
Antioxidant and Immune Stimulating Effects of Allium cepa Skin in the RAW 264.7 Cells and in the C57BL/6 Mouse Immunosuppressed by Cyclophosphamide
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Plant Material and Reagents
2.1.1. Sample Preparation for the Experiment
2.1.2. Reagents for the Experiment
2.2. Evaluation of Functional Compounds and Antioxidant Activity
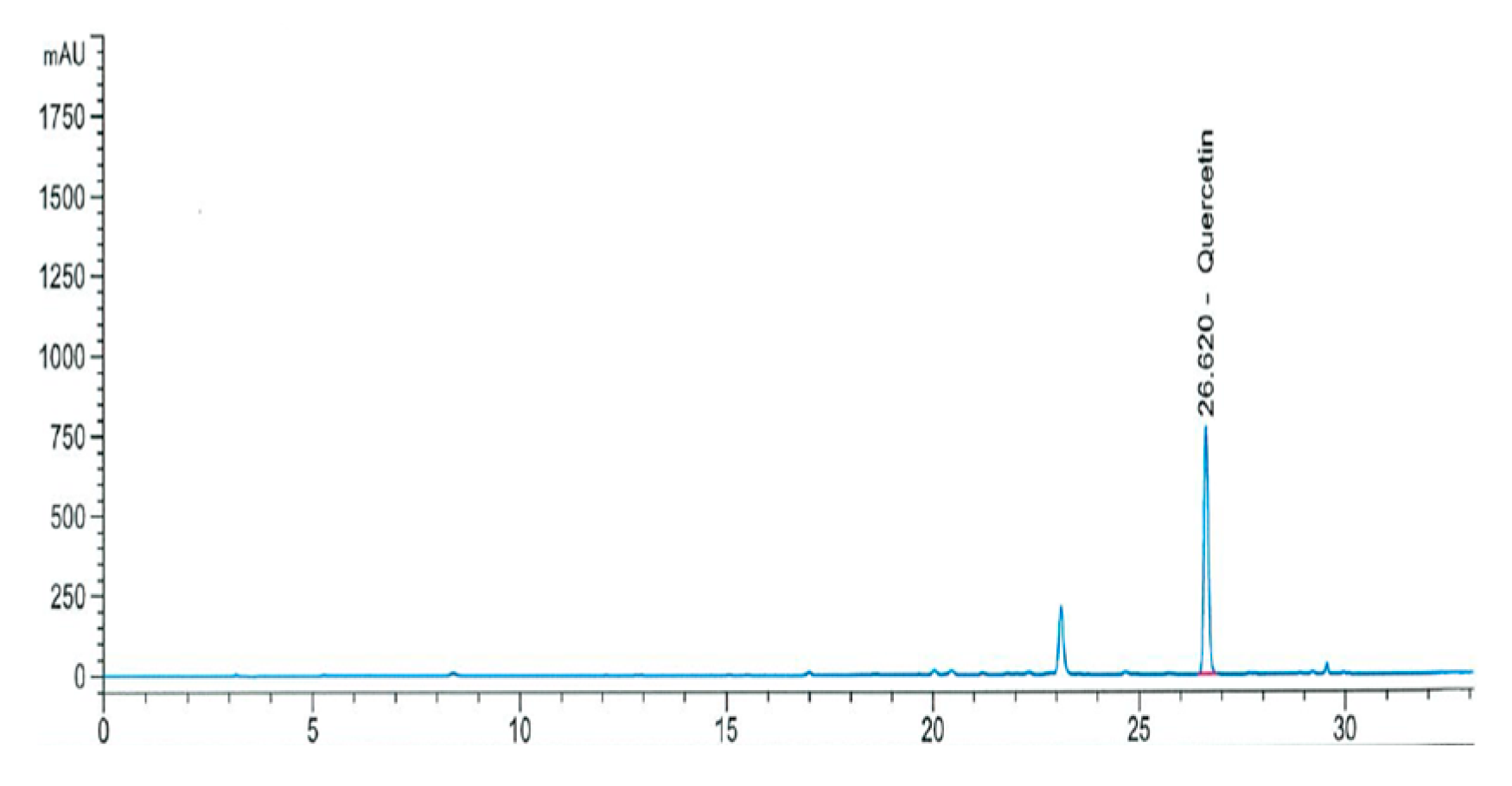
2.2.1. Quercetin Concentration
2.2.2. Total Phenolic Content
2.2.3. DPPH Radical Scavenging Activity
2.2.4. ABTS Radical Scavenging Activity
2.2.5. Xanthine Oxidase Inhibitory Activity
2.3. Cell Experiment for Evaluations of Antioxidant and Immunomodulatory Effects
2.3.1. Cell Viability
2.3.2. Superoxide Dismutase Activity
2.3.3. Catalase Activity
2.3.4. Nitric Oxide Concentration
2.3.5. Cytokine Concentrations
2.3.6. Cytokine Expression
2.4. Animal Experiment
2.4.1. Experimental Design
- Group 1: NOR (normal control, distilled water (DW)) (n = 10);
- Group 2: NC (negative control, CPA, DW) (n = 10);
- Group 3: PC (positive control, CPA, β-glucan 50 mg/kg BW) (n = 10);
- Group 4: OS100 (CPA, OS extract 100 mg/kg BW) (n = 10);
- Group 5: OS200 (CPA, OS extract 200 mg/kg BW) (n = 10).
