

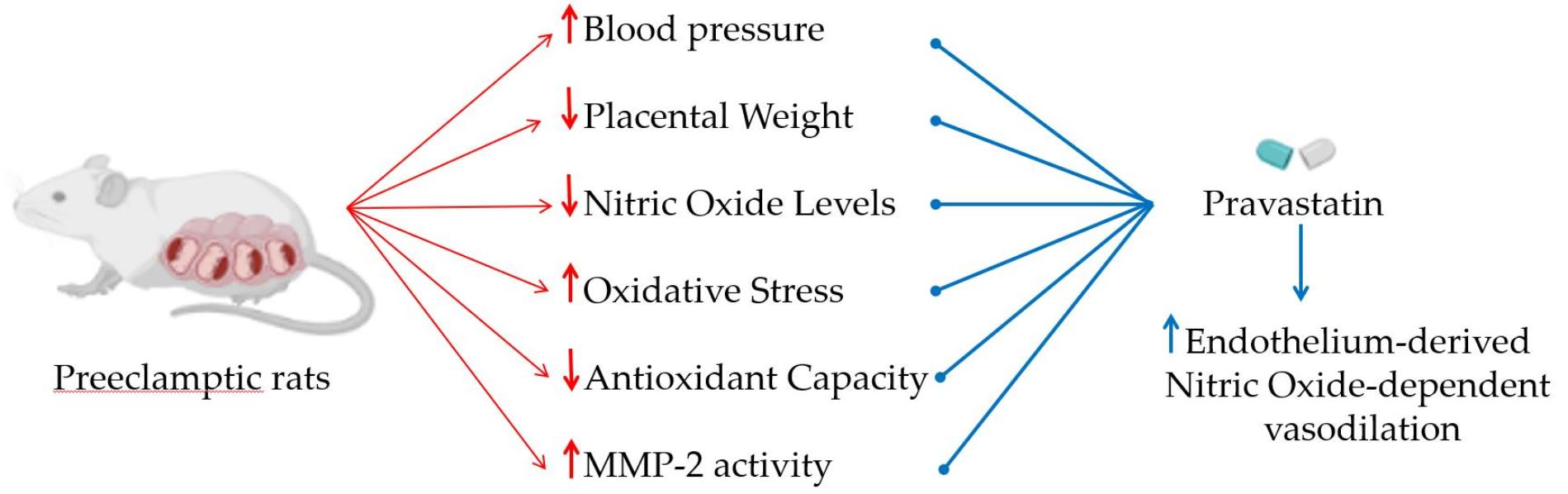
Pravastatin Prevents Increases in Activity of Metalloproteinase-2 and Oxidative Stress, and Enhances Endothelium-Derived Nitric Oxide-Dependent Vasodilation in Gestational Hypertension
 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Protocols
- Normotensive Pregnant rats (Norm-Preg group): saline (0.9% NaCl) solution (0.3–0.45 mL) was intraperitoneally (i.p.) administered on days 1, 7, and 14, and saline was administered by gavage from pregnancy day 10 until 19 (n = 8).
- Normotensive pregnant rats treated with pravastatin (Norm-Preg + Prava group): saline was i.p. administered on days 1, 7, and 14, and pravastatin (10 mg/kg/day) was administrated by gavage from pregnancy day 10 until 19 (n = 8).
- Hypertensive pregnant rats (HTN-Preg group): hypertension was induced by i.p. administration of 12.5 mg of DOCA on the first day of pregnancy, followed by i.p. injection of 6.5 mg of DOCA on days 7 and 14 of pregnancy; drinking water was replaced by saline from pregnancy day 1 until 19; and saline was administered by gavage from pregnancy day 10 until 19 (n = 8).
- Hypertensive pregnant rats treated with pravastatin (HTN-Preg + Prava group): hypertension was induced by i.p. administration of 12.5 mg of DOCA on the first day of pregnancy, followed by i.p. injection of 6.5 mg of DOCA on days 7 and 14 of pregnancy; drinking water was replaced by saline from pregnancy day 1 until 19; and pravastatin (10 mg/kg/day) was administrated by gavage from pregnancy day 10 until 19 (n = 8).
2.2. Institutional Review Board Statement
2.3. Blood Pressure Measurements
2.4. Vascular Reactivity
2.5. Determination of NO Metabolites (Nitrite/Nitrate: NOx) Levels in Plasma
2.6. Determination of Lipid Peroxide Levels in Plasma
2.7. Determination of Plasma Antioxidant Capacity
2.8. Zymography for MMP-2 Activity
2.9. Data Analysis and Statistics
