
Early-Life Exposure to Commercial Formulation Containing Deltamethrin and Cypermethrin Insecticides Impacts Redox System and Induces Unexpected Regional Effects in Rat Offspring Brain

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Early-Life Treatment and Sample Collection
2.4. Sample Preparation
2.5. Oxidative Stress Study in the Brain
2.5.1. Superoxide Dismutase Determination
2.5.2. Catalase Activity Determination
2.5.3. Glutathione Determination
2.5.4. Lipid Peroxidation Determination
2.5.5. Protein Carbonyl Determination
2.6. Immunoblotting Analysis
2.7. Statistical Analysis
3. Results
3.1. Antioxidant Enzymes
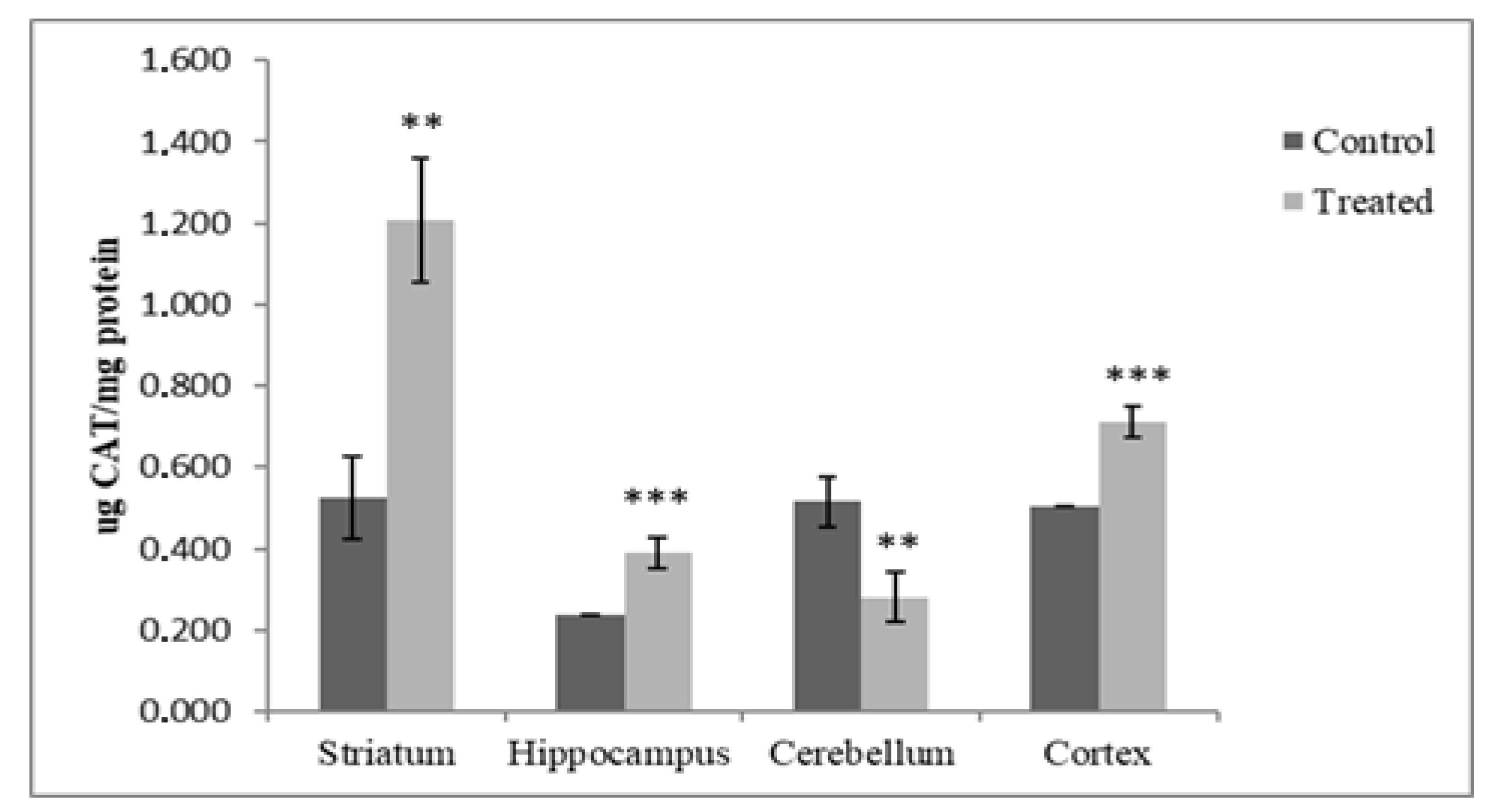
3.1.1. Catalase Analysis
3.1.2. SOD Analysis
3.2. GSH Content
3.3. Lipid Peroxidation
3.4. Protein Carbonyl
3.5. Western Blot Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Casida, J.E. Pyrethrum flowers and pyrethroid insecticides. Environ. Health Perspect. 1980, 34, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Soderlund, D.M. Molecular mechanisms of pyrethroid insecticide neurotoxicity: Recent advances. Arch. Toxicol. 2012, 86, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.A.; Wang, K.; Olson, J.R.; Bonner, M.R.; Hendy, O.; Abdel Rasoul, G.; Rohlman, D.S. The impact of repeated organophosphorus pesticide exposure on biomarkers and neurobehavioral outcomes among adolescent pesticide applicators. J. Toxicol. Environ. Health. 2017, 80 Pt A, 542–555. [Google Scholar] [CrossRef]
- Hocine, L.; Merzouk, H.; Merzouk, S.A.; Ghorzi, H.; Youbi, M.; Narce, M. The effects of alpha-cypermethrin exposure on biochemical and redox parameters in pregnant rats and their newborns. Pestic. Biochem. Physiol. 2016, 134, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Mekircha, F.; Chebab, S.; Gabbianelli, R.; Leghouchi, E. The possible ameliorative effect of Olea europaea L. oil against deltamethrin-induced oxidative stress and alterations of serum concentrations of thyroid and reproductive hormones in adult female rats. Ecotoxicol. Environ. Saf. 2018, 161, 374–382. [Google Scholar] [CrossRef]
- Nasuti, C.; Carloni, M.; Fedeli, D.; Gabbianelli, R.; Di Stefano, A.; Serafina, C.L.; Silva, I.; Domingues, V.; Ciccocioppo, R. Effects of early life permethrin exposure on spatial working memory and on monoamine levels in different brain areas of pre-senescent rats. Toxicology 2013, 303, 162–168. [Google Scholar] [CrossRef]
- Vadhana, M.S.D.; Arumugam, S.S.; Carloni, M.; Nasuti, C.; Gabbianelli, R. Early life permethrin treatment leads to long-term cardiotoxicity. Chemosphere 2013, 93, 1029–1034. [Google Scholar] [CrossRef]
- Kumar, A.; Sasmal, D.; Sharma, N. An insight into deltamethrin induced apoptotic calcium, p53 and oxidative stress signalling pathways. Toxicol. Environ. Health Sci. 2015, 7, 25–34. [Google Scholar] [CrossRef]
- Syed, F.; Chandravanshi, L.P.; Khanna, V.K.; Soni, I. Beta-cyfluthrin induced neurobehavioral impairments in adult rats. Chem. Biol. Interact. 2016, 243, 19–28. [Google Scholar] [CrossRef]
