
Lignan-Rich Sesame (Sesamum indicum L.) Cultivar Exhibits In Vitro Anti-Cholinesterase Activity, Anti-Neurotoxicity in Amyloid-β Induced SH-SY5Y Cells, and Produces an In Vivo Nootropic Effect in Scopolamine-Induced Memory Impaired Mice

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material and Chemicals
2.2. Preparation of Crude Extracts and Sesame Oil
2.3. Functional Compound Analysis of Different Sesame Varieties
2.4. Antioxidant and Cholinesterase Inhibitory Activities of Different Sesame Varieties In Vitro
2.5. Neuroprotective Effect in Amyloid-β Induced SH-SY5Y Cells of Lignan-Rich Sesame Cultivars
2.6. Treatment of Scopolamine-Induced ICR Mice with Extracts and Oil from Lignan-Rich Sesame Cultivars
2.7. Memory Evaluation Using a Passive Avoidance Test and Water Maze Test Using ICR Mice
2.8. Immunohistochemistry of AChE, ACh, TNF-α, IL-6, and IL-1β Levels in Mouse Brain Tissue Extracts
2.9. Western Blot Analysis of Protein Expression Related to Amyloid-β Accumulation, Neuronal Regeneration, and Inflammation in Mouse Brain Tissue Extracts
3. Results and Discussion
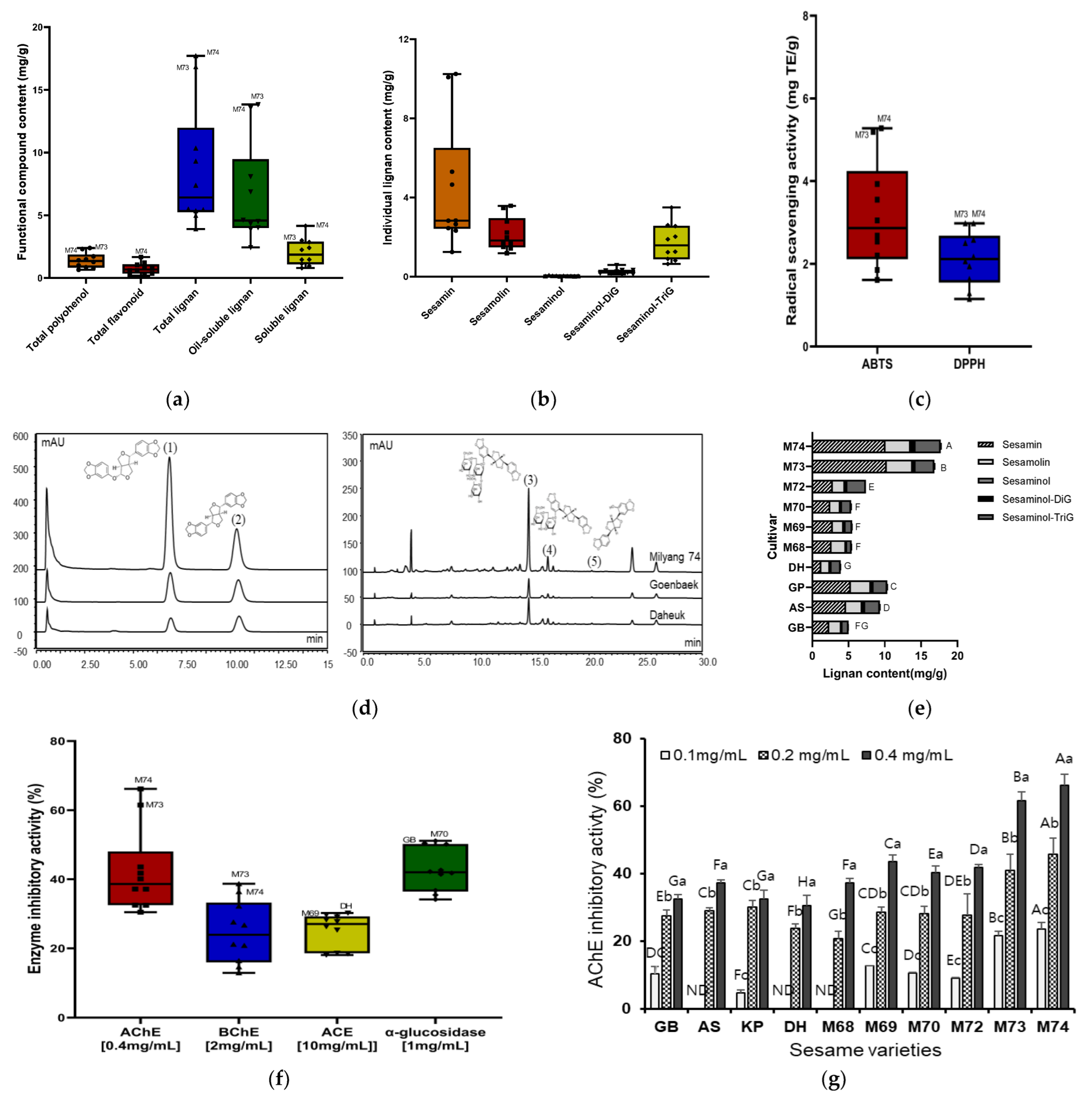
3.1. Screening of Functional Compounds in Different Sesame Varieties
3.2. Screening of Antioxidant Activity and Enzyme Inhibitory Activity in Different Sesame Cultivars
3.3. Neuroprotective Effect of Lignan-Rich Sesame Cultivar in Amyloid-β Induced SH-SY5Y Cell
3.4. Effect of Lignan-Rich Sesame Cultivar Pretreatments on the Improvements of Impaired Memory Functions in Scopolamine-Treated ICR Mice
3.5. Effect of Lignan-Rich Sesame Cultivar Pretreatments on Acetylcholine (ACh) Concentration and Acetylcholinesterase (AChE) Activity in Mouse Brain Tissue Extracts
3.6. Effect of Lignan-Rich Sesame Cultivar Pretreatments on Protein Expression Related to Amyloid-β Accumulation in Mouse Brain Tissue Extracts
3.7. Effect of Lignan-Rich Sesame Cultivar Pretreatments on Protein Expression Related to Neuronal Regeneration in Mouse Brain Tissue Extracts
3.8. Effect of Lignan-Rich Sesame Cultivar Pretreatments on Protein Expression Related to Neuronal Inflammation in Mouse Brain Tissue Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barichello, T.; Giridharan, V.V.; Dal-Pizzol, F. A cerebrospinal fluid biosignature for the diagnosis of alzheimer’s disease. Braz. J. Psychiatry 2019, 41, 467–468. [Google Scholar] [CrossRef] [PubMed]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s disease. Eur. J. Neurol. 2017, 25, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Attems, J.; Jellinger, K.A. The overlap between vascular disease and alzheimer’s disease—Lessons from pathology. BMC Med. 2014, 12, 206. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the cholinergic system. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef]
- Bagheri, M.; Joghataei, M.-T.; Mohseni, S.; Roghani, M. Genistein ameliorates learning and memory deficits in amyloid β(1–40) rat model of alzheimer’s disease. Neurobiol. Learn Mem. 2011, 95, 270–276. [Google Scholar] [CrossRef]
