
Deubiquitylase OTUD3 Mediates Endoplasmic Reticulum Stress through Regulating Fortilin Stability to Restrain Dopaminergic Neurons Apoptosis
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plasmids and Viruses
2.2. Cell Culture and Transfection
2.3. Cell Viability Assessment
2.4. Trypan Blue Assay
2.5. Propidium Iodide (PI) Assay
2.6. Measurement of Intracellular MDA Levels
2.7. Flow Cytometric Measurement of Apoptosis
2.8. Animals and In Vivo Treatment
2.9. RT-PCR and Quantitative PCR
2.10. Transmission Electron Microscopy Measurement of Changes in Endoplasmic Reticulum
2.11. Western Blotting and Immunoprecipitation
2.12. In Vitro Ubiquitin Conjugation Assay
2.13. Immunofluorescence Staining
2.14. Regents
2.15. Statistical Analysis
3. Results
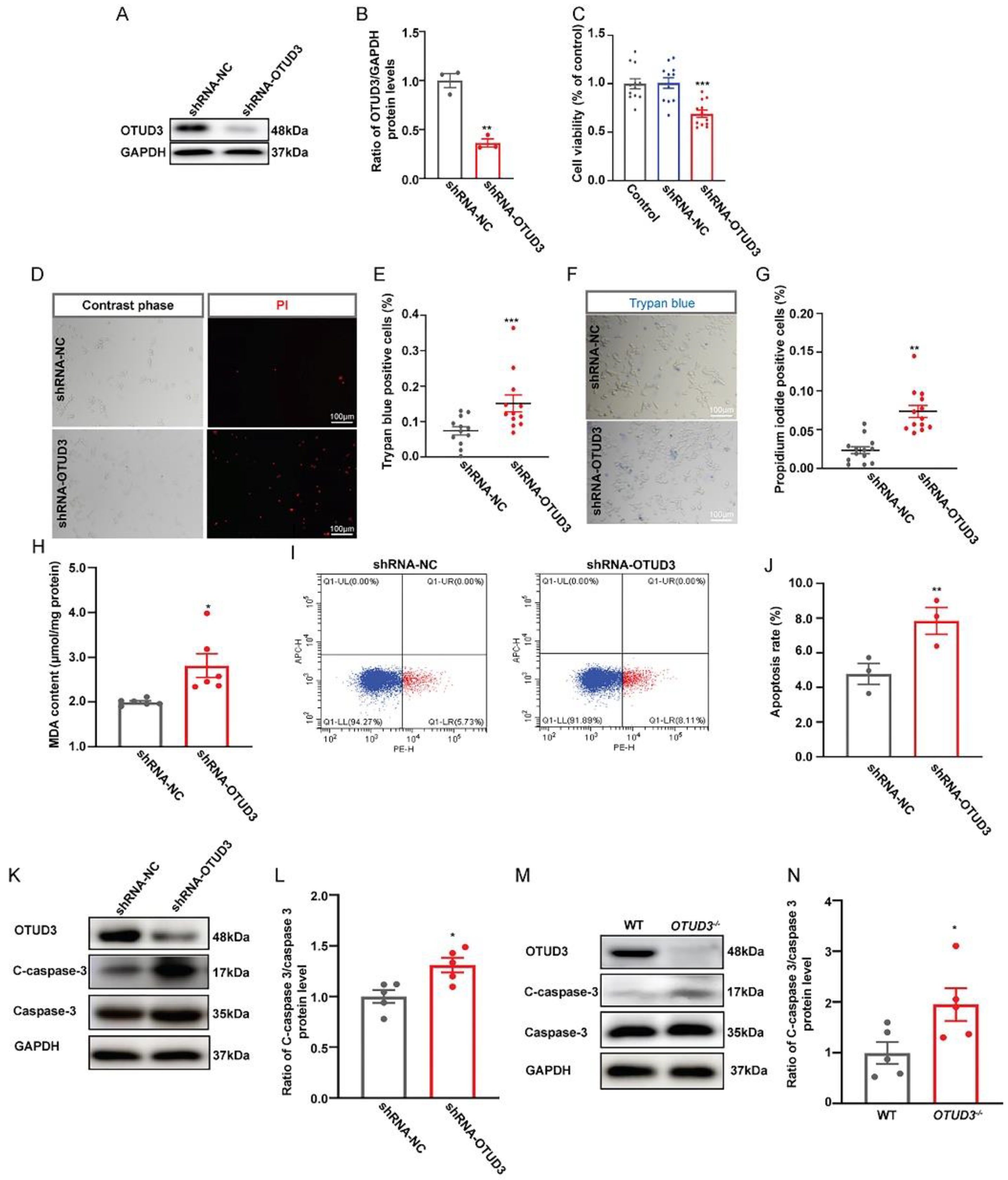
3.1. OTUD3 Deletion Induced Neuronal Apoptosis
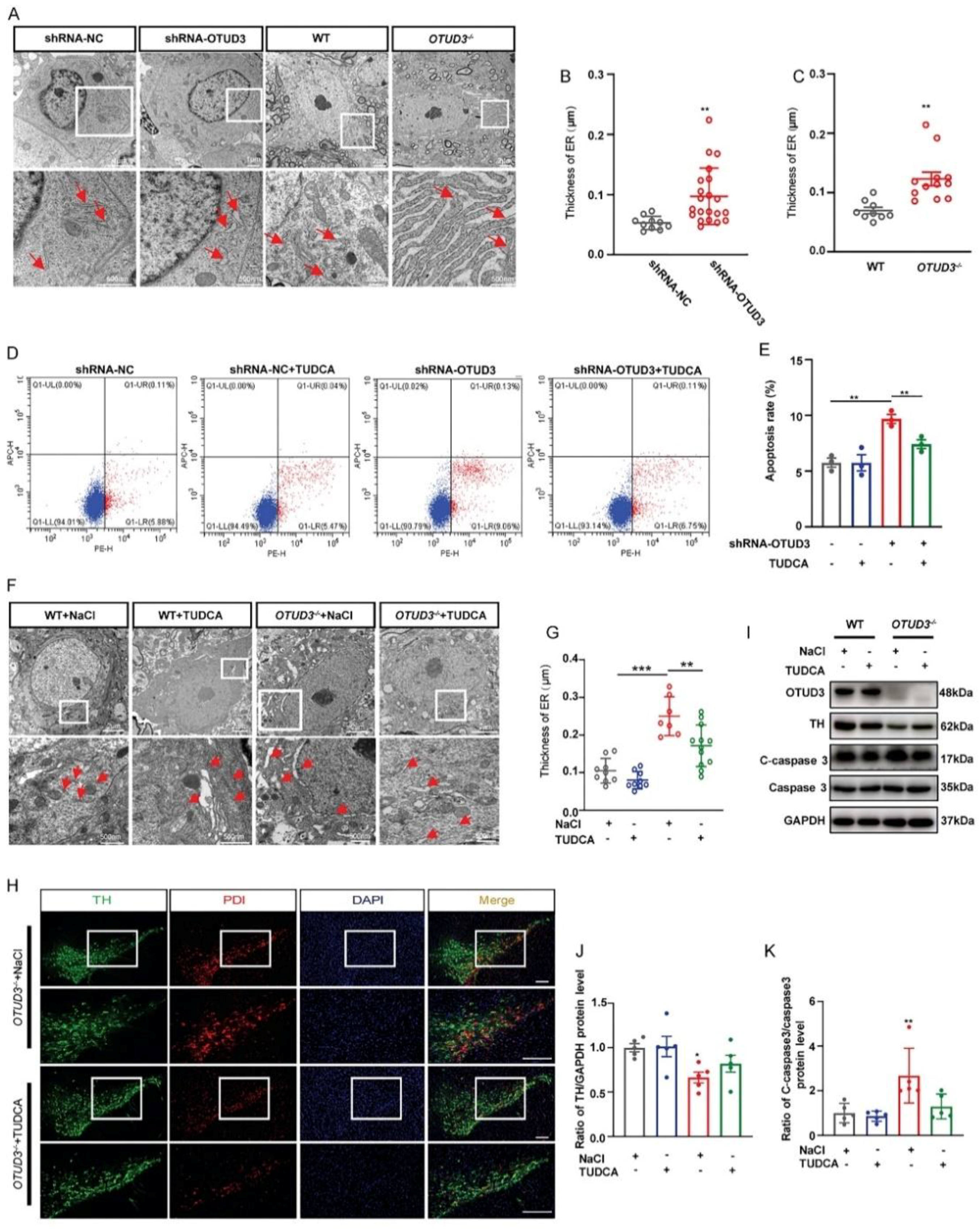
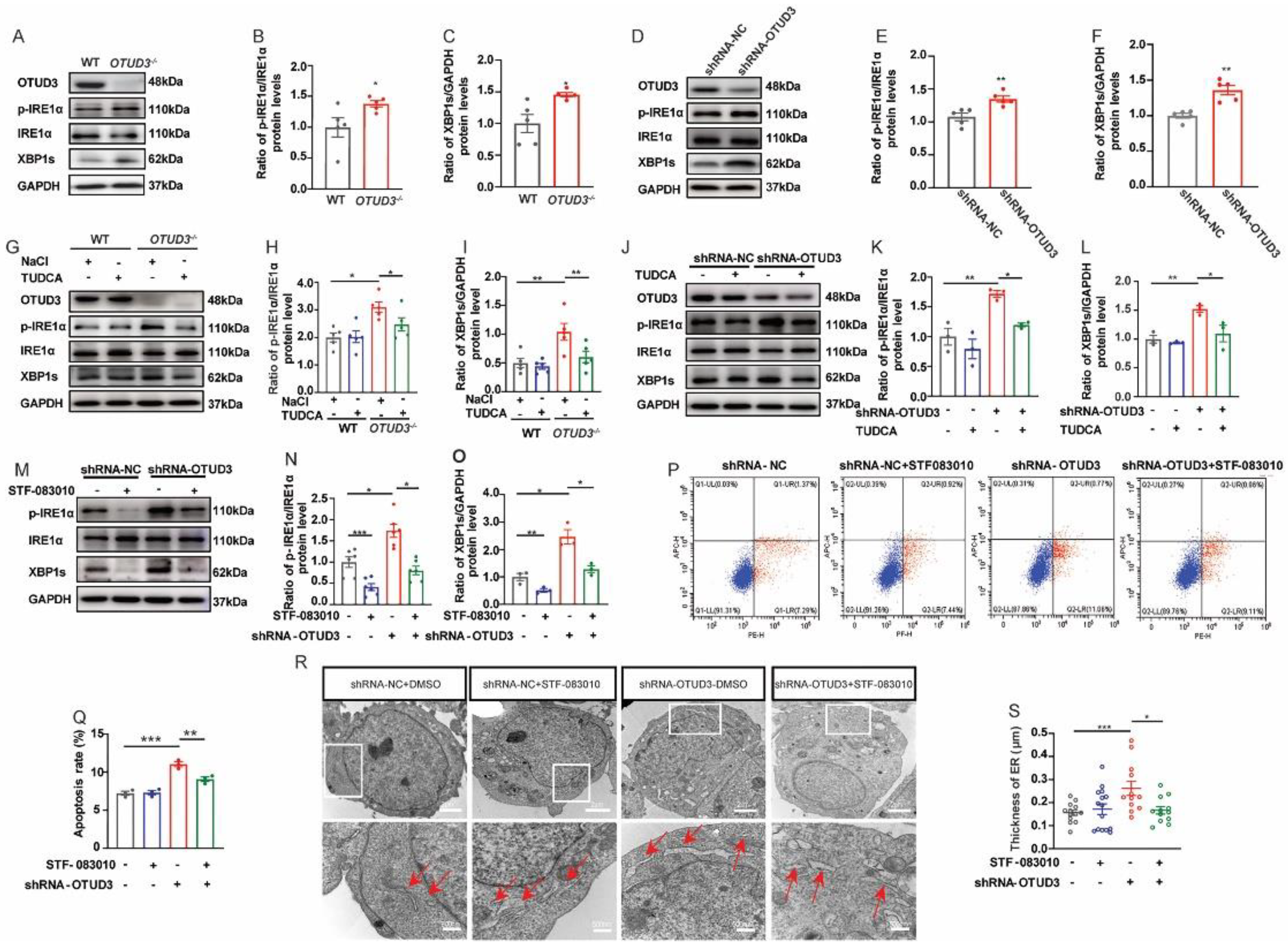
3.2. OTUD3 Knockdown Induced ER Stress
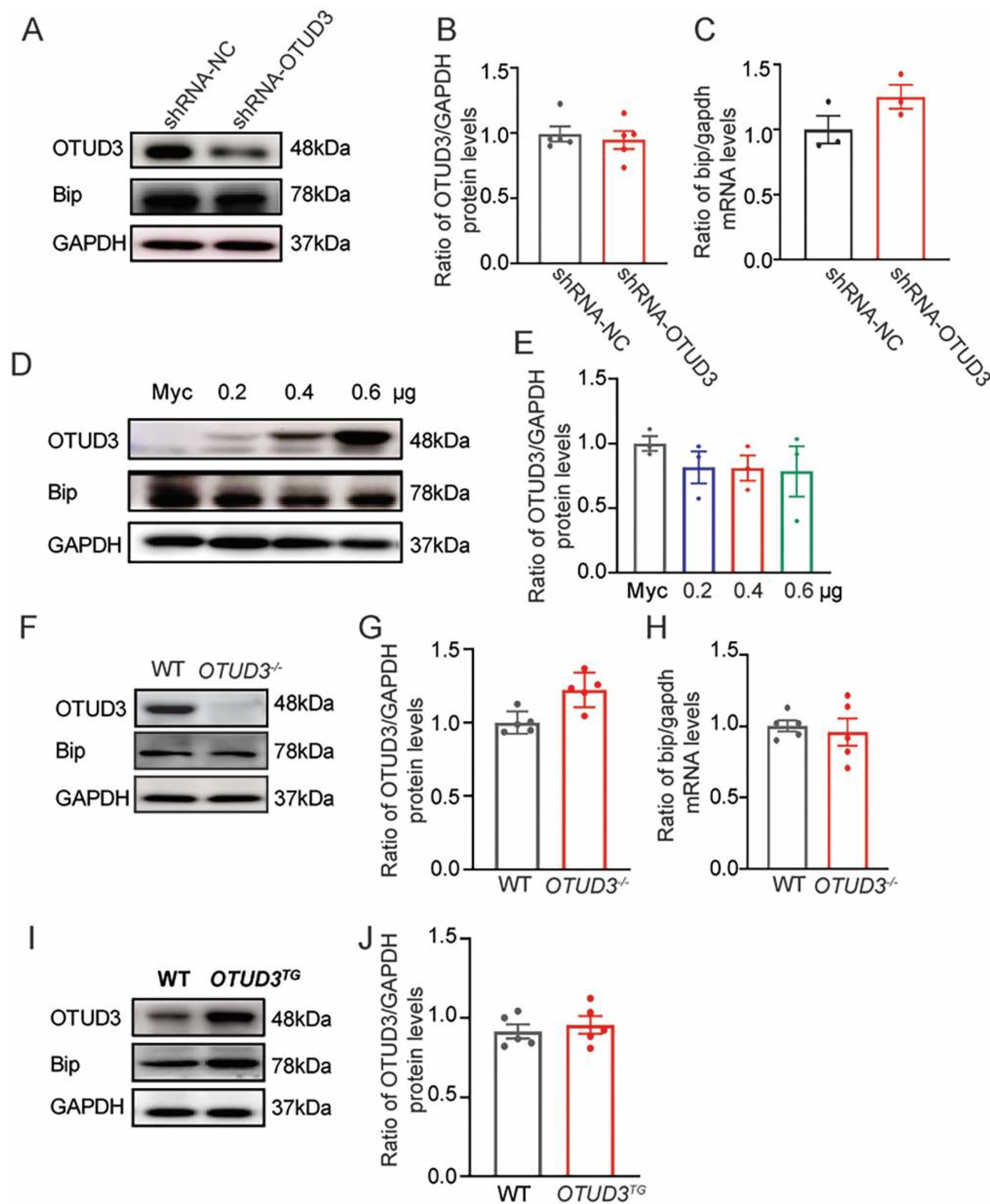
3.3. The Expression of Bip Was Not Changed In Vivo and In Vitro
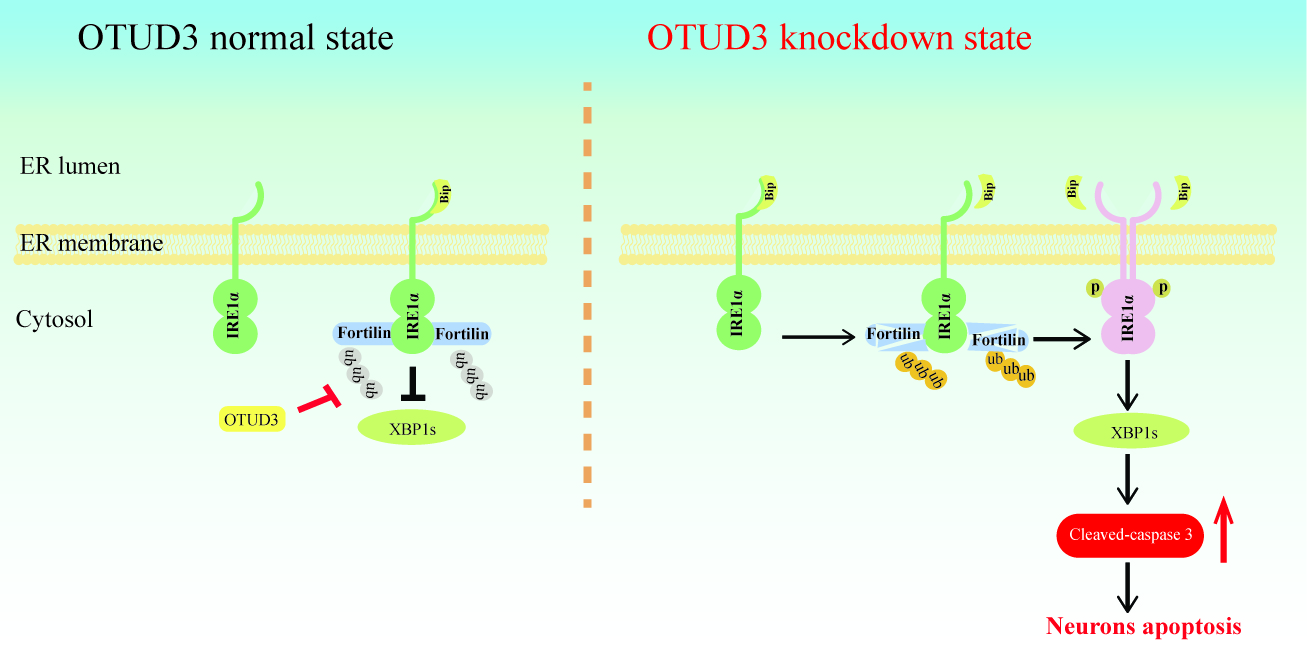
3.4. OTUD3 Knockdown Induced ER Stress via Activating IRE1α Pathway
3.5. OTUD3 Was Involved in ER Stress by Regulating the Ubiquitination Level of IRE1α Binding Protein Fortilin
3.6. Fortilin Alleviated OTUD3 Knockdown Induced ER Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tolosa, E.; Garrido, A. Challenges in the diagnosis of Parkinson’s disease. Lancet Neurol. 2021, 20, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Bloem, B.R.; Okun, M.S. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef] [PubMed]
