
Antioxidant Effect Assessment and Trans Epithelial Analysis of New Hempseed Protein Hydrolysates
 , , , , ,
, , , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hempseed Hydrolysates (HHs) Production
2.2. In Vitro Antioxidant Activities
2.2.1. 1-Diphenyl-2-picrylhydrazyl Radical (DPPH) Assay
2.2.2. Trolox Equivalent Antioxidant Capacity (TEAC)
2.2.3. Ferric Reducing Antioxidant Power (FRAP) Assay
2.2.4. Oxygen Radical Absorbance Capacity (ORAC) Assay
2.3. Mass Spectrometry Analysis (HPLC Chip ESI-MS/MS)
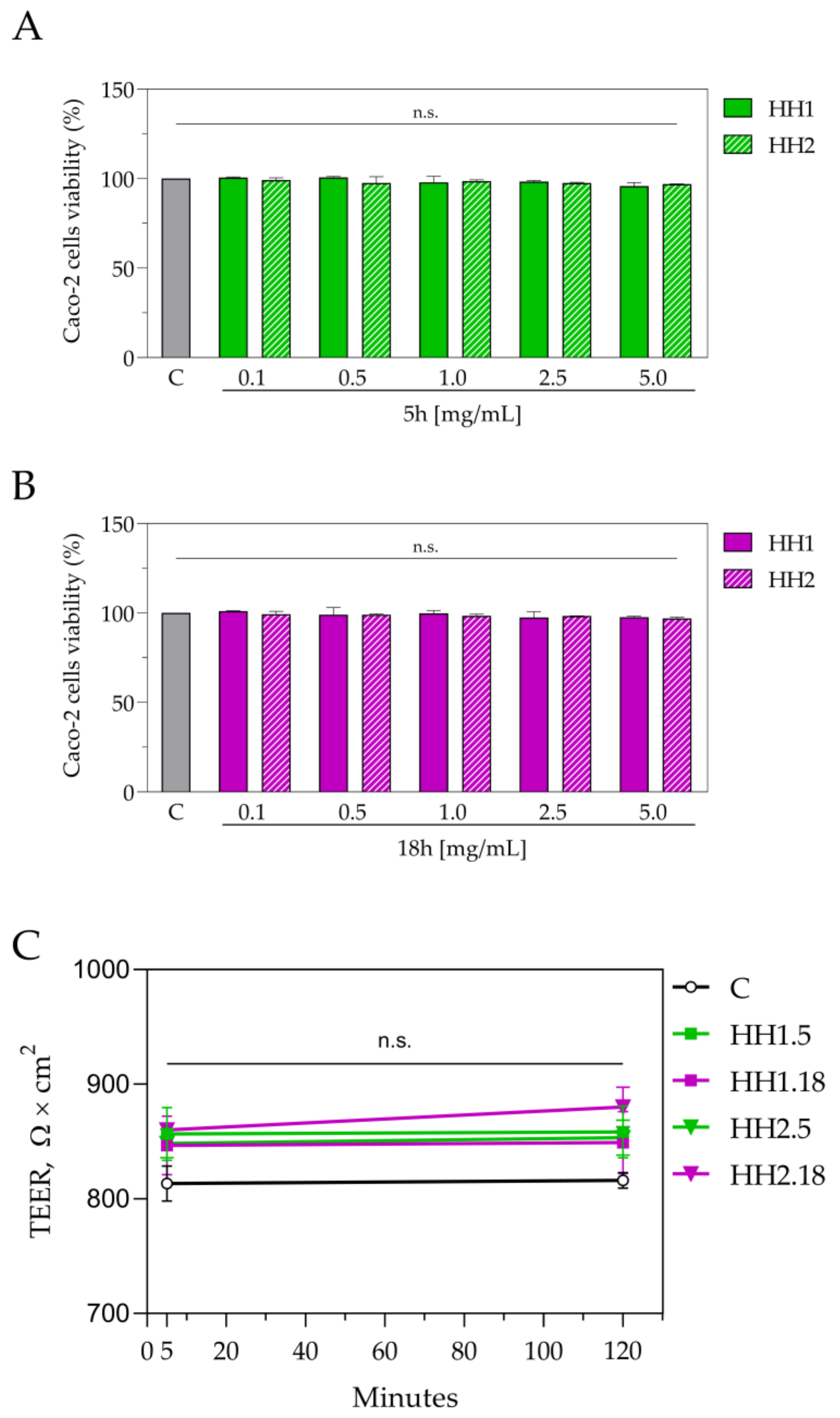
2.4. Cell Culture and Cell Monolayers Integrity Evaluation
2.5. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT) Assay
2.6. Trans-Epithelial Transport of Hemp Hydrolysates
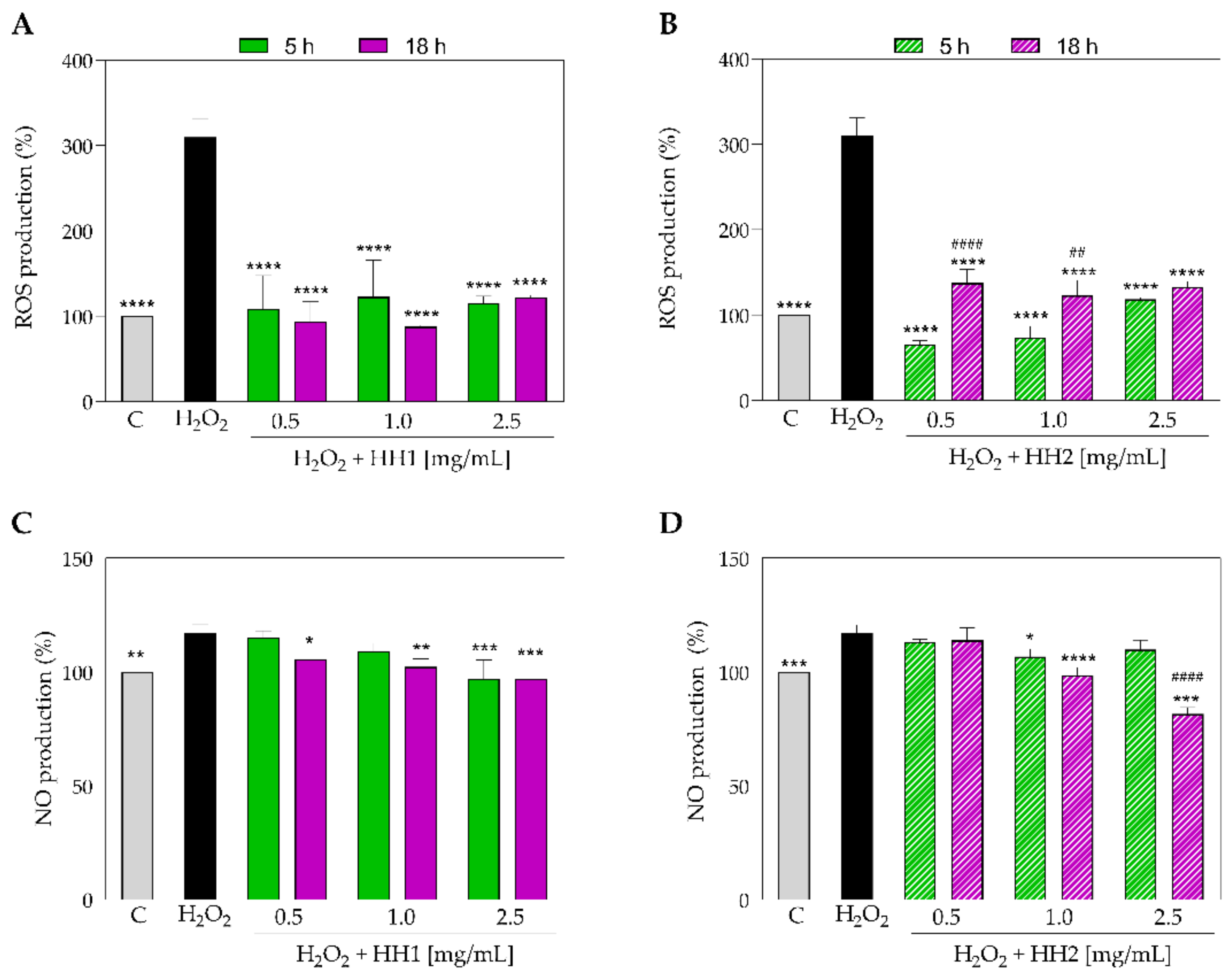
2.7. Fluorometric Intracellular ROS Assay
2.8. Nitric Oxide Level Evaluation on Caco-2 Cells
2.9. DPPH Assay in BL Solution
2.10. Trolox Equivalent Antioxidant Capacity (TEAC) in BL Solution
2.11. Statistical Analysis
3. Results
3.1. In Vitro Antioxidant Capacity
3.1.1. HHs have DPPH Radical Scavenging Activity
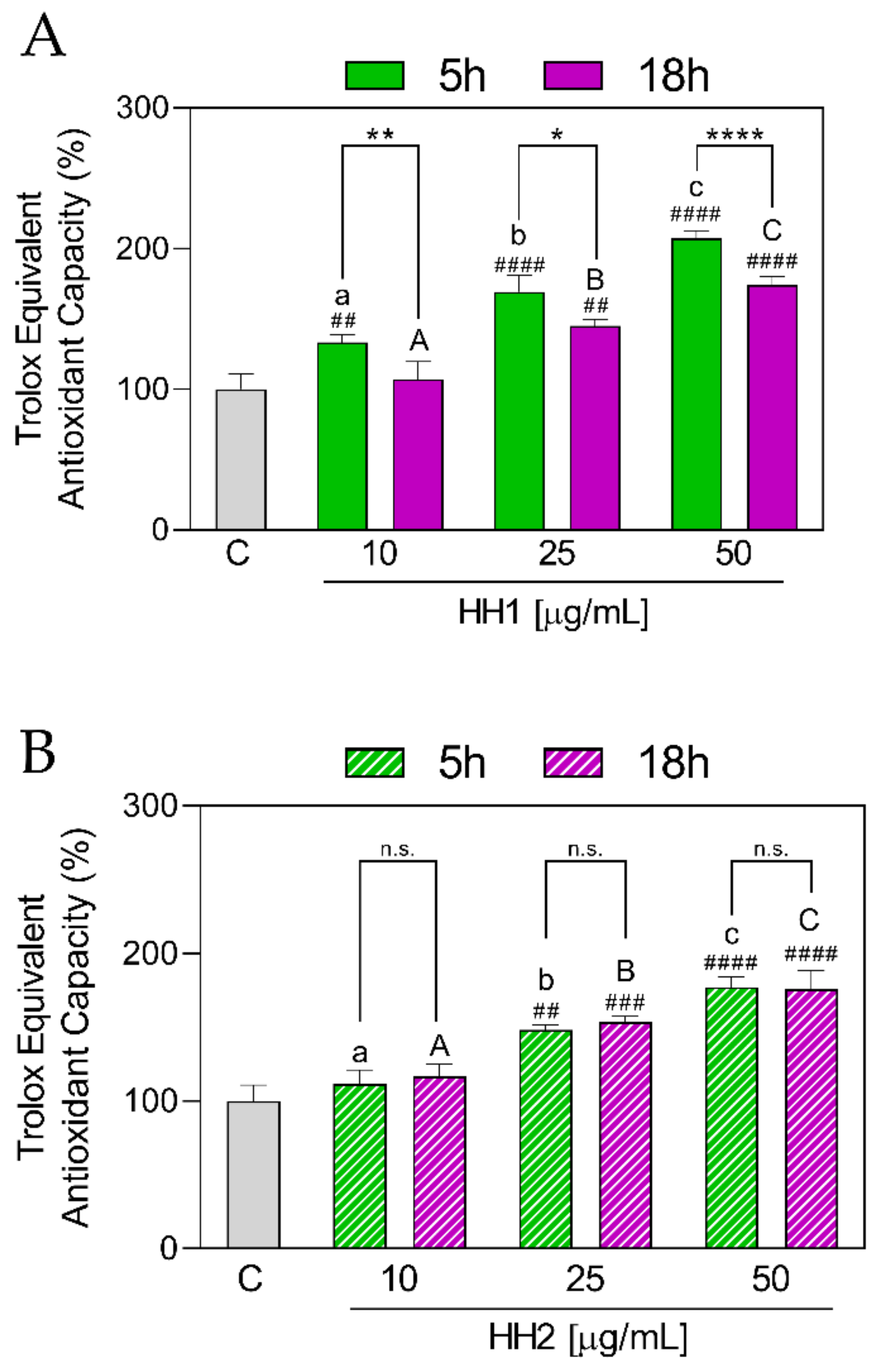
3.1.2. HHs Improve the Antioxidant Capacity by TEAC Radical Scavenging
3.1.3. HHs Improve the Ferric Reducing Antioxidant Power
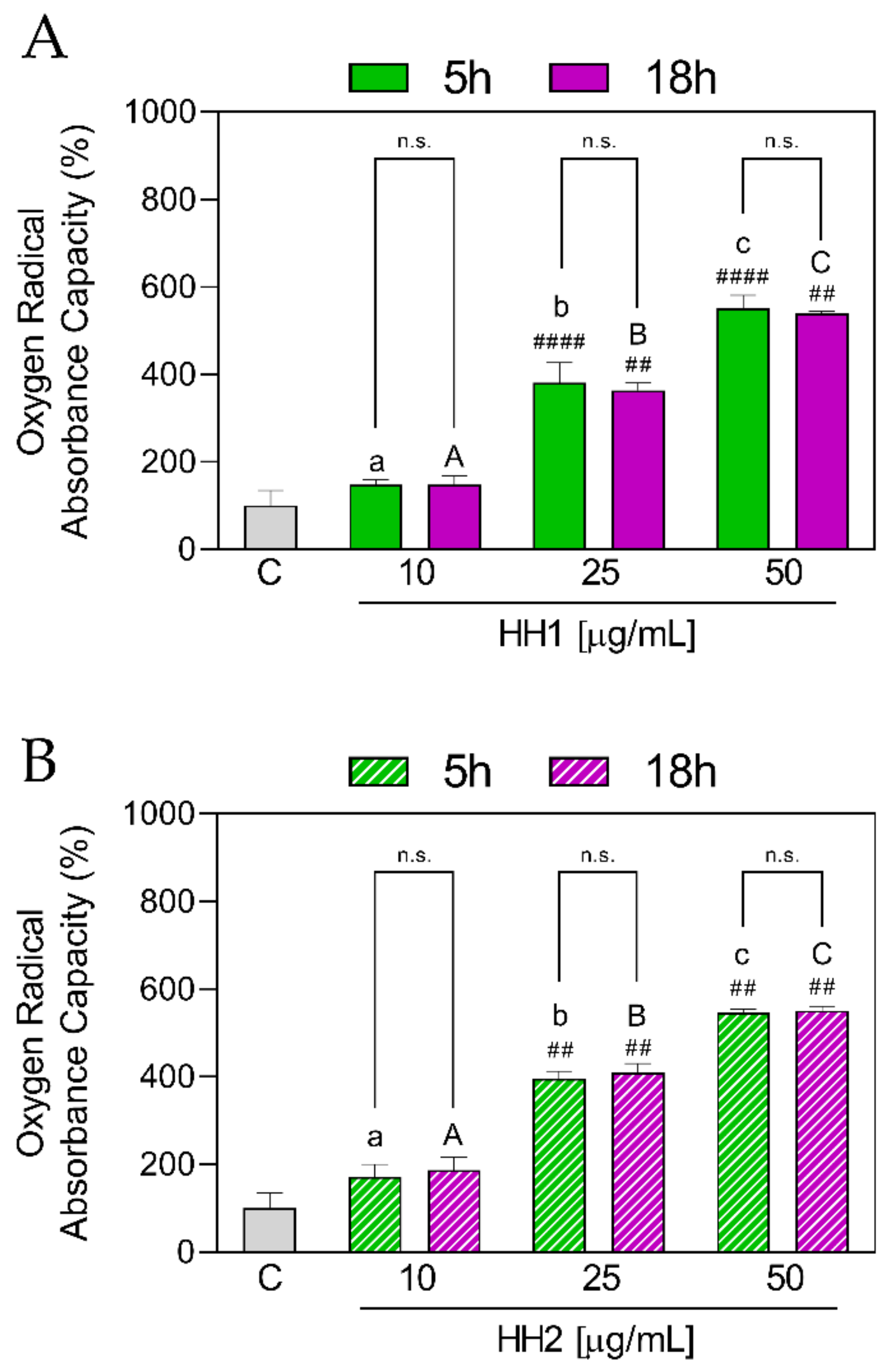
3.1.4. HHs Improve Antioxidant Capacity by AAPH Radical Scavenging
3.1.5. HH1 and HH2 Show Differences in the Antioxidant Activity
3.2. Analysis and Characterization of HHs Composition
3.3. The HHs Reduce the H2O2-Induced Oxidative Stress in Caco-2 Cells
3.4. Trans-Epithelial Transported HHs Exert Antioxidant Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ulug, S.K.; Jahandideh, F.; Wu, J. Novel technologies for the production of bioactive peptides. Trends Food Sci. Technol. 2021, 108, 27–39. [Google Scholar] [CrossRef]