- Shankar, G.M.; Li, S.; Mehta, T.H.; Garcia-Munoz, A.; Shepardson, N.E.; Smith, I.; Brett, F.M.; Farrell, M.A.; Rowan, M.J.; Lemere, C.A.; et al. Amyloid-β protein dimers isolated directly from Alzheimer’s brains impair synaptic plasticity and memory. Nat. Med. 2008, 14, 837–842. [Google Scholar] [CrossRef]
- Zeng, K.-W.; Wang, X.-M.; Ko, H.; Kwon, H.C.; Cha, J.W.; Yang, H.O. Hyperoside protects primary rat cortical neurons from neurotoxicity induced by amyloid β-protein via the PI3K/AKT/bad/BCLXL-regulated mitochondrial apoptotic pathway. Eur. J. Pharmacol. 2011, 672, 45–55. [Google Scholar] [CrossRef]
- Welikovitch, L.A.; Do Carmo, S.; Maglóczky, Z.; Szocsics, P.; Lőke, J.; Freund, T.; Cuello, A.C. Evidence of intraneuronal Aβ accumulation preceding tau pathology in the Entorhinal Cortex. Acta Neuropathol. 2018, 136, 901–917. [Google Scholar] [CrossRef]
- Cha, M.-Y.; Han, S.-H.; Son, S.M.; Hong, H.-S.; Choi, Y.-J.; Byun, J.; Mook-Jung, I. Mitochondria-specific accumulation of amyloid β induces mitochondrial dysfunction leading to apoptotic cell death. PLoS ONE 2012, 7, e34929. [Google Scholar] [CrossRef]
- Ben, E.V.; Michael, W. Volume 3: Chemical constituents, traditional and modern medicinal uses. In Medicinal Plants of the World; CABI: Wallingford, UK, 2017. [Google Scholar]
- Yen, G.-C.; Shyu, S.-L. Oxidative stability of sesame oil prepared from sesame seed with different roasting temperatures. Food Chem. 1989, 31, 215–224. [Google Scholar] [CrossRef]
- Kamal-Eldin, A.; Moazzami, A.; Washi, S. Sesame seed lignans: Potent physiological modulators and possible ingredients in functional foods & nutraceuticals. Recent Patents Food Nutr. Agric. 2011, 3, 17–29. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Massri, M.; Nasrallah, G.K. A comprehensive review on the anti-cancer properties and mechanisms of action of sesamin, a lignan in sesame seeds (Sesamum indicum). Eur. J. Pharmacol. 2017, 815, 512–521. [Google Scholar] [CrossRef]
- Dalibalta, S.; Majdalawieh, A.F.; Manjikian, H. Health benefits of sesamin on cardiovascular disease and its associated risk factors. Saudi Pharm. J. 2020, 28, 1276–1289. [Google Scholar] [CrossRef]
- Keowkase, R.; Shoomarom, N.; Bunargin, W.; Sitthithaworn, W.; Weerapreeyakul, N. Sesamin and sesamolin reduce amyloid-β toxicity in a transgenic caenorhabditis elegans. Biomed. Pharmacother. 2018, 107, 656–664. [Google Scholar] [CrossRef]
- Ruankham, W.; Suwanjang, W.; Wongchitrat, P.; Prachayasittikul, V.; Prachayasittikul, S.; Phopin, K. Sesamin and sesamol attenuate H2O2-induced oxidative stress on human neuronal cells via the SIRT1-SIRT3-Foxo3a signaling pathway. Nutr. Neurosci. 2019, 24, 90–101. [Google Scholar] [CrossRef]
- Hou, R.C.-W.; Chen, Y.-S.; Chen, C.-H.; Chen, Y.-H.; Jeng, K.-C.G. Protective effect of 1,2,4-benzenetriol on LPS-induced NO production by BV2 microglial cells. J. Biomed. Sci. 2005, 13, 89–99. [Google Scholar] [CrossRef]