2.4.2. Collecting Blood and Hematological Analysis
2.4.3. Serum IgG, Cytokines, and NK Cell Activity
2.4.4. T-Cell and B-Cell Proliferations in Splenocytes
2.5. Statistical Analysis
3. Results and Discussion
3.1. Functional Compounds Concentration and Antioxidant Activities of OS Extract
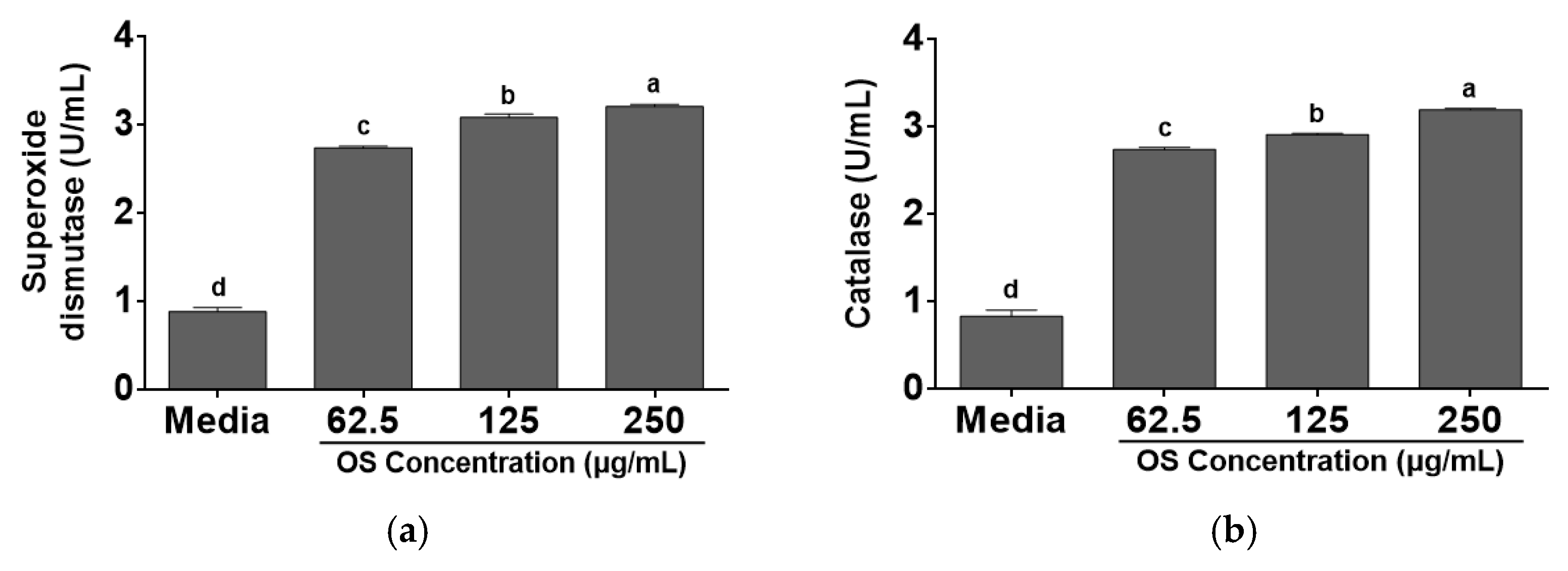
3.2. Effects of OS Extracts on Antioxidant Enzymes in RAW 264.7 Cells
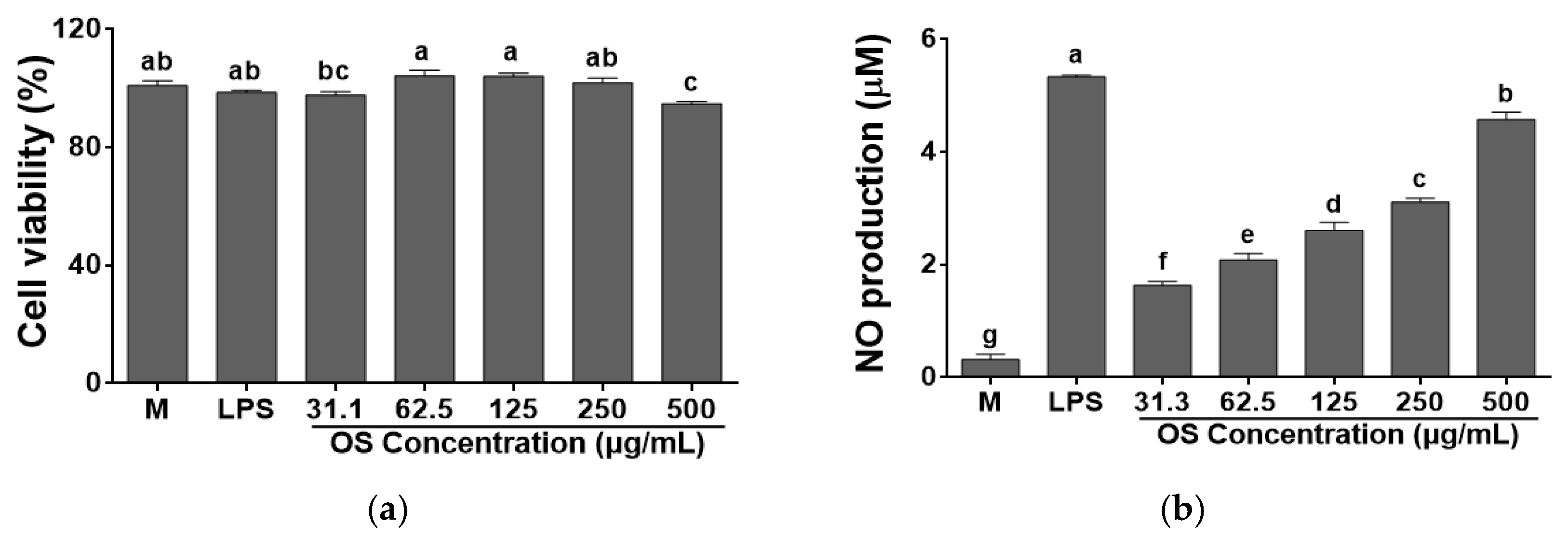
3.3. Effects of OS Extract on Cell Viability and NO Production by RAW 264.7 Cells
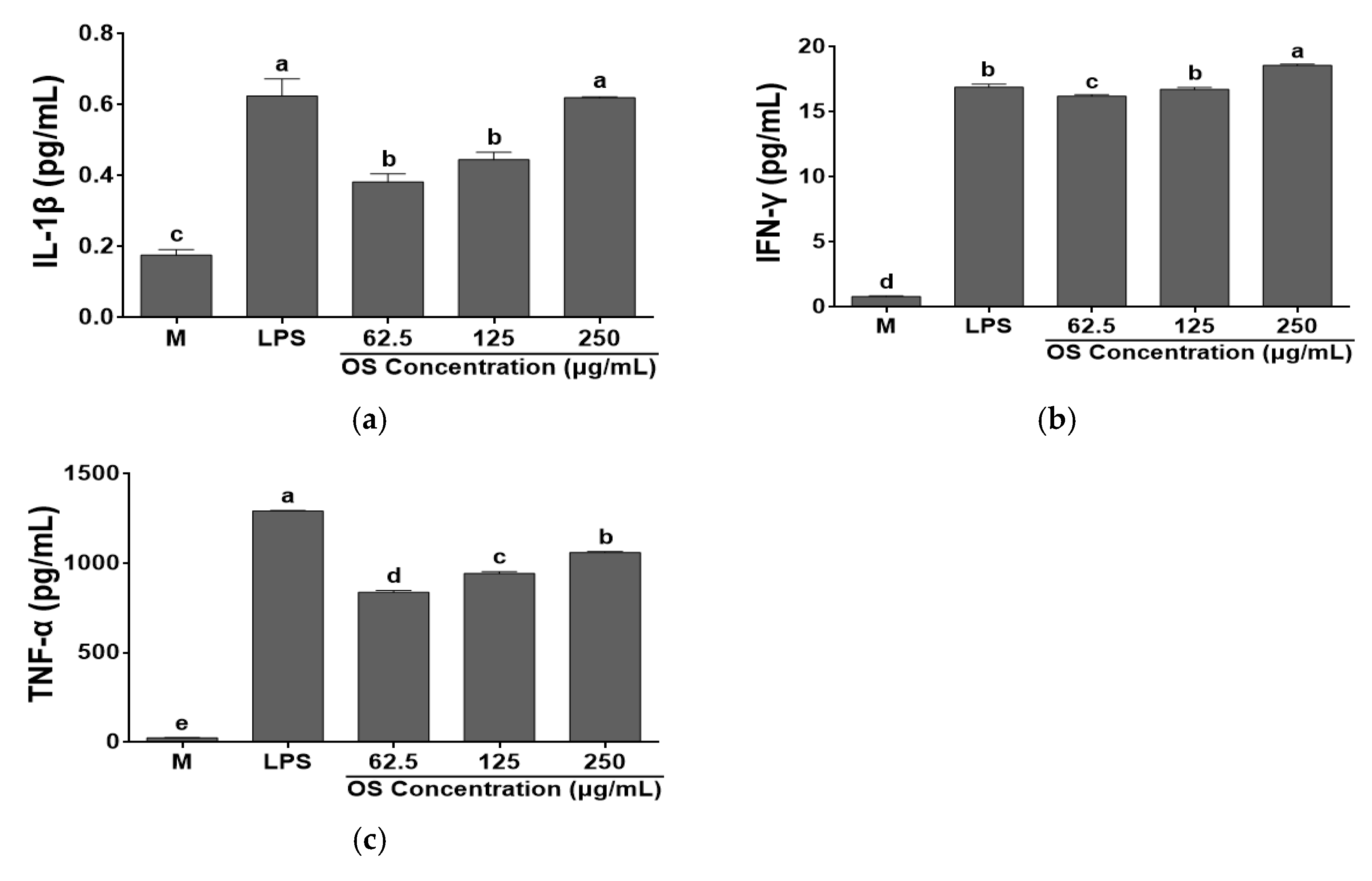
3.4. Effects of OS Extracts on Cytokine Productions by RAW 264.7 Cells
3.5. Effects of OS Extracts on Body Weight Gain and Hematological Factors of the Immunosuppressed Mice
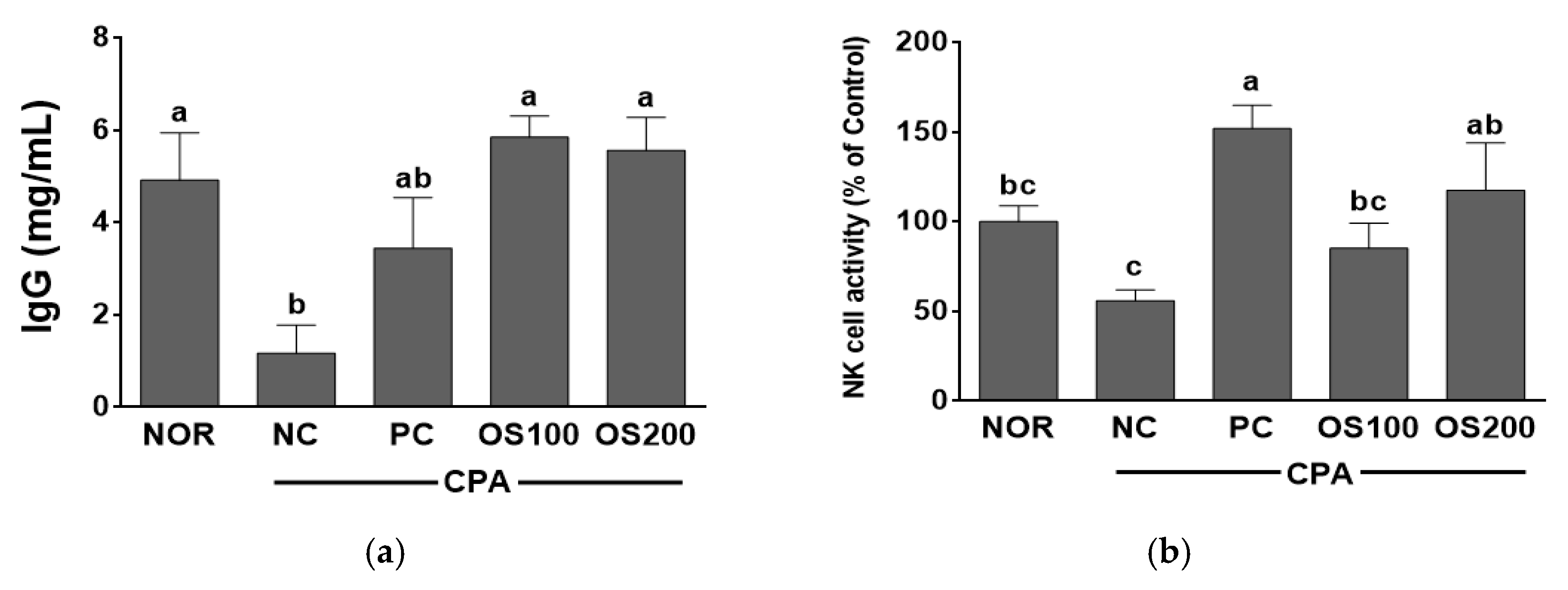
3.6. Effects of OS Extracts on Serum Immunoglobulin and NK Cell Activity in Immunosuppressed Mice
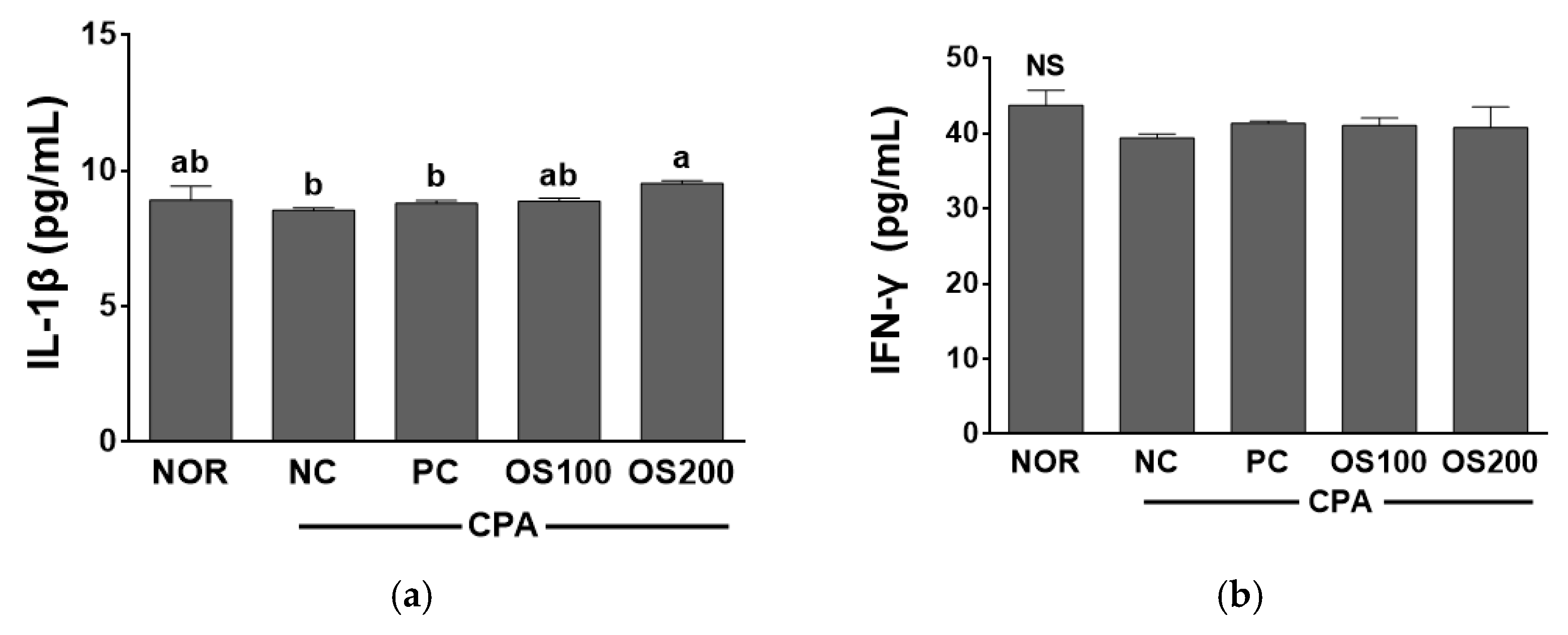
3.7. Effects of OS Extracts on Serum Cytokine Concentration in Immunosuppressed Mice
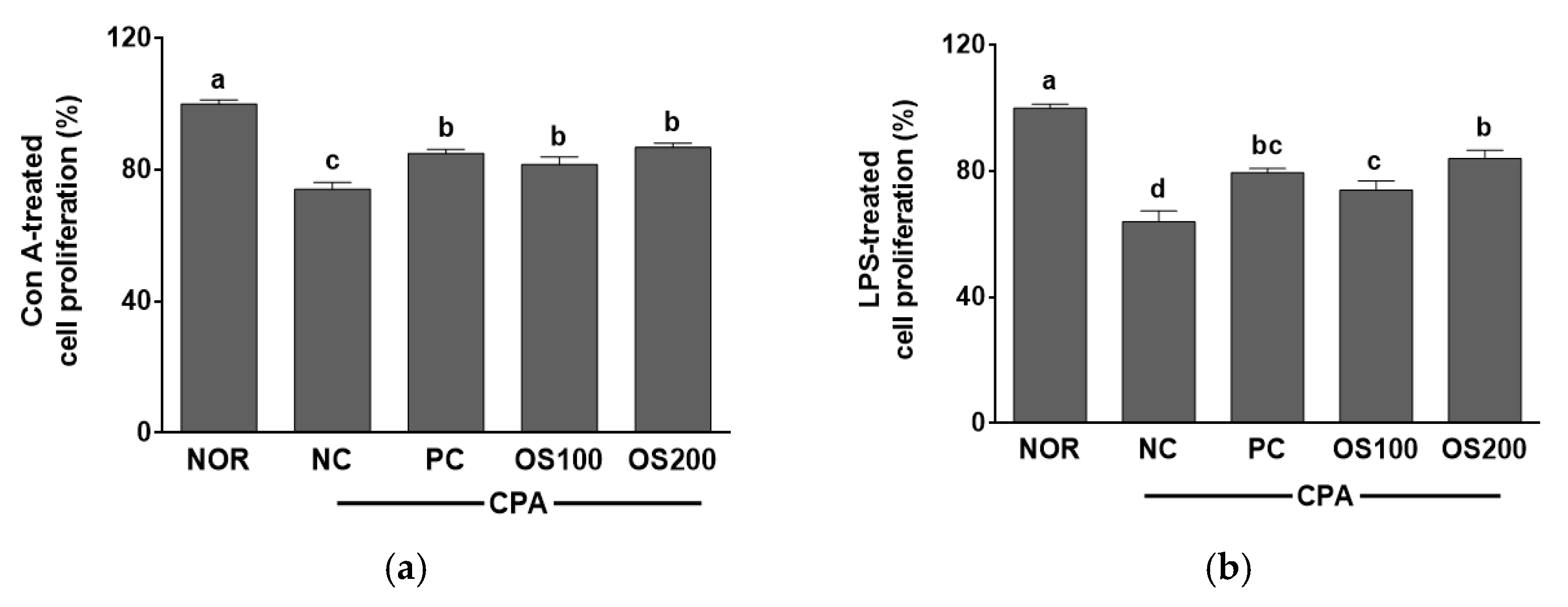
3.8. Effects of OS Extracts on the Proliferation of Mice Splenocytes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bratovic, A. Antioxidant enzymes and their role in preventing cell damage. Act. Sci. Nutr. Health 2020, 4, 1–7. [Google Scholar] [CrossRef]
- Singh, B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Singh, D.P.; Sarma, B.K.; Upadhyay, G.; Singh, H.B. Polyphenolics from various extracts/fractions of red onion (Allium cepa) peel with potent antioxidant and antimutagenic activities. Food Chem. Toxicol. 2009, 47, 1161–1167. [Google Scholar] [CrossRef]
- Lee, C.Y.; Park, H.S.; Kong, D.H.; Kim, Y.K.; Cho, W.J. Immunomodulatory effect of the water extract of Aster tataricus through mitogen-activated protein kinase signaling pathway. J. Nutr. Health 2020, 53, 452–463. [Google Scholar] [CrossRef]
- Hong, S.H.; Ku, J.M.; Kim, H.I.; Ahn, C.W.; Park, S.H.; Seo, H.S.; Shin, Y.C.; Ko, S.G. The immune-enhancing activity of Cervus Nippon manchuricus extract (NGE) in RAW264.7 macrophage cells and immunosuppressed mice. Int. Food Res. J. 2017, 99, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; La, G. The molecular mechanism of natural killer cells function and its importance in cancer immunotherapy. Front. Immunol. 2017, 8, 1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, T.J. Effect of the water extracts from root of Taraxacum officinale on innate and adaptive immune responses in mice. Korean J. Food Nutr. 2008, 21, 275–282. [Google Scholar]