- Fedeli, D.; Montani, M.; Bordoni, L.; Galeazzi, R.; Nasuti, C.; Correia-Sa, L.; Domingues, V.F.; Jayant, M.; Brahmachari, V.; Massaccesi, L. In vivo and in silico studies to identify mechanisms associated with Nurr1 modulation following early life exposure to permethrin in rats. Neuroscience 2017, 340, 411–423. [Google Scholar] [CrossRef]
- Chambers-Richards, T.; Su, Y.; Chireh, B.; D’Arcy, C. Exposure to toxic occupations and their association with Parkinson’s disease: A systematic review with meta-analysis. Rev. Environ. Health 2021, 18, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Graaf, L.; Boulanger, M.; Bureau, M.; Bouvier, G.; Meryet-Figuiere, M.; Tual, S.; Baldi, I. Occupational pesticide exposure, cancer and chronic neurological disorders: A systematic review of epidemiological studies in greenspace workers. Environ. Res. 2022, 203, 111822. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, H.; Ghassemi-Barghi, N.; Malakshah, O.; Ashari, S. Pyrethroid exposure and neurotoxicity: A mechanistic approach. Arh. Hig. Rada Toksikol. 2019, 70, 74–89. [Google Scholar] [CrossRef] [PubMed]
- Nandipati, S.; Litvan, I. Environmental Exposures and Parkinson’s Disease. Int. J. Environ. Res. Public Health 2016, 13, 881. [Google Scholar] [CrossRef]
- Furlong, M.A.; Paul, K.C.; Cockburn, M.; Bronstein, J.; Keener, A.; Rosario, I.D.; Folle, A.D.; Ritz, B. Ambient Pyrethroid Pesticide Exposures in Adult Life and Depression in Older Residents of California’s Central Valley. Environ. Epidemiol. 2020, 4, e123. [Google Scholar] [CrossRef]
- Kannarkat, G.T.; Cook, D.A.; Lee, J.K.; Chang, J.; Chung, J.; Sandy, E.; Paul, K.C.; Ritz, B.; Bronstein, J.; Factor, S.A.; et al. Common Genetic Variant Association with Altered HLA Expression, Synergy with Pyrethroid Exposure, and Risk for Parkinson’s Disease: An Observational and Case-Control Study. NPJ Parkinsons Dis. 2015, 1, 15002. [Google Scholar] [CrossRef]
- Rice, D.; Barone, S. Critical periods of vulnerability for the developing nervous system: Evidence from humans and animal models. Environ. Health Perspect. 2000, 108 (Suppl. S3), 511–533. [Google Scholar] [CrossRef]
- Dayal, M.; Parmar, D.; Ali, M.; Dhawan, A.; Dwivedi, U.N.; Seth, P.K. Induction of rat brain cytochrome P450s (P450s) by deltamethrin: Regional specificity and correlation with neurobehavioral toxicity. Neurotox. Res. 2001, 3, 351–357. [Google Scholar] [CrossRef]
- Mani, V.M.; Asha, S.; Sadiq, A.M.M. Pyrethroid deltamethrin-induced developmental neurodegenerative cerebral injury and ameliorating effect of dietary glycoside naringin in male wistar rats. Biomed. Aging Pathol. 2014, 4, 1–8. [Google Scholar] [CrossRef]
- Agrawal, S.; Dixit, A.; Singh, A.; Tripathi, P.; Singh, D.; Patel, D.K.; Singh, M.P. Cyclosporine A and MnTMPyP alleviate α-synuclein expression and aggregation in cypermethrin-induced Parkinsonism. Mol. Neurobiol. 2015, 52, 1619–1628. [Google Scholar] [CrossRef]
- Vaccari, C.; El Dib, R.; Gomaa, H.; Lopes, L.C.; de Camargo, J.L. Paraquat and Parkinson’s disease: A systematic review and meta-analysis of observational studies. J. Toxicol. Environ. Health B Crit. Rev. 2019, 22, 172–202. [Google Scholar] [CrossRef] [PubMed]