- Cruz-Chamorro, I.; Santos-Sánchez, G.; Álvarez-López, A.I.; Pedroche, J.; Lardone, P.J.; Arnoldi, A.; Lammi, C.; Carrillo-Vico, A. Pleiotropic biological effects of Lupinus spp. protein hydrolysates. Trends Food Sci. Technol. 2023, 133, 244–266. [Google Scholar] [CrossRef]
- Santos-Sánchez, G.; Cruz-Chamorro, I.; Álvarez-Ríos, A.I.; Álvarez-Sánchez, N.; Rodríguez-Ortiz, B.; Álvarez-López, A.I.; Fernández-Pachón, M.-S.; Pedroche, J.; Millán, F.; Millan-Linares, M.d.C. Bioactive Peptides from Lupin (Lupinus angustifolius) Prevent the Early Stages of Atherosclerosis in Western Diet-Fed ApoE–/–Mice. J. Agric. Food Chem. 2022, 70, 8243–8253. [Google Scholar] [CrossRef] [PubMed]
- Santos-Sánchez, G.; Cruz-Chamorro, I.; Álvarez-Ríos, A.I.; Fernández-Santos, J.M.; Vázquez-Román, M.V.; Rodríguez-Ortiz, B.; Álvarez-Sánchez, N.; Álvarez-López, A.I.; Millán-Linares, M.d.C.; Millán, F. Lupinus angustifolius protein hydrolysates reduce abdominal adiposity and ameliorate metabolic associated fatty liver disease (MAFLD) in Western diet fed-ApoE−/− mice. Antioxidants 2021, 10, 1222. [Google Scholar] [CrossRef] [PubMed]
- Santos-Sánchez, G.; Cruz-Chamorro, I.; Bollati, C.; Bartolomei, M.; Pedroche, J.; Millán, F.; del Carmen Millán-Linares, M.; Capriotti, A.L.; Cerrato, A.; Laganà, A. A Lupinus angustifolius protein hydrolysate exerts hypocholesterolemic effects in Western diet-fed ApoE−/− mice through the modulation of LDLR and PCSK9 pathways. Food Funct. 2022, 13, 4158–4170. [Google Scholar] [CrossRef]
- Wong, F.-C.; Xiao, J.; Wang, S.; Ee, K.-Y.; Chai, T.-T. Advances on the antioxidant peptides from edible plant sources. Trends Food Sci. Technol. 2020, 99, 44–57. [Google Scholar] [CrossRef]
- Aluko, R.E. Structure and function of plant protein-derived antihypertensive peptides. Curr. Opin. Food Sci. 2015, 4, 44–50. [Google Scholar] [CrossRef]
- Guha, S.; Majumder, K. Structural-features of food-derived bioactive peptides with anti-inflammatory activity: A brief review. J. Food Biochem. 2019, 43, e12531. [Google Scholar] [CrossRef]