- Sharma, S.; Kaur, I.P. Development and evaluation of sesamol as an antiaging agent. Int. J. Dermatol. 2006, 45, 200–208. [Google Scholar] [CrossRef]
- Cheng, Y.; Shao, Y.; Yan, W. Sesamin and Sesamolin Contents in various commercial sesame oils. Planta Med. 2011, 77, PJ5. [Google Scholar] [CrossRef]
- Prasad, N.R.; Mahesh, T.; Menon, V.P.; Jeevanram, R.; Pugalendi, K.V. Photoprotective effect of sesamol on UVB-radiation induced oxidative stress in human blood lymphocytes in vitro. Environ. Toxicol. Pharmacol. 2005, 20, 1–5. [Google Scholar] [CrossRef]
- Kim, S.; Lee, M.H.; Oh, E.; Kim, J.I.; Bae, S.B.; Jo, G.S.; Oh, G.W.; Ha, T.J.; Jeong, C.S. A New Sesame Cultivar ‘Milyang 74’ with Enhanced Lignan Content and Use Therof. Korea Patent 10-2020-0133774, 15 October 2020. [Google Scholar]
- Woo, K.S.; Ko, J.Y.; Jeong, H.S. Effect of milling time on antioxidant compounds and activities of methanol extracts of sorghum [Sorghum bicolor (L.) Moench]. Food Sci. Biotechnol. 2014, 23, 1741–1746. [Google Scholar] [CrossRef]
- Park, J.E.; Kim, J.I.; Lee, M.H.; Kim, S.; Oh, E.; Cho, K.S.; Oh, K.W. Influence of roasting temperature on the functional components of Perilla and sesame oils. J. Korean Soc. Food Sci. Nutr. 2021, 50, 149–154. [Google Scholar] [CrossRef]
- Maliszewski, D.; Wróbel, A.; Kolesińska, B.; Frączyk, J.; Drozdowska, D. 1,3,5-triazine nitrogen mustards with different peptide group as innovative candidates for ACHE and BACE1 inhibitors. Molecules 2021, 26, 3942. [Google Scholar] [CrossRef]
- Ishiyama, M.; Tominaga, H.; Shiga, M.; Sasamoto, K.; Ohkura, Y.; Ueno, K. A combined assay of cell viability and in vitro cytotoxicity with a highly water-soluble tetrazolium salt, neutral red and crystal violet. Biol. Pharm. Bull. 1996, 19, 1518–1520. [Google Scholar] [CrossRef]
- Wang, H.; Joseph, J.A. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef]
- Kim, M.Y.; Kim, H.J.; Lee, Y.Y.; Kim, M.H.; Lee, J.Y.; Kang, M.S.; Koo, B.C.; Lee, B.W. Antioxidant and anti-inflammatory effects of peanut (arachishypogaea). skin extracts of various cultivars in oxidative-damaged hepg2 cells and lps-induced raw 264.7 macrophages. Food Sci. Nutr. 2020, 9, 973–984. [Google Scholar] [CrossRef]
- Shi, L.-K.; Liu, R.-J.; Jin, Q.-Z.; Wang, X.-G. The contents of Lignans in sesame seeds and commercial sesame oils of China. J. Am. Oil Chem. Soc. 2017, 94, 1035–1044. [Google Scholar] [CrossRef]
- Siriwarin, B.; Weerapreeyakul, N. Sesamol induced apoptotic effect in lung adenocarcinoma cells through both intrinsic and extrinsic pathways. Chem. Interact. 2016, 254, 109–116. [Google Scholar] [CrossRef]
- Netzel, M.; Strass, G.; Bitsch, I.; Könitz, R.; Christmann, M.; Bitsch, R. Effect of grape processing on selected antioxidant phenolics in red wine. J. Food Eng. 2003, 56, 223–228. [Google Scholar] [CrossRef]
- Ben Othman, S.; Katsuno, N.; Kanamaru, Y.; Yabe, T. Water-soluble extracts from defatted sesame seed flour show antioxidant activity in vitro. Food Chem. 2015, 175, 306–314. [Google Scholar] [CrossRef]