- Mahoney-Sánchez, L.; Bouchaoui, H. Ferroptosis and its potential role in the physiopathology of Parkinson’s Disease. Prog. Neurobiol. 2021, 196, 101890. [Google Scholar] [CrossRef]
- Bi, M.; Du, X. Deficient immunoproteasome assembly drives gain of α-synuclein pathology in Parkinson’s disease. Redox Biol. 2021, 47, 102167. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Li, H. Deubiquitylase OTUD3 prevents Parkinson’s disease through stabilizing iron regulatory protein 2. Cell Death Dis. 2022, 13, 418. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Hong, M. Genome-wide association study of ulcerative colitis in Koreans suggests extensive overlapping of genetic susceptibility with Caucasians. Inflamm. Bowel Dis. 2013, 19, 954–966. [Google Scholar] [CrossRef]
- Garshott, D.M.; Sundaramoorthy, E. Distinct regulatory ribosomal ubiquitylation events are reversible and hierarchically organized. Elife 2020, 9, e54023. [Google Scholar] [CrossRef]
- Zhang, Z.; Fang, X. Acetylation-Dependent Deubiquitinase OTUD3 Controls MAVS Activation in Innate Antiviral Immunity. Mol. Cell 2020, 79, 304–319.e7. [Google Scholar] [CrossRef]
- Fagone, P.; Jackowski, S. Membrane phospholipid synthesis and endoplasmic reticulum function. J. Lipid Res. 2009, 50, S311–S316. [Google Scholar] [CrossRef] [Green Version]
- van Anken, E.; Braakman, I. Versatility of the endoplasmic reticulum protein folding factory. Crit. Rev. Biochem. Mol. Biol. 2005, 40, 191–228. [Google Scholar] [CrossRef]
