
Radical Scavenging and Cellular Antioxidant Activity of the Cocoa Shell Phenolic Compounds after Simulated Digestion

 , ,
, ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. CSF and CSE Processing
2.3. CSF and CSE In Vitro Gastrointestinal Digestion
2.4. Phytochemical Profile Analysis by HPLC-PDA-ESI/MSn
2.5. Extraction of Free and Bound Phenolic Compounds
2.6. Assessment of the Total Phenolic Content
2.7. Evaluation of the In Vitro Antioxidant Capacity
2.7.1. ABTS Antioxidant Capacity
2.7.2. Ferric Reducing Antioxidant Power (FRAP)
2.8. Evaluation of the ROS and RNS Scavenging Capacity
2.8.1. Superoxide Anion Radical (O2•−) Scavenging Capacity
2.8.2. Hydrogen Peroxide (H2O2) Scavenging Capacity
2.8.3. Nitric Oxide (NO) Scavenging Capacity
2.8.4. Peroxynitrite (ONOO−) Scavenging Capacity
2.9. Antioxidant Activity in Cell Culture-Based Experiments
2.9.1. Cell Viability
2.9.2. Intracellular Reactive Oxygen Species (ROS) Measurement
2.9.3. Cell Lysates
2.9.4. Reduced Glutathione (GSH)
2.9.5. Thiol Groups
2.9.6. Catalase (CAT) and Superoxide Dismutase (SOD) Activity
2.10. Statistical Analysis
3. Results and Discussion
3.1. The Cocoa Shell Contained a High Content of Phenolic Compounds and Methylxanthines Which Were Released during Digestion
3.2. Digestion Affected Total Phenolic Content and Antioxidant Capacity Depending on the Cocoa Shell Matrix
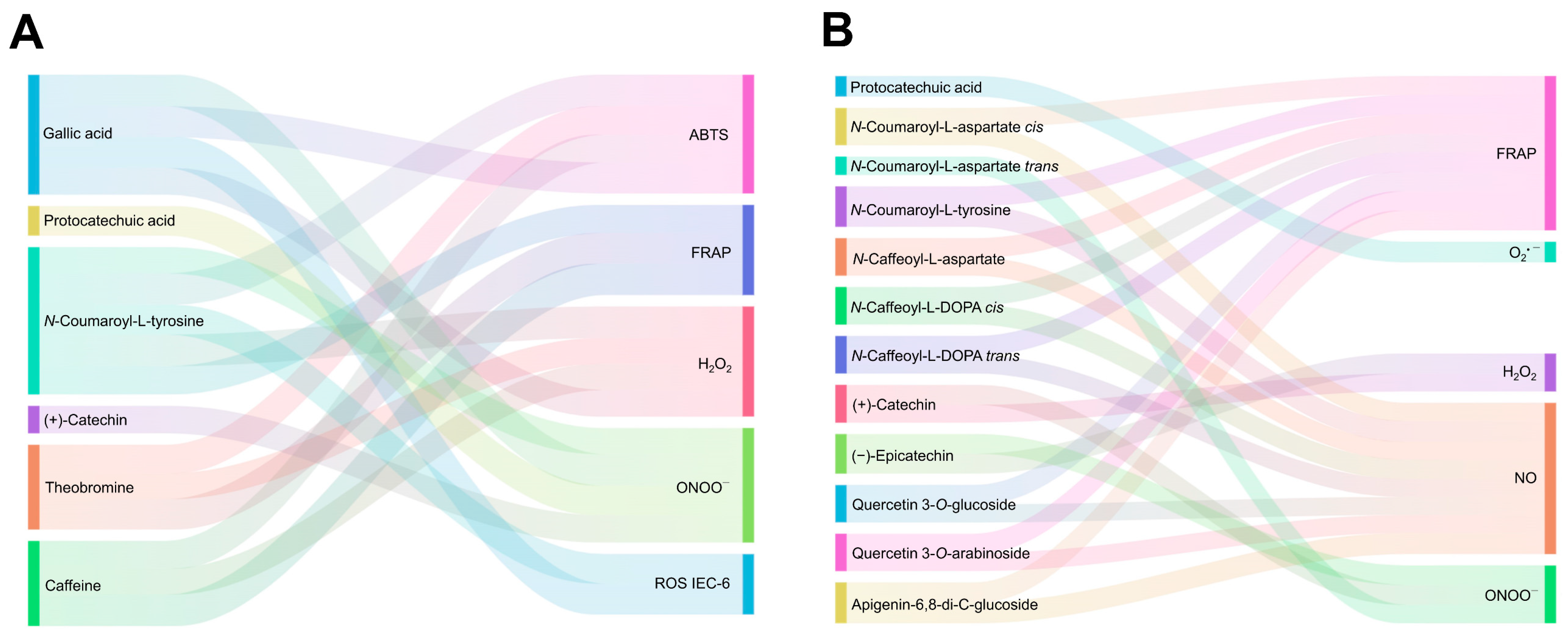
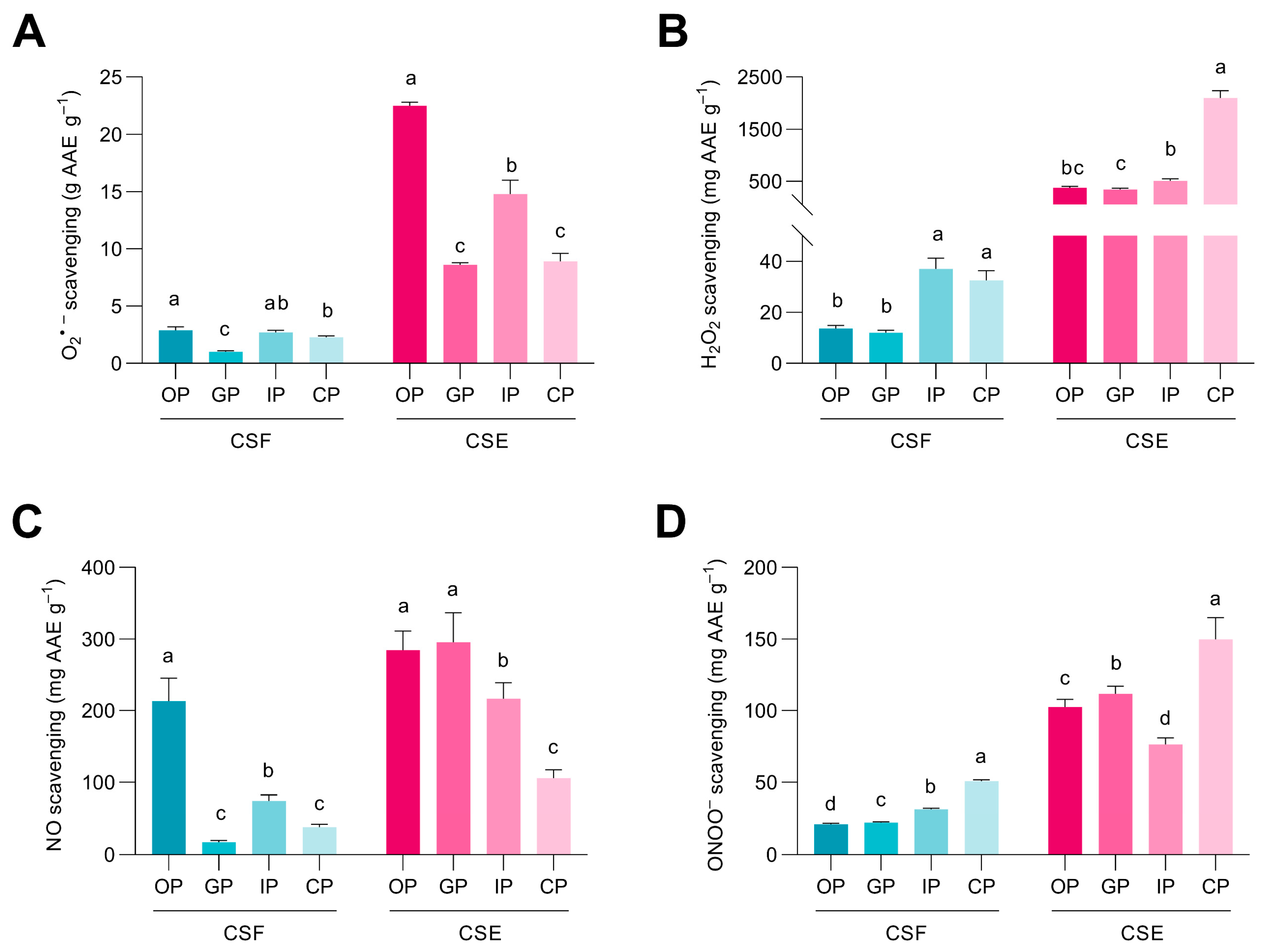
3.3. Phenolic Compounds from the Cocoa Shell May Retain Their Free Radicals Scavenge after Digestion
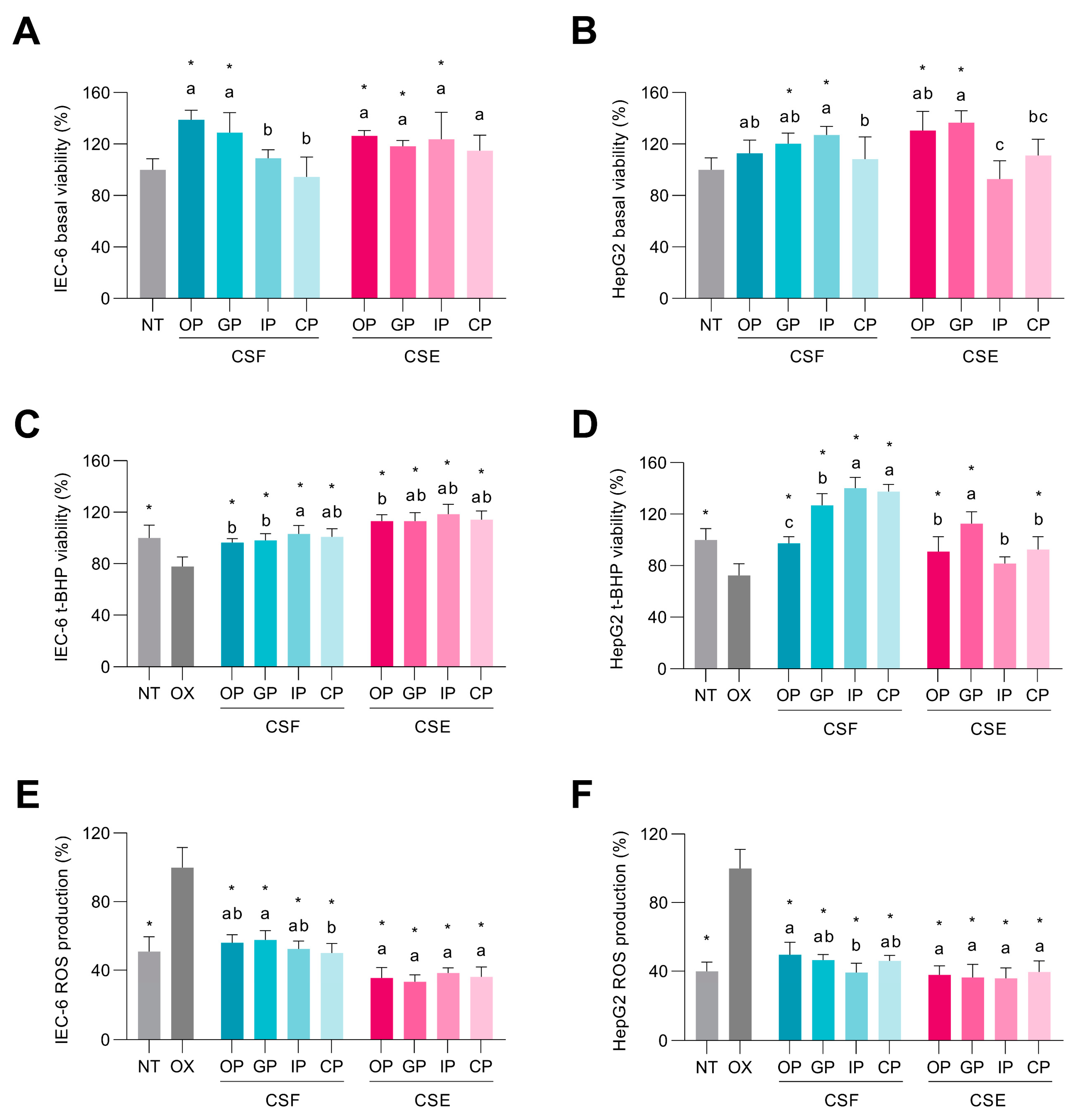
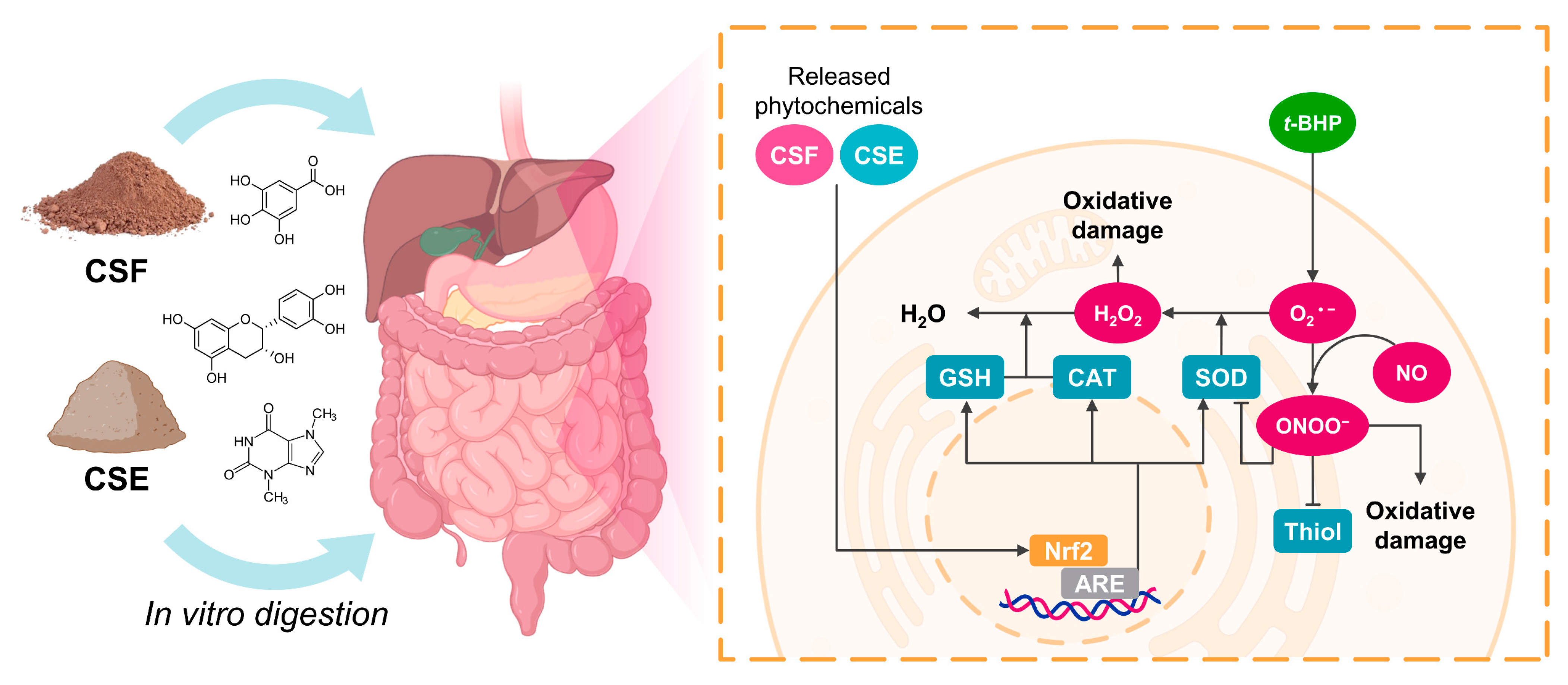
3.4. In Vitro Simulated Digestion Maintained the Cocoa Shell’s Capacity to Decrease ROS Production in Intestinal and Hepatic Cells
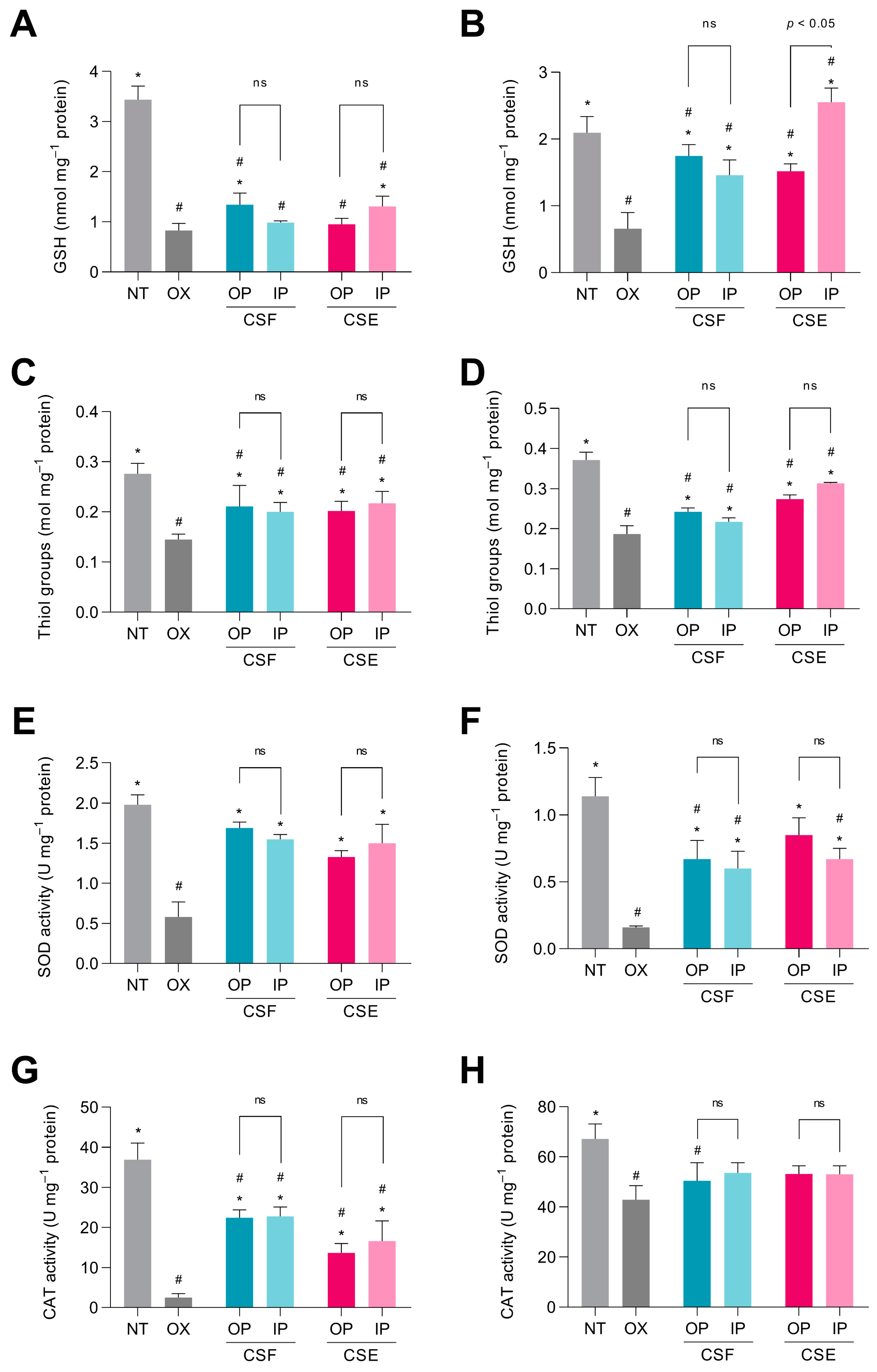
3.5. The Digested Cocoa Shell Activated the Cellular Antioxidant Defense System in Intestinal and Hepatic Cells
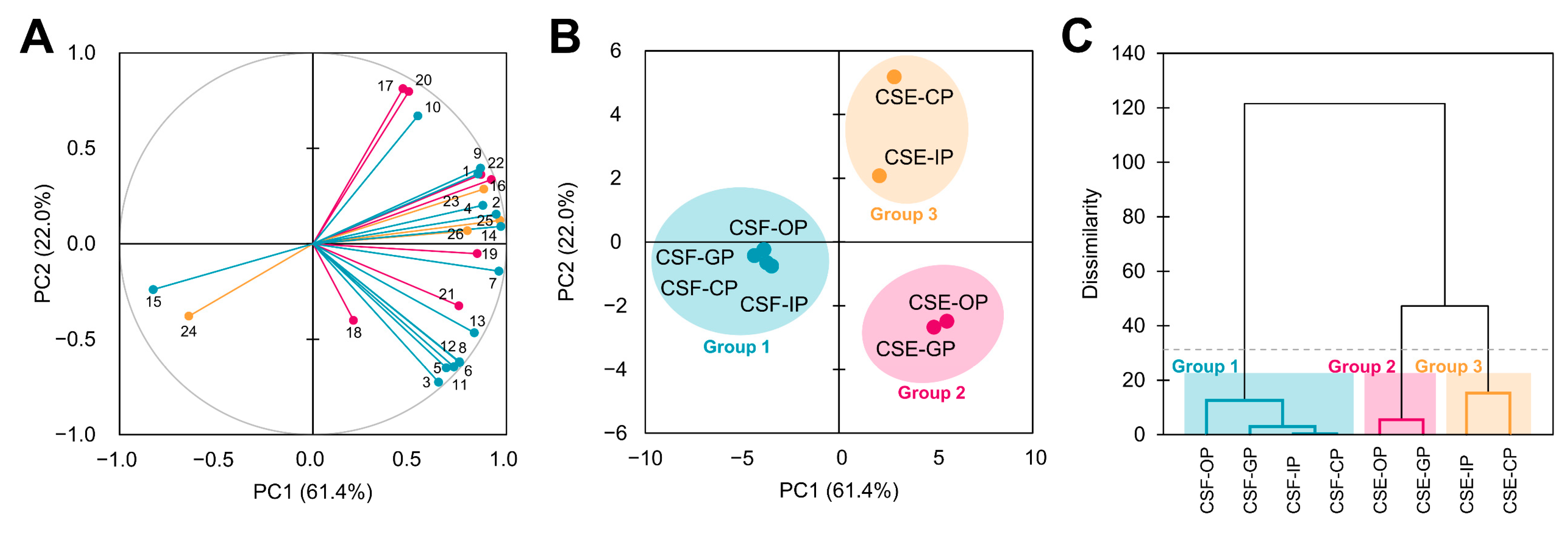
3.6. The Antioxidant and Radical Scavenging Capacities of the Cocoa Shell Are Influence by the Phytochemical Composition, Digestion, and Matrix
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Micheli, F.; Guiltinan, M.; Gramacho, K.P.; Wilkinson, M.J.; de Figueira, A.V.O.; de Cascardo, J.C.M.; Maximova, S.; Lanaud, C. Functional Genomics of Cacao. Adv. Bot. Res. 2010, 55, 119–177. [Google Scholar] [CrossRef]
- Ramón, V.; Hernández, H.E.; Polania, P.; Suárez, J.C. Spatial Distribution of Cocoa Quality: Relationship between Physicochemical, Functional and Sensory Attributes of Clones from Southern Colombia. Agronomy 2022, 13, 15. [Google Scholar] [CrossRef]