- Kardani, K.; Bolhassani, A. Antimicrobial/anticancer peptides: Bioactive molecules and therapeutic agents. Immunotherapy 2021, 13, 669–684. [Google Scholar] [CrossRef]
- Lü, J.M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell. Mol. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Zou, T.-B.; He, T.-P.; Li, H.-B.; Tang, H.-W.; Xia, E.-Q. The structure-activity relationship of the antioxidant peptides from natural proteins. Molecules 2016, 21, 72. [Google Scholar] [CrossRef]
- Wang, B.; Xie, N.; Li, B. Influence of peptide characteristics on their stability, intestinal transport, and in vitro bioavailability: A review. J. Food Biochem. 2019, 43, e12571. [Google Scholar] [CrossRef]
- House, J.D.; Neufeld, J.; Leson, G. Evaluating the quality of protein from hemp seed (Cannabis sativa L.) products through the use of the protein digestibility-corrected amino acid score method. J. Agric. Food Chem. 2010, 58, 11801–11807. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xiong, Y.L. Processing, nutrition, and functionality of hempseed protein: A review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 936–952. [Google Scholar] [CrossRef] [PubMed]
- El-Sohaimy, S.A.; Androsova, N.V.; Toshev, A.D.; El Enshasy, H.A. Nutritional Quality, Chemical, and Functional Characteristics of Hemp (Cannabis sativa ssp. Sativa) Protein Isolate. Plants 2022, 11, 2825. [Google Scholar] [CrossRef] [PubMed]
- Santos-Sánchez, G.; Álvarez-López, A.I.; Ponce-España, E.; Carrillo-Vico, A.; Bollati, C.; Bartolomei, M.; Lammi, C.; Cruz-Chamorro, I. Hempseed (Cannabis sativa) protein hydrolysates: A valuable source of bioactive peptides with pleiotropic health-promoting effects. Trends Food Sci. Technol. 2022, 127, 303–318. [Google Scholar] [CrossRef]
- Malomo, S.A.; Onuh, J.O.; Girgih, A.T.; Aluko, R.E. Structural and antihypertensive properties of enzymatic hemp seed protein hydrolysates. Nutrients 2015, 7, 7616–7632. [Google Scholar] [CrossRef]