- Pobre, K.F.R.; Poet, G.J. The endoplasmic reticulum (ER) chaperone BiP is a master regulator of ER functions: Getting by with a little help from ERdj friends. J. Biol. Chem. 2019, 294, 2098–2108. [Google Scholar] [CrossRef] [Green Version]
- Hendershot, L.M.; Valentine, V.A. Localization of the gene encoding human BiP/GRP78, the endoplasmic reticulum cognate of the HSP70 family, to chromosome 9q34. Genomics 1994, 20, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Haas, I.G. BiP—A heat shock protein involved in immunoglobulin chain assembly. Curr. Top. Microbiol. Immunol. 1991, 167, 71–82. [Google Scholar] [PubMed]
- Gething, M.J.; Sambrook, J. Protein folding in the cell. Nature 1992, 355, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Little, E.; Ramakrishnan, M. The glucose-regulated proteins (GRP78 and GRP94): Functions, gene regulation, and applications. Crit. Rev. Eukaryot. Gene Expr. 1994, 4, 1–18. [Google Scholar] [CrossRef]
- Li, F.; Zhang, D. Characterization of fortilin, a novel antiapoptotic protein. J. Biol. Chem. 2001, 276, 47542–47549. [Google Scholar] [CrossRef] [Green Version]
- Susini, L.; Besse, S. TCTP protects from apoptotic cell death by antagonizing bax function. Cell Death Differ. 2008, 15, 1211–1220. [Google Scholar] [CrossRef] [Green Version]