- Okiyama, D.C.G.; Navarro, S.L.B.; Rodrigues, C.E.C. Cocoa shell and its compounds: Applications in the food industry. Trends Food Sci. Technol. 2017, 63, 103–112. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Cañas, S.; Braojos, C.; Cano-Muñoz, P.; Martín-Cabrejas, M.A. Cocoa Shell. In Molecular Mechanisms of Functional Food; Wiley: Hoboken, NJ, USA, 2022; pp. 485–519. [Google Scholar]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The Role of Oxidative Stress in Physiopathology and Pharmacological Treatment with Pro- And Antioxidant Properties in Chronic Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 2082145. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Arancibia-Hernández, Y.L.; Hernández-Cruz, E.Y.; Pedraza-Chaverri, J. RONS and Oxidative Stress: An Overview of Basic Concepts. Oxygen 2022, 2, 437–478. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 283. [Google Scholar] [CrossRef]
- Poljsak, B.; Kovač, V.; Milisav, I. Antioxidants, food processing and health. Antioxidants 2021, 10, 433. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Cañas, S.; Taladrid, D.; Bartolomé, B.; Aguilera, Y.; Martin-Cabrejas, M.A. Extraction of phenolic compounds from cocoa shell: Modeling using response surface methodology and artificial neural networks. Sep. Purif. Technol. 2021, 270, 118779. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Aguilera, Y.; Martin-Cabrejas, M.A.; Gonzalez de Mejia, E. Phytochemicals from the Cocoa Shell Modulate Mitochondrial Function, Lipid and Glucose Metabolism in Hepatocytes via Activation of FGF21/ERK, AKT, and mTOR Pathways. Antioxidants 2022, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; de Mejia, E.G. Cocoa Shell Aqueous Phenolic Extract Preserves Mitochondrial Function and Insulin Sensitivity by Attenuating Inflammation between Macrophages and Adipocytes In vitro. Mol. Nutr. Food Res. 2019, 63, 1801413. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, P.; Ragusky, K.; Phuthong, S.; Ruvira, S.; Ramiro-Cortijo, D.; Cañas, S.; Rebollo-Hernanz, M.; Morales, M.D.; López de Pablo, Á.L.; Martín-Cabrejas, M.A.; et al. Vasoactive Properties of a Cocoa Shell Extract: Mechanism of Action and Effect on Endothelial Dysfunction in Aged Rats. Antioxidants 2022, 11, 429. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; Gonzalez de Mejia, E. Relationship of the phytochemicals from coffee and cocoa by-products with their potential to modulate biomarkers of metabolic syndrome in vitro. Antioxidants 2019, 8, 279. [Google Scholar] [CrossRef]
- Saura-Calixto, F.; Serrano, J.; Goñi, I. Intake and bioaccessibility of total polyphenols in a whole diet. Food Chem. 2007, 101, 492–501. [Google Scholar] [CrossRef]
- Cañas, S.; Rebollo-Hernanz, M.; Braojos, C.; Benítez, V.; Ferreras-Charro, R.; Dueñas, M.; Aguilera, Y.; Martín-Cabrejas, M.A. Gastrointestinal fate of phenolic compounds and amino derivatives from the cocoa shell: An in vitro and in silico approach. Food Res. Int. 2022, 162, 112117. [Google Scholar] [CrossRef]