- Girgih, A.T.; Udenigwe, C.C.; Li, H.; Adebiyi, A.P.; Aluko, R.E. Kinetics of enzyme inhibition and antihypertensive effects of hemp seed (Cannabis sativa L.) protein hydrolysates. J. Am. Oil Chem. Soc. 2011, 88, 1767–1774. [Google Scholar] [CrossRef]
- Orio, L.P.; Boschin, G.; Recca, T.; Morelli, C.F.; Ragona, L.; Francescato, P.; Arnoldi, A.; Speranza, G. New ACE-inhibitory peptides from hemp seed (Cannabis sativa L.) proteins. J. Agric. Food Chem. 2017, 65, 10482–10488. [Google Scholar] [CrossRef]
- Ren, Y.; Liang, K.; Jin, Y.; Zhang, M.; Chen, Y.; Wu, H.; Lai, F. Identification and characterization of two novel α-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- Cerrato, A.; Lammi, C.; Capriotti, A.L.; Bollati, C.; Cavaliere, C.; Montone, C.M.; Bartolomei, M.; Boschin, G.; Li, J.; Piovesana, S. Isolation and functional characterization of hemp seed protein-derived short-and medium-chain peptide mixtures with multifunctional properties for metabolic syndrome prevention. Food Res. Int. 2023, 163, 112219. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, C.; Aiello, G.; Arnoldi, A.; Lammi, C. Hempseed peptides exert hypocholesterolemic effects with a statin-like mechanism. J. Agric. Food Chem. 2017, 65, 8829–8838. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Chamorro, I.; Santos-Sánchez, G.; Bollati, C.; Bartolomei, M.; Li, J.; Arnoldi, A.; Lammi, C. Hempseed (Cannabis sativa) peptides WVSPLAGRT and IGFLIIWV exert anti-inflammatory activity in the LPS-stimulated human hepatic cell line. J. Agric. Food Chem. 2022, 70, 577–583. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Malomo, S.; Offengenden, M.; Wu, J.; Aluko, R.E. Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J. Funct. Foods 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Bollati, C.; Cruz-Chamorro, I.; Aiello, G.; Li, J.; Bartolomei, M.; Santos-Sánchez, G.; Ranaldi, G.; Ferruzza, S.; Sambuy, Y.; Arnoldi, A. Investigation of the intestinal trans-epithelial transport and antioxidant activity of two hempseed peptides WVSPLAGRT (H2) and IGFLIIWV (H3). Food Res. Int. 2022, 152, 110720. [Google Scholar] [CrossRef]
- Girgih, A.T.; Alashi, A.M.; He, R.; Malomo, S.A.; Raj, P.; Netticadan, T.; Aluko, R.E. A novel hemp seed meal protein hydrolysate reduces oxidative stress factors in spontaneously hypertensive rats. Nutrients 2014, 6, 5652–5666. [Google Scholar] [CrossRef]
- Gao, J.; Li, T.; Chen, D.; Gu, H.; Mao, X. Identification and molecular docking of antioxidant peptides from hemp seed protein hydrolysates. LWT 2021, 147, 111453. [Google Scholar] [CrossRef]
- Rodriguez-Martin, N.M.; Toscano, R.; Villanueva, A.; Pedroche, J.; Millan, F.; Montserrat-de la Paz, S.; Millan-Linares, M.C. Neuroprotective protein hydrolysates from hemp (Cannabis sativa L.) seeds. Food Funct. 2019, 10, 6732–6739. [Google Scholar] [CrossRef]
- Logarušić, M.; Slivac, I.; Radošević, K.; Bagović, M.; Redovniković, I.R.; Srček, V.G. Hempseed protein hydrolysates’ effects on the proliferation and induced oxidative stress in normal and cancer cell lines. Mol. Biol. Rep. 2019, 46, 6079–6085. [Google Scholar] [CrossRef]
- Mahbub, R.; Callcott, E.; Rao, S.; Ansari, O.; Waters, D.L.; Blanchard, C.L.; Santhakumar, A.B. The effect of selected hemp seed protein hydrolysates in modulating vascular function. Food Biosci. 2022, 45, 101504. [Google Scholar] [CrossRef]
- Wang, Q.; Xiong, Y.L. Zinc-binding behavior of hemp protein hydrolysates: Soluble versus insoluble zinc-peptide complexes. J. Funct. Foods 2018, 49, 105–112. [Google Scholar] [CrossRef]
- Lammi, C.; Aiello, G.; Bollati, C.; Li, J.; Bartolomei, M.; Ranaldi, G.; Ferruzza, S.; Fassi, E.M.A.; Grazioso, G.; Sambuy, Y. Trans-epithelial transport, metabolism, and biological activity assessment of the multi-target lupin peptide LILPKHSDAD (P5) and its metabolite LPKHSDAD (P5-Met). Nutrients 2021, 13, 863. [Google Scholar] [CrossRef] [PubMed]