- Cans, C.; Passer, B.J. Translationally controlled tumor protein acts as a guanine nucleotide dissociation inhibitor on the translation elongation factor eEF1A. Proc. Natl. Acad. Sci. USA 2003, 100, 13892–13897. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Yao, L. TCTP promotes glioma cell proliferation in vitro and in vivo via enhanced β-catenin/TCF-4 transcription. Neuro-Oncology 2014, 16, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Qin, Y. Serum translationally controlled tumor protein is involved in rat liver regeneration after hepatectomy. Hepatol. Res. 2016, 46, 1392–1401. [Google Scholar] [CrossRef]
- Pinkaew, D.; Chattopadhyay, A. Fortilin binds IRE1α and prevents ER stress from signaling apoptotic cell death. Nat. Commun. 2017, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Jeong, M.; Jeong, M.H. TCTP protein degradation by targeting mTORC1 and signaling through S6K, Akt, and Plk1 sensitizes lung cancer cells to DNA-damaging drugs. Sci. Rep. 2021, 11, 20812. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.Y.; Wang, H.J. miR-27b-regulated TCTP as a novel plasma biomarker for oral cancer: From quantitative proteomics to post-transcriptional study. J. Proteomics 2012, 77, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Yoshitake, Y.; Shinozaki, D. Autophagy triggered by iron-mediated ER stress is an important stress response to the early phase of Pi starvation in plants. Plant J. 2022, 110, 1370–1381. [Google Scholar] [CrossRef]
- da Silva, D.C.; Valentão, P. Endoplasmic reticulum stress signaling in cancer and neurodegenerative disorders: Tools and strategies to understand its complexity. Pharmacol. Res. 2020, 155, 104702. [Google Scholar] [CrossRef]
