83 ± 0.43 c | 3.62 ± 0.65 j–l | 65.75 c–e | 2.12 ± 0.12 k–n | 1.35 ± 0.13 m–o | 20.95 ± 1.11 cd | 2.98 ± 0.23 cd | 4.35 ± 0.65 e–g | 31.75 d–f |
| Ś17 | 1.24 ± 0.33 m–o | 8.42 ± 0.88 j–l | 1.97 ± 0.43 q–v | 18.29 ± 0.88 e–g | 1.79 ± 0.65 q–u | 1.38 ± 0.16 jk | 1.43 ± 0.11 g–k | 12.88 ± 1.32 b | 47.41 h–k | 2.73 ± 0.32 j–l | 0.64 ± 0.02 q–u | 11.81 ± 0.88 e–g | 0.58 ± 0.07 q–u | 5.50 ± 0.33 c | 21.26 k–n |
| Ś18 | 1.41 ± 0.11 l–n | 18.55 ± 1.72 fg | 9.01 ± 1.21 hi | 41.61 ± 0.34 a | 11.77 ± 1.11 b | 2.92 ± 0.32 ef | 4.00 ± 0.88 de | 7.53 ± 0.92 d | 96.80 b | 6.02 ± 0.25 fg | 2.92 ± 0.12 hi | 26.86 ± 0.77 a | 3.82 ± 0.23 b | 5.15 ± 0.54 cd | 44.77 b |
| Ś19 | 3.63 ± 0.24 cd | 4.37 ± 0.32 m–r | 1.39 ± 0.46 s–v | 16.72 ± 1.12 f–i | 1.56 ± 0.21 r–u | nd o | 0.76 ± 0.39 l–r | 4.28 ± 0.19 g–j | 32.70 m–p | 1.42 ± 0.32 m–r | 0.45 ± 0.07 s–u | 10.79 ± 0.76 f–i | 0.51 ± 0.11 r–u | 2.81 ± 0.26 j–m | 15.98 o–q |
| Ś20 | 1.38 ± 0.43 l–n | 3.66 ± 0.37 n–t | 2.05 ± 0.29 q–v | 10.08 ± 1.08 n–r | 5.26 ± 0.31 g–i | nd o | 1.35 ± 0.52 i–m | 14.59 ± 0.32 a | 38.37 k–n | 1.19 ± 0.15 n–t | 0.66 ± 0.06 q–u | 6.51 ± 1.00 n–r | 1.71 ± 0.21 g–i | 5.62 ± 0.41 c | 15.69 o–q |
| Ś21 | 1.24 ± 0.48 m–o | 0.75 ± 0.01 t | 0.65 ± 0.08 v | 7.60 ± 0.55 r–t | 1.00 ± 0.11 t–v | nd o | 1.36 ± 0.19 h–l | 1.38 ± 0.21 p–t | 13.98 tu | 0.24 ± 0.02 u | 0.21 ± 0.03 u | 4.91 ± 0.10 r–t | 0.32 ± 0.03 uv | 1.29 ± 0.43 q–u | 6.98 u–x |
| Ś22 | 4.45 ± 0.29 b | 1.88 ± 0.21 q–t | 7.26 ± 0.58 jk | 18.78 ± 0.99 ef | 24.12 ± 0.73 a | nd o | 9.99 ± 0.42 a | 5.31 ± 1.03 fg | 71.79 cd | 0.61 ± 0.4 q–u | 2.36 ± 0.29 jk | 12.12 ± 0.43 ef | 7.83 ± 0.54 a | 6.41 ± 0.73 b | 29.32 f–h |
| Ś23 | 2.86 ± 0.54 ef | 3.42 ± 0.11 o–t | 2.51 ± 0.58 p–t | 7.68 ± 0.29 r–t | 3.60 ± 0.66 k–o | 1.68 ± 0.11 j | 1.54 ± 0.33 g–j | 1.90 ± 0.11 n–q | 25.19 p–s | 1.11 ± 0.09 o–u | 0.81 ± 0.08 p–t | 4.96 ± 0.26 r–t | 1.17 ± 0.27 k–o | 2.59 ± 0.37 k–n | 10.64 r–u |
| Ś24 | nd t | 4.90 ± 0.54 m–p | 1.85 ± 0.29 q–v | 4.66 ± 0.48 tu | 1.11 ± 0.12 t–v | 0.40 ± 0.12 no | 0.19 ± 0.10 q–s | 0.46 ± 0.19 s–u | 13.58 tu | 1.59 ± 0.12 m–p | 0.60 ± 0.01 q–u | 3.01 ± 0.77 tu | 0.36 ± 0.02 t–v | 0.34 ± 0.11 v | 5.90 v–x |