- Ide, T.; Azechi, A.; Kitade, S.; Kunimatsu, Y.; Suzuki, N.; Nakajima, C.; Ogata, N. Comparative effects of sesame seeds differing in Lignan contents and composition on fatty acid oxidation in rat liver. J. Oleo Sci. 2015, 64, 211–222. [Google Scholar] [CrossRef]
- Orhan, I.E. Enzyme inhibitors as the attractive targets for the treatment of various diseases. Curr. Med. Chem. 2019, 26, 3206–3207. [Google Scholar] [CrossRef] [PubMed]
- Ballard, C.; Greig, N.; Guillozet-Bongaarts, A.; Enz, A.; Darvesh, S. Cholinesterases: Roles in the brain during health and disease. Curr. Alzheimer Res. 2005, 2, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Shal, B.; Ding, W.; Ali, H.; Kim, Y.S.; Khan, S. Anti-neuroinflammatory potential of natural products in attenuation of Alzheimer’s disease. Front. Pharmacol. 2018, 9, 548. [Google Scholar] [CrossRef]
- Karavelioglu, Z.; Cakir-Koc, R. Preparation of chitosan nanoparticles as Ginkgo biloba extract carrier: In vitro neuroprotective effect on oxidative stress-induced human neuroblastoma cells (SH-SY5Y). Int. J. Biol. Macromol. 2021, 192, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Parhizkar, S.; Arzberger, T.; Brendel, M.; Kleinberger, G.; Deussing, M.; Focke, C.; Nuscher, B.; Xiong, M.; Ghasemigharagoz, A.; Katzmarski, N.; et al. Loss of TREM2 function increases amyloid seeding but reduces plaque-associated ApoE. Nat. Neurosci. 2019, 22, 191–204. [Google Scholar] [CrossRef]
- Ben Othman, S.; Katsuno, N.; Kitayama, A.; Fujimura, M.; Kitaguchi, K.; Yabe, T. Water-soluble fractions from defatted sesame seeds protect human neuroblast cells against peroxyl radicals and hydrogen peroxide-induced oxidative stress. Free. Radic. Res. 2016, 50, 949–958. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, Y.; Yang, D.; Yuan, F.; Ding, J.; Chen, H.; Tian, H. Sesamin protects SH-SY5Y cells against mechanical stretch injury and promoting cell survival. BMC Neurosci. 2017, 18, 57. [Google Scholar] [CrossRef]
- Mohamed, E.A.; Ahmed, H.I.; Zaky, H.S.; Badr, A.M. Sesame oil mitigates memory impairment, oxidative stress, and neurodegeneration in a rat model of alzheimer’s disease. A pivotal role of NF-ΚB/p38mapk/BDNF/PPAR-γ pathways. J. Ethnopharmacol. 2021, 267, 113468. [Google Scholar] [CrossRef]
- Asliranifam, N.; Najafzadeh, H.; Papahn, A.A.; Moazedi, A.A.; Pourmahdi, M. Effect of sesame oil consumption on the passive avoidance memory of rat offspring during pregnancy. Pharmacol. Physiol. 2011, 15, 268–276. [Google Scholar]
- Fathi, F.; Hatami, H.; Ali Hemmati, A.R.; Banan Khojasteh, S.M. Effects of sesame oil on improving spatial memory in Alzheimers disease. J. Babol. Univ. Med. Sci. 2014, 16, 34–41. [Google Scholar]
- Zare, K.; Fatemi Tabatabaei, S.R.; Shahriari, A.; Jafari, R.A. Effect of butter and sesame oils on avoidance memory of diabetic rats. Iran. J. Diabetes Obes. 2011, 3, 65–71. [Google Scholar]