- Hoozemans, J.J.; van Haastert, E.S. Activation of the unfolded protein response in Parkinson’s disease. Biochem. Biophys. Res. Commun. 2007, 354, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Ryu, E.J.; Harding, H.P. Endoplasmic reticulum stress and the unfolded protein response in cellular models of Parkinson’s disease. J. Neurosci. 2002, 22, 10690–10698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtonen, Š.; Jaronen, M. Inhibition of Excessive Oxidative Protein Folding Is Protective in MPP(+) Toxicity-Induced Parkinson’s Disease Models. Antioxid. Redox Signal. 2016, 25, 485–497. [Google Scholar] [CrossRef]
- Holtz, W.A.; O’Malley, K.L. Parkinsonian mimetics induce aspects of unfolded protein response in death of dopaminergic neurons. J. Biol. Chem. 2003, 278, 19367–19377. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.W.; Yen, J.H. Luteolin modulates 6-hydroxydopamine-induced transcriptional changes of stress response pathways in PC12 cells. PLoS ONE 2014, 9, e97880. [Google Scholar] [CrossRef]
- Devi, L.; Ohno, M. PERK mediates eIF2α phosphorylation responsible for BACE1 elevation, CREB dysfunction and neurodegeneration in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2014, 35, 2272–2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pytel, D.; Majsterek, I. Tumor progression and the different faces of the PERK kinase. Oncogene 2016, 35, 1207–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Alam, G.N. The unfolded protein response induces the angiogenic switch in human tumor cells through the PERK/ATF4 pathway. Cancer Res. 2012, 72, 5396–5406. [Google Scholar] [CrossRef] [Green Version]