- Liu, R.H.; Finley, J. Potential cell culture models for antioxidant research. J. Agric. Food Chem. 2005, 53, 4311–4314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Y.; Zhang, X.; Lu, Y.; Chen, H. New insights in intestinal oxidative stress damage and the health intervention effects of nutrients: A review. J. Funct. Foods 2020, 75, 104248. [Google Scholar] [CrossRef]
- Sadasivam, N.; Kim, Y.J.; Radhakrishnan, K.; Kim, D.K. Oxidative Stress, Genomic Integrity, and Liver Diseases. Molecules 2022, 27, 3159. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Bringe, N.A.; Gonzalez de Mejia, E. Selected Soybean Varieties Regulate Hepatic LDL-Cholesterol Homeostasis Depending on Their Glycinin:β-Conglycinin Ratio. Antioxidants 2022, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Benítez, V.; Rebollo-Hernanz, M.; Aguilera, Y.; Bejerano, S.; Cañas, S.; Martín-Cabrejas, M.A. Extruded coffee parchment shows enhanced antioxidant, hypoglycaemic, and hypolipidemic properties by releasing phenolic compounds from the fibre matrix. Food Funct. 2021, 12, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Fernández-Gómez, B.; Herrero, M.; Aguilera, Y.; Martín-Cabrejas, M.A.; Uribarri, J.; Del Castillo, M.D. Inhibition of the Maillard reaction by phytochemicals composing an aqueous coffee silverskin extract via a mixed mechanism of action. Foods 2019, 8, 438. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Aguilera, Y.; Herrera, T.; Cayuelas, L.T.; Dueñas, M.; Rodríguez-Rodríguez, P.; Ramiro-Cortijo, D.; Arribas, S.M.; Martín-Cabrejas, M.A. Bioavailability of melatonin from lentil sprouts and its role in the plasmatic antioxidant status in rats. Foods 2020, 9, 330. [Google Scholar] [CrossRef]
- Panzella, L.; Cerruti, P.; Ambrogi, V.; Agustin-Salazar, S.; D’Errico, G.; Carfagna, C.; Goya, L.; Ramos, S.; Martín, M.A.; Napolitano, A.; et al. A Superior All-Natural Antioxidant Biomaterial from Spent Coffee Grounds for Polymer Stabilization, Cell Protection, and Food Lipid Preservation. ACS Sustain. Chem. Eng. 2016, 4, 1169–1179. [Google Scholar] [CrossRef]
- Grancieri, M.; Martino, H.S.D.; Gonzalez de Mejia, E. Digested total protein and protein fractions from chia seed (Salvia hispanica L.) had high scavenging capacity and inhibited 5-LOX, COX-1-2, and iNOS enzymes. Food Chem. 2019, 289, 204–214. [Google Scholar] [CrossRef]
- Robinson, K.M.; Beckman, J.S. Synthesis of peroxynitrite from nitrite and hydrogen peroxide. Methods Enzymol. 2005, 396, 207–214. [Google Scholar]
- Patil, P.S.; Tatke, P.A.; Gabhe, S.Y. In vitro Antioxidant and Free Radical Scavenging Activity of Extracts of Rosa damascena Flower Petals. Am. J. Phytomed. Clin. Ther. 2015, 3, 589–601. [Google Scholar]
- Hissin, P.J.; Hilf, R. A fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Morgan, P.E.; Davies, M.J. Quantification of protein modification by oxidants. Free Radic. Biol. Med. 2009, 46, 965–988. [Google Scholar] [CrossRef] [PubMed]