- Van Maele-Fabry, G.; Hoet, P.; Vilain, F.; Lison, D. Occupational exposure to pesticides and Parkinson’s disease: A systematic review and meta-analysis of cohort studies. Environ. Int. 2012, 46, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.T.; Levy, L.S. Parkinson’s disease and pesticide exposure—A new assessment. Crit. Rev. Toxicol. 2013, 43, 515–534. [Google Scholar] [CrossRef] [PubMed]
- Nasuti, C.; Fattoretti, P.; Carloni, M.; Fedeli, D.; Ubaldi, M.; Ciccocioppo, R.; Gabbianelli, R. Neonatal exposure to permethrin pesticide causes lifelong fear and spatial learning deficits and alters hippocampal morphology of synapses. J. Neurodev. Disord. 2014, 6, 1–11. [Google Scholar] [CrossRef]
- Carloni, M.; Nasuti, C.; Fedeli, D.; Montani, M.; Vadhana, M.S.D.; Amici, A.; Gabbianelli, R. Early life permethrin exposure induces long-term brain changes in Nurr1, NF-kB and Nrf-2. Brain Res. 2013, 1515, 19–28. [Google Scholar] [CrossRef]
- Vadhana, M.S.D.; Carloni, M.; Nasuti, C.; Fedeli, D.; Gabbianelli, R. Early life permethrin insecticide treatment as origin of heart damage in adult rats. Exp. Gerontol. 2011, 46, 731–738. [Google Scholar] [CrossRef]
- Carloni, M.; Nasuti, C.; Fedeli, D.; Montani, M.; Amici, A.; Vadhana, D.M.S.; Gabbianelli, R. The impact of early life permethrin exposure on development of neurodegeneration in adulthood. Exp. Gerontol. 2012, 47, 60–66. [Google Scholar] [CrossRef]
- Bordoni, L.; Nasuti, C.; Di Stefano, A.; Marinelli, L.; Gabbianelli, R. Epigenetic memory of early-life parental perturbation: Dopamine decrease and DNA methylation changes in offspring. Oxid. Med. Cell. Longev. 2019, 2019, 1472623. [Google Scholar] [CrossRef]
- Bordoni, L.; Nasuti, C.; Fedeli, D.; Galeazzi, R.; Laudadio, E.; Massaccesi, L.; Lopez-Rodas, G.; Gabbianelli, R. Early impairment of epigenetic pattern in neurodegeneration: Additional mechanisms behind pyrethroid toxicity. Exp. Gerontol. 2019, 124, 110629. [Google Scholar] [CrossRef]
- Rossnerova, A.; Izzotti, A.; Pulliero, A.; Bast, A.; Rattan, S.I.S.; Rossner, P. The Molecular Mechanisms of Adaptive Response Related to Environmental Stress. Int. J. Mol. Sci. 2020, 21, 7053. [Google Scholar] [CrossRef]
- Pickering, A.M.; Staab, T.A.; Tower, J.; Sieburth, D.; Davies, K.J.A. A conserved role for the 20S proteasome and Nrf2 transcription factor in oxidative stress adaptation in mammals, Caenorhabditis elegans and Drosophila melanogaster. J. Exp. Biol. 2013, 216, 543–553. [Google Scholar] [CrossRef]
- Bordoni, L.; Fedeli, D.; Nasuti, C.; Capitani, M.; Fiorini, D.; Gabbianelli, R. Permethrin pesticide induces NURR1 up-regulation in dopaminergic cell line: Is the pro-oxidant effect involved in toxicant-neuronal damage? Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 201, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Rabhi, K.K.; Esancy, K.; Voisin, A.; Crespin, L.; Le Corre, J.; Tricoire-Leignel, H.; Anton, S.; Gadenne, C. Unexpected effects of low doses of a neonicotinoid insecticide on behavioral responses to sex pheromone in a pest insect. PLoS ONE 2014, 9, e114411. [Google Scholar] [CrossRef]