- Nicastro, H.L.; Ross, S.A.; Milner, J.A. Garlic and onions: Their cancer prevention properties. Cancer Prev. Res. 2015, 8, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Tang, G.Y.; Meng, X.; Li, Y.; Zhao, C.N.; Liu, Q.; Li, H.B. Effects of vegetables on cardiovascular diseases and related mechanisms. Nutrients 2017, 9, 857–881. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Noh, S.K. Hypolipidemic effect on onion peel extract in rats exposed to cigarette smoke extract with a high-fat diet. J. Korean Soc. Food Sci. Nutr. 2016, 45, 161–166. [Google Scholar] [CrossRef]
- Park, S.K.; Lee, U.; Kang, J.Y.; Kim, J.M.; Shin, E.J.; Heo, H.J. Ameliorative effect of onion (Allium cepa L.) flesh and peel on amyloid-β-induced cognitive dysfunction via mitochondrial activation. Korean J. Food Sci. Technol. 2020, 52, 263–273. [Google Scholar]
- Chae, M.R.; Kang, S.J.; Lee, K.P.; Choi, B.R.; Kim, H.L.; Park, J.K.; Kim, C.Y.; Lee, S.W. Onion (A. cepa L.) peel extract (OPE) regulates human sperm motility via protein kinase C-mediated activation of the human voltage-gated proton channel. Andrology 2017, 5, 979–989. [Google Scholar] [CrossRef] [Green Version]
- Chia, P.W.; Lim, B.S.; Yong, F.; Poh, S.; Kan, S.Y. An efficient synthesis of bisenols in water extract of waste onion peel ash. Environ. Chem. Lett. 2018, 16, 1493–1499. [Google Scholar] [CrossRef]
- Sagar, N.A.; Pareek, S.; Gonzalez-Aguilar, G.A. Quantification of flavonoids, total phenols and antioxidant properties of onion skin: A comparative study of fifteen Indian cultivars. J. Food Sci. Technol. 2020, 57, 2423–2432. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.K.; Kim, K.B.W.R.; Ahn, N.K.; Choi, Y.U.; Kim, M.; Bark, S.W.; Kim, B.R.; Park, J.H.; Bae, N.Y.; Ahn, D.H. Anti-inflammatory effects of onion (A. cepa) peel hot water extract in vitro and in vivo. KSBB J. 2015, 30, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Ra, K.S.; Suh, H.J.; Chung, S.H.; Son, J.Y. Antioxidant activity of solvent extract from onion skin. Korean J. Food Sci. Technol. 1997, 29, 595–600. [Google Scholar]
- Eloff, J.N. Which extractant should be used for the screening and isolation of antimicrobial components from plants? J. Ethnopharmacol. 1998, 60, 1–8. [Google Scholar] [CrossRef]
- Lee, W.J.; Kang, J.E.; Choi, J.H.; Jeong, S.T.; Kim, M.K.; Choi, H.S. Comparison of the flavonoid and urushiol content in different parts of Rhus verniciflua stokes grown in Wonju and Okcheon. Korean J. Food Sci. Technol. 2015, 47, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Jeong, U.Y.; Jung, J.; Lee, E.B.; Choi, J.H.; Kim, J.S.; Jang, H.H.; Park, S.Y.; Lee, S.H. Antioxidant and immune stimulating effects of Allium hookeri extracts in the RAW 264.7 cells and immune-depressed C57BL/6 mice. Antioxidants 2022, 11, 1927. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; RiceEvans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Alizadeh, S.R.; Ebrahimzadeh, M.A. Quercetin derivatives: Drug design, development, and biological activities, a review. Eur. J. Med. Chem. 2022, 229, 114068. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Kim, K.T.; Kim, H.J.; Chung, M.S.; Chang, P.S.; Park, H.; Paik, H.D. Antioxidant activities of onion (Allium cepa L.) peel extracts produced by ethanol, hot water, and subcritical water extraction. Food Sci. Biotechnol. 2014, 23, 615–621. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Total phenolics, antioxidant and xanthine oxidase inhibitory activity of three colored onions. Front. Life Sci. 2013, 7, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Barbhai, M.D.; Hasan, M.; Dhumal, S.; Singh, S.; Pandiselvam, R.; Rais, N.; Natta, S.; Senapathy, M.; Sinha, N.; et al. Onion (Allium cepa L.) peel: A review on the extraction of bioactive compounds, its antioxidant potential, and its application as a functional food ingredient. J. Food Sci. 2022, 87, 4289–4311. [Google Scholar] [CrossRef]
- Sultana, B.; Anwar, F.; Ashraf, M. Effect of extraction solvent/technique on the antioxidant activity of selected medicinal plant extracts. Molecules 2009, 14, 2167–2180. [Google Scholar] [CrossRef] [Green Version]
- Flieger, J.; Flieger, M. The [DPPH/DPPH-H]-HPLC-DAD method on tracking the antioxidant activity of pure antioxidants and goutweed (Aegopodium podagraria L.) hydroalcoholic extracts. Molecules 2020, 25, 6005. [Google Scholar] [CrossRef]
- Jeong, D.S.; Back, D.A.; Kwon, Y.R.; Kwon, G.M.; Youn, K.S. Quality characteristics and antioxidant activity of onion peel extracts by extraction methods. Korean J. Food Preserv. 2015, 22, 267–274. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, J.S.; Kim, S.H.; Jeong, S.H.; Jeong, U.Y.; Jung, J.E.; Lee, S.K.; Lee, S.H. Antioxidant and anti-inflammatory effects of ethanol extract from whole onion (Allium cepa L.) with leaves. Agriculture 2022, 12, 963. [Google Scholar] [CrossRef]
- Ouyang, H.; Hou, K.; Peng, W.; Liu, Z.; Deng, H. Antioxidant and xanthine oxidase inhibitory activities of total polyphenols from onion. Saudi J. Biol. Sci. 2018, 25, 1509–1513. [Google Scholar] [CrossRef]
- Herrera-Pool, E.; Ramos-Díaz, A.L.; Lizardi-Jiménez, M.A.; RechCohuo, S.; Ayora-Talavera, T.; Cuevas-Bernardino, J.C.; García-Cruz, U.; Pacheco, N. Effect of solvent polarity on the ultrasound assisted extraction and antioxidant activity of phenolic compounds from habanero pepper leaves (Capsicum chinense) and its identification by UPLC-PDA-ESI-MS/MS. Ultrason. Sonochem. 2021, 76, 105658. [Google Scholar] [CrossRef]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP decolorization assay of antioxidant capacity reaction pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marefati, N.; Ghorani, V.; Shakeri, F.; Boskabady, M.; Kianian, F.; Rezaee, R.; Boskabady, M.H. A review of anti-inflammatory, antioxidant, and immunomodulatory effects of Allium cepa and its main constituents. Pharm. Biol. 2021, 59, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Gulcin, I. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markhali, F.S.; Teixerira, J.A.; Rocha, C.M.R. Effect of ohmic heating on the extraction yield, polyphenol content and antioxidant activity of olive mill leaves. Clean Technol. 2022, 4, 512–528. [Google Scholar] [CrossRef]
- Duke, E.J.; Joyce, P.; Ryan, J.P. Characterization of alternative molecular forms of xanthine oxidase in the mouse. Biochem. J. 1973, 131, 187–190. [Google Scholar] [CrossRef] [Green Version]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Ra, K.S.; Bae, S.H.; Son, H.S.; Chung, S.H.; Suh, H.J. Inhibition of xanthine oxidase by flavonols from onion skin. J. Korean Soc. Food Sci. Nutr. 1998, 27, 693–697. [Google Scholar]
- Nile, S.H.; Nile, A.S.; Keum, Y.S.; Sharma, K. Utilization of quercetin and quercetin glycosides from onion (Allium cepa L.) solid waste as an antioxidant, urease and xanthine oxidase inhibitors. Food Chem. 2017, 235, 119–126. [Google Scholar] [CrossRef]
- Ryu, H.W.; Lee, J.H.; Kang, J.E.; Jin, Y.M.; Park, K.H. Inhibition of xanthine oxidase by phenolic phytochemicals from Broussonetia papyrifera. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 587–594. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free Radic. Biol. Med. 2017, 104, 144–164. [Google Scholar] [CrossRef]
- Marefati, N.; Eftekhar, N.; Kaveh, M.; Boskabadi, J.; Beheshti, F.; Boskabady, M.H. The effect of Allium cepa extract on lung oxidant, antioxidant, and immunological biomarkers in ovalbumin-sensitized rats. Med. Princ. Pract. 2018, 27, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Masood, S.; Rehman, A.; Bashir, S.; Shazly, M.E.; Imran, N.; Khalil, P.; Ifthikar, F.; Jaffar, H.M.; Khursheed, T. Investigation of the anti-hyperglycemic and antioxidant effects of wheat bread supplemented with onion peel extract and onion powder in diabetic rats. J. Diabetes Metab. Disord. 2021, 20, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Chernukha, I.; Fedulova, L.; Vasilevskaya, E.; Kulikovskii, A.; Kupaeva, N.; Kotenkova, E. Antioxidant effect of ethanolic onion (Allium cepa) husk extract in ageing rats. Saudi J. Biol. Sci. 2021, 28, 2877–2885. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Wu, J.; Zhao, M.; Sun, W.; Sun, J.; Li, H.; Huang, M. Immunomodulatory activity of a novel polysaccharide extracted from Huangshui on THP-1 cells through NO production and increased IL-6 and TNF-α expression. Food Chem. 2020, 330, 127257. [Google Scholar] [CrossRef]
- Park, K.U.; Kim, J.Y.; Cho, Y.S.; Yee, S.T.; Jeong, C.H.; Kang, K.S.; Seo, K.I. Anticancer and immune-activity of onion Kimchi methanol extract. J. Korean Soc. Food Sci. Nutr. 2004, 33, 1439–1444. [Google Scholar]