3. Results
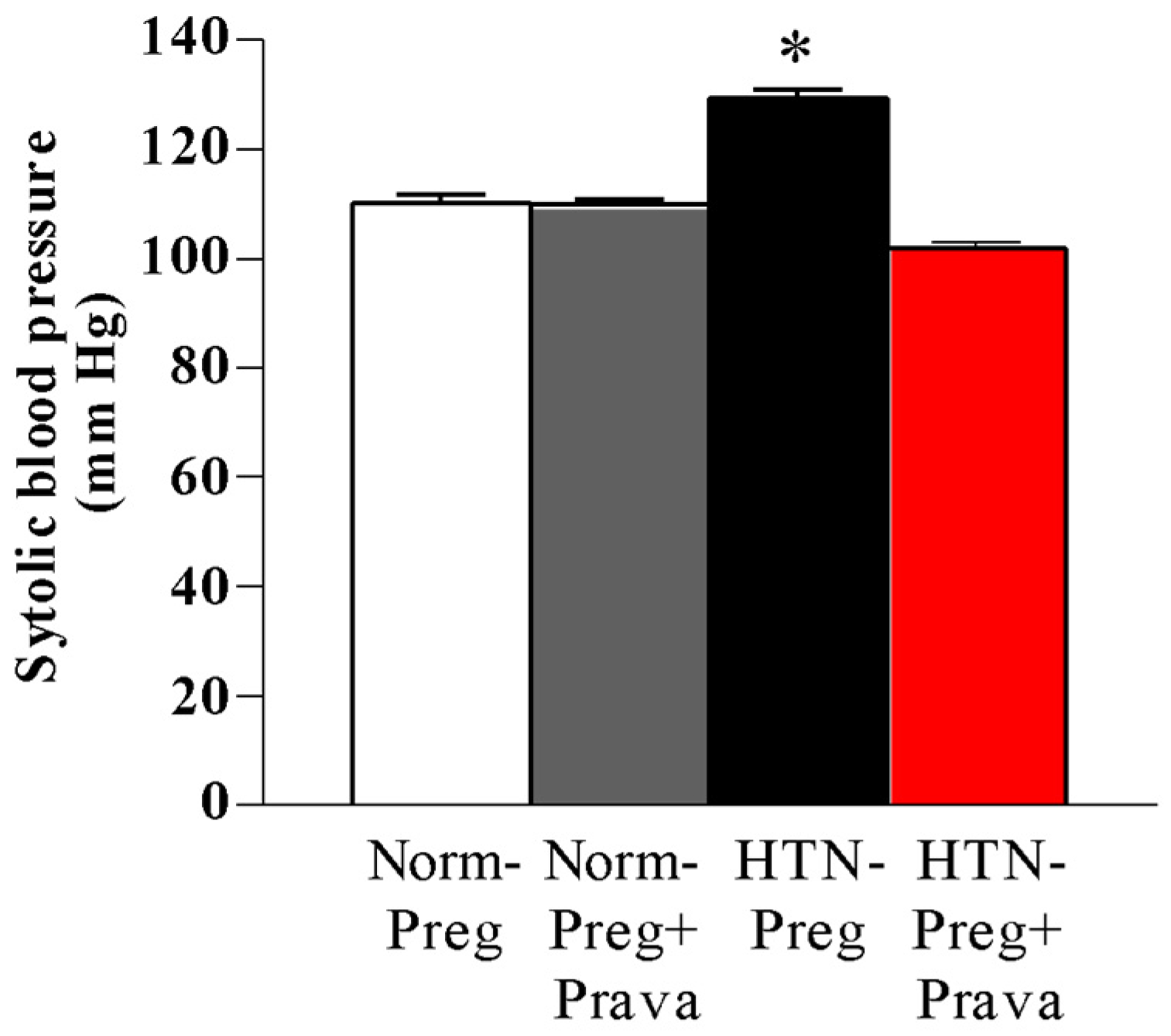
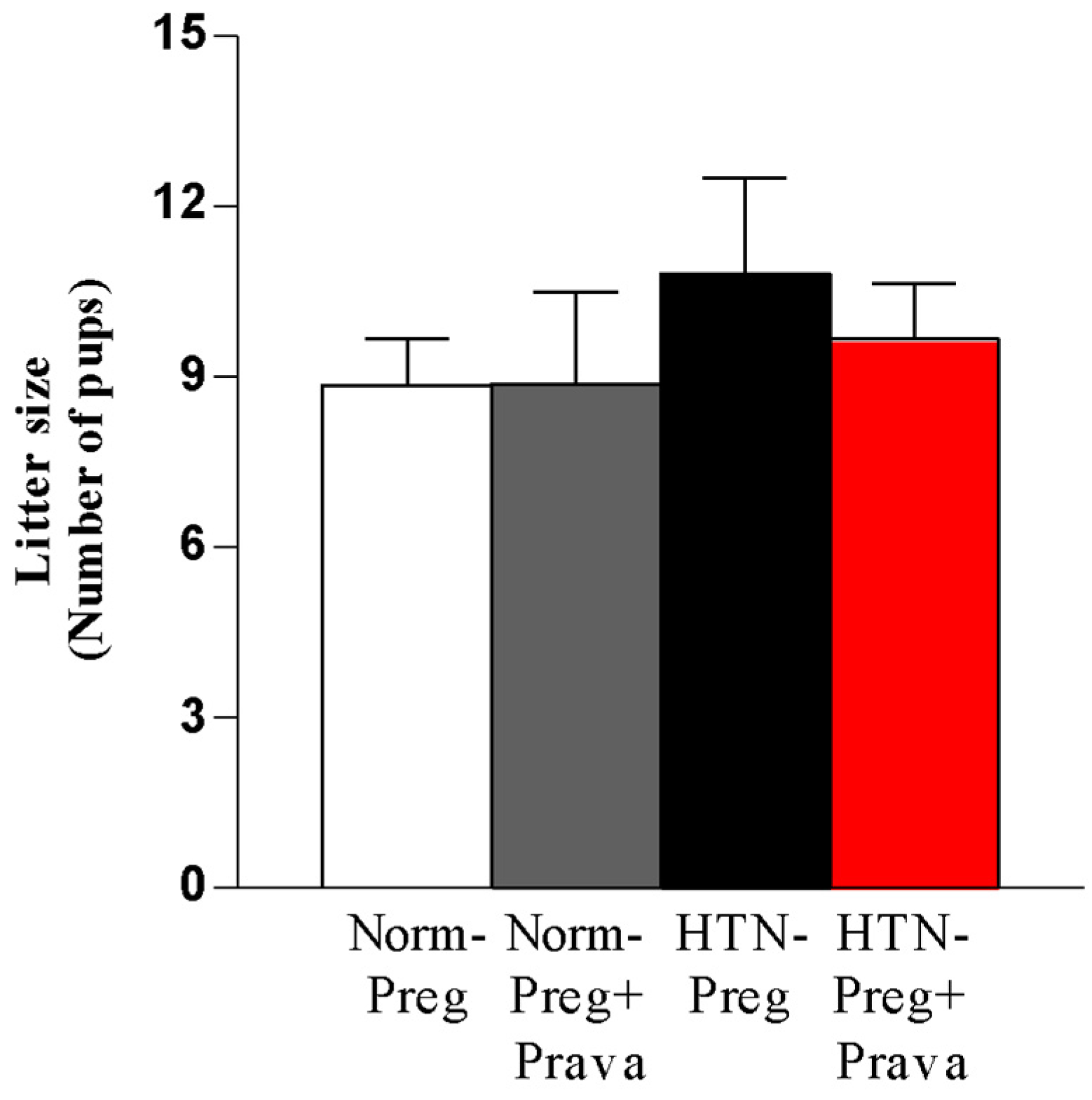
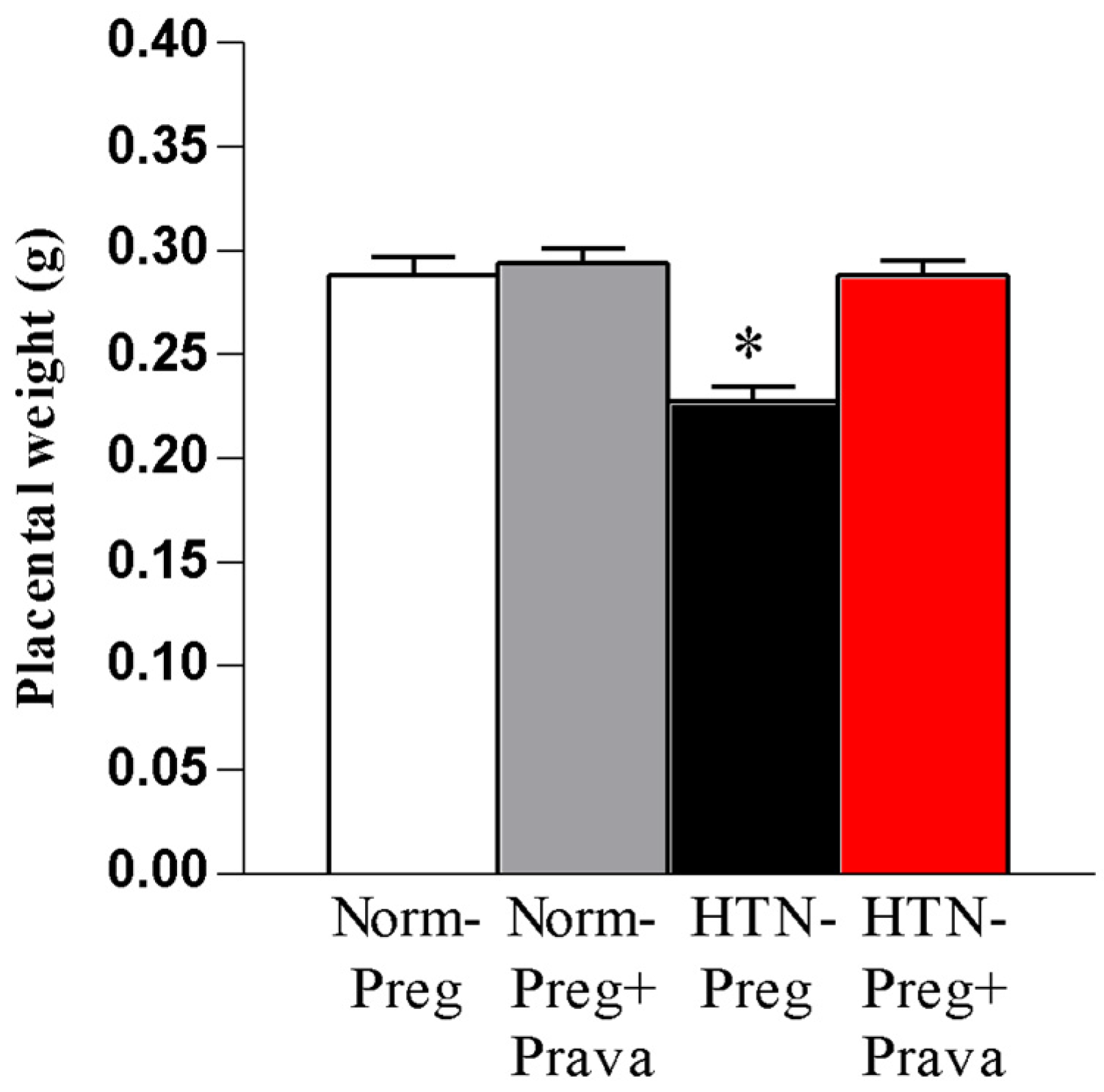
3.1. Pravastatin Prevents Both Hypertensive Pregnancy and Placental Growth Restriction
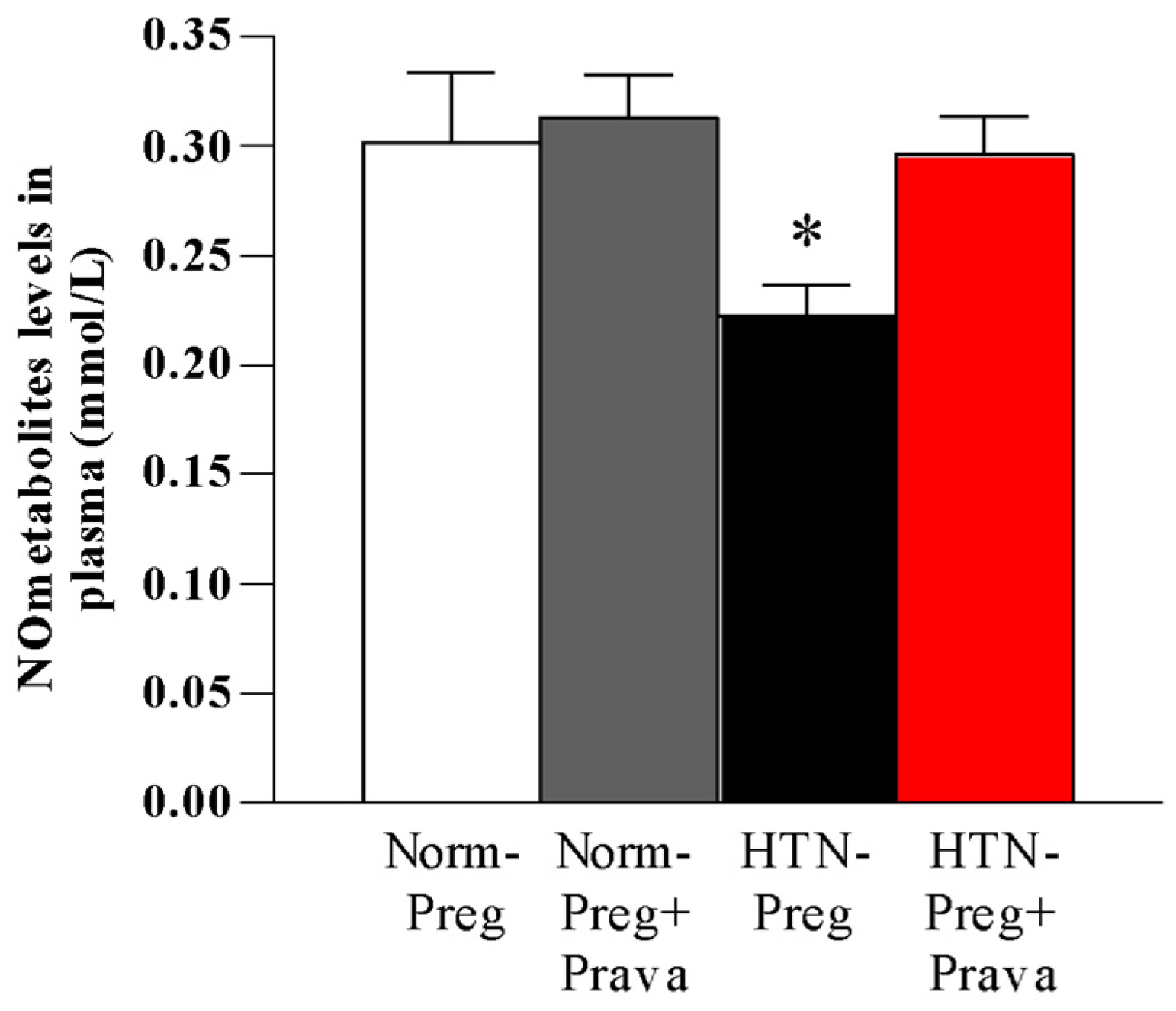
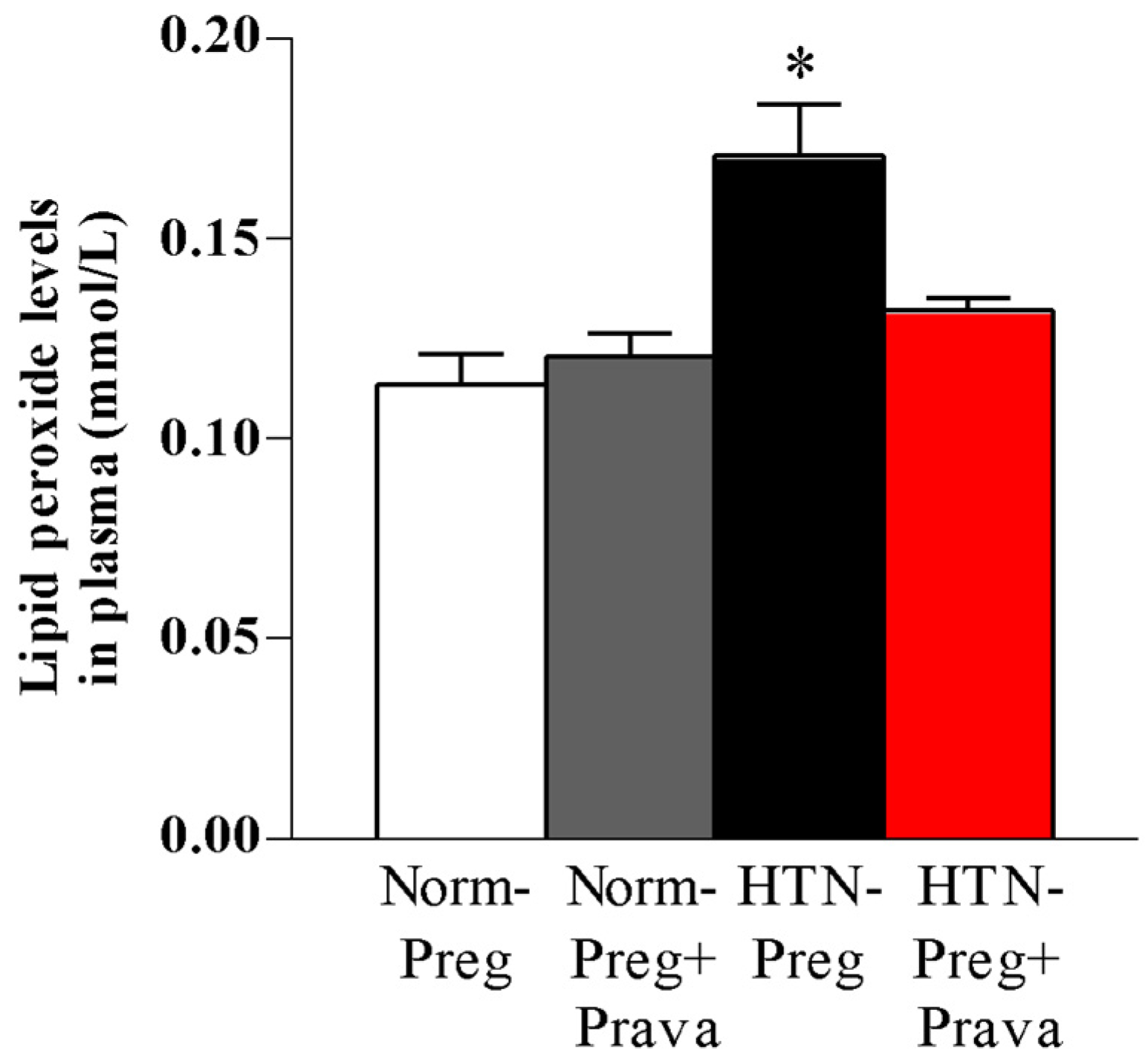
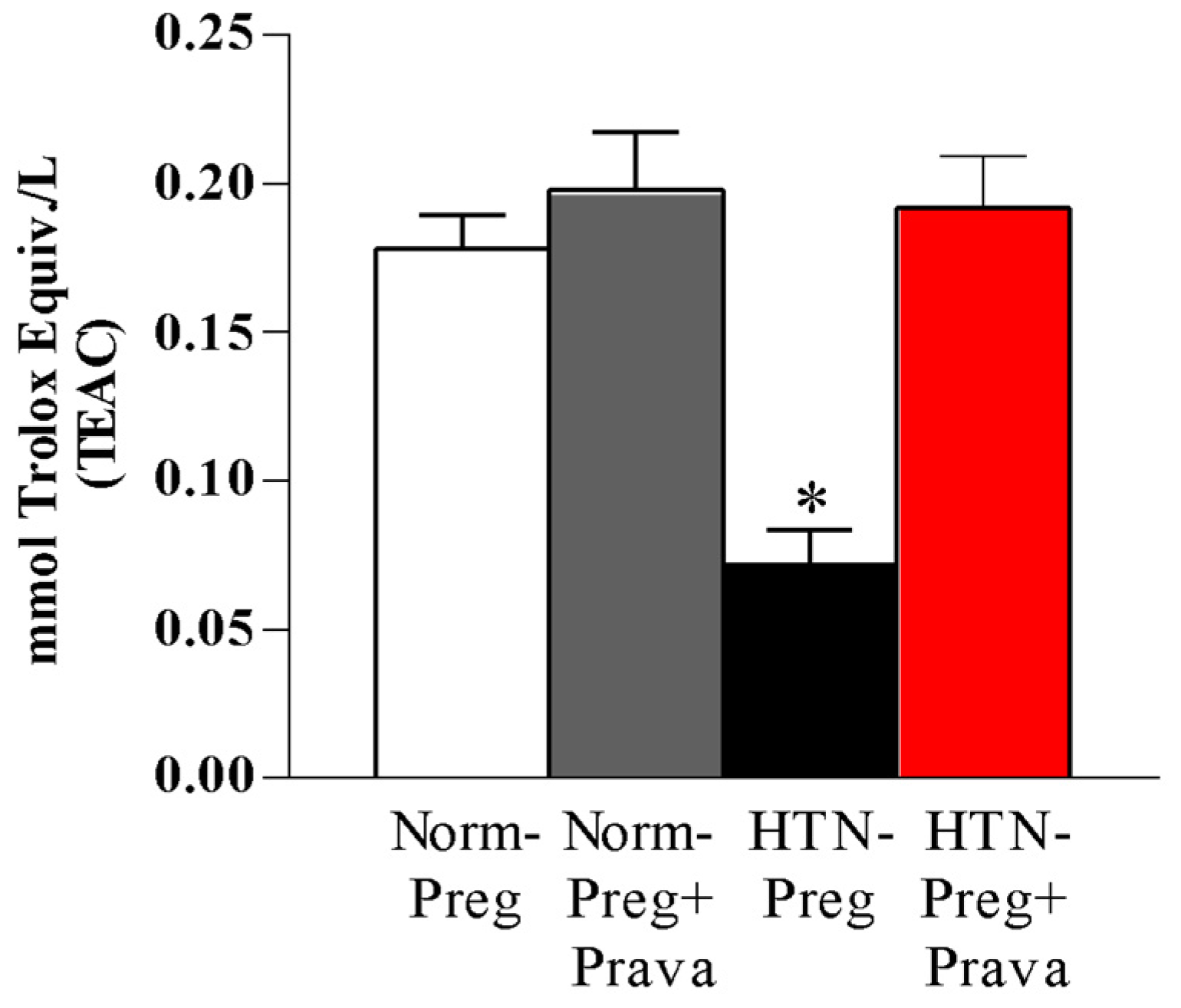
3.2. Pravastatin Prevents Both Reductions in Circulating Levels of NO Metabolites and Increases in Lipid Peroxide Levels in Plasma of Hypertension in Pregnancy, and Restores Plasma Antioxidant Capacity
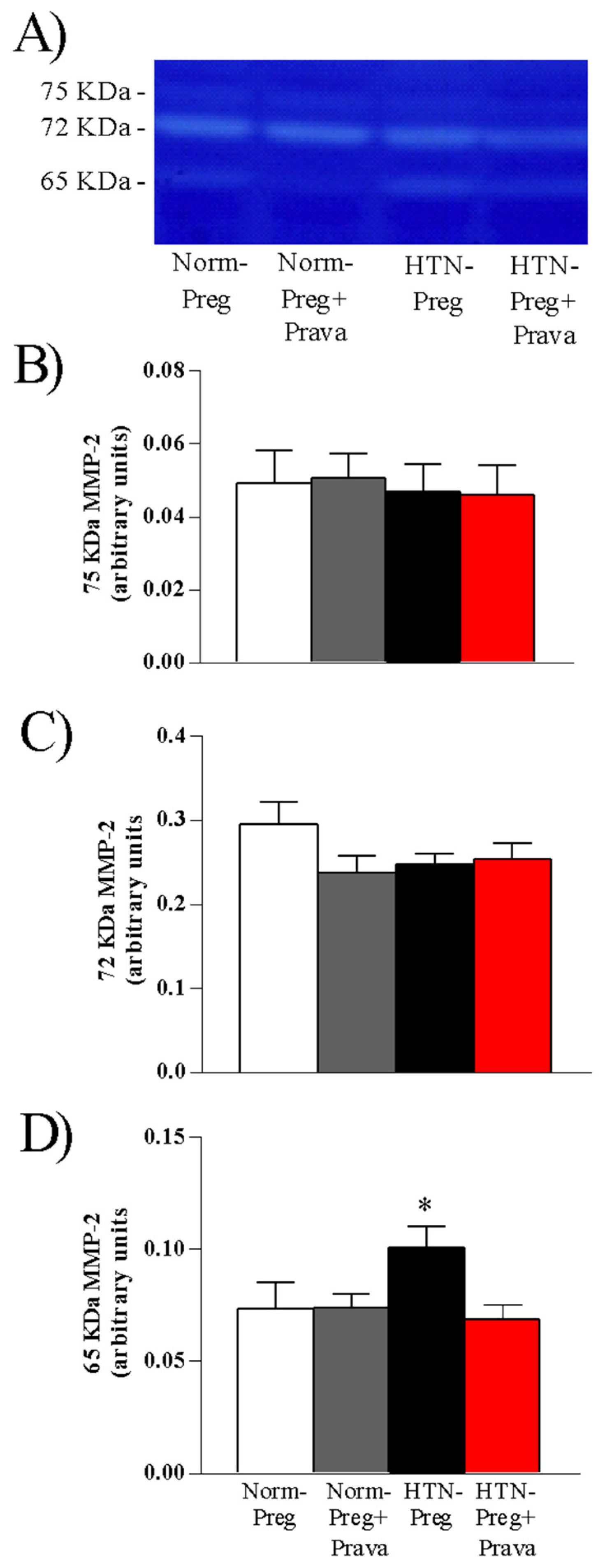
3.3. Pravastatin Prevents Increases in Activity of MMP-2 in Placenta Induced by Hypertensive Pregnancy
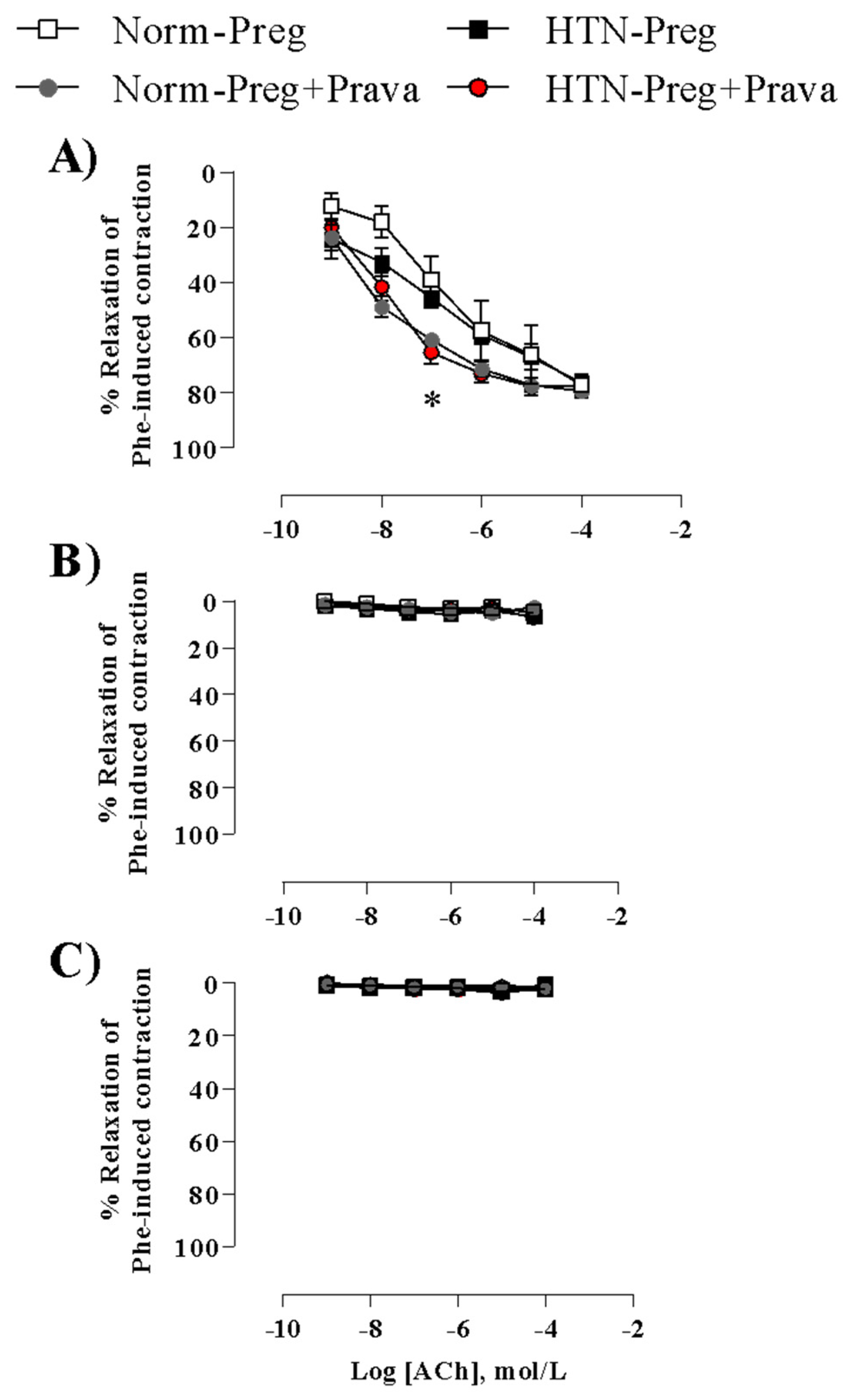
3.4. Pravastatin Enhances Endothelium-Derived NO-Dependent Vasodilation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davignon, J. Beneficial Cardiovascular Pleiotropic Effects of Statins. Circulation 2004, 109 (Suppl. 1), III39–III43. [Google Scholar] [CrossRef] [PubMed]
- Kumasawa, K.; Iriyama, T.; Nagamatsu, T.; Osuga, Y.; Fujii, T. Pravastatin for Preeclampsia: From Animal to Human. J. Obstet. Gynaecol. Res. 2020, 46, 1255–1262. [Google Scholar] [CrossRef]
- Costantine, M.M.; Cleary, K.; Hebert, M.F.; Ahmed, M.S.; Brown, L.M.; Ren, Z.; Easterling, T.R.; Haas, D.M.; Haneline, L.S.; Caritis, S.N.; et al. Safety and Pharmacokinetics of Pravastatin Used for the Prevention of Preeclampsia in High-Risk Pregnant Women: A Pilot Randomized Controlled Trial. Am. J. Obstet Gynecol. 2016, 214, 720.e1–720.e17. [Google Scholar] [CrossRef] [PubMed]
- Chimini, J.S.; Possomato-Vieira, J.S.; da Silva, M.L.S.; Dias-Junior, C.A. Placental Nitric Oxide Formation and Endothelium-dependent Vasodilation Underlie Pravastatin Effects against Angiogenic Imbalance, Hypertension in Pregnancy and Intrauterine Growth Restriction. Basic. Clin. Pharmacol. Toxicol. 2019, 124, 385–393. [Google Scholar] [CrossRef]