- Tabari, S.-S.S.; Babri, S.; Mirzaie, F.; Farajdokht, F.; Mohaddes, G. Enduring amnesia induced by ICV scopolamine is reversed by sesame oil in male rats. Acta Cir. Bras. 2016, 31, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Giacobini, E.; Sugaya, K.; Elble, R.J. Markers of Cholinergic Dysfunction in Alzheimer Disease. In Alzheimer’s Disease; CRC Press: Boca Raton, FL, USA, 2020; pp. 137–156. [Google Scholar]
- Hasselmo, M.E. The role of acetylcholine in learning and memory. Curr. Opin. Neurobiol. 2006, 16, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Francis, P.T.; Palmer, A.M.; Snape, M.; Wilcock, G.K. The cholinergic hypothesis of alzheimer’s disease: A review of progress. J. Neurol. Neurosurg Psychiatry 1999, 66, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Birks, J.S. Cholinesterase inhibitors for alzheimer’s disease. Cochrane Database Syst. Rev. 2006, 2016, CD005593. [Google Scholar] [CrossRef]
- Klinkenberg, I.; Blokland, A. The validity of scopolamine as a pharmacological model for cognitive impairment: A Review of Animal Behavioral Studies. Neurosci. Biobehav. Rev. 2010, 34, 1307–1350. [Google Scholar] [CrossRef]
- Ito, N.; Saito, H.; Seki, S.; Ueda, F.; Asada, T. Effects of composite supplement containing Astaxanthin and SESAMIN on cognitive functions in people with mild cognitive impairment: A randomized, double-blind, placebo-controlled trial. J. Alzheimers Dis. 2018, 62, 1767–1775. [Google Scholar] [CrossRef]
- Kuhad, A.; Chopra, K. Effect of sesamol on diabetes-associated cognitive decline in rats. Exp. Brain Res. 2007, 185, 411–420. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, Y.; Qiao, Q.; Sun, Y.; Liu, Q.; Ren, B.; Liu, X. Sesamol supplementation prevents systemic inflammation-induced memory impairment and amyloidogenesis via inhibition of nuclear factor kappab. Mol. Nutr. Food Res. 2016, 61, 1600734. [Google Scholar] [CrossRef]
- Yun, D.; Wang, Y.; Zhang, Y.; Jia, M.; Xie, T.; Zhao, Y.; Yang, C.; Chen, W.; Guo, R.; Liu, X.; et al. Sesamol attenuates scopolamine-induced cholinergic disorders, neuroinflammation, and cognitive deficits in mice. J. Agric. Food Chem. 2022, 70, 13602–13614. [Google Scholar] [CrossRef]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Demuro, A.; Mina, E.; Kayed, R.; Milton, S.C.; Parker, I.; Glabe, C.G. Calcium dysregulation and membrane disruption as a ubiquitous neurotoxic mechanism of soluble amyloid oligomers. J. Biol. Chem. 2005, 280, 17294–17300. [Google Scholar] [CrossRef] [PubMed]
- Scholz, D.; Chernyshova, Y.; Ückert, A.-K.; Leist, M. Reduced AΒ secretion by human neurons under conditions of strongly increased BACE activity. J. Neurochem. 2018, 147, 256–274. [Google Scholar] [CrossRef] [PubMed]
- Kimberly, W.; Xia, W.; Rahmati, T.; Wolfe, M.S.; Selkoe, D.J. The transmembrane aspartates in Presenilin 1 and 2 are obligatory for γ-secretase activity and amyloid β-protein generation. J. Biol. Chem. 2000, 275, 3173–3178. [Google Scholar] [CrossRef]
