
The Synergy between Glutathione and Phenols—Phenolic Antioxidants Repair Glutathione: Closing the Virtuous Circle—A Theoretical Insight



Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
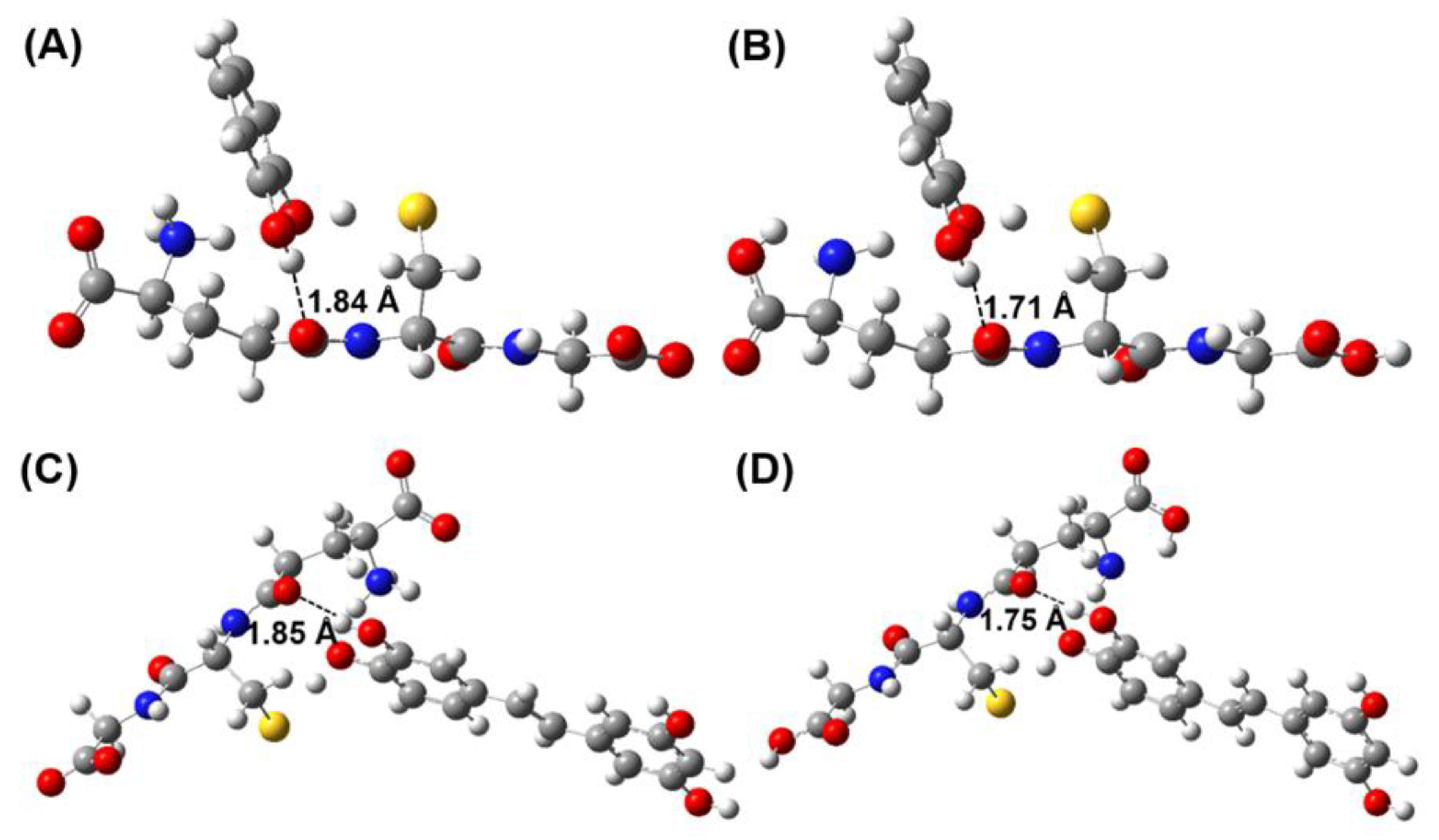
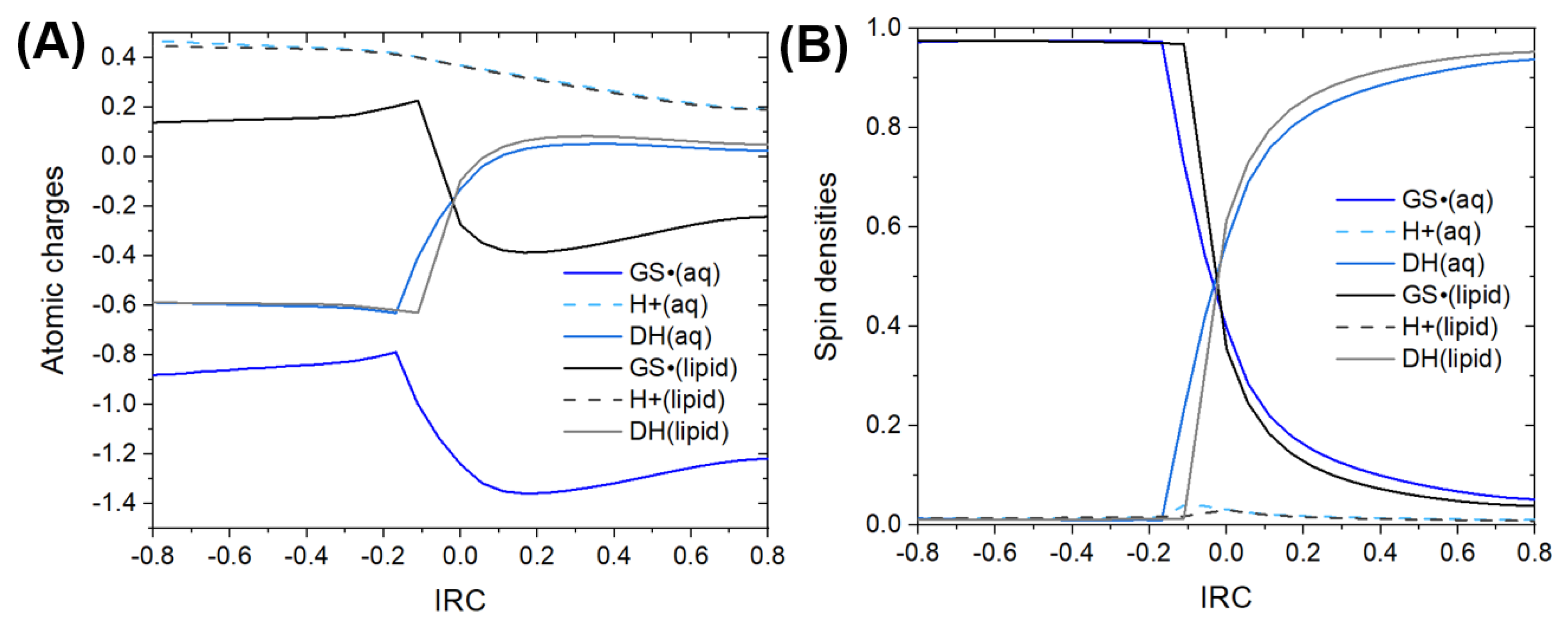
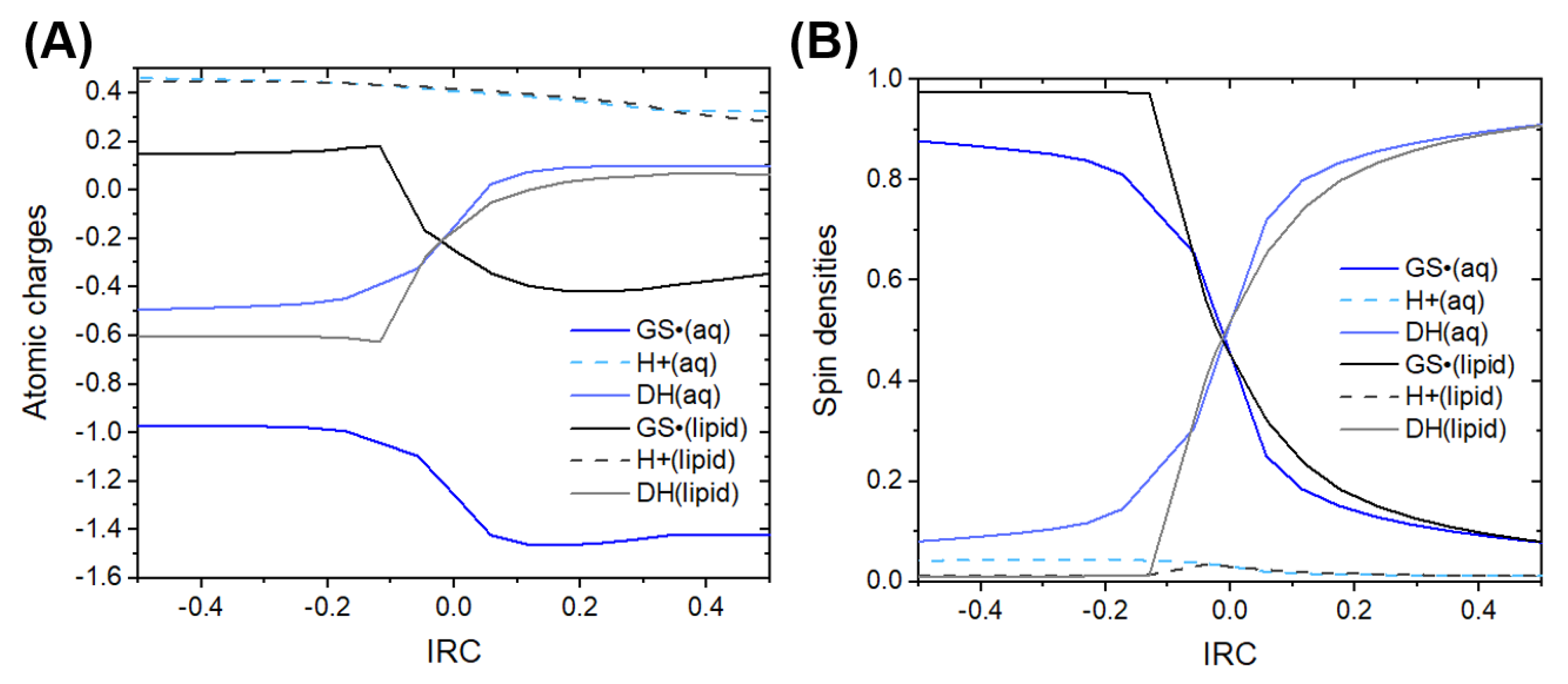
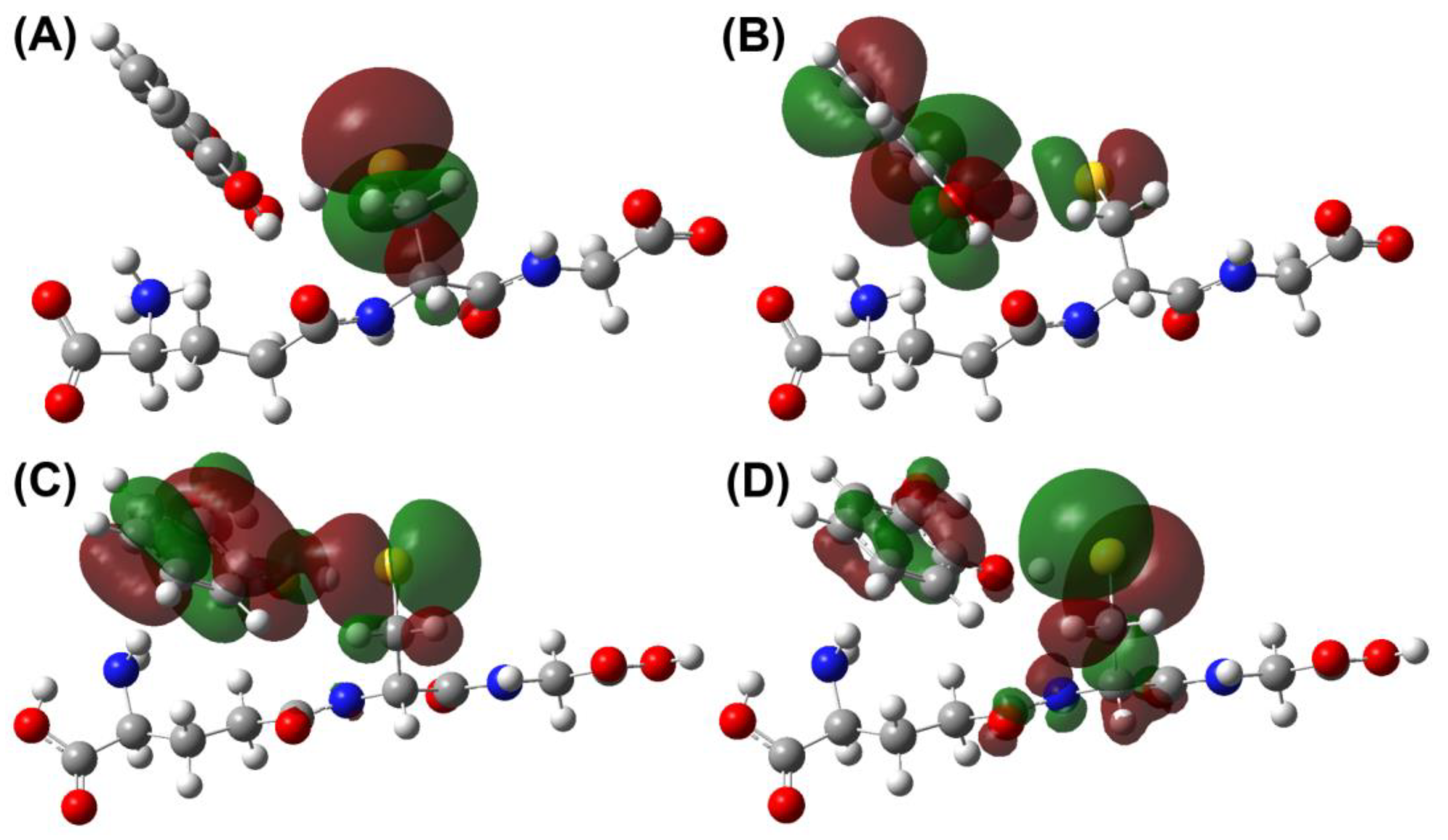
3.1. Thermodynamics and Kinetics
3.2. Mechanistic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halliwell, B. Reactive Species and Antioxidants. Redox Biology Is a Fundamental Theme of Aerobic Life. Plant. Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Biochemistry of Oxidative Stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Genestra, M. Oxyl Radicals, Redox-Sensitive Signalling Cascades and Antioxidants. Cell. Signal. 2007, 19, 1807–1819. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free Radicals, Antioxidants in Disease and Health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Sies, H. Oxidative Stress: Oxidants and Antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Halliwell, B.; Whiteman, M. Measuring Reactive Species and Oxidative Damage in Vivo and in Cell Culture: How Should You Do It and What Do the Results Mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef]
- Stephens, N.G.; Parsons, A.; Brown, M.J.; Schofield, P.M.; Kelly, F.; Cheeseman, K.; Mitchinson, M. Randomised Controlled Trial of Vitamin E in Patients with Coronary Disease: Cambridge Heart Antioxidant Study (CHAOS). Lancet 1996, 347, 781–786. [Google Scholar] [CrossRef]
- Floyd, R.A. Antioxidants, Oxidative Stress, and Degenerative Neurological Disorders. Proc. Soc. Exp. Biol. Med. 1999, 222, 236–245. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, Oxidative Stress and the Biology of Ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of Oxygen Radicals in DNA Damage and Cancer Incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef]
- Sayre, L.M.; Perry, G.; Smith, M.A. Oxidative Stress and Neurotoxicity. Chem. Res. Toxicol. 2008, 21, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Hybertson, B.M.; Gao, B.; Bose, S.K.; McCord, J.M. Oxidative Stress in Health and Disease: The Therapeutic Potential of Nrf2 Activation. Mol. Aspects Med. 2011, 32, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Kelly, F.J.; Fussell, J.C. Role of Oxidative Stress in Cardiovascular Disease Outcomes Following Exposure to Ambient Air Pollution. Free Radic. Biol. Med. 2017, 110, 345–367. [Google Scholar] [CrossRef]
- Halliwell, B. Role of Free Radicals in the Neurodegenerative Diseases: Therapeutic Implications for Antioxidant Treatment. Drugs Aging 2001, 18, 685–716. [Google Scholar] [CrossRef]
- Vasanthi, P.; Nalini, G.; Rajasekhar, G. Status of Oxidative Stress in Rheumatoid Arthritis. Int. J. Rheum. Dis. 2009, 12, 29–33. [Google Scholar] [CrossRef]