- Rebollo-Hernanz, M.; Kusumah, J.; Bringe, N.A.; Shen, Y.; de Mejia, E.G. Peptide release, radical scavenging capacity, and antioxidant responses in intestinal cells are determined by soybean variety and gastrointestinal digestion under simulated conditions. Food Chem. 2023, 405, 134929. [Google Scholar] [CrossRef]
- Oñatibia-Astibia, A.; Franco, R.; Martínez-Pinilla, E. Health benefits of methylxanthines in neurodegenerative diseases. Mol. Nutr. Food Res. 2017, 61, 1600670. [Google Scholar] [CrossRef] [PubMed]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of In vitro Digestion on Composition, Bioaccessibility and Antioxidant Activity of Food Polyphenols-A Non-Systematic Review. Nutrients 2020, 12, 1401. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Reginio, F.C.; Thuengtung, S.; Ogawa, Y. Changes in bioactive compounds and antioxidant activity of plant-based foods by gastrointestinal digestion: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 4684–4705. [Google Scholar] [CrossRef]
- Cañas, S.; Rebollo-Hernanz, M.; Braojos, C.; Benítez, V.; Ferreras-Charro, R.; Dueñas, M.; Aguilera, Y.; Martín-Cabrejas, M.A. Understanding the Gastrointestinal Behavior of the Coffee Pulp Phenolic Compounds under Simulated Conditions. Antioxidants 2022, 11, 1818. [Google Scholar] [CrossRef]
- La Mantia, A.; Ianni, F.; Schoubben, A.; Cespi, M.; Lisjak, K.; Guarnaccia, D.; Sardella, R.; Blasi, P. Effect of Cocoa Roasting on Chocolate Polyphenols Evolution. Antioxidants 2023, 12, 469. [Google Scholar] [CrossRef]
- Platzer, M.; Kiese, S.; Herfellner, T.; Schweiggert-Weisz, U.; Eisner, P. How does the phenol structure influence the results of the folin-ciocalteu assay? Antioxidants 2021, 10, 811. [Google Scholar] [CrossRef]
- Platzer, M.; Kiese, S.; Herfellner, T.; Schweiggert-Weisz, U.; Miesbauer, O.; Eisner, P. Common trends and differences in antioxidant activity analysis of phenolic substances using single electron transfer based assays. Molecules 2021, 26, 1244. [Google Scholar] [CrossRef]
- de Mudenuti, N.V.R.; de Camargo, A.C.; Shahidi, F.; Madeira, T.B.; Hirooka, E.Y.; Grossmann, M.V.E. Soluble and insoluble-bound fractions of phenolics and alkaloids and their antioxidant activities in raw and traditional chocolate: A comparative study. J. Funct. Foods 2018, 50, 164–171. [Google Scholar] [CrossRef]
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The Versatility of Antioxidant Assays in Food Science and Safety—Chemistry, Applications, Strengths, and Limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef] [PubMed]
- Hayyan, M.; Hashim, M.A.; Alnashef, I.M. Superoxide Ion: Generation and Chemical Implications. Chem. Rev. 2016, 116, 3029–3085. [Google Scholar] [CrossRef]
- René, A.; Abasq, M.L.; Hauchard, D.; Hapiot, P. How do phenolic compounds react toward superoxide ion? A simple electrochemical method for evaluating antioxidant capacity. Anal. Chem. 2010, 82, 8703–8710. [Google Scholar] [CrossRef] [PubMed]