- Lee, Y.; Lee, S.H.; Jeong, M.S.; Kim, J.B.; Jang, H.H.; Choe, J.S.; Kim, D.W.; Lillehoj, H.S. In vitro analysis of the immunomodulating effects of Allium hookeri on lymphocytes, macrophages, and tumour cells. J. Poult. Sci. 2017, 54, 142–148. [Google Scholar] [CrossRef] [Green Version]
- García-Ortiz, A.; Serrador, J.M. Nitric oxide signaling in T cell-mediated immunity. Trends Mol. Med. 2018, 24, 412–427. [Google Scholar] [CrossRef]
- Shreshtha, S.; Sharma, P.; Kumar, P.; Sharma, R.; Singh, S. nitric oxide: It’s role in immunity. J. Clin. Diagn. Res. 2018, 12, BE1–BE5. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, Y.S.; Lim, E.M. Immune enhancing effect of Houttuybuae herba on mouse macrophage. Korean J. Obstet. Gynecol. 2012, 25, 12–22. [Google Scholar]
- Cha, J.H.; Kim, Y.S.; Lee, E.M. Effects of Prunellae spica water extract on immune response in macrophage cells. Korean J. Obstet. Gynecol. 2010, 23, 91–100. [Google Scholar]
- Ma, T.; Li, C.; Zhao, F.; Cao, J.; Zhang, X.; Shen, X. Effects of co-fermented collagen peptide-jackfruit juice on the immune response and gut microbiota in immunosuppressed mice. Food Chem. 2021, 365, 130487. [Google Scholar] [CrossRef]
- Ahn, N.K.; Kang, B.K.; Kim, K.B.W.R.; Kim, M.J.; Bae, N.Y.; Park, J.H.; Park, S.H.; Ahn, D.H. Anti-inflammatory effect of ethanol extract from onion (Allium cepa L.) peel on lipopolysaccharide-induced inflammatory responses in RAW 264.7 cells and mice ears. J. Korean Soc. Food Sci. 2015, 44, 1612–1620. [Google Scholar] [CrossRef]
- Ueda, H.; Takeuchi, A.; Wako, T. Activation of immune response in mice by an oral administration of bunching onion (Allium fistulosum) mucus. Biosci. Biotechnol. Biochem. 2013, 77, 1809–1813. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.H.; Lee, Y.S.; Ku, S.K.; Lee, H.J. Phellinus baumii enhances the immune response in cyclophosphamide-induced immunosuppressed mice. Nutr. Res. 2020, 75, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Kumar, W.P.; Venkatesh, Y.P. Alleviation of cyclophosphamide-induced immunosuppression in Wistar rats by onion lectin (Allium cepa agglutinin). J. Ethnopharmacol. 2016, 186, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Mo, S.; Shen, M.; Chen, Y.; Yu, Q.; Li, Z.; Xie, J. Sulfated modification enhances the immunomodulatory effect of Cyclocarya paliurus polysaccharide on cyclophosphamide-induced immunosuppressed mice through MyD88-dependent MAPK/NF-κB and PI3K-Akt signaling pathways. Int. Food Res. J. 2021, 150, 110756. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, X.; Yi, R.; Li, G.; Sun, P.; Qian, Y.; Zhao, X. Immunomodulatory effect on tremella polysaccharides aginst cyclophosphamide-induced immunosuppression in mice. Molecules 2018, 23, 239. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, M.; Chen, J.; Tang, Y.; Dou, J.; Yu, J.; Xi, T.; Zhou, C. A polysaccharide from Strongylocentrotus nudus eggs protects against myelosuppression and immunosuppression in cyclophosphamide-treated mice. Int. Immunopharmacol. 2011, 11, 1946–1953. [Google Scholar] [CrossRef]