- Galano, A. Free Radicals Induced Oxidative Stress at a Molecular Level: The Current Status, Challenges and Perspectives of Computational Chemistry Based Protocols. J. Mex. Chem. Soc. 2017, 59, 231–262. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive Oxygen Species in Living Systems: Source, Biochemistry, and Role in Human Disease. Am. J. Med. 1991, 91, S14–S22. [Google Scholar] [CrossRef]
- Prieto, M.A.; Murado, M.A.; Vázquez, J.A. Quantification, Characterization and Description of Synergy and Antagonism in the Antioxidant Response. Food Res. Int. 2014, 60, 218–229. [Google Scholar] [CrossRef]
- Prieto, M.A.; Curran, T.P.; Gowen, A.; Vázquez, J.A. An Efficient Methodology for Quantification of Synergy and Antagonism in Single Electron Transfer Antioxidant Assays. Food Res. Int. 2015, 67, 284–298. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free Radicals, Metals and Antioxidants in Oxidative Stress-Induced Cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Ran, L.; Chi, Y.; Huang, Y.; He, Q.; Ren, Y. Synergistic Antioxidant Effect of Glutathione and Edible Phenolic Acids and Improvement of the Activity Protection by Coencapsulation into Chitosan-Coated Liposomes. LWT 2020, 127, 109409. [Google Scholar] [CrossRef]
- Marinova, E.; Toneva, A.; Yanishlieva, N. Synergistic Antioxidant Effect of α-Tocopherol and Myricetin on the Autoxidation of Triacylglycerols of Sunflower Oil. Food Chem. 2008, 106, 628–633. [Google Scholar] [CrossRef]
- Hidalgo, M.; Sánchez-Moreno, C.; de Pascual-Teresa, S. Flavonoid–Flavonoid Interaction and Its Effect on Their Antioxidant Activity. Food Chem. 2010, 121, 691–696. [Google Scholar] [CrossRef]
- Casadey, R.; Challier, C.; Senz, A.; Criado, S. Antioxidant Ability of Tyrosol and Derivative-Compounds in the Presence of O2(1Δg)-Species. Studies of Synergistic Antioxidant Effect with Commercial Antioxidants. Food Chem. 2019, 285, 275–281. [Google Scholar] [CrossRef]
- Quiroga, P.R.; Nepote, V.; Baumgartner, M.T. Contribution of Organic Acids to α-Terpinene Antioxidant Activity. Food Chem. 2019, 277, 267–272. [Google Scholar] [CrossRef]
- Marteau, C.; Favier, D.; Nardello-Rataj, V.; Aubry, J.-M. Dramatic Solvent Effect on the Synergy between α-Tocopherol and BHT Antioxidants. Food Chem. 2014, 160, 190–195. [Google Scholar] [CrossRef]
- Bottari, N.B.; Baldissera, M.D.; Tonin, A.A.; Rech, V.C.; Alves, C.B.; D’Avila, F.; Thomé, G.R.; Guarda, N.S.; Moresco, R.N.; Camillo, G.; et al. Synergistic Effects of Resveratrol (Free and Inclusion Complex) and Sulfamethoxazole-Trimetropim Treatment on Pathology, Oxidant/Antioxidant Status and Behavior of Mice Infected with Toxoplasma Gondii. Microb. Pathog. 2016, 95, 166–174. [Google Scholar] [CrossRef]
- Wang, D.; Jiang, Y.; Sun-Waterhouse, D.; Zhai, H.; Guan, H.; Rong, X.; Li, F.; Yu, J.; Li, D. MicroRNA-Based Regulatory Mechanisms Underlying the Synergistic Antioxidant Action of Quercetin and Catechin in H2O2-Stimulated HepG2 Cells: Roles of BACH1 in Nrf2-Dependent Pathways. Free Rad. Biol. Med. 2020, 153, 122–131. [Google Scholar] [CrossRef] [PubMed]