- Zhao, G.; Tan, J.; Mao, G.; Cui, M.-Z.; Xu, X. The same γ-secretase accounts for the multiple intramembrane cleavages of App. J. Neurochem. 2006, 100, 1234–1246. [Google Scholar] [CrossRef]
- Katayama, S.; Sugiyama, H.; Kushimoto, S.; Uchiyama, Y.; Hirano, M.; Nakamura, S. Effects of sesaminol feeding on brain aβ accumulation in a senescence-accelerated mouse-prone 8. J. Agric. Food Chem. 2016, 64, 4908–4913. [Google Scholar] [CrossRef]
- Spires, T.L.; Orne, J.D.; SantaCruz, K.; Pitstick, R.; Carlson, G.A.; Ashe, K.H.; Hyman, B.T. Region-specific dissociation of neuronal loss and neurofibrillary pathology in a mouse model of tauopathy. Am. J. Pathol. 2006, 168, 1598–1607. [Google Scholar] [CrossRef]
- Lamar, M.; Boots, E.A.; Arfanakis, K.; Barnes, L.L.; Schneider, J.A. Brain structural alterations common to cardiovascular disease risk factors and alzheimer’s dementia. In Vascular Disease, Alzheimer’s Disease, and Mild Cognitive Impairment; Oxford Academic: Oxford, UK, 2020; pp. 241–258. [Google Scholar]
- Xu, C.-J.; Wang, J.-L.; Jin, W.-L. The emerging therapeutic role of NGF in alzheimer’s disease. Neurochem. Res. 2016, 41, 1211–1218. [Google Scholar] [CrossRef]
- Zhang, Y.-wu; Chen, Y.; Liu, Y.; Zhao, Y.; Liao, F.-F.; Xu, H. App regulates NGF receptor trafficking and NGF-mediated neuronal differentiation and survival. PLoS ONE 2013, 8, e80571. [Google Scholar] [CrossRef]
- Chen, S.-D.; Wu, C.-L.; Hwang, W.-C.; Yang, D.-I. More insight into BDNF against neurodegeneration: Anti-apoptosis, anti-oxidation, and suppression of autophagy. Int. J. Mol. Sci. 2017, 18, 545. [Google Scholar] [CrossRef]
- Tanila, H. The role of BDNF in Alzheimer’s disease. Neurobiol. Dis. 2017, 97, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Jiang, X.; Wang, R.; Zhai, S.; Zhang, Y.; Wang, D. Forsythoside B attenuates memory impairment and neuroinflammation via inhibition on NF-ΚB signaling in alzheimer’s disease. J. Neuroinflamm. 2020, 17, 305. [Google Scholar] [CrossRef] [PubMed]
- Bartolotti, N.; Lazarov, O. CREB signals as PBMC-based biomarkers of cognitive dysfunction: A novel perspective of the brain-immune axis. Brain Behav. Immun. 2019, 78, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Q.; Jia, M.; Fu, S.; Pan, J.; Chu, C.; Liu, X.; Liu, X.; Liu, Z. (+)-SESAMIN attenuates chronic unpredictable mild stress-induced depressive-like behaviors and memory deficits via suppression of neuroinflammation. J. Nutr. Biochem. 2018, 64, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Proenca, C.C.; Song, M.; Lee, F.S. Differential effects of BDNF and neurotrophin 4 (NT4) on endocytic sorting of TrkB receptors. J. Neurochem. 2016, 138, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms underlying inflammation in neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H. Inflammation and alzheimer’s disease. Neurobiol. Aging 2000, 21, 383–421. [Google Scholar] [CrossRef]
- Dong, Y.; Benveniste, E.N. Immune function of astrocytes. Glia 2001, 36, 180–190. [Google Scholar] [CrossRef]