- Park, H.Y.; Oh, M.J.; Choi, I.W. Immunomodulatory activities of Corchorus olitorius leaf extract: Beneficial effects in macrophage and NK cell activation immunosuppressed mice. J. Funct. Foods 2018, 46, 220–226. [Google Scholar] [CrossRef]
- Choi, J.H.; Kim, S.H.; Lee, E.B.; Kim, J.S.; Jung, J.E.; Jeong, U.Y.; Lee, S.H. Anti-diabetic effects of Allium hookeri extracts prepared by different methods in type 2 C57BL/J-db/db mice. Pharmaceuticals 2022, 15, 486. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, V.K.; Venkatesh, Y.P. Characterization of onion lectin (Allium cepa agglutinin) as an immunomodulatory protein including Th1-type immune response in vitro. Int. Immunopharmacol. 2015, 26, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y. A study on the biological activity of Allium cepa extract in vivo. J. Life Sci. 2020, 30, 267–277. [Google Scholar]
- Zheng, Y.; Guan, J.; Wang, L.; Luo, X.; Zhang, X. Comparative proteomic analysis of spleen reveals key immune-related proteins in the yak (Bos grunniens) at different growth stages. Comp. Biochem. Physiol. Part D Genom. Proteom. 2022, 42, 100968. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | |||
|---|---|---|---|
| Instrument | Agilent 1260 Infinity Binary LC | ||
| Column | Zorbax Eclipse Plus C18 UG 120 (4.6 × 250 mm, 5 μm) | ||
| Mobile phase | Mobile phase A: 5% formic acid Mobile phase B: MeOH | ||
| Time (min) | A (%) | B (%) | |
| 0 | 100 | 0 | |
| 25 | 40 | 60 | |
| 26 | 0 | 100 | |
| 30 | 40 | 60 | |
| 31 | 80 | 20 | |
| 37 | 80 | 20 | |
| Gradient program | Gradient elution of mobile phase A and B | ||
| UV length | 360 nm | ||
| Injection volume | 10 μL | ||
| Column temperature | 40 °C | ||
| Flow rate Run time | 0.8 mL/min 37 min | ||
| Sample | Quercetin (mg/g) | Total Phenolic Content (mg GAE/g) |
|---|---|---|
| OS extract | 37.9 ± 0.6 | 65.6 ± 0.1 |
| Sample | Concentration (μg/mL) | DPPH Radical Scavenging Activity (%) | ABTS Radical Scavenging Activity (%) |
|---|---|---|---|
| OS extract | 62.5 | 14.78 ± 0.44 f | 76.85 ± 0.23 e |
| 125 | 18.67 ± 0.26 e | 90.78 ± 0.12 d | |
| 250 | 26.53 ± 0.46 d | 95.90 ± 0.09 c | |
| 500 | 36.19 ± 0.93 c | 98.82 ± 0.06 b | |
| 1000 | 46.89 ± 0.94 b | 99.68 ± 0.06 a | |
| 2000 | 68.07 ± 0.99 a | 99.68 ± 0.07 a | |
| IC50 (1) | 945.7 ± 32.3 | 26.5 ± 0.2 |
| Sample | Concentration (μg/mL) | Xanthine Oxidase Inhibitory Activity (%) |