- El-Agamey, A.; Lowe, G.M.; McGarvey, D.J.; Mortensen, A.; Phillip, D.M.; Truscott, T.G.; Young, A.J. Carotenoid Radical Chemistry and Antioxidant/pro-Oxidant Properties. Arch. Biochem. Biophys. 2004, 430, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Poljßak, B.; Raspor, P. The Antioxidant and Pro-Oxidant Activity of Vitamin C and Troloxin Vitro: A Comparative Study. J. Appl. Toxicol. 2008, 28, 183–188. [Google Scholar] [CrossRef] [PubMed]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Pawlowska, E.; Szczepanska, J.; Blasiak, J. Pro- and Antioxidant Effects of Vitamin C in Cancer in Correspondence to Its Dietary and Pharmacological Concentrations. Oxid. Med. Cell Longev. 2019, 2019, 7286737. [Google Scholar] [CrossRef]
- Halliwell, B. Dietary Polyphenols: Good, Bad, or Indifferent for Your Health? Cardiovasc. Res. 2007, 73, 341–347. [Google Scholar] [CrossRef]
- de la Lastra, C.A.; Villegas, I. Resveratrol as an Antioxidant and Pro-Oxidant Agent: Mechanisms and Clinical Implications. Biochem. Soc. Trans. 2007, 35, 1156–1160. [Google Scholar] [CrossRef]
- Halliwell, B. Are Polyphenols Antioxidants or Pro-Oxidants? What Do We Learn from Cell Culture and in Vivo Studies? Arch. Biochem. Biophys. 2008, 476, 107–112. [Google Scholar] [CrossRef]
- Von Sonntag, C. Topics in Free Radical-Mediated DNA. Damage: Purines and Damage Amplification—Superoxic Reactions—Bleomycin, the Incomplete Radiomimetic. Int. J. Radiat. Biol. 1994, 66, 485–490. [Google Scholar] [CrossRef]
- Misiaszek, R.; Crean, C.; Joffe, A.; Geacintov, N.E.; Shafirovich, V. Oxidative DNA Damage Associated with Combination of Guanine and Superoxide Radicals and Repair Mechanisms via Radical Trapping. J. Biol. Chem. 2004, 279, 32106–32115. [Google Scholar] [CrossRef]
- Osakada, Y.; Kawai, K.; Tachikawa, T.; Fujitsuka, M.; Tainaka, K.; Tero-Kubota, S.; Majima, T. Generation of Singlet Oxygen during Photosensitized One-Electron Oxidation of DNA. Chem. Eur. J. 2012, 18, 1060–1063. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Rugeles, L.; Galano, A.; Alvarez-Idaboy, J.R. The Other Side of the Superoxide Radical Anion: Its Ability to Chemically Repair DNA Oxidized Sites. Chem. Commun. 2018, 54, 13710–13713. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Pérez-González, A. On the free radical scavenging mechanism of protocatechuic acid, regeneration of the catechol group in aqueous solution. Theor. Chem. Acc. 2012, 131, 1265. [Google Scholar] [CrossRef]
- Galano, A.; Francisco Marquez, M.; Pérez-González, A. Ellagic Acid: An Unusually Versatile Protector against Oxidative Stress. Chem. Res. Toxicol. 2014, 27, 904–918. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Diduk, R.; Galano, A. Adrenaline and Noradrenaline: Protectors against Oxidative Stress or Molecular Targets? J. Phys. Chem. B 2015, 119, 3479–3491. [Google Scholar] [CrossRef]