- Aiello, G.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A.; Vistoli, G.; Lammi, C. Behavior of three hypocholesterolemic peptides from soy protein in an intestinal model based on differentiated Caco-2 cell. J. Funct. Foods 2018, 45, 363–370. [Google Scholar] [CrossRef]
- Karami, Z.; Akbari-Adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef]
- Ji, D.; Xu, M.; Udenigwe, C.C.; Agyei, D. Physicochemical characterisation, molecular docking, and drug-likeness evaluation of hypotensive peptides encrypted in flaxseed proteome. Curr. Res. Food Sci. 2020, 3, 41–50. [Google Scholar] [CrossRef]
- Clemente, A. Enzymatic protein hydrolysates in human nutrition. Trends Food Sci. Technol. 2000, 11, 254–262. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The role of oxidative stress in physiopathology and pharmacological treatment with pro-and antioxidant properties in chronic diseases. Oxidative Med. Cell. Longev. 2020, 2020, 2082145. [Google Scholar] [CrossRef]
- Teh, S.-S.; Bekhit, A.E.-D.A.; Carne, A.; Birch, J. Antioxidant and ACE-inhibitory activities of hemp (Cannabis sativa L.) protein hydrolysates produced by the proteases AFP, HT, Pro-G, actinidin and zingibain. Food Chem. 2016, 203, 199–206. [Google Scholar] [PubMed]
- Aguilar-Toalá, J.; Quintanar-Guerrero, D.; Liceaga, A.; Zambrano-Zaragoza, M. Encapsulation of bioactive peptides: A strategy to improve the stability, protect the nutraceutical bioactivity and support their food applications. RSC Adv. 2022, 12, 6449–6458. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Hong, H.; Wu, J.; Yan, X. Bioavailability of bioactive peptides derived from food proteins across the intestinal epithelial membrane: A review. Trends Food Sci. Technol. 2019, 86, 399–411. [Google Scholar] [CrossRef]