| Ś25 | 0.42 ± 0.02 r–t | 12.10 ± 0.99 h | 17.35 ± 0.21 c | 8.40 ± 0.29 q–s | 7.73 ± 0.21 e | nd o | 0.80 ± 0.22 l–q | 6.37 ± 0.88 e | 53.17 f–i | 3.93 ± 0.32 h | 5.63 ± 0.32 c | 5.42 ± 0.29 q–s | 2.51 ± 0.18 e | 2.47 ± 0.22 l–o | 19.95 l–o |
| Ś26 | 2.30 ± 0.20 g–i | 4.64 ± 0.48 m–q | 11.40 ± 0.42 fg | 11.04 ± 0.54 l–r | 4.11 ± 0.32 i–m | 8.09 ± 0.11 a | 10.06 ± 1.06 a | 14.87 ± 1.32 a | 66.50 c–e | 1.50 ± 0.12 m–q | 3.70 ± 0.12 fg | 7.12 ± 0.88 l–r | 1.33 ± 0.56 i–m | 11.46 ± 0.67 a | 25.12 h–k |
| Ś27 | 0.24 ± 0.11 st | 1.13 ± 0.29 st | 0.42 ± 0.01 v | 6.02 ± 0.55 s–u | 0.90 ± 0.13 uv | nd o | nd s | 0.51 ± 0.01 r–u | 9.21 u | 0.37 ± 0.01 s–u | 0.14 ± 0.01 u | 3.89 ± 0.39 s–u | 0.29 ± 0.02 uv | 0.24 ± 0.04 v | 4.92 wx |
| Ś28 | nd t | 3.51 ± 0.73 o–t | 0.82 ± 0.01 uv | 2.40 ± 0.64 u | 0.15 ± 0.29 v | 0.52 ± 0.05 n | 0.15 ± 0.02 rs | 0.07 ± 0.01 u | 7.60 u | 1.14 ± 0.12 o–u | 0.26 ± 0.02 u | 1.55 ± 0.44 u | 0.05 ± 0.00 v | 0.24 ± 0.01 v | 3.24 x |
| Ś29 | 2.00 ± 0.10 i–k | 16.40 ± 0.71 g | 12.92 ± 0.39 ef | 20.88 ± 0.49 e | 11.97 ± 0.92 b | 2.17 ± 0.23 h | 1.15 ± 0.05 j–n | 3.61 ± 0.12 j–l | 71.09 cd | 5.32 ± 0.43 g | 4.19 ± 0.03 ef | 13.48 ± 0.88 e | 3.88 ± 0.34 b | 2.90 ± 0.55 j–l | 29.77 e–g |
| Ś30 | nd t | 9.98 ± 0.48 h–j | 8.54 ± 0.32 ij | 38.99 ± 2.73 ab | 2.30 ± 0.13 o–t | nd o | 1.85 ± 0.18 g–i | 4.72 ± 0.32 f–i | 66.37 c–e | 3.24 ± 0.26 h–j | 2.77 ± 0.12 ij | 25.16 ± 0.66 ab | 0.75 ± 0.06 o–t | 2.13 ± 0.18 m–p | 34.05 c–e |
| Ś31 | 1.30 ± 0.11 m–o | 28.02 ± 0.49 c | 11.02 ± 0.43 g | 30.80 ± 0.38 d | 7.97 ± 0.21 de | 2.60 ± 0.32 f–h | 1.60 ± 0.21 g–j | 3.91 ± 0.12 h–k | 87.21 b | 9.09 ± 0.33 c | 3.58 ± 0.65 g | 19.88 ± 0.32 d | 2.59 ± 0.03 de | 3.05 ± 0.45 i–l | 38.19 c |
| Ś32 | 4.12 ± 0.32 b | 53.67 ± 0.39 a | 20.34 ± 0.25 b | 35.35 ± 1.88 bc | 9.72 ± 0.45 c | nd o | 1.56 ± 0.19 g–j | 4.01 ± 0.54 h–k | 128.77 a | 17.42 ± 0.54 a | 6.60 ± 0.66 b | 22.82 ± 0.99 bc | 3.16 ± 0.43 c | 3.15 ± 0.45 i–l | 53.14 a |