- Guo, Y.; Baschieri, A.; Mollica, F.; Valgimigli, L.; Cedrowski, J.; Litwinienko, G.; Amorati, R. Hydrogen Atom Transfer from HOO• to ortho-Quinones Explains the Antioxidant Activity of Polydopamine. Angew. Chem. Int. Ed. 2021, 60, 15220–15224. [Google Scholar] [CrossRef]
- Baschieri, A.; Valgimigli, L.; Gabbanini, S.; DiLabio, G.A.; Romero-Montalvo, E.; Amorati, R. Extremely Fast Hydrogen Atom Transfer between Nitroxides and HOO• Radicals and Implication for Catalytic Coantioxidant Systems. J. Am. Chem. Soc. 2018, 140, 10354–10362. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Álvarez-Diduk, R.; Ramírez-Silva, M.T.; Alarcón-Ángeles, G.; Rojas-Hernández, A. Role of the Reacting Free Radicals on the Antioxidant Mechanism of Curcumin. Chem. Phys. 2009, 363, 13–23. [Google Scholar] [CrossRef]
- Qian, Y.P.; Shang, Y.J.; Teng, Q.F.; Chang, J.; Fan, G.J.; Wei, X.; Li, R.R.; Li, H.P.; Yao, X.J.; Dai, F.; et al. Hydroxychalcones as potent antioxidants: Structure–activity relationship analysis and mechanism considerations. Food Chem. 2011, 126, 241–248. [Google Scholar] [CrossRef]
- Xue, Y.; Liu, Y.; Luo, Q.; Wang, H.; Chen, R.; Liu, Y.; Li, Y. Antiradical Activity and Mechanism of Coumarin-Chalcone Hybrids: Theoretical Insights. J. Phys. Chem. A 2018, 122, 8520–8529. [Google Scholar] [CrossRef]
- Vo, Q.V.; Bay, M.V.; Nam, P.C.; Quang, D.T.; Flavel, M.; Hoa, N.T.; Mechler, A. Theoretical and Experimental Studies of the Antioxidant and Antinitrosant Activity of Syringic Acid. J. Org. Chem. 2020, 85, 15514–15520. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Vashistha, V.K.; Das, D.K. Recent Advances in the Antioxidant Activity and Mechanisms of Chalcone Derivatives: A Computational Review. Free Radic. Res. 2022, 56, 378–397. [Google Scholar] [CrossRef] [PubMed]
- Robillard, B.; Hughes, L.; Slaby, M.; Lindsay, D.A.; Ingold, K.U. Synthesis of 2-Substituted 5,7,8-Trimethyl-6-Hydroxythiochromans and Purported Syntheses of Sulfur-Containing Analogs of Vitamin E. J. Org. Chem. 1986, 51, 1700–1704. [Google Scholar] [CrossRef]
- Giles, G.I.; Tasker, K.M.; Jacob, C. Hypothesis: The Role of Reactive Sulfur Species in Oxidative Stress. Free Rad. Biol. Med. 2001, 31, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.; Giles, G.I.; Giles, N.M.; Sies, H. Sulfur and Selenium: The Role of Oxidation State in Protein Structure and Function. Angew. Chem. Int. Ed. 2003, 42, 4742–4758. [Google Scholar] [CrossRef]
- Townsend, D.M.; Tew, K.D.; Tapiero, H. Sulfur Containing Amino Acids and Human Disease. Biomed. Pharmacother. 2004, 58, 47–55. [Google Scholar] [CrossRef]
- Battin, E.E.; Brumaghim, J.L. Antioxidant Activity of Sulfur and Selenium: A Review of Reactive Oxygen Species Scavenging, Glutathione Peroxidase, and Metal-Binding Antioxidant Mechanisms. Cell Biochem. Biophys. 2009, 55, 1–23. [Google Scholar] [CrossRef]