- Pillai, S. Birth pangs: The stressful origins of lymphocytes. J. Clin. Investig. 2005, 115, 224–227. [Google Scholar] [CrossRef]
- Endres, K.; Reinhardt, S. ER-stress in Alzheimer’s disease: Turning the scale? Am. J. Neurodegener. Dis. 2013, 2, 247–265. [Google Scholar] [PubMed]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Shaffer, A.L.; Shapiro-Shelef, M. XBP1, downstream of Blimp-1, expands the secretory apparatus and other organelles, and increases protein synthesis in plasma cell differentiation. Immunity 2004, 21, 81–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.H.; Chu, G.C. XBP-1 is required for biogenesis of cellular secretory machinery of exocrine glands. Embo J. 2005, 24, 4368–4380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xicoy, H.; Wieringa, B. The SH-SY5Y cell line in Parkinson’s disease research: A systematic review. Mol. Neurodegener. 2017, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Li, H. The deubiquitylase OTUD3 stabilizes GRP78 and promotes lung tumorigenesis. Nat. Commun. 2019, 10, 2914. [Google Scholar] [CrossRef] [Green Version]
- Pu, Q.; Lv, Y.R. Tumor suppressor OTUD3 induces growth inhibition and apoptosis by directly deubiquitinating and stabilizing p53 in invasive breast carcinoma cells. BMC Cancer 2020, 20, 583. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Lv, Y. Deubiquitylase OTUD3 regulates PTEN stability and suppresses tumorigenesis. Nat. Cell Biol. 2015, 17, 1169–1181. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wei, L. Integration of Hippo signalling and the unfolded protein response to restrain liver overgrowth and tumorigenesis. Nat. Commun. 2015, 6, 6239. [Google Scholar] [CrossRef] [Green Version]
- Hetzer, S.M.; Guilhaume-Correa, F. Traumatic Optic Neuropathy Is Associated with Visual Impairment, Neurodegeneration, and Endoplasmic Reticulum Stress in Adolescent Mice. Cells 2021, 10, 996. [Google Scholar] [CrossRef]
- Xie, P.; Chen, Y. The deubiquitinase OTUD3 stabilizes ACTN4 to drive growth and metastasis of hepatocellular carcinoma. Aging (Albany NY) 2021, 13, 19317–19338. [Google Scholar] [CrossRef]
- Hetz, C.; Zhang, K. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Papa, F.R. The Unfolded Protein Response and Cell Fate Control. Mol. Cell 2018, 69, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Hetz, C.; Martinon, F. The unfolded protein response: Integrating stress signals through the stress sensor IRE1α. Physiol. Rev. 2011, 91, 1219–1243. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Y. Microglia-derived IL-1β promoted neuronal apoptosis through ER stress-mediated signaling pathway PERK/eIF2α/ATF4/CHOP upon arsenic exposure. J. Hazard. Mater. 2021, 417, 125997. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhuang, Y. Endoplasmic reticulum stress aggravates copper-induced apoptosis via the PERK/ATF4/CHOP signaling pathway in duck renal tubular epithelial cells. Environ. Pollut. 2021, 272, 115981. [Google Scholar] [CrossRef]