|---|---|---|
| OS extract | 6.3 | 32.20 ± 0.87 d |
| 12.5 | 53.18 ± 0.34 c | |
| 25 | 75.70 ± 0.29 b | |
| 50 | 89.74 ± 0.70 a | |
| 100 | 90.66 ± 0.76 a | |
| IC50 (1) | 10.7 ± 0.2 |
| Group | NOR (1) | NC | PC | OS100 | OS200 | |
|---|---|---|---|---|---|---|
| Body weight gain (g/mice) Red blood cell (RBC, ×106 cells/μL) | 2.74 ± 0.39 a 10.2 ± 0.2 ab | 0.98 ± 0.26 b 9.98 ± 0.10 ab | 1.28 ± 0.26 b 9.97 ± 0.13 ab | 1.30 ± 0.21 b 9.85 ± 0.16 b | 1.93 ± 0.21 ab 10.3 ± 0.1 a | |
| Hemoglobin (Hb, g/dL) | 14.9 ± 0.3 NS | 14.4 ± 0.2 | 14.6 ± 0.2 | 14.3 ± 0.2 | 14.8 ± 0.1 | |
| RBC indexes | MCV (fL) (2) | 49.5 ± 0.6 b | 48.7 ± 0.5 b | 49.5 ± 0.5 bc | 49.6 ± 0.2 b | 51.6 ± 0.5 a |
| MCH (pg) | 14.6 ± 0.0 ab | 14.5 ± 0.0 abc | 14.6 ± 0.1 a | 14.3 ± 0.1 c | 14.41 ± 0.0 bc | |
| MCHC (g/dL) | 29.8 ± 0.3 a | 29.6 ± 0.3 ab | 29.0 ± 0.3 abc | 28.8 ± 0.2 bc | 28.5 ± 0.3 c | |
| Hematocrit (%) | 50.3 ± 1.1 b | 47.8 ± 0.6 c | 49.7 ± 0.4 bc | 49.3 ± 0.5 bc | 52.4 ± 0.3 a | |
| Platelet (×10 3 cells/μL) | 1226 ± 44 NS | 907.9 ± 104.8 | 993.8 ± 116.8 | 1010 ± 97 | 1026 ± 107 | |
| White blood cell (WBC, ×103 cells/μL) | 4.40 ± 0.33 a | 2.36 ± 0.43 c | 3.05 ± 0.44 bc | 3.29 ± 0.29 b | 4.37 ± 0.21 a | |
| WBC differential counting (%) | Neutrophil | 7.53 ± 0.07 b | 19.8 ± 0.8 a | 15.6 ± 0.4 a | 15.4 ± 0.9 a | 18.7 ± 2.7 a |
| Lymphocyte | 83.1 ± 0.8 a | 68.7 ± 2.5 b | 74.8 ± 2.0 b | 75.1 ± 1.7 b | 73.9 ± 2.0 b | |
| Monocyte | 3.67 ± 0.52 NS | 3.78 ± 0.31 | 3.98 ± 0.41 | 4.18 ± 0.63 | 3.88 ± 0.55 | |
| Eosinophil | 7.43 ± 0.74 a | 1.73 ± 0.67 b | 5.30 ± 0.86 a | 7.47 ± 1.48 a | 5.27 ± 1.64 a | |
| Basophil | 0.12 ± 0.05 NS | 0.00 ± 0.00 | 0.08 ± 0.05 | 0.12 ± 0.07 | 0.10 ± 0.04 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-S.; Lee, E.-B.; Choi, J.-H.; Jung, J.; Jeong, U.-Y.; Bae, U.-J.; Jang, H.-H.; Park, S.-Y.; Cha, Y.-S.; Lee, S.-H. Antioxidant and Immune Stimulating Effects of Allium cepa Skin in the RAW 264.7 Cells and in the C57BL/6 Mouse Immunosuppressed by Cyclophosphamide. Antioxidants 2023, 12, 892. https://doi.org/10.3390/antiox12040892
Kim J-S, Lee E-B, Choi J-H, Jung J, Jeong U-Y, Bae U-J, Jang H-H, Park S-Y, Cha Y-S, Lee S-H. Antioxidant and Immune Stimulating Effects of Allium cepa Skin in the RAW 264.7 Cells and in the C57BL/6 Mouse Immunosuppressed by Cyclophosphamide. Antioxidants. 2023; 12(4):892. https://doi.org/10.3390/antiox12040892
Chicago/Turabian StyleKim, Ji-Su, Eun-Byeol Lee, Ji-Hye Choi, Jieun Jung, Un-Yul Jeong, Ui-Jin Bae, Hwan-Hee Jang, Shin-Young Park, Youn-Soo Cha, and Sung-Hyen Lee. 2023. "Antioxidant and Immune Stimulating Effects of Allium cepa Skin in the RAW 264.7 Cells and in the C57BL/6 Mouse Immunosuppressed by Cyclophosphamide" Antioxidants 12, no. 4: 892. https://doi.org/10.3390/antiox12040892