- Ahmad, S.; Elsherbiny, N.M.; Haque, R.; Khan, M.B.; Ishrat, T.; Shah, Z.A.; Ali, M.; Jamal, A.; Katare, D.P.; Liou, G.I.; et al. Sesamin attenuates neurotoxicity in mouse model of ischemic brain stroke. Neurotoxicology 2014, 45, 100–110. [Google Scholar] [CrossRef]
- Testa, G.; Gamba, P.; Badilli, U.; Gargiulo, S.; Maina, M.; Guina, T.; Calfapietra, S.; Biasi, F.; Cavalli, R.; Poli, G.; et al. Loading into nanoparticles improves quercetin’s efficacy in preventing neuroinflammation induced by oxysterols. PLoS ONE 2014, 9, e96795. [Google Scholar] [CrossRef]
- Jo, S.-H.; Kang, T.-B.; Koppula, S.; Cho, D.-Y.; Kim, J.-S.; Kim, I.-S.; Choi, D.-K. Mitigating effect of Lindera obtusiloba Blume extract on neuroinflammation in microglial cells and scopolamine-induced amnesia in mice. Molecules 2021, 26, 2870. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Yuan, T.; Zhang, X.; Wang, L.; Pan, J.; Liu, Y.; Zhao, B.; Zhao, W.; Liu, Z.; Liu, X. Protective effects of Sesamol on systemic inflammation and cognitive impairment in aging mice. J. Agric. Food Chem. 2020, 68, 3099–3111. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | 1 MDA | 2 GR | 3 GPX | 4 CAT | 5 SOD |
|---|---|---|---|---|---|
| Control | 0.6 ± 0.02 | 10.32 ± 1.21 | 42.92 ± 1.97 | 5.26 ± 0.84 | 1.3 ± 0.26 |
| Aβ | 1.15 ± 0.11 ### | 29.92 ± 1.37 ### | 92.49 ± 2.52 ### | 23.92 ± 2.66 ### | 14.46 ± 2.65 ### |
| Aβ + DH 100 | 0.79 ± 0.01 a | 18.69 ± 1.97 a | 63.82 ± 3.51 a | 18.22 ± 2.00 a | 4.54 ± 0.88 a |
| Aβ + GB 100 | 0.85 ± 0.05 a | 17.47 ± 0.98 a | 54.71 ± 3.50 b | 14.71 ± 0.74 b | 3.48 ± 0.47 a |
| Aβ + M74 100 | 0.68 ± 0.03 b | 13.36 ± 0.86 b | 39.36 ± 1.56 c | 11.29 ± 1.07 c | 2.17 ± 0.18 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-Y.; Kim, S.; Lee, J.; Kim, J.-I.; Oh, E.; Kim, S.-W.; Lee, E.; Cho, K.-S.; Kim, C.-S.; Lee, M.-H. Lignan-Rich Sesame (Sesamum indicum L.) Cultivar Exhibits In Vitro Anti-Cholinesterase Activity, Anti-Neurotoxicity in Amyloid-β Induced SH-SY5Y Cells, and Produces an In Vivo Nootropic Effect in Scopolamine-Induced Memory Impaired Mice. Antioxidants 2023, 12, 1110. https://doi.org/10.3390/antiox12051110
Kim M-Y, Kim S, Lee J, Kim J-I, Oh E, Kim S-W, Lee E, Cho K-S, Kim C-S, Lee M-H. Lignan-Rich Sesame (Sesamum indicum L.) Cultivar Exhibits In Vitro Anti-Cholinesterase Activity, Anti-Neurotoxicity in Amyloid-β Induced SH-SY5Y Cells, and Produces an In Vivo Nootropic Effect in Scopolamine-Induced Memory Impaired Mice. Antioxidants. 2023; 12(5):1110. https://doi.org/10.3390/antiox12051110
Chicago/Turabian StyleKim, Min-Young, Sungup Kim, Jeongeun Lee, Jung-In Kim, Eunyoung Oh, Sang-Woo Kim, Eunsoo Lee, Kwang-Soo Cho, Choon-Song Kim, and Myoung-Hee Lee. 2023. "Lignan-Rich Sesame (Sesamum indicum L.) Cultivar Exhibits In Vitro Anti-Cholinesterase Activity, Anti-Neurotoxicity in Amyloid-β Induced SH-SY5Y Cells, and Produces an In Vivo Nootropic Effect in Scopolamine-Induced Memory Impaired Mice" Antioxidants 12, no. 5: 1110. https://doi.org/10.3390/antiox12051110