- Bendary, E.; Francis, R.R.; Ali, H.M.G.; Sarwat, M.I.; El Hady, S. Antioxidant and structure–activity relationships (SARs) of some phenolic and anilines compounds. Ann. Agric. Sci. 2013, 58, 173–181. [Google Scholar] [CrossRef]
- Martemucci, G.; Costagliola, C.; Mariano, M.; D’andrea, L.; Napolitano, P.; D’Alessandro, A.G. Free Radical Properties, Source and Targets, Antioxidant Consumption and Health. Oxygen 2022, 2, 48–78. [Google Scholar] [CrossRef]
- Rodrigues, E.; Mariutti, L.R.B.; Mercadante, A.Z. Carotenoids and phenolic compounds from Solanum sessiliflorum, an unexploited amazonian fruit, and their scavenging capacities against reactive oxygen and nitrogen species. J. Agric. Food Chem. 2013, 61, 3022–3029. [Google Scholar] [CrossRef] [PubMed]
- Kotha, R.R.; Tareq, F.S.; Yildiz, E.; Luthria, D.L. Oxidative Stress and Antioxidants—A Critical Review on In Vitro Antioxidant Assays. Antioxidants 2022, 11, 2388. [Google Scholar] [CrossRef] [PubMed]
- Li, C.X.; Wang, F.R.; Zhang, B.; Deng, Z.Y.; Li, H.Y. Stability and antioxidant activity of phenolic compounds during in vitro digestion. J. Food Sci. 2023, 88, 696–716. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, S.; Huang, Y.; Huang, J.; Li, Y. Effect of in vitro gastrointestinal digestion on phenolic compounds and antioxidant properties of soluble and insoluble dietary fibers derived from hulless barley. J. Food Sci. 2021, 86, 628–634. [Google Scholar] [CrossRef]
- Mehta, J.; Rayalam, S.; Wang, X. Cytoprotective Effects of Natural Compounds against Oxidative Stress. Antioxidants 2018, 7, 147. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Venkidasamy, B.; Subramanian, U.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Girish, S.; Thangavel, S.; Dhanapal, A.R.; Fedoseeva, N.; et al. Bioactive compounds in oxidative stress-mediated diseases: Targeting the Nrf2/ARE signaling pathway and epigenetic regulation. Antioxidants 2021, 10, 1859. [Google Scholar] [CrossRef] [PubMed]
- Demirci-Çekiç, S.; Özkan, G.; Avan, A.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of Oxidative Stress and Antioxidant Defense. J. Pharm. Biomed. Anal. 2022, 209, 114477. [Google Scholar] [CrossRef] [PubMed]
- Kučera, O.; Endlicher, R.; Roušar, T.; Lotková, H.; Garnol, T.; Drahota, Z.; Červinková, Z. The effect of tert -butyl hydroperoxide-induced oxidative stress on lean and steatotic rat hepatocytes in vitro. Oxid. Med. Cell. Longev. 2014, 2014, 752506. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative stress mitigation by antioxidants—An overview on their chemistry and influences on health status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar] [CrossRef]
- Moskaug, J.O.; Carlsen, H.; Myhrstad, M.C.W.; Blomhoff, R. Polyphenols and glutathione synthesis regulation. Am. J. Clin. Nutr. 2005, 81, 277S–283S. [Google Scholar] [CrossRef] [PubMed]
- Baba, S.P.; Bhatnagar, A. Role of thiols in oxidative stress. Curr. Opin. Toxicol. 2018, 7, 133–139. [Google Scholar] [CrossRef]
