
Differential Apoptotic Effects of Bee Product Mixtures on Normal and Cancer Hepatic Cells


 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Samples
2.3. Cell Culture
2.4. MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide) Assay
2.5. Cell Cycle Analysis
2.6. TdT-dUTP Terminal Nick-End Labeling (TUNEL) Assay
2.7. Assay for Apoptotic Pathways Identification
2.8. Total Extracts and Caspase Activity Assay
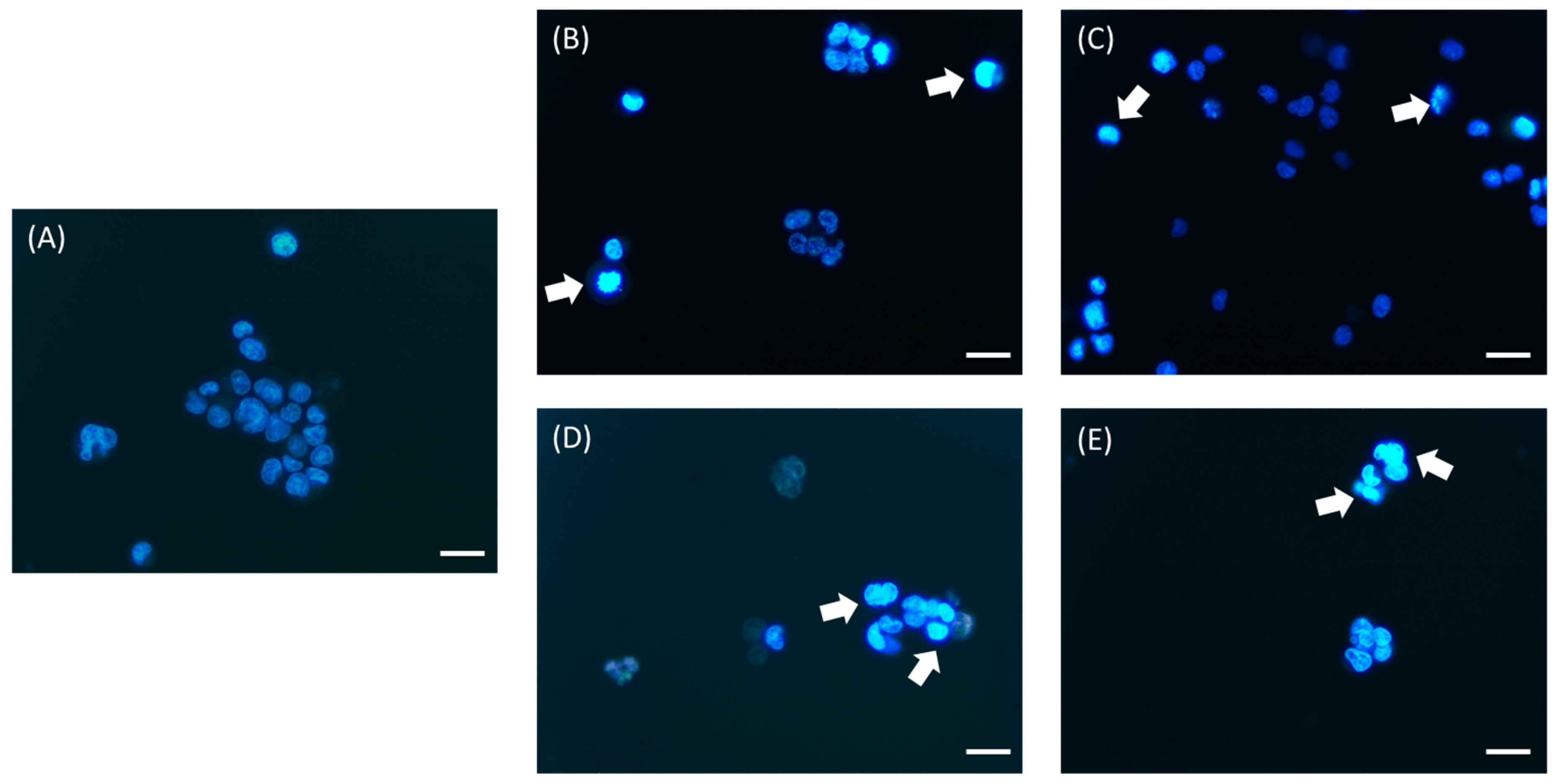
2.9. Morphological Evaluation of Cell Death
2.10. Statistical Analyses
3. Results and Discussion
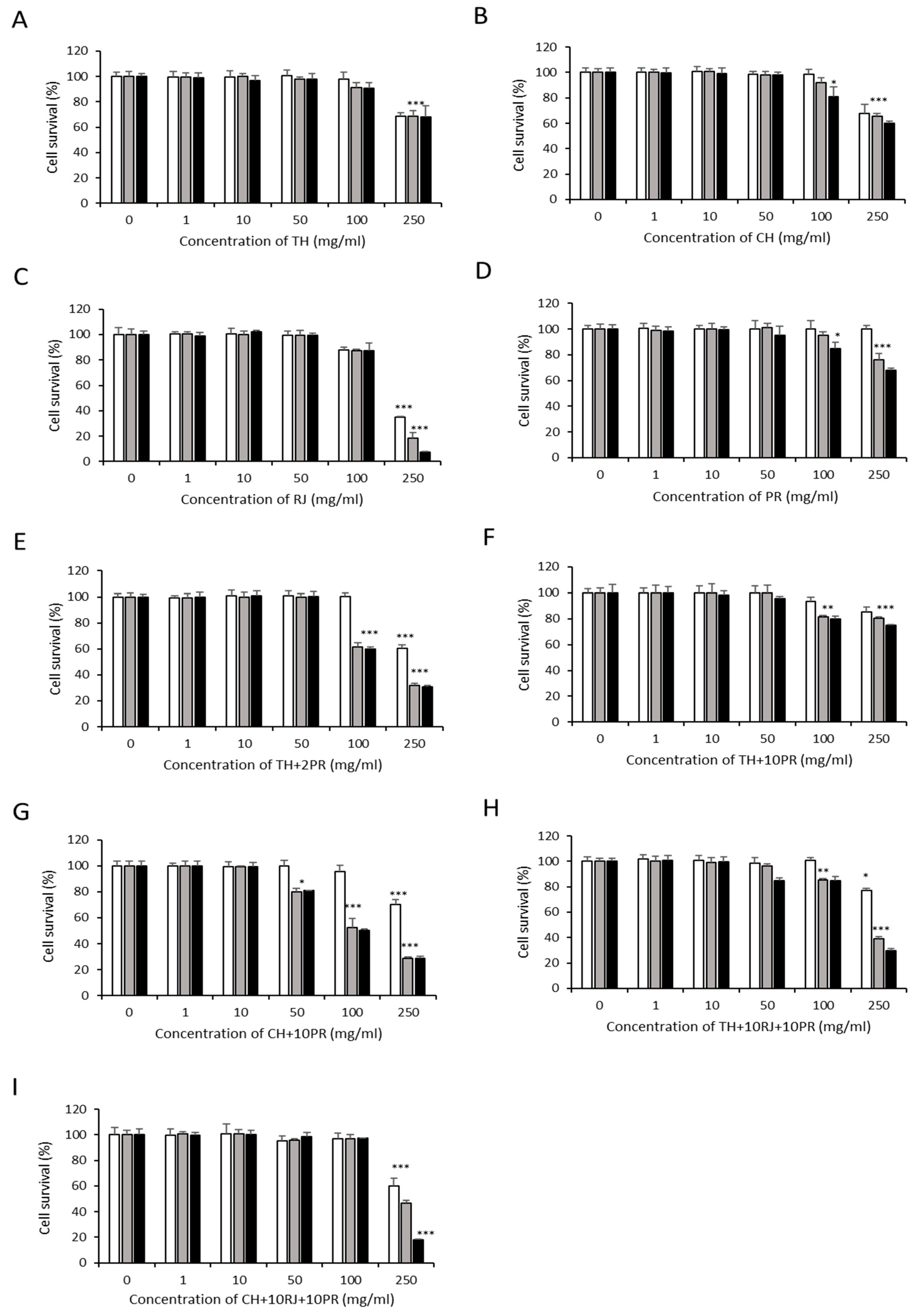