- Merksamer, P.I.; Papa, F.R. The UPR and cell fate at a glance. J. Cell Sci. 2010, 123 Pt 7, 1003–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Fujita, T. Physical and functional antagonism between tumor suppressor protein p53 and fortilin, an anti-apoptotic protein. J. Biol. Chem. 2011, 286, 32575–32585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Li, F. Physical and functional interaction between myeloid cell leukemia 1 protein (MCL1) and Fortilin. The potential role of MCL1 as a fortilin chaperone. J. Biol. Chem. 2002, 277, 37430–37438. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Huan, X.; Jia, F.; Zhang, Z.; Bi, M.; Fu, L.; Du, X.; Chen, X.; Yan, C.; Jiao, Q.; et al. Deubiquitylase OTUD3 Mediates Endoplasmic Reticulum Stress through Regulating Fortilin Stability to Restrain Dopaminergic Neurons Apoptosis. Antioxidants 2023, 12, 809. https://doi.org/10.3390/antiox12040809
Chen L, Huan X, Jia F, Zhang Z, Bi M, Fu L, Du X, Chen X, Yan C, Jiao Q, et al. Deubiquitylase OTUD3 Mediates Endoplasmic Reticulum Stress through Regulating Fortilin Stability to Restrain Dopaminergic Neurons Apoptosis. Antioxidants. 2023; 12(4):809. https://doi.org/10.3390/antiox12040809
Chicago/Turabian StyleChen, Ling, Xuejie Huan, Fengju Jia, Zhen Zhang, Mingxia Bi, Lin Fu, Xixun Du, Xi Chen, Chunling Yan, Qian Jiao, and et al. 2023. "Deubiquitylase OTUD3 Mediates Endoplasmic Reticulum Stress through Regulating Fortilin Stability to Restrain Dopaminergic Neurons Apoptosis" Antioxidants 12, no. 4: 809. https://doi.org/10.3390/antiox12040809