- Pickering, A.M.; Vojtovich, L.; Tower, J.; Davies, K.J.A. Oxidative stress adaptation with acute, chronic, and repeated stress. Free Radic. Biol. Med. 2013, 55, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Beghoul, A.; Kebieche, M.; Gasmi, S.; Chouit, Z.; Amiour, C.; Lahouel, A.; Soulimani, R. Impairment of mitochondrial integrity and redox status in brain regions during a low-dose long-term exposition of rats to pyrethrinoïds: The preventive effect of quercetin. Environ. Sci. Pollut. Res. 2017, 24, 19714–19722. [Google Scholar] [CrossRef]
- Arora, D.; Siddiqui, M.H.; Sharma, P.K.; Singh, S.P.; Tripathi, A.; Mandal, P.; Singh, U.S.; Singh, P.K.; Shukla, Y. Evaluation and physiological correlation of plasma proteomic fingerprints for deltamethrin-induced hepatotoxicity in Wistar rats. Life Sci. 2016, 160, 72–83. [Google Scholar] [CrossRef]
- Barlow, S.M.; Sullivan, F.M.; Lines, J. Risk assessment of the use of deltamethrin on bednets for the prevention of malaria. Food Chem. Toxicol. 2001, 39, 407–422. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Yadav, S.; Srivastava, V.; Kumar, R.; Singh, D.; Sethumadhavan, R.; Parmar, D. Imprinting of cerebral and hepatic cytochrome P450s in rat offsprings exposed prenatally to low doses of cypermethrin. Mol. Neurobiol. 2013, 48, 128–140. [Google Scholar] [CrossRef]
- Nasuti, C.; Gabbianelli, R.; Falcioni, M.L.; Di Stefano, A.; Sozio, P.; Cantalamessa, F. Dopaminergic system modulation, behavioral changes, and oxidative stress after neonatal administration of pyrethroids. Toxicology 2007, 229, 194–205. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Concetti, A.; Massei, P.; Rotilio, G.; Brunori, M.; Rachmilewitz, E.A. Superoxide dismutase in red blood cells: Method of assay and enzyme content in normal subjects and in patients with β-thalassemia (major and intermedia). J. Lab. Clin. Med. 1976, 87, 1057–1064. [Google Scholar]
- Misra, H.P.; Fridovich, I. The generation of superoxide radical during the autoxidation of hemoglobin. J. Biol. Chem. 1972, 247, 6960–6962. [Google Scholar] [CrossRef]
- Luck, H. Catalase. In Method of Enzymatic Analysis; Bergmayer, M.V., Ed.; Academic Press: New York, NY, USA, 1974; p. 885. [Google Scholar]
- Butler, R.N.; Butler, W.J.; Moraby, Z.; Fettmann, M.J.; Khoo, K.K.; Roberts-Thomson, I.C. Glutathione concentrations and glutathione S-transferase activity in human colonic neoplasms. J. Gastroenterol. Hepatol. 1994, 9, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Shibata, M.; Niki, E. Estimation of lipid peroxidation of live cells using a fluorescent probe, diphenyl-1-pyrenylphosphine. Free Radic. Biol. Med. 2001, 31, 164–174. [Google Scholar] [CrossRef]
- Soglia, F.; Petracci, M.; Ertbjerg, P. Novel DNPH-based method for determination of protein carbonylation in muscle and meat. Food Chem. 2016, 197, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar] [PubMed]
- Sies, H. Oxidative eustress: On constant alert for redox homeostasis. Redox Biol. 2021, 41, 101867. [Google Scholar] [CrossRef]