- Kumasawa, K.; Ikawa, M.; Kidoya, H.; Hasuwa, H.; Saito-Fujita, T.; Morioka, Y.; Takakura, N.; Kimura, T.; Okabe, M. Pravastatin Induces Placental Growth Factor (PGF) and Ameliorates Preeclampsia in a Mouse Model. Proc. Natl. Acad. Sci. USA 2011, 108, 1451–1455. [Google Scholar] [CrossRef] [PubMed]
- Khan, B.; Allah Yar, R.; Khakwani, A.k.; Karim, S.; Arslan Ali, H. Preeclampsia Incidence and Its Maternal and Neonatal Outcomes With Associated Risk Factors. Cureus 2022, 14, e31143. [Google Scholar] [CrossRef]
- Baumwell, S.; Karumanchi, S.A. Pre-Eclampsia: Clinical Manifestations and Molecular Mechanisms. Nephron. Clin. Pract. 2007, 106, c72–c81. [Google Scholar] [CrossRef]
- Lopez-Jaramillo, P.; Barajas, J.; Rueda-Quijano, S.M.; Lopez-Lopez, C.; Felix, C. Obesity and Preeclampsia: Common Pathophysiological Mechanisms. Front. Physiol. 2018, 9, 1838. [Google Scholar] [CrossRef]
- Perry, A.; Stephanou, A.; Rayman, M.P. Dietary Factors That Affect the Risk of Pre-Eclampsia. BMJ Nutr. Prev. Health 2022, 5, 118–133. [Google Scholar] [CrossRef]
- Garovic, V.D.; August, P. Preeclampsia and the Future Risk of Hypertension: The Pregnant Evidence. Curr. Hypertens Rep. 2013, 15, 114–121. [Google Scholar] [CrossRef]
- Hodžić, J.; Izetbegović, S.; Muračević, B.; Iriškić, R.; Štimjanin Jović, H. Nitric Oxide Biosynthesis during Normal Pregnancy and Pregnancy Complicated by Preeclampsia. Med. Glas. 2017, 14, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Myatt, L.; Webster, R.P. Vascular Biology of Preeclampsia. JTH J. Thromb. Haemost. 2009, 7, 375–384. [Google Scholar] [CrossRef]
- Possomato-Vieira, J.S.; Khalil, R.A. Mechanisms of Endothelial Dysfunction in Hypertensive Pregnancy and Preeclampsia. Adv. Pharmacol. 2016, 77, 361–431. [Google Scholar] [CrossRef] [PubMed]