| Ś33 | 3.01 ± 0.10 e | 18.67 ± 0.29 fg | 6.16 ± 0.12 kl | 16.67 ± 0.39 f–i | 5.52 ± 0.44 f–h | 2.71 ± 0.43 fg | 1.37 ± 0.29 h–l | 2.42 ± 0.65 m–o | 56.53 e–h | 6.06 ± 0.99 fg | 2.00 ± 0.65 kl | 10.76 ± 0.63 f–i | 1.79 ± 0.25 f–h | 3.08 ± 0.29 i–l | 23.70 j–l |
| Ś34 | 0.36 ± 0.02 r–t | 22.71 ± 0.83 de | 14.22 ± 0.32 de | 14.55 ± 1.32 h–l | 4.67 ± 0.29 h–l | 3.26 ± 0.29 de | 0.82 ± 0.11 k–p | 1.17 ± 0.29 q–t | 61.75 d–f | 7.37 ± 0.65 de | 4.61 ± 0.55 de | 9.39 ± 0.65 h–l | 1.52 ± 0.43 g–l | 1.82 ± 0.19 o–s | 24.71 i–k |
| Ś35 | 2.93 ± 0.38 ef | 4.11 ± 0.77 m–r | 5.02 ± 0.86 lm | 9.88 ± 0.74 o–r | 1.97 ± 0.12 p–u | 2.33 ± 0.33 gh | 3.99 ± 0.32 de | 0.61 ± 0.05 r–u | 30.83 m–q | 1.33 ± 0.36 n–r | 1.63 ± 0.13 lm | 6.37 ± 0.82 o–r | 0.64 ± 0.52 p–u | 3.20 ± 0.39 i–k | 13.18 q–s |
| Ś36 | 1.65 ± 0.32 k–m | 20.58 ± 1.43 ef | 12.73 ± 0.19 ef | 16.59 f–j | 4.90 ± 0.74 g–k | 6.20 ± 0.62 b | 2.87 ± 0.17 f | 1.46 ± 0.32 o–s | 66.98 c–e | 6.68 ± 0.43 ef | 4.13 ± 0.43 ef | 10.71± 0.69 f–j | 1.59 ± 0.29 g–k | 3.96 ± 0.38 f–h | 27.06 g–j |
| Ś37 | 0.88 ± 0.06 o–q | 16.55 ± 1.76 g | 23.17 ± 0.32 a | 12.56 k–p | 11.71 ± 1.32 b | nd o | 1.14 ± 0.21 j–o | 9.52 ± 0.99 c | 75.52 c | 5.37 ± 0.67 g | 7.52 ± 0.77 a | 8.11 ± 0.73 k–p | 3.80 ± 0.54 b | 3.74 ± 0.19 g–i | 28.54 f–i |
| Ś38 | 2.20 ± 0.19 g–j | 1.76 ± 0.53 q–t | 0.70 ± 0.11 v | 29.52 d | 5.07 ± 0.37 g–j | 0.44 ± 0.05 no | 4.58 ± 0.11 cd | 4.89 ± 0.43 f–h | 49.16 g–j | 0.57 ± 0.09 q–u | 0.23 ± 0.06 u | 19.05 ± 0.88 d | 1.65 ± 0.29 g–j | 3.93 ± 0.44 f–h | 25.42 g–k |
| Ś39 | 0.56 ± 0.03 q–s | 39.72 ± 0.38 b | 15.78 ± 0.98 cd | 19.48 ef | 6.75 ± 0.52 ef | 5.26 ± 0.29 c | 1.02 ± 0.32 j–o | 3.81 ± 0.76 i–k | 92.38 b | 12.89 ± 0.87 b | 5.12 ± 0.56 cd | 12.57 ± 0.62 ef | 2.19 ± 0.29 ef | 3.46 ± 0.10 h–j | 36.23 cd |
| Ś40 | 3.19 ± 0.11 de | 3.96 ± 0.43 n–s | 1.95 ± 0.28 q–v | 14.52 h–l | 1.79 ± 0.10 q–u | 1.15 ± 0.18 k–m | 1.37 ± 0.29 h–l | 5.55 ± 0.39 ef | 33.49 m–p | 1.29 ± 0.88 n–s | 0.63 ± 0.05 q–u | 9.37 ± 0.99 h–l | 0.58 ± 0.05 q–u | 3.66 ± 0.29 g–i | 15.53 o–q |