- Miao, L.; St Clair, D.K. Regulation of superoxide dismutase genes: Implications in disease. Free Rad. Biol Med. 2009, 47, 344–356. [Google Scholar] [CrossRef]
- Hermes-Lima, M.; Storey, J.M.; Storey, K.B. Antioxidant defenses and animal adaptation to oxygen availability during environmental stress. In Cell and Molecular Response to Stress; Storey, K.B., Storey, J.M., Eds.; Elsevier Press: Amsterdam, The Netherlands, 2001; Volume 2, pp. 263–287. [Google Scholar]
- Pamok, S.; Aengwanich, W.; Komutrin, T. Adaptation to oxidative stress and impact of chronic oxidative stress on immunity in heat-stressed broilers. J. Therm. Biol. 2009, 34, 353–357. [Google Scholar] [CrossRef]
- Haddad, J.J.; Harb, H.L. L-gamma-Glutamyl-L-cysteinyl-glycine (glutathione; GSH) and GSH-related enzymes in the regulation of pro- and anti-inflammatory cytokines: A signaling transcriptional scenario for redox(y) immunologic sensor(s)? Mol. Immunol. 2005, 42, 987–1014. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Aspects Med. 2009, 30, 1–12. [Google Scholar] [CrossRef]
- Deneke, S.M.; Fanburg, B.L. Regulation of cellular glutathione. Am. J. Physiol. Cell. Mol. Physiol. 1989, 257, L163–L173. [Google Scholar] [CrossRef]
- Halliwell, B. Antioxidant defence mechanisms: From the beginning to the end (of the beginning). Free Radic. Res. 1999, 31, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Limón-Pacheco, J.; Gonsebatt, M.E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat. Res. Toxicol. Environ. Mutagen. 2009, 674, 137–147. [Google Scholar] [CrossRef]
- Kehm, R.; Baldensperger, T.; Raupbach, J.; Höhn, A. Protein oxidation—Formation mechanisms, detection and relevance as biomarkers in human diseases. Redox Biol. 2021, 42, 101901. [Google Scholar] [CrossRef]
- Gonos, E.S.; Kapetanou, M.; Sereikaite, J.; Bartosz, G.; Naparło, K.; Grzesik, M.; Sadowska-Bartosz, I. Origin and pathophysiology of protein carbonylation, nitration and chlorination in age-related brain diseases and aging. Aging 2018, 10, 868–901. [Google Scholar] [CrossRef]
- Suzuki, Y.J.; Carini, M.; Butterfield, D.A. Protein carbonylation. Antioxid. Redox Signal. 2010, 12, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Chondrogianni, N.; Tzavelas, C.; Pemberton, A.J.; Nezis, I.P.; Rivett, A.J.; Gonos, E.S. Overexpression of Proteasome β5 Assembled Subunit Increases the Amount of Proteasome and Confers Ameliorated Response to Oxidative Stress and Higher Survival Rates. J. Biol. Chem. 2005, 12, 11840–11850. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A. The ubiquitin–proteasome pathway: On protein death and cell life. EMBO J. 1998, 24, 7151–7160. [Google Scholar] [CrossRef]
- De Martino, G.N.; Slaughter, C.A. The proteasome, a novel protease regulated by multiple mechanisms. J. Biol. Chem. 1999, 32, 22123–22126. [Google Scholar] [CrossRef]
- Burré, J.; Sharma, M.; Südhof, T.C. Cell Biology and Pathophysiology of α-Synuclein. Cold Spring Harb Perspect Med. 2018, 8, a024091. [Google Scholar] [CrossRef] [PubMed]