- Benhar, M. Oxidants, antioxidants and thiol Redox switches in the control of regulated cell death pathways. Antioxidants 2020, 9, 309. [Google Scholar] [CrossRef]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. 2018, 12, 88–93. [Google Scholar]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Albano, G.D.; Gagliardo, R.P.; Montalbano, A.M.; Profita, M. Overview of the Mechanisms of Oxidative Stress: Impact in Inflammation of the Airway Diseases. Antioxidants 2022, 11, 2237. [Google Scholar] [CrossRef]
- Pérez-Torres, I.; Manzano-Pech, L.; Rubio-Ruíz, M.E.; Soto, M.E.; Guarner-Lans, V. Nitrosative Stress and Its Association with Cardiometabolic Disorders. Molecules 2020, 25, 2555. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | ND | OP | GP | IP † | CP | |
|---|---|---|---|---|---|---|
| Cocoa shell flour | ||||||
| Hydroxybenzoic acids | ||||||
| 3,4,5-Trihydroxybenzoic acid (Gallic acid) | 16.0 ± 0.4 a | 7.2 ± 0.6 c | 7.1 ± 0.4 c | 11.8 ± 0.8 b | (0.7) | 16.5 ± 1.5 a |
| 3,4-Dihydroxybenzoic acid (Protocatechuic acid) | 6.6 ± 0.6 b | 2.7 ± 0.1 d | 5.4 ± 0.0 bc | 4.6 ± 0.3 c | (0.7) | 10.8 ± 1.0 a |
| Total | 22.6 ± 1.0 b | 9.9 ± 0.6 d | 12.5 ± 0.4 d | 16.5 ± 1.0 c | (0.7) | 27.3 ± 2.5 a |
| N-Phenylpropenoyl-L-amino acids | ||||||
| N-(3-(4-Hydroxycinnamoyl)-L-aspartic acid (N-Coumaroyl-L-aspartate) cis | 0.9 ± 0.0 b | 0.9 ± 0.0 b | 0.8 ± 0.1 b | 1.1 ± 0.1 a | (1.2) | 0.8 ± 0.1 b |
| N-(3-(4-Hydroxycinnamoyl)-L-aspartic acid (N-Coumaroyl-L-aspartate) trans | 0.5 ± 0.0 b | 0.6 ± 0.0 b | 0.5 ± 0.1 b | 1.5 ± 0.1 a | (3.1) | n.d |
| N-(3-(4-Hydroxycinnamoyl)-L-tyrosine (N-Coumaroyl-L-tyrosine) | n.d. | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 0.4 ± 0.0 a | n.d. | 0.4 ± 0.0 a |
| N-(3-(3,4-Dihydroxycinnamoyl)-L-aspartic acid (N-Caffeoyl-L-aspartate) | 5.1 ± 0.3 | n.d. | n.d. | n.d. | (0.0) | n.d. |
| N-(3-(3,4-Dihydroxycinnamoyl)-L-3,4- DOPA (N-Caffeoyl-L-DOPA) cis | 0.9 ± 0.0 a | 0.6 ± 0.0 c | 0.6 ± 0.0 bc | 0.4 ± 0.1 d | (0.5) | 0.7 ± 0.1 b |
| Total | 7.4 ± 0.4 a | 2.1 ± 0.1 c | 2.0 ± 0.1 c | 3.4 ± 0.3 b | (0.5) | 2.0 ± 0.2 c |
| Flavan-3-ols | ||||||
| 2,3,4′,5,7-Pentahydroxyflavan-3,4-diol ((+)-Catechin) | 11.5 ± 0.8 b | 4.0 ± 0.4 d | 10.0 ± 0.2 b | 7.2 ± 0.6 c | (0.6) | 16.5 ± 1.4 a |
| 2,3′,4,5′,7-Pentahydroxyflavan-3,4-diol ((−)-Epicatechin) | 1.3 ± 0.1 b | 1.5 ± 0.1 b | 2.0 ± 0.1 a | 2.3 ± 0.1 a | (1.7) | n.d. |
| Total | 12.8 ± 0.9 b | 5.5 ± 0.5 d | 12.0 ± 0.3 b | 9.4 ± 0.7 c | (0.7) | 16.5 ± 1.4 a |
| Flavonols | ||||||
| 3,3′,4′,5,7-Pentahydroxyflavone 3-β-glucoside (Quercetin 3-O-glucoside) | 0.3 ± 0.0 a | 0.1 ± 0.0 b | 0.1 ± 0.0 b | n.d. | (0.0) | n.d. |
| 3,3′,4′,5,7-Pentahydroxyflavone 3-β-arabinoside (Quercetin 3-O-arabinoside) | 0.3 ± 0.0 a | 0.1 ± 0.0 b | 0.1 ± 0.0 c | n.d. | (0.0) | n.d. |
| Total | 0.6 ± 0.0 a | 0.2 ± 0.0 b | 0.2 ± 0.0 b | n.d. | (0.0) | n.d. |
| Methylxanthines | ||||||