| Ś41 | 4.07 ± 0.32 bc | 3.59 ± 0.29 o–t | 4.17 ± 0.79 m–o | 35.92 bc | 12.42 ± 1.86 b | 1.52 ± 0.55 jk | 7.48 ± 0.72 b | 4.23 ± 0.64 h–j | 73.39 c | 1.17 ± 0.43 o–u | 1.35 ± 0.31 m–o | 23.18 ± 1.34 bc | 4.03 ± 0.29 b | 5.61 ± 0.55 c | 35.35 cd |
| Ś42 | 0.37 ± 0.05 r–t | 3.75 ± 0.54 n–s | 3.73 ± 0.54 m–p | 5.74 s–u | 3.14 ± 0.51 m–p | 1.17 ± 0.43 k–m | 0.73 ± 0.43 m–r | 1.14 ± 0.49 q–t | 19.79 r–t | 1.22 ± 0.76 n–t | 1.21 ± 0.23 m–p | 3.71 ± 0.34 s–u | 1.02 ± 0.38 m–p | 1.11 ± 0.16 s–u | 8.27 t–w |
| Ś43 | 6.91 ± 0.25 a | 4.21 ± 0.77 m–r | 2.47 ± 0.49 p–u | 17.62 e–h | 1.78 ± 1.63 r–u | 1.42 j± 0.28 k | 2.97 ± 0.49 f | 3.30 ± 0.29 j–m | 40.67 j–m | 1.37 ± 0.56 m–r | 0.80 ± 0.03 p–t | 11.37 ± 1.93 e–h | 0.58 ± 0.01 q–u | 4.74 ± 0.39 de | 18.86 m–o |
| Sample | Antioxidant Activity | Antidiabetic Acitvity † | Antiobesity Activity † | 15-LOX † | Anti-Aging Activity † | |||
|---|---|---|---|---|---|---|---|---|
| ABTSo+ | FRAP | α-Amylase | α-Glucosidase | Pancreatic lipase | AChE | BuChE | ||
| Ś1 | 5.38 ± 0.33 c–h | 8.32 ± 0.29 g–l | nd r | 2.92 ± 0.11 q | 7.94 ± 0.02 a | 9.27 ± 1.01 m–r | 31.32 ± 0.72 j–m | 48.60 ± 1.32 g–k |
| Ś2 | 6.97 ± 0.68 ab | 11.71 ± 0.40 ab | nd r | nd r | 2.98 ± 0.09 j–o | 16.71 ± 0.43 e–f | 37.68 ±1.03 f–k | 56.33 ± 1.65 c–h |
| Ś3 | 4.53 ± 0.23 g–m | 8.01 ± 0.60 h–m | nd r | 2.06 ± 0.32 q–r | 2.56 ± 0.11 n–t | 11.58 ± 0.23 i–m | nd r | 41.92 ± 2.33 j–n |
| Ś4 | 3.71 ± 0.51 m–o | 6.12 ± 0.18 op | 32.57 ± 2.15 g–k | 1.84 ± 0.11 q–r | 3.11 ± 0.06 i–o | 9.54 ± 0.28 m–r | 41.73 ± 1.64 e–h | 52.73 ± 2.54 e–i |
| Ś5 | 3.63 ± 0.04 m–o | 6.06 ± 0.26 op | 31.70 ± 1.76 h–l | 0.45 ± 0.31 r | 4.23 ± 0.04 c–f | 8.63 ± 0.54 n–s | 42.71 ± 1.33 d–g | 56.16 ± 1.42 c–h |
| Ś6 | 4.71 ± 0.59 e–l | 8.45 ± 0.16 g–l | 25.97 ± 2.41 m–o | nd r | 5.60 ± 0.23 b | 5.90 ± 0.99 t–v | 25.19 ± 1.43 m–o | 25.43 ± 1.76 p |
| Ś7 | 4.73 ± 0.61 e–l | 8.75 ± 0.69 f–k | 30.98 ± 2.51 i–m | nd r | 3.70 ± 0.41 f–i | 7.01 ± 0.29 r–u | 38.85 ± 2.17 f–j | 53.89 ± 2.54 d–i |
| Ś8 | 5.58 ± 0.09 c–f | 9.47 ± 0.76 d–i | nd r | nd r | 4.69 ± 0.23 cd | 20.67 ± 0.76 c | 36.70 ± 2.43 g–k | 53.04 ± 1.65 e–i |