- Henderson, M.X.; Trojanowski, J.Q.; Lee, V.M. α-Synuclein pathology in Parkinson’s disease and related α-synucleinopathies. Neurosci Lett. 2019, 14, 134316. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Courte, J.; Bousset, L.; Von Boxberg, Y.; Villard, C.; Melki, R.; Peyrin, J.-M. The expression level of alpha-synuclein in different neuronal populations is the primary determinant of its prion-like seeding. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Sokratian, A.; Ziaee, J.; Kelly, K.; Chang, A.; Bryant, N.; Wang, S.; Xu, E.; Li, J.Y.; Wang, S.H.; Ervin, J.; et al. Heterogeneity in α-synuclein fibril activity correlates to disease phenotypes in Lewy body dementia. Acta Neuropathol. 2021, 141, 547–564. [Google Scholar] [CrossRef]
- Kou, J.; Gillette, J.S.; Bloomquist, J.R. Neurotoxicity in murine striatal dopaminergic pathways following co-application of permethrin, chlorpyrifos, and MPTP. Pestic. Biochem. Physiol. 2006, 85, 68–75. [Google Scholar] [CrossRef]
- Mitra, S.; Chakrabarti, N.; Bhattacharyya, A. Differential regional expression patterns of α-synuclein, TNF-α, and IL-1β; and variable status of dopaminergic neurotoxicity in mouse brain after Paraquat treatment. J. Neuroinflammation 2011, 8, 1–22. [Google Scholar] [CrossRef]
- Manning-Bog, A.B.; McCormack, A.L.; Li, J.; Uversky, V.N.; Fink, A.L.; Di Monte, D.A. The herbicide paraquat causes up-regulation and aggregation of α-synuclein in mice: Paraquat and α-synuclein. J. Biol. Chem. 2002, 277, 1641–1644. [Google Scholar] [CrossRef]
- Hashimoto, M.; Hsu, L.J.; Rockenstein, E.; Takenouchi, T.; Mallory, M.; Masliah, E. α-Synuclein protects against oxidative stress via inactivation of the c-Jun N-terminal kinase stress-signaling pathway in neuronal cells. J. Biol. Chem. 2002, 277, 11465–11472. [Google Scholar] [CrossRef]
- Skibinski, G.; Hwang, V.; Ando, D.M.; Daub, A.; Lee, A.K.; Ravisankar, A.; Modan, S.; Finucane, M.M.; Shaby, B.A.; Finkbeiner, S. Nrf2 mitigates LRRK2-and α-synuclein–induced neurodegeneration by modulating proteostasis. Proc. Natl. Acad. Sci. USA 2017, 114, 1165–1170. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mekircha, F.; Fedeli, D.; Nasuti, C.; Kecies, H.; Gabbianelli, R.; Bordoni, L. Early-Life Exposure to Commercial Formulation Containing Deltamethrin and Cypermethrin Insecticides Impacts Redox System and Induces Unexpected Regional Effects in Rat Offspring Brain. Antioxidants 2023, 12, 1047. https://doi.org/10.3390/antiox12051047
Mekircha F, Fedeli D, Nasuti C, Kecies H, Gabbianelli R, Bordoni L. Early-Life Exposure to Commercial Formulation Containing Deltamethrin and Cypermethrin Insecticides Impacts Redox System and Induces Unexpected Regional Effects in Rat Offspring Brain. Antioxidants. 2023; 12(5):1047. https://doi.org/10.3390/antiox12051047
Chicago/Turabian StyleMekircha, Fatiha, Donatella Fedeli, Cinzia Nasuti, Hadjer Kecies, Rosita Gabbianelli, and Laura Bordoni. 2023. "Early-Life Exposure to Commercial Formulation Containing Deltamethrin and Cypermethrin Insecticides Impacts Redox System and Induces Unexpected Regional Effects in Rat Offspring Brain" Antioxidants 12, no. 5: 1047. https://doi.org/10.3390/antiox12051047