- Jao, C.-L.; Huang, S.-L.; Hsu, K.-C. Angiotensin I-converting enzyme inhibitory peptides: Inhibition mode, bioavailability, and antihypertensive effects. BioMedicine 2012, 2, 130–136. [Google Scholar] [CrossRef]
- Liu, D.; Guo, Y.; Ma, H. Production, bioactivities and bioavailability of bioactive peptides derived from walnut origin by-products: A review. Crit. Rev. Food Sci. Nutr. 2022, 1–16. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | EC Number | Family | Enzyme | HH1 | HH2 |
|---|---|---|---|---|---|
| Aspergillus niger | 4.2.2.10 | Polysaccharidases | Pectin lyase | ✓ | ✓ |
| 3.2.1.15 | Glycosidases | Polygalacturonase | ✓ | ✓ | |
| 3.2.1.4 | Cellulase | ✓ | ✓ | ||
| Aspergillus oryzae | 3.4.21.63 | Peptidase S8 | Oryzin | ✓ | |
| 3.4.11.1 | Peptidase M17 | Leucyl aminopeptidase | ✓ | ||
| Bacillus licheniformis | 3.4.21.62 | Peptidase S8 | Subtilisin | ✓ |
| Assay | Concentration | 5 h | p-Value | 18 h | p-Value | ||
|---|---|---|---|---|---|---|---|
| HH1 | HH2 | HH1 | HH2 | ||||
| DPPH | 0.5 mg/mL | 54.50 ± 3.43 | 83.80 ± 5.96 | 0.005 | 82.70 ± 0.96 | 83.10 ± 4.52 | >0.999 |
| 1.0 mg/mL | 40.10 ± 2.08 | 62.70 ± 2.06 | 0.008 | 58.80 ± 4.07 | 60.90 ± 3.16 | 0.961 | |
| 2.5 mg/mL | 68.00 ± 6.37 | 40.00 ± 1.22 | 0.047 | 41.00 ± 0.89 | 35.70 ± 1.25 | 0.088 | |
| TEAC | 10 µg/mL | 133 ± 5.70 | 111 ± 9.69 | 0.009 | 107 ± 12.30 | 117 ± 8.13 | 0.183 |
| 25 µg/mL | 169 ± 11.80 | 148 ± 3.50 | 0.021 | 145 ± 4.61 | 154 ± 3.74 | 0.044 | |
| 50 µg/mL | 205 ± 5.28 | 177 ± 6.93 | 0.0001 | 174 ± 5.56 | 175 ± 12.7 | >0.999 | |
| FRAP | 10 µg/mL | 235 ± 60.70 | 192 ± 93.00 | 0.785 | 196 ± 94.90 | 216 ± 86.60 | 0.999 |
| 25 µg/mL | 646 ± 36.80 | 589 ± 37.30 | 0.376 | 566 ± 73.40 | 575 ± 69.60 | >0.999 | |
| 50 µg/mL | 1578 ± 39.00 | 1397 ± 150.00 | 0.141 | 1186 ± 25.90 | 1392 ± 195.00 | 0.209 | |
| ORAC | 10 µg/mL | 148 ± 12.00 | 172 ± 27.60 | 0.100 | 148 ± 20.00 | 187 ± 28.70 | 0.001 |