- Ingenbleek, Y.; Kimura, H. Nutritional Essentiality of Sulfur in Health and Disease. Nutr. Rev. 2013, 71, 413–432. [Google Scholar] [CrossRef]
- Soares Netto, L.E.; De Oliveira, M.A.; Monteiro, G.; Dias Demasi, A.P.; Rosa Cussiol, J.R.; Fulan Discola, K.; Demasi, M.; Monteiro Silva, G.; Vidigal Alves, S.; Genu Faria, V.; et al. Reactive Cysteine in Proteins: Protein Folding, Antioxidant Defense, Redox Signaling and More. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 180–193. [Google Scholar] [CrossRef]
- Altomare, A.; Baron, G.; Brioschi, M.; Longoni, M.; Butti, R.; Valvassori, E.; Tremoli, E.; Carini, M.; Agostoni, P.; Vistoli, G.; et al. N-Acetyl-Cysteine Regenerates Albumin Cys34 by a Thiol-Disulfide Breaking Mechanism: An Explanation of Its Extracellular Antioxidant Activity. Antioxidants 2020, 9, 367. [Google Scholar] [CrossRef]
- McIntyre, N.R.; Lowe, E.W.; Chew, G.H.; Owen, T.C.; Merkler, D.J. Thiorphan, Tiopronin, and Related Analogs as Substrates and Inhibitors of Peptidylglycine α-Amidating Monooxygenase (PAM). FEBS Lett. 2006, 580, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Meister, A. Glutathione Metabolism and Its Selective Modification. J. Biol. Chem. 1988, 263, 17205–17208. [Google Scholar] [CrossRef] [PubMed]
- Kerksick, C.; Willoughby, D. The Antioxidant Role of Glutathione and N-Acetyl-Cysteine Supplements and Exercise-Induced Oxidative Stress. J. Int. Soc. Sports. Nutr. 2005, 2, 38–44. [Google Scholar] [CrossRef]
- Marí, M.; Morales, A.; Colell, A.; Garcıía-Ruiz, C.; Fernández-Checa, J.C. Mitochondrial Glutathione, a Key Survival Antioxidant. Antioxid. Redox Signal. 2009, 11, 2685–2700. [Google Scholar] [CrossRef] [PubMed]
- Gaucher, C.; Boudier, A.; Bonetti, J.; Clarot, I.; Leroy, P.; Parent, M. Glutathione: Antioxidant Properties Dedicated to Nanotechnologies. Antioxidants 2018, 7, 62. [Google Scholar] [CrossRef]
- Alvarez-Idaboy, J.R.; Galano, A. On the Chemical Repair of DNA Radicals by Glutathione: Hydrogen vs Electron Transfer. J. Phys. Chem. B 2012, 116, 9316–9325. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Barberán, F.A.; Andrés-Lacueva, C. Polyphenols and Health: Current State and Progress. J. Agric. Food Chem. 2012, 60, 8773–8775. [Google Scholar] [CrossRef]
- Carreon-Gonzalez, M.; Muñoz-Rugeles, L.; Vivier-Bunge, A.; Alvarez-Idaboy, J.R. Chemical Repair of Damaged Leucine and Tryptophane by Thiophenols at Close to Diffusion-controlled Rates: Mechanisms and Kinetics. J. Comput. Chem. 2022, 43, 556–567. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 289, Catechol. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Catechol (accessed on 10 May 2023).
- Kocaçalışkan, I.; Talan, I.; Terzi, I. Antimicrobial Activity of Catechol and Pyrogallol as Allelochemicals. Z. Naturforsch. C. J. Biosci. 2006, 61, 639–642. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 1057, Pyrogallol. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Pyrogallol (accessed on 10 May 2023).