- Possomato-Vieira, J.S.; Chimini, J.S.; da Silva, M.L.S.; Dias-Junior, C.A. Increases in Placental Nitric Oxide, but Not Nitric Oxide-Mediated Relaxation, Underlie the Improvement in Placental Efficiency and Antihypertensive Effects of Hydrogen Sulphide Donor in Hypertensive Pregnancy. Clin. Exp. Pharmacol. Physiol. 2018, 45, 1118–1127. [Google Scholar] [CrossRef]
- Sandrim, V.C.; Montenegro, M.F.; Palei, A.C.T.; Metzger, I.F.; Sertorio, J.T.C.; Cavalli, R.C.; Tanus-Santos, J.E. Increased Circulating Cell-Free Hemoglobin Levels Reduce Nitric Oxide Bioavailability in Preeclampsia. Free Radic. Biol. Med. 2010, 49, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, R.A.; Possomato-Vieira, J.S.; Bonacio, G.F.; Rizzi, E.; Dias-Junior, C.A. Reductions of Circulating Nitric Oxide Are Followed by Hypertension during Pregnancy and Increased Activity of Matrix Metalloproteinases-2 and -9 in Rats. Cells 2019, 8, 1402. [Google Scholar] [CrossRef] [PubMed]
- Sahay, A.S.; Jadhav, A.T.; Sundrani, D.P.; Wagh, G.N.; Mehendale, S.S.; Joshi, S.R. Matrix Metalloproteinases-2 (MMP-2) and Matrix Metalloproteinases -9 (MMP-9) Are Differentially Expressed in Different Regions of Normal and Preeclampsia Placentae. J. Cell. Biochem. 2018, 119, 6657–6664. [Google Scholar] [CrossRef]
- Svineng, G.; Ravuri, C.; Rikardsen, O.; Huseby, N.-E.; Winberg, J.-O. The Role of Reactive Oxygen Species in Integrin and Matrix Metalloproteinase Expression and Function. Connect Tissue Res. 2008, 49, 197–202. [Google Scholar] [CrossRef]
- Chen, J.; Khalil, R.A. Matrix Metalloproteinases in Normal Pregnancy and Preeclampsia. Prog. Mol. Biol. Transl. Sci. 2017, 148, 87–165. [Google Scholar] [CrossRef]
- Laskowska, M. Altered Maternal Serum Matrix Metalloproteinases MMP-2, MMP-3, MMP-9, and MMP-13 in Severe Early- and Late-Onset Preeclampsia. BioMed Res. Int. 2017, 2017, 6432426. [Google Scholar] [CrossRef]