| Ś9 | 4.83 ± 0.30 e–l | 8.90 ± 0.89 e–j | nd r | nd r | 3.54 ± 0.24 g–k | 6.59 ± 0.99 s–v | 18.83 ± 1.43 op | 32.22 ± 1.27 n–p |
| Ś10 | 3.16 ± 0.24 op | 5.81 ± 0.30 op | 30.70 ± 1.11 i–m | 0.68 ± 0.40 r | 3.58 ± 0.54 f–k | 7.26 ± 0.69 q–u | 27.87 ± 2.52 l–n | 36.26 ± 2.54 no |
| Ś11 | 7.71 ± 0.47 a | 13.28 ± 0.30 a | nd r | 6.93 ± 0.65 m–o | 3.20 ± 0.32 h–n | 4.19 ± 0.67 v | 40.97 ± 2.43 e–h | 60.54 ± 1.52 b–f |
| Ś12 | 6.25 ± 0.64 bc | 10.51 ± 0.22 b–e | nd r | 0.19 ± 0.23 r | 1.94 ± 0.26 t–w | 8.28 ± 0.58 o–t | 31.98 ± 1.11 i–m | 37.15 ± 1.75 l–o |
| Ś13 | 6.59 ± 0.56 b | 11.75 ± 0.29 ab | nd r | 3.50 ± 1.12 p–q | 2.50 ± 0.54 o–t | 14.77 ± 0.76 f–h | 42.29 ± 2.12 e–g | 61.37 ± 1.99 b–e |
| Ś14 | 4.67 ± 0.47 f–l | 9.08 ± 0.43 d–i | 29.78 ± 2.88 i–m | 10.44 ± 1.01 i–k | 2.66 ± 0.83 n–s | 10.73 ± 0.99 k–o | 39.43 ± 3.21 f–i | 51.88 ± 2.72 e–i |
| Ś15 | 2.45 ± 0.24 pq | 3.57 ± 0.30 q | 42.34 ± 2.71 c–e | 14.98 ± 2.43 d–f | 4.82 ± 0.88 c | 18.65 ± 0.73 c–e | 61.72 ± 3.41 a | 64.72 ± 2.41 bc |
| Ś16 | 4.18 ± 0.33 j–n | 7.13 ± 0.16 k–p | 5.24 ± 0.31 r | 5.14 ± 0.32 o–p | 2.77 ± 0.34 l–q | 11.40 ± 0.67 j–m | 33.11 ± 2.12 i–l | 46.90 ± 1.99 h–l |
| Ś17 | 3.10 ± 0.20 o–q | 5.59 ± 0.08 p | 47.44 ± 2.54 c | 19.72 ± 2.11 b | 3.63 ± 0.88 f–j | 24.67 ±0.78 b | 44.33 ± 2.16 c–f | 56.10 ± 1.62 c–h |
| Ś18 | 5.46 ± 0.45 c–g | 9.85 ± 0.38 c–g | 27.15 ± 1.11 k–n | 8.56 ± 0.67 k–m | 2.09 ± 0.75 r–u | 12.40 ± 0.65 h–k | 20.62 ± 2.11 n–p | 39.34 ± 1.61 k–n |
| Ś19 | 4.94 ± 0.25 e–k | 8.88 ± 0.18 e–j | 21.89 ± 1.43 n–p | 1.89 ± 0.12 qr | 1.65 ± 0.54 u–v | 9.94 ± 0.48 k–p | 7.62 ± 0.54 q | 29.06 ± 2.84 op |
| Ś20 | 6.07 ± 0.62 b–d | 11.33 ± 0.28 bc | 32.68 ± 2.52 g–j | 11.75 ± 1.12 g–i | 2.68 ± 0.78 n–r | 9.83 ± 0.99 k–q | 33.03 ± 2.54 i–l | 48.11 ± 2.91 g–k |
| Ś21 | 3.25 ± 0.12 n–p | 6.88 ± 0.24 l–p | 53.57 ± 1.54 b | 11.49 ± 0.32 g–i | 3.44 ± 0.45 g–l | 17.07 ± 0.69 d–f | 50.77 ± 1.12 bc | 48.76 ± 1.63 g–k |
| Ś22 | 2.64 ± 0.4 pq | 6.06 ± 0.17 op | 37.69 ± 1.11 e–g | 11.67 ± 1.01 g–i | 1.40 ± 0.56 vw | 8.40 ± 0.67 o–t | 30.36 ± 1.25 k–m | 28.93 ± 1.45 op |