- Maroyi, A. From Traditional Usage to Pharmacological Evidence: Systematic Review of Gunnera Perpensa L. Evid. Based Complement. Alternat. Med. 2016, 2016, 1720123. [Google Scholar] [CrossRef]
- Boubertakh, B.; Liu, X.G.; Cheng, X.L.; Li, P. A Spotlight on Chemical Constituents and Pharmacological Activities of Nigella Glandulifera Freyn et Sint Seeds. J. Chem. 2013, 2013, 820183. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, Q.; Hu, X. A Comprehensive Review of Ethnopharmacological Uses, Phytochemistry, Biological Activities, and Future Prospects of Nigella Glandulifera. Med. Chem. Res. 2020, 29, 1168–1186. [Google Scholar] [CrossRef]
- Yang, C.J.; Wang, C.S.; Hung, J.Y.; Huang, H.W.; Chia, Y.C.; Wang, P.H.; Weng, C.F.; Huang, M.S. Pyrogallol Induces G2-M Arrest in Human Lung Cancer Cells and Inhibits Tumor Growth in an Animal Model. Lung Cancer 2009, 66, 162–168. [Google Scholar] [CrossRef]
- Yilmaz, Y.; Toledo, R.T. Health Aspects of Functional Grape Seed Constituents. Trends. Food Sci. Technol. 2004, 15, 422–433. [Google Scholar] [CrossRef]
- Wenzel, E.; Somoza, V. Metabolism and Bioavailability Oftrans-Resveratrol. Mol. Nutr. Food Res. 2005, 49, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Cottart, C.H.; Nivet-Antoine, V.; Laguillier-Morizot, C.; Beaudeux, J.L. Resveratrol Bioavailability and Toxicity in Humans. Mol. Nutr. Food Res. 2010, 54, 7–16. [Google Scholar] [CrossRef]
- Gambini, J.; López-Grueso, R.; Olaso-González, G.; Inglés, M.; Abdelazid, K.; El Alami, M.; Bonet-Costa, V.; Borrás, C.; Viña, J. Resveratrol: Distribución, propiedades y perspectivas. Rev. Esp. Geriatr. Gerontol. 2013, 48, 79–88. [Google Scholar] [CrossRef]
- Kershaw, J.; Kim, K.-H. The Therapeutic Potential of Piceatannol, a Natural Stilbene, in Metabolic Diseases: A Review. J. Med. Food 2017, 20, 427–438. [Google Scholar] [CrossRef]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary Phenolics: Chemistry, Bioavailability and Effects on Health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the Polyphenols: Status and Controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef]
- Bohn, T. Dietary Factors Affecting Polyphenol Bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, A.; Inai, M.; Masuda, T. Chemical Evidence for the Synergistic Effect of a Cysteinyl Thiol on the Antioxidant Activity of Caffeic and Dihydrocaffeic Esters. Food Chem. 2013, 138, 1483–1492. [Google Scholar] [CrossRef]
- Amorati, R.; Valgimigli, L.; Dinér, P.; Bakhtiari, K.; Saeedi, M.; Engman, L. Multi-Faceted Reactivity of Alkyltellurophenols Towards Peroxyl Radicals: Catalytic Antioxidant Versus Thiol-Depletion Effect. Chem. Eur. J. 2013, 19, 7510–7522. [Google Scholar] [CrossRef]
- Alfieri, M.L.; Panzella, L.; Amorati, R.; Cariola, A.; Valgimigli, L.; Napolitano, A. Role of Sulphur and Heavier Chalcogens on the Antioxidant Power and Bioactivity of Natural Phenolic Compounds. Biomolecules 2022, 12, 90. [Google Scholar] [CrossRef] [PubMed]
- Milligan, J.R.; Tran, N.Q.; Ly, A.; Ward, J.F. Peptide Repair of Oxidative DNA Damage. Biochemistry 2004, 43, 5102–5108. [Google Scholar] [CrossRef]
- Gebicki, J.M.; Nauser, T.; Domazou, A.; Steinmann, D.; Bounds, P.L.; Koppenol, W.H. Reduction of Protein Radicals by GSH and Ascorbate: Potential Biological Significance. Amino Acids 2010, 39, 1131–1137. [Google Scholar] [CrossRef]
- Houée-Lévin, C.; Bobrowski, K.; Horakova, L.; Karademir, B.; Schöneich, C.; Davies, M.J.; Spickett, C.M. Exploring Oxidative Modifications of Tyrosine: An Update on Mechanisms of Formation, Advances in Analysis and Biological Consequences. Free Rad. Res. 2015, 49, 347–373. [Google Scholar] [CrossRef]
- Baranowska, M.; Suliborska, K.; Chrzanowski, W.; Kusznierewicz, B.; Namieśnik, J.; Bartoszek, A. The Relationship between Standard Reduction Potentials of Catechins and Biological Activities Involved in Redox Control. Redox Biol. 2018, 17, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Pittman, R.N. Chapter 4: Oxygen Transport. In Regulation of Tissue Oxygenation; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2011. Available online: https://www.ncbi.nlm.nih.gov/books/NBK54103/ (accessed on 10 May 2023).