- Timokhina, E.; Strizhakov, A.; Ibragimova, S.; Gitel, E.; Ignatko, I.; Belousova, V.; Zafiridi, N. Matrix Metalloproteinases MMP-2 and MMP-9 Occupy a New Role in Severe Preeclampsia. J. Pregnancy 2020, 2020, 8369645. [Google Scholar] [CrossRef] [PubMed]
- Aouache, R.; Biquard, L.; Vaiman, D.; Miralles, F. Oxidative Stress in Preeclampsia and Placental Diseases. Int. J. Mol. Sci. 2018, 19, 1496. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M.; Briones, A.M. Reactive Oxygen Species and Vascular Biology: Implications in Human Hypertension. Hypertens. Res. 2011, 34, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, É.; Castro, M.M.; Prado, C.M.; Silva, C.A.; Fazan, R.; Rossi, M.A.; Tanus-Santos, J.E.; Gerlach, R.F. Matrix Metalloproteinase Inhibition Improves Cardiac Dysfunction and Remodeling in 2-Kidney, 1-Clip Hypertension. J. Card Fail. 2010, 16, 599–608. [Google Scholar] [CrossRef]
- Erel, O. A Novel Automated Direct Measurement Method for Total Antioxidant Capacity Using a New Generation, More Stable ABTS Radical Cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef]
- Rizzi, E.; Ceron, C.S.; Guimaraes, D.A.; Prado, C.M.; Rossi, M.A.; Gerlach, R.F.; Tanus-Santos, J.E. Temporal Changes in Cardiac Matrix Metalloproteinase Activity, Oxidative Stress, and TGF-β in Renovascular Hypertension-Induced Cardiac Hypertrophy. Exp. Mol. Pathol. 2013, 94, 1–9. [Google Scholar] [CrossRef]
- Costantine, M.M.; Cleary, K. Pravastatin for the Prevention of Preeclampsia in High-Risk Pregnant Women. Obstet. Gynecol. 2013, 121, 349–353. [Google Scholar] [CrossRef]
- Li, W.; Mata, K.M.; Mazzuca, M.Q.; Khalil, R.A. Altered Matrix Metalloproteinase-2 and -9 Expression/Activity Links Placental Ischemia and Anti-Angiogenic SFlt-1 to Uteroplacental and Vascular Remodeling and Collagen Deposition in Hypertensive Pregnancy. Biochem. Pharmacol. 2014, 89, 370–385. [Google Scholar] [CrossRef]