| 3,7-Dimethylxanthine (Theobromine) | 525.8 ± 4.9 a | 236.0 ± 2.0 d | 382.2 ± 41.1 c | 470.1 ± 12.8 b | (0.9) | 510.6 ± 3.1 ab |
| 1,3,7-Trimethylxanthine (Caffeine) | 169.4 ± 0.4 a | 55.8 ± 0.1 d | 101.0 ± 1.1 c | 130.3 ± 1.0 b | (0.8) | 132.9 ± 3.1 b |
| Total | 695.2 ± 5.2 a | 291.8 ± 2.1 d | 483.2 ± 42.2 c | 600.5 ± 13.8 b | (0.9) | 643.5 ± 6.2 b |
| Cocoa shell extract | ||||||
| Hydroxybenzoic acids | ||||||
| 3,4,5-Trihydroxybenzoic acid (Gallic acid) | 73.9 ± 2.2 a | 64.1 ± 1.7 b | 31.1 ± 2.4 d | 39.2 ± 2.6 c | (0.5) | 66.7 ± 6.3 ab |
| 3,4-Dihydroxybenzoic acid (Protocatechuic acid) | 34.9 ± 2.4 b | 40.2 ± 0.7 a | 27.7 ± 0.7 d | 33.4 ± 2.3 bc | (1.0) | 31.0 ± 1.1 cd |
| Total | 108.9 ± 4.7 a | 104.3 ± 2.4 ab | 58.8 ± 3.2 d | 72.6 ± 4.9 c | (0.7) | 97.7 ± 7.5 b |
| N-Phenylpropenoyl-L-amino acids | ||||||
| N-(3-(4-Hydroxycinnamoyl)-L-aspartic acid (N-Coumaroyl-L-aspartate) cis | 6.2 ± 0.6 a | 5.9 ± 0.0 a | 6.4 ± 6.0 a | n.d. | (0.0) | n.d. |
| N-(3-(4-Hydroxycinnamoyl)-L-aspartic acid (N-Coumaroyl-L-aspartate) trans | 4.5 ± 0.3 bc | 4.9 ± 0.4 b | 4.1 ± 0.0 c | 1.7 ± 0.0 d | (0.4) | 5.6 ± 0.3 a |
| N-(3-(4-Hydroxycinnamoyl)-L-tyrosine (N-Coumaroyl-L-tyrosine) | 0.8 ± 0.2 bc | 1.0 ± 0.0 b | 1.3 ± 0.1 a | 0.6 ± 0.1 c | (0.8) | n.d. |
| N-(3-(3,4-Dihydroxycinnamoyl)-L-aspartic acid (N-Caffeoyl-L-aspartate) | 19.1 ± 1.8 ab | 21.1 ± 0.7 a | 17.3 ± 2.0 b | n.d. | (0.0) | n.d. |
| N-(3-(3,4-Dihydroxycinnamoyl)-L-3,4-DOPA (N-Caffeoyl-L-DOPA) cis | 3.8 ± 0.1 b | 3.8 ± 0.4 b | 4.7 ± 0.5 a | 3.1 ± 0.3 c | (0.8) | 2.4 ± 0.0 c |
| N-(3-(3,4-Dihydroxycinnamoyl)-L-3,4-DOPA (N-Caffeoyl-L-DOPA) trans | 1.1 ± 0.0 a | 1.1 ± 0.1 a | 1.0 ± 0.0 a | n.d. | (0.0) | n.d. |
| Total | 35.4 ± 3.0 a | 37.8 ± 1.6 a | 34.7 ± 3.3 a | 5.4 ± 0.4 b | (0.2) | 8.0 ± 0.3 b |
| Flavan-3-ols | ||||||
| 2,3,4′,5,7-Pentahydroxyflavan-3,4-diol ((+)-Catechin) | 46.1 ± 1.4 bc | 41.6 ± 3.8 c | 56.1 ± 5.6 b | 40.3 ± 0.2 c | (0.9) | 71.4 ± 8.3 a |
| 2,3′,4,5′,7-Pentahydroxyflavan-3,4-diol ((−)-Epicatechin) | 3.5 ± 0.1 c | 3.0 ± 0.4 c | 6.0 ± 0.5 b | 3.3 ± 0.3 c | (1.0) | 13.8 ± 1.5 a |
| Total | 49.5 ± 1.5 c | 44.6 ± 4.2 c | 62.1 ± 6.2 b | 43.7 ± 0.6 c | (0.9) | 85.1 ± 9.8 a |
| Flavonols | ||||||
| 3,3′,4′,5,7-Pentahydroxyflavone 3-β-glucoside (Quercetin 3-O-glucoside) | 1.4 ± 0.0 a | 1.3 ± 0.0 b | 1.2 ± 0.0 b | n.d. | (0.0) | n.d. |
| 3,3′,4′,5,7-Pentahydroxyflavone 3-β-arabinoside (Quercetin 3-O-arabinoside) | 1.3 ± 0.0 a | 1.3 ± 0.1 a | 1.3 ± 0.0 a | n.d. | (0.0) | n.d. |
| Total | 2.7 ± 0.1 a | 2.5 ± 0.2 a | 2.5 ± 0.1 a | – | (0.0) | – |
| Flavones | ||||||
| 5,7,4′-Trihydroxyflavone-6,8-di-C-glucoside (Apigenin-6,8-di-C-glucoside) | 2.9 ± 0.0 b | 2.8 ± 0.2 b | 3.4 ± 0.1 a | 1.9 ± 0.2 c | (0.7) | n.d. |
| Methylxanthines | ||||||
| 3,7-Dimethylxanthine (Theobromine) | 2605.3 ± 125.5 a | 2253.0 ± 20.6 b | 1759.8 ± 125.9 c | 1249.8 ± 49.0 d | (0.5) | 1919.3 ± 86.3 c |