- Sharma, S.; Hashmi, M.F. Partial Pressure Of Oxygen; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK493219/ (accessed on 10 May 2023).
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A. (Eds.) Gaussian 09, Revision E.01; Gaussian Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Zhao, Y.; Truhlar, D.G. The M06 Suite of Density Functionals for Main Group Thermochemistry, Thermochemical Kinetics, Noncovalent Interactions, Excited States, and Transition Elements: Two New Functionals and Systematic Testing of Four M06-Class Functionals and 12 Other Functionals. Theor. Chem. Acc. 2008, 120, 215–241. [Google Scholar] [CrossRef]
- Marenich, A.V.; Cramer, C.J.; Truhlar, D.G. Universal Solvation Model Based on Solute Electron Density and on a Continuum Model of the Solvent Defined by the Bulk Dielectric Constant and Atomic Surface Tensions. J. Phys. Chem. B 2009, 113, 6378–6396. [Google Scholar] [CrossRef]
- Okuno, Y. Theoretical Investigation of the Mechanism of the Baeyer-Villiger Reaction in Nonpolar Solvents. Chem. Eur. J. 1997, 3, 212–218. [Google Scholar] [CrossRef]
- Eyring, H. The Activated Complex in Chemical Reactions. J. Chem. Phys. 1935, 2, 107–115. [Google Scholar] [CrossRef]
- Evans, M.G.; Polanyi, M. Some Applications of the Transition State Method to the Calculation of Reaction Velocities, Especially in Solution. Trans. Faraday Soc. 1935, 31, 875–894. [Google Scholar] [CrossRef]
- Eckart, C. The Penetration of a Potential Barrier by Electrons. Phys. Rev. 1930, 35, 1303–1309. [Google Scholar] [CrossRef]
- Marcus, R.A. On the Theory of Oxidation-Reduction Reactions Involving Electron Transfer. I. J. Chem. Phys. 1956, 24, 966–978. [Google Scholar] [CrossRef]
- Marcus, R.A. On the Theory of Oxidation-Reduction Reactions Involving Electron Transfer. II. Applications to Data on the Rates of Isotopic Exchange Reactions. J. Chem. Phys. 1957, 26, 867–871. [Google Scholar] [CrossRef]
- Marcus, R.A. On the Theory of Oxidation-Reduction Reactions Involving Electron Transfer. III. Applications to Data on the Rates of Organic Redox Reactions. J. Chem. Phys. 1957, 26, 872–877. [Google Scholar] [CrossRef]
- Marcus, R.A.; Sutin, N. Electron Transfers in Chemistry and Biology. Biochim. Biophys. Acta 1985, 811, 265–322. [Google Scholar] [CrossRef]
- Marcus, R.A. Electron Transfer Reactions in Chemistry. Theory and Experiment. Rev. Mod. Phys. 1993, 65, 599–610. [Google Scholar] [CrossRef]
- Marcus, R.A. Electron transfer reactions in chemistry. Theory and experiment. J. Electroanal. Chem. 1997, 438, 251–259. [Google Scholar] [CrossRef]
- Collins, F.C.; Kimball, G.E. Diffusion-Controlled Reaction Rates. J. Colloid. Sci. 1949, 4, 425–437. [Google Scholar] [CrossRef]
- Smoluchowski, M.v. Versuch einer mathematischen Theorie der Koagulationskinetik kolloider Lösungen. Z. Phys. Chem. 1918, 92, 129–168. [Google Scholar] [CrossRef]
- Einstein, A. Über die von der molekularkinetischen Theorie der Wärme geforderte Bewegung von in ruhenden Flüssigkeiten suspendierten Teilchen. Ann. Phys. 1905, 322, 549–560. [Google Scholar] [CrossRef]
- Truhlar, D.G. Nearly Encounter-Controlled Reactions: The Equivalence of the Steady-State and Diffusional Viewpoints. J. Chem. Educ. 1985, 62, 104–106. [Google Scholar] [CrossRef]
- Charkoudian, L.K.; Franz, K.J. Fe(III)-Coordination Properties of Neuromelanin Components: 5,6-Dihydroxyindole and 5,6-Dihydroxyindole-2-Carboxylic Acid. Inorg. Chem. 2006, 45, 3657–3664. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, H.; Lu, X.; Chen, C.; Cao, Y.; Xin, Z. Effect of Pyrogallol on the Ring-Opening Polymerization and Curing Kinetics of a Fully Bio-Based Benzoxazine. Thermochim. Acta 2020, 694, 178787–178794. [Google Scholar] [CrossRef]
- Flores da Silva, L.; Crivellaro Guerra, C.; Klein, D.; Bergold, A.M. Solid cation exchange phase to remove interfering anthocyanins in the analysis of other bioactive phenols in red wine. Food Chem. 2017, 227, 158–165. [Google Scholar] [CrossRef]