- Hatanaka, T. Clinical Pharmacokinetics of Pravastatin. Clin. Pharmacokinet. 2000, 39, 397–412. [Google Scholar] [CrossRef]
- Zarek, J.; DeGorter, M.K.; Lubetsky, A.; Kim, R.B.; Laskin, C.A.; Berger, H.; Koren, G. The Transfer of Pravastatin in the Dually Perfused Human Placenta. Placenta 2013, 34, 719–721. [Google Scholar] [CrossRef]
- Nanovskaya, T.N.; Patrikeeva, S.L.; Paul, J.; Costantine, M.M.; Hankins, G.D.V.; Ahmed, M.S. Transplacental Transfer and Distribution of Pravastatin. Am. J. Obstet Gynecol. 2013, 209, 373.e1–373.e5. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Williams, D.; Cheed, V.; Middleton, L.; Ahmad, S.; Wang, K.; Vince, A.; Hewett, P.; Spencer, K.; Khan, K.; et al. Pravastatin for Early-onset Pre-eclampsia: A Randomised, Blinded, Placebo-controlled Trial. BJOG Int. J. Obstet Gy. 2020, 127, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Fruci, S.; Salvi, S.; Moresi, S.; Gallini, F.; Dell’Aquila, M.; Arena, V.; Di Stasio, E.; Ferrazzani, S.; De Carolis, S.; Lanzone, A. Pravastatin for Severe Preeclampsia with Growth Restriction: Placental Findings and Infant Follow-Up. Eur. J. Obstet. Gynecol. Reprod. Biol. 2023, 283, 37–42. [Google Scholar] [CrossRef]
- Redecha, P.; van Rooijen, N.; Torry, D.; Girardi, G. Pravastatin Prevents Miscarriages in Mice: Role of Tissue Factor in Placental and Fetal Injury. Blood 2009, 113, 4101–4109. [Google Scholar] [CrossRef] [PubMed]
- Pánczél, Z.; Kukor, Z.; Supák, D.; Kovács, B.; Kecskeméti, A.; Czizel, R.; Djurecz, M.; Alasztics, B.; Csomó, K.B.; Hrabák, A.; et al. Pravastatin Induces NO Synthesis by Enhancing Microsomal Arginine Uptake in Healthy and Preeclamptic Placentas. BMC Pregnancy Childbirth 2019, 19, 426. [Google Scholar] [CrossRef]