| 1,3,7-Trimethylxanthine (Caffeine) | 34.0 ± 2.0 a | 28.1 ± 0.8 b | 25.2 ± 1.6 bc | 20.0 ± 1.7 d | (0.6) | 24.8 ± 1.0 c |
| Total | 2639.3 ± 127.4 a | 2281.1 ± 21.3 b | 1785.0 ± 127.5 c | 1269.8 ± 50.7 d | (0.5) | 1944.2 ± 87.3 c |
| Digestion Phase | CSF | CSE | ||||
|---|---|---|---|---|---|---|
| Digested Fraction | Non-Digested Fraction | Digested Fraction | ||||
| Free | Bound | Total | ||||
| TPC (mg GAE g−1) | Raw Material | – | 19.7 ± 0.9 a | 14.5 ± 0.3 a | 34.2 ± 1.1 a | 46.3 ± 2.5 c |
| Oral Phase | 3.1 ± 0.3 c | 18.0 ± 0.6 b | 14.2 ± 0.6 a | 32.2 ± 1.2 b | 47.2 ± 2.0 bc | |
| Gastric Phase | 2.4 ± 0.3 d | 15.1 ± 0.5 c | 13.5 ± 0.3 a | 28.6 ± 0.8 c | 50.1 ± 1.4 b | |
| Intestinal Phase | 7.7 ± 0.4 a | 10.5 ± 0.5 d | 13.9 ± 1.3 a | 24.4 ± 1.3 d | 58.0 ± 1.8 a | |
| Colonic Phase | 6.6 ± 0.5 b | 14.7 ± 0.5 c | 10.1 ± 0.5 b | 24.8 ± 1.1 d | 58.1 ± 2.2 a | |
| ABTS (mg TE g−1) | Raw Material | – | 38.6 ± 0.7 a | 34.8 ± 1.0 ab | 73.4 ± 1.7 a | 85.0 ± 1.7 d |
| Oral Phase | 2.0 ± 1.0 d | 37.4 ± 0.4 a | 34.3 ± 1.1 b | 71.7 ± 1.5 a | 88.4 ± 2.1 d | |
| Gastric Phase | 4.0 ± 0.3 c | 31.9 ± 1.3 c | 34.2 ± 1.3 b | 66.1 ± 2.6 b | 96.8 ± 2.1 c | |
| Intestinal Phase | 30.9 ± 0.7 b | 25.6 ± 1.7 d | 36.2 ± 1.3 a | 61.8 ± 3.0 c | 633.0 ± 10.4 a | |
| Colonic Phase | 38.9 ± 0.7 a | 33.9 ± 0.6 b | 25.8 ± 0.8 c | 59.7 ± 1.4 d | 601.4 ± 2.8 b | |
| FRAP (mmol TE g−1) | Raw Material | – | 48.9 ± 2.9 a | 27.5 ± 1.8 d | 76.4 ± 4.8 b | 23.5 ± 1.6 a |
| Oral Phase | 2.0 ± 0.2 d | 47.6 ± 1.7 a | 30.2 ± 1.4 c | 77.8 ± 3.0 ab | 22.4 ± 1.8 a | |
| Gastric Phase | 13.8 ± 0.6 c | 42.2 ± 1.2 b | 37.8 ± 1.5 b | 80.0 ± 2.7 a | 20.5 ± 0.8 b | |
| Intestinal Phase | 26.8 ± 1.6 a | 27.1 ± 1.5 d | 40.3 ± 1.6 a | 67.4 ± 3.1 c | 16.3 ± 1.1 c | |
| Colonic Phase | 22.5 ± 1.3 b | 35.9 ± 1.0 c | 24.6 ± 0.6 e | 60.5 ± 1.6 d | 12.0 ± 0.8 d | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cañas, S.; Rebollo-Hernanz, M.; Bermúdez-Gómez, P.; Rodríguez-Rodríguez, P.; Braojos, C.; Gil-Ramírez, A.; Benítez, V.; Aguilera, Y.; Martín-Cabrejas, M.A. Radical Scavenging and Cellular Antioxidant Activity of the Cocoa Shell Phenolic Compounds after Simulated Digestion. Antioxidants 2023, 12, 1007. https://doi.org/10.3390/antiox12051007
Cañas S, Rebollo-Hernanz M, Bermúdez-Gómez P, Rodríguez-Rodríguez P, Braojos C, Gil-Ramírez A, Benítez V, Aguilera Y, Martín-Cabrejas MA. Radical Scavenging and Cellular Antioxidant Activity of the Cocoa Shell Phenolic Compounds after Simulated Digestion. Antioxidants. 2023; 12(5):1007. https://doi.org/10.3390/antiox12051007
Chicago/Turabian StyleCañas, Silvia, Miguel Rebollo-Hernanz, Patricia Bermúdez-Gómez, Pilar Rodríguez-Rodríguez, Cheyenne Braojos, Alicia Gil-Ramírez, Vanesa Benítez, Yolanda Aguilera, and María A. Martín-Cabrejas. 2023. "Radical Scavenging and Cellular Antioxidant Activity of the Cocoa Shell Phenolic Compounds after Simulated Digestion" Antioxidants 12, no. 5: 1007. https://doi.org/10.3390/antiox12051007