| Ś23 | 5.04 ± 0.35 e–j | 10.60 ± 0.65 b–d | 28.23 ± 0.65 j–m | 7.68 ± 0.13 l–n | 2.76 ± 0.54 m–r | 12.30 ± 0.73 h–l | 41.97 ± 1.23 e–g | 51.07 ± 1.63 f–j |
| Ś24 | 5.59 ± 0.89 c–f | 9.36 ± 0.14 d–i | nd r | 15.65 ± 1.23 d | 3.53 ± 0.65 g–k | 6.09 ± 0.56 s–v | 38.77 ± 1.32 f–j | 57.01 ± 1.52 c–g |
| Ś25 | 4.84 ± 0.20 e–k | 8.68 ± 0.12 f–k | 19.79 ± 1.52 pq | 10.31 ± 1.16 i–k | 4.45 ± 0.48 c–e | 4.80 ± 0.48 uv | 50.87 ±1.45 bc | 65.08 ± 2.72 bc |
| Ś26 | 3.18 ± 0.25 op | 6.48 ± 0.31 m–p | 38.90 ± 1.43 d–f | 19.62 ± 0.21 b | 2.79 ± 0.76 l–p | 12.30 ± 0.99 h–l | 47.47 ± 2.15 b–e | 47.94 ± 1.99 g–p |
| Ś27 | 5.43 ± 0.70 c–h | 10.36 ± 0.36 b–f | 17.18 ± 1.59 pq | 8.66 ± 0.54 km | 3.38 ± 0.41 g–m | 14.01 ± 0.65 g–i | 21.26 ± 2.43 no | 36.64 ± 2.61 m–o |
| Ś28 | 5.08 ± 0.58 e–j | 8.85 ± 0.35 e–j | nd r | 13.15 ± 1.43 f–h | 2.18 ± 0.54 p–u | 7.75 ± 0.46 p–t | 39.41 ± 2.55 f–i | 70.26 ± 2.71 ab |
| Ś29 | 4.52 ± 0.50 h–m | 8.63 ± 0.29 g–k | 21.13 ± 0.54 op | 9.59 ± 1.00 i–l | 1.98 ± 0.43 t–v | 10.51 ± 0.59 k–o | 13.32 ± 2.43 pq | 15.60 ± 2.61 q |
| Ś30 | 4.42 ± 0.29 i–m | 7.81 ± 0.14 i–n | 32.67 ± 2.41 g–j | 13.36 ± 1.01 e–g | 2.25 ± 0.32 p–u | 10.41 ± 0.68 k–o | 18.07 ± 1.65 op | 35.52 ± 1.11 no |
| Ś31 | 2.48 ± 0.11 pq | 3.34 ± 0.08 q | 30.97 ± 2.54 i–m | 14.50 ± 2.11 d–f | 2.94 ± 0.53 k–o | 19.41 ± 0.69 c–d | 40.77 ± 1.67 e–h | 51.43 ± 2.61 f–j |
| Ś32 | 4.90 ± 0.59 e–k | 11.26 ± 0.44 bc | 37.02 ± 1.11 e–h | 16.27 ± 0.43 cd | 1.39 ± 0.75 vw | 9.79 ± 0.62 l–q | 35.38 ± 1.79 g–l | 35.29 ± 2.88 no |
| Ś33 | 4.01 ± 0.18 k–o | 6.23 ± 0.19 n–p | nd r | 10.19 ± 0.41 i–k | 1.99 ± 0.65 s–v | 32.67 ± 0.59 a | 34.26 ± 2.43 h–l | 50.49 ± 2.99 g–i |
| Ś34 | 3.90 ± 0.56 l–o | 6.94 ± 0.47 l–p | 2.63 ± 0.11 r | 6.01 ±0.32 n–o | 3.81 ± 0.47 e–h | 10.26 ± 0.49 k–p | 50.13 ± 2.11 b–d | 64.12 ± 3.99 bc |
| Ś35 | 3.12 ± 0.43 o–q | 6.54 ± 0.21 m–p | 26.44 ± 0.22 l–o | 13.35 ±0.78 e–g | 0.44 ± 0.65 x | 6.00 ± 0.59 t–v | 19.65 ± 2.43 op | nd r |
| Ś36 | 5.64 ± 0.36 c–e | 9.75 ± 0.06 c–g | nd r | 15.50 ± 1.32 de | 4.89 ± 0.56 c | 4.98 ± 0.59 uv | 61.82 ± 1.11 a | 75.73 ± 3.11 a |