- Guerby, P.; Tasta, O.; Swiader, A.; Pont, F.; Bujold, E.; Parant, O.; Vayssiere, C.; Salvayre, R.; Negre-Salvayre, A. Role of Oxidative Stress in the Dysfunction of the Placental Endothelial Nitric Oxide Synthase in Preeclampsia. Redox Biol. 2021, 40, 101861. [Google Scholar] [CrossRef]
- Onat, T.; Aydoğan Kırmızı, D.; Başer, E.; Ercan, M.; Demir Çaltekin, M.; Yalçın, S.; Kara, M.; Esinler, D.; Yalvaç, E.S. The Relationship between Oxidative Stress and Preeclampsia. The Serum Ischemia-Modified Albumin Levels and Thiol/Disulfide Homeostasis. Turk. J. Obstet. Gynecol. 2020, 17, 102–107. [Google Scholar] [CrossRef]
- Bauer, A.J.; Banek, C.T.; Needham, K.; Gillham, H.; Capoccia, S.; Regal, J.F.; Gilbert, J.S. Pravastatin Attenuates Hypertension, Oxidative Stress, and Angiogenic Imbalance in Rat Model of Placental Ischemia-Induced Hypertension. Hypertension 2013, 61, 1103–1111. [Google Scholar] [CrossRef]
- Matsubara, K.; Matsubara, Y.; Hyodo, S.; Katayama, T.; Ito, M. Role of Nitric Oxide and Reactive Oxygen Species in the Pathogenesis of Preeclampsia. J. Obstet. Gynaecol. Res. 2010, 36, 239–247. [Google Scholar] [CrossRef]
- Eleuterio, N.M.; Palei, A.C.T.; Rangel Machado, J.S.; Tanus-Santos, J.E.; Cavalli, R.C.; Sandrim, V.C. Positive Correlations between Circulating Adiponectin and MMP2 in Preeclampsia Pregnant. Pregnancy Hypertens. 2015, 5, 205–208. [Google Scholar] [CrossRef]
- Blascke de Mello, M.M.; Parente, J.M.; Schulz, R.; Castro, M.M. Matrix Metalloproteinase (MMP)-2 Activation by Oxidative Stress Decreases Aortic Calponin-1 Levels during Hypertrophic Remodeling in Early Hypertension. Vascul. Pharmacol. 2019, 116, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.; Oh, H.-R.; Unsworth, E.J.; Bregonzio, C.; Saavedra, J.M.; Stetler-Stevenson, W.G.; Cuttitta, F. Matrix Metalloproteinase-2 Cleavage of Adrenomedullin Produces a Vasoconstrictor out of a Vasodilator. Biochem. J. 2004, 383, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Patron, C.; Radomski, M.W.; Davidge, S.T. Vascular Matrix Metalloproteinase-2 Cleaves Big Endothelin-1 Yielding a Novel Vasoconstrictor. Circ. Res. 1999, 85, 906–911. [Google Scholar] [CrossRef]