| 25 µg/mL | 381 ± 45.50 | 396 ± 16.80 | 0.997 | 363 ± 17.20 | 409 ± 20.7 | 0.001 | |
| 50 µg/mL | 552 ± 28.90 | 547 ± 7.83 | >0.999 | 539 ± 4.70 | 551 ± 10.00 | 0.103 | |
| Protein | Sequence | m/z (Da) | Sample |
|---|---|---|---|
| HH1.5 | |||
| acyl-activating enzyme 12 | (H)AHGRADDTMNLGGIKVSSVE(I) | 686.43 | 1.5 |
| (Y)HAHGRADDT(M) | 327.32 | 1.5BL | |
| acyl-activating enzyme 8 | (D)GGHKPGSVGKPVGQEMAILDQ(N) | 702.54 | 1.5 |
| (G)HKPGSVGKPVGQEMAILDQ(N) | 664.56 | 1.5BL | |
| HH1.18 | |||
| RNA polymerase beta subunit | (L)GVPSRMNVGQIFECSLGLAGEL(L) | 759.61 | 1.18 |
| (D)MVFNPLGVPSRMNVGQIFECSL(G) | 813.24 | 1.18 | |
| (V)PSRMNVGQIFE(C) | 426.74 | 1.18BL | |
| ATP synthase CF1 alpha subunit (chloroplast) | (V)LMGDGLLIQEGSSVKATGRIAQIPVSEAFL(G) | 1034.28 | 1.18 |
| (A)TGRIAQIPVSEAF(L) | 463.69 | 1.18BL | |
| ATP synthase CF1 beta subunit (chloroplast) | (G)LMRGMDVIDTGAPLSVPVGGATLGRIF(N) | 914.98 | 1.18 |
| (P)LSVPVGGATLGRI(F) | 620.66 | 1.18BL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Sánchez, G.; Aiello, G.; Rivardo, F.; Bartolomei, M.; Bollati, C.; Arnoldi, A.; Cruz-Chamorro, I.; Lammi, C. Antioxidant Effect Assessment and Trans Epithelial Analysis of New Hempseed Protein Hydrolysates. Antioxidants 2023, 12, 1099. https://doi.org/10.3390/antiox12051099
Santos-Sánchez G, Aiello G, Rivardo F, Bartolomei M, Bollati C, Arnoldi A, Cruz-Chamorro I, Lammi C. Antioxidant Effect Assessment and Trans Epithelial Analysis of New Hempseed Protein Hydrolysates. Antioxidants. 2023; 12(5):1099. https://doi.org/10.3390/antiox12051099
Chicago/Turabian StyleSantos-Sánchez, Guillermo, Gilda Aiello, Fabrizio Rivardo, Martina Bartolomei, Carlotta Bollati, Anna Arnoldi, Ivan Cruz-Chamorro, and Carmen Lammi. 2023. "Antioxidant Effect Assessment and Trans Epithelial Analysis of New Hempseed Protein Hydrolysates" Antioxidants 12, no. 5: 1099. https://doi.org/10.3390/antiox12051099