| Ś37 | 4.88 ± 0.56 e–k | 9.53 ± 0.46 d–h | 19.47 ± 0.76 pq | 15.97 ± 1.54 d | 3.00 ± 0.47 j–o | 18.52 ± 0.50 c–e | 41.82 ± 2.12 e–h | 62.91 ± 1.64 b–d |
| Ś38 | 5.27 ± 0.34 d–i | 9.68 ± 0.28 c–h | 15.26 ± 1.43 q | 11.07 ± 2.13 h–j | 4.77 ± 0.57 c | 16.81 ± 0.47 ef | 52.47 ± 1.11 b | 53.65 ± 2.61 d–i |
| Ś39 | 6.08 ± 0.67 b–d | 10.36 ± 0.58 b–f | nd r | 9.15 ± 2.11 j–l | 1.28 ± 0.54 w | 15.59 ± 0.38 fg | 37.65 ± 2.65 f–k | 32.30 ± 2.75 n–p |
| Ś40 | 3.35 ± 0.07 n–p | 6.26 ± 0.22 n–p | 43.62 ± 3.16 cd | 18.39 ± 3.01 bc | 2.10 ± 0.54 q–u | 13.49 ± 0.56 g–j | 50.83 ± 2.43 bc | 46.22 ± 1.75 i–m |
| Ś41 | 2.20 ± 0.24 q | 3.25 ± 0.05 q | 61.53 ± 2.47 a | 24.07 ± 2.74 a | 4.03 ± 0.38 d–g | 16.99 ± 0.49 d–f | 45.20 ± 2.13 b–f | 60.45 ± 2.94 b–f |
| Ś42 | 3.61 ± 0.23 m–o | 7.35 ± 0.06 j–o | 40.36 ± 1.11 de | 16.33 ± 2.01 cd | 3.95 ± 0.58 e–g | 11.08 ± 0.39 j–n | 45.15 ± 2.43 b–f | 52.71 ± 1.59 e–i |
| Ś43 | 5.55 ± 0.71 c–f | 9.19 ± 0.47 d–i | 34.58 ± 3.54 f–i | nd r | 4.45 ± 0.54 c–e | 5.81 ± 0.59 t–v | 44.46 ± 0.65 c–f | 61.68 ± 2.01 b–e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rybak, M.; Wojdyło, A. Inhibition of α-Amylase, α-Glucosidase, Pancreatic Lipase, 15-Lipooxygenase and Acetylcholinesterase Modulated by Polyphenolic Compounds, Organic Acids, and Carbohydrates of Prunus domestica Fruit. Antioxidants 2023, 12, 1380. https://doi.org/10.3390/antiox12071380
Rybak M, Wojdyło A. Inhibition of α-Amylase, α-Glucosidase, Pancreatic Lipase, 15-Lipooxygenase and Acetylcholinesterase Modulated by Polyphenolic Compounds, Organic Acids, and Carbohydrates of Prunus domestica Fruit. Antioxidants. 2023; 12(7):1380. https://doi.org/10.3390/antiox12071380
Chicago/Turabian StyleRybak, Martyna, and Aneta Wojdyło. 2023. "Inhibition of α-Amylase, α-Glucosidase, Pancreatic Lipase, 15-Lipooxygenase and Acetylcholinesterase Modulated by Polyphenolic Compounds, Organic Acids, and Carbohydrates of Prunus domestica Fruit" Antioxidants 12, no. 7: 1380. https://doi.org/10.3390/antiox12071380