- Costantine, M.M.; Tamayo, E.; Lu, F.; Bytautiene, E.; Longo, M.; Hankins, G.D.V.; Saade, G.R. Using Pravastatin to Improve the Vascular Reactivity in a Mouse Model of Soluble Fms-Like Tyrosine Kinase-1–Induced Preeclampsia. Obstet. Gynecol. 2010, 116, 114–120. [Google Scholar] [CrossRef]
- Xu, P.; Alfaidy, N.; Challis, J.R.G. Expression of Matrix Metalloproteinase (MMP)-2 and MMP-9 in Human Placenta and Fetal Membranes in Relation to Preterm and Term Labor. J. Clin. Endocrinol. Metab. 2002, 87, 1353–1361. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toghi, C.J.; Martins, L.Z.; Pacheco, L.L.; Caetano, E.S.P.; Mattos, B.R.; Rizzi, E.; Dias-Junior, C.A. Pravastatin Prevents Increases in Activity of Metalloproteinase-2 and Oxidative Stress, and Enhances Endothelium-Derived Nitric Oxide-Dependent Vasodilation in Gestational Hypertension. Antioxidants 2023, 12, 939. https://doi.org/10.3390/antiox12040939
Toghi CJ, Martins LZ, Pacheco LL, Caetano ESP, Mattos BR, Rizzi E, Dias-Junior CA. Pravastatin Prevents Increases in Activity of Metalloproteinase-2 and Oxidative Stress, and Enhances Endothelium-Derived Nitric Oxide-Dependent Vasodilation in Gestational Hypertension. Antioxidants. 2023; 12(4):939. https://doi.org/10.3390/antiox12040939
Chicago/Turabian StyleToghi, Cristal Jesus, Laisla Zanetoni Martins, Leonardo Lopes Pacheco, Edileia Souza Paula Caetano, Bruna Rahal Mattos, Elen Rizzi, and Carlos Alan Dias-Junior. 2023. "Pravastatin Prevents Increases in Activity of Metalloproteinase-2 and Oxidative Stress, and Enhances Endothelium-Derived Nitric Oxide-Dependent Vasodilation in Gestational Hypertension" Antioxidants 12, no. 4: 939. https://doi.org/10.3390/antiox12040939