- Cordova-Gomez, M.; Galano, A.; Alvarez-Idaboy, J.R. Piceatannol, a Better Peroxyl Radical Scavenger than Resveratrol. RSC Adv. 2013, 3, 20209–20218. [Google Scholar] [CrossRef]
- Galano, A.; Alvarez-Idaboy, J.R. A Computational Methodology for Accurate Predictions of Rate Constants in Solution: Application to the Assessment of Primary Antioxidant Activity. J. Comput. Chem. 2013, 34, 2430–2445. [Google Scholar] [CrossRef]
- Litwinienko, G.; Ingold, K.U. Abnormal Solvent Effects on Hydrogen Atom Abstractions. 1. The Reactions of Phenols with 2,2-Diphenyl-1-Picrylhydrazyl (Dpph•) in Alcohols. J. Org. Chem. 2003, 68, 3433–3438. [Google Scholar] [CrossRef]
- Litwinienko, G.; Ingold, K.U. Abnormal Solvent Effects on Hydrogen Atom Abstraction. 2. Resolution of the Curcumin Antioxidant Controversy. The Role of Sequential Proton Loss Electron Transfer. J. Org. Chem. 2004, 69, 5888–5896. [Google Scholar] [CrossRef] [PubMed]
- Litwinienko, G.; Ingold, K.U. Abnormal Solvent Effects on Hydrogen Atom Abstraction. 3. Novel Kinetics in Sequential Proton Loss Electron Transfer Chemistry. J. Org. Chem. 2005, 70, 8982–8990. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.M.; Hrovat, D.A.; Thomas, J.L.; Borden, W.T. Proton-Coupled Electron Transfer versus Hydrogen Atom Transfer in Benzyl/Toluene, Methoxyl/Methanol, and Phenoxyl/Phenol Self-Exchange Reactions. J. Am. Chem. Soc. 2002, 124, 11142–11147. [Google Scholar] [CrossRef]
- Muñoz-Rugeles, L.; Galano, A.; Raúl Alvarez-Idaboy, J. Non-Covalent π-π Stacking Interactions Turn off Non-Adiabatic Effects in Proton-Coupled Electron Transfer Reactions. Phys. Chem. Chem. Phys. 2017, 19, 6969–6972. [Google Scholar] [CrossRef]
- Sirjoosingh, A.; Hammes-Schiffer, S. Proton-Coupled Electron Transfer versus Hydrogen Atom Transfer: Generation of Charge-Localized Diabatic States. J. Phys. Chem. A 2011, 115, 2367–2377. [Google Scholar] [CrossRef]
- Muñoz-Rugeles, L.; Alvarez-Idaboy, J.R. A Proton–Electron Sequential Transfer Mechanism: Theoretical Evidence about Its Biological Relevance. Phys. Chem. Chem. Phys. 2015, 17, 28525–28528. [Google Scholar] [CrossRef]
- Muñoz-Rugeles, L.; Galano, A.; Alvarez-Idaboy, J.R. The Role of Acid–Base Equilibria in Formal Hydrogen Transfer Reactions: Tryptophan Radical Repair by Uric Acid as a Paradigmatic Case. Phys. Chem. Chem. Phys. 2017, 19, 15296–15309. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
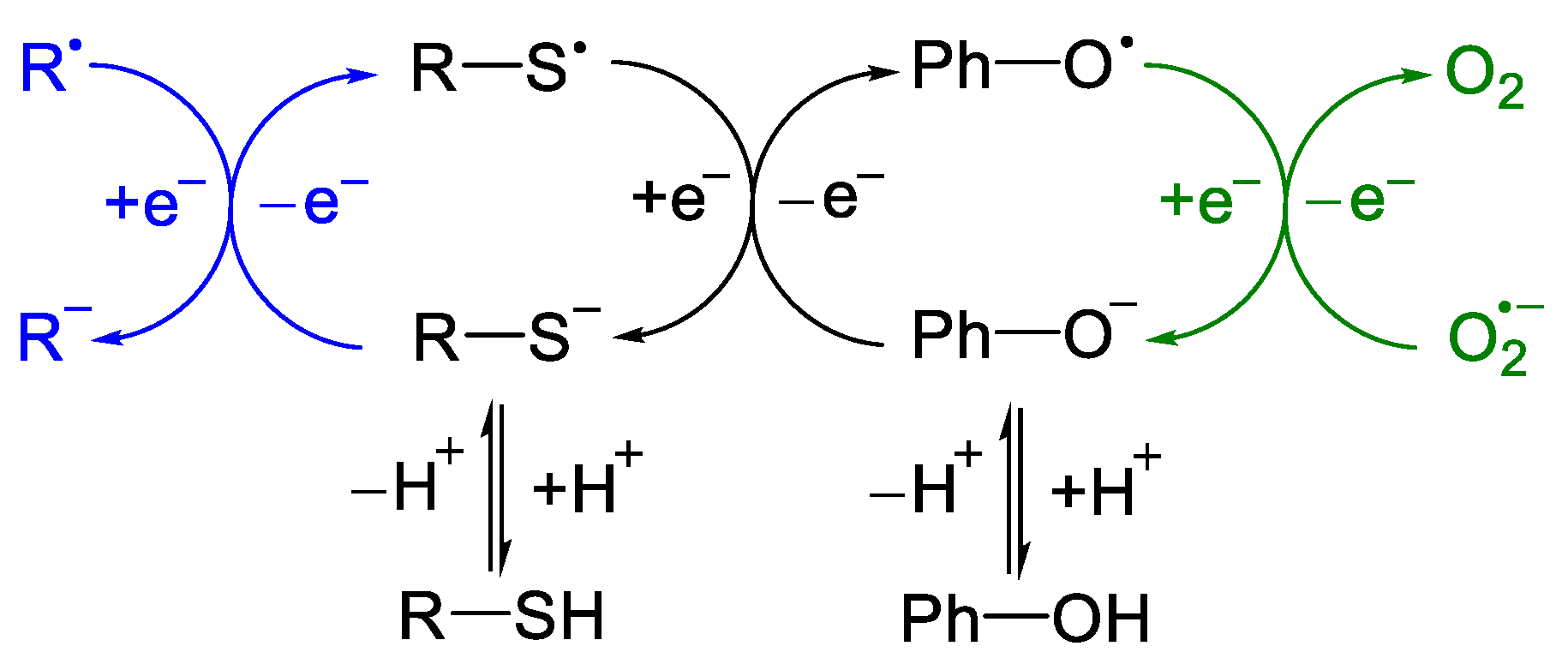{kind=link}
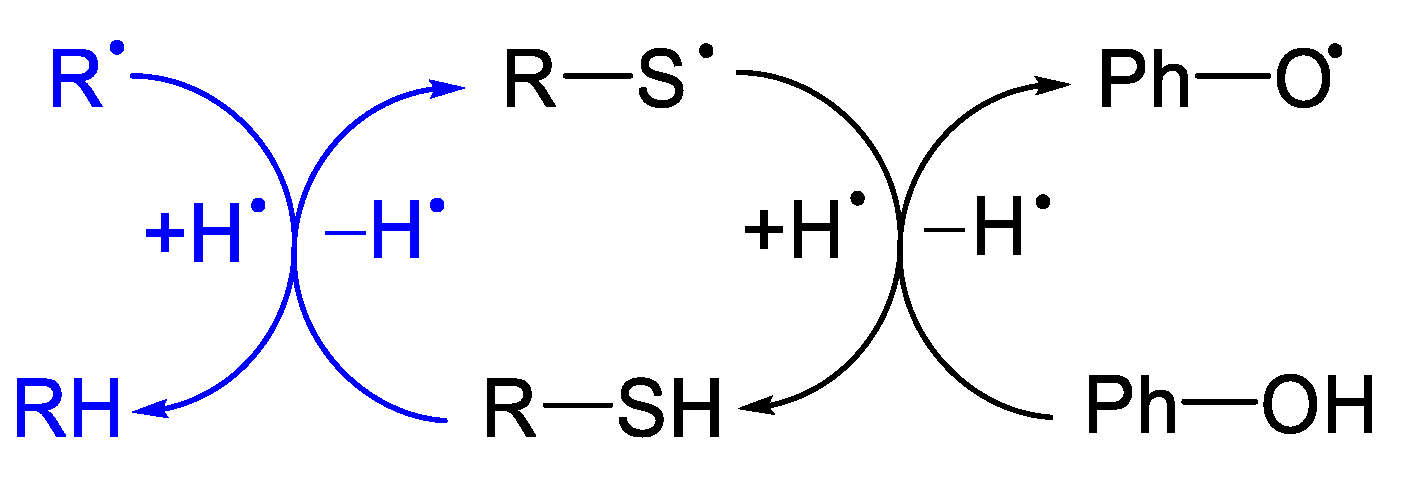{kind=link}
| SET | FHT | ||||||
|---|---|---|---|---|---|---|---|
| Phenol | ktotal [b] | ktotal [b] | koverall [b] | ||||
| Cat | −6.01 | 6.02 | 3.21 × 106 | −2.99 | 13.90 [c] | 1.62 × 104 [c] | 3.21 × 106 [c] |
| 15.17 [d] | 4.21 × 103 [d] | 3.21 × 106 [d] | |||||
| Pyr | −7.19 | 5.64 | 1.16 × 107 | −6.65 | 11.59 | 1.28 × 105 | 1.17 × 107 |
| Res | −6.23 | 4.76 | 3.79 × 107 | −1.15 | 16.46 | 2.31 × 104 | 3.79 × 107 |
| Pic | −8.25 | 4.32 | 6.65 × 108 | −5.81 | 13.23 [c] | 2.15 × 104 [c] | 6.65 × 108 [c] |
| 14.90 [d] | 7.78 × 103 [d] | 6.65 × 108 [d] | |||||
| Phenol | koverall [b] | ||
|---|---|---|---|
| Cat | −5.04 | 7.99 [c] | 8.64 × 106 [c] |
| 11.84 [d] | 1.30 × 104 [d] | ||
| Pyr | −9.22 | 6.75 | 7.04 × 107 |
| Res | −2.28 | 12.82 | 2.80 × 104 |
| Pic | −8.22 | 6.89 [c] | 5.53 × 107 [c] |
| 11.05 [d] | 3.67 × 104 [d] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carreon-Gonzalez, M.; Alvarez-Idaboy, J.R. The Synergy between Glutathione and Phenols—Phenolic Antioxidants Repair Glutathione: Closing the Virtuous Circle—A Theoretical Insight. Antioxidants 2023, 12, 1125. https://doi.org/10.3390/antiox12051125
Carreon-Gonzalez M, Alvarez-Idaboy JR. The Synergy between Glutathione and Phenols—Phenolic Antioxidants Repair Glutathione: Closing the Virtuous Circle—A Theoretical Insight. Antioxidants. 2023; 12(5):1125. https://doi.org/10.3390/antiox12051125
Chicago/Turabian StyleCarreon-Gonzalez, Mirzam, and Juan Raúl Alvarez-Idaboy. 2023. "The Synergy between Glutathione and Phenols—Phenolic Antioxidants Repair Glutathione: Closing the Virtuous Circle—A Theoretical Insight" Antioxidants 12, no. 5: 1125. https://doi.org/10.3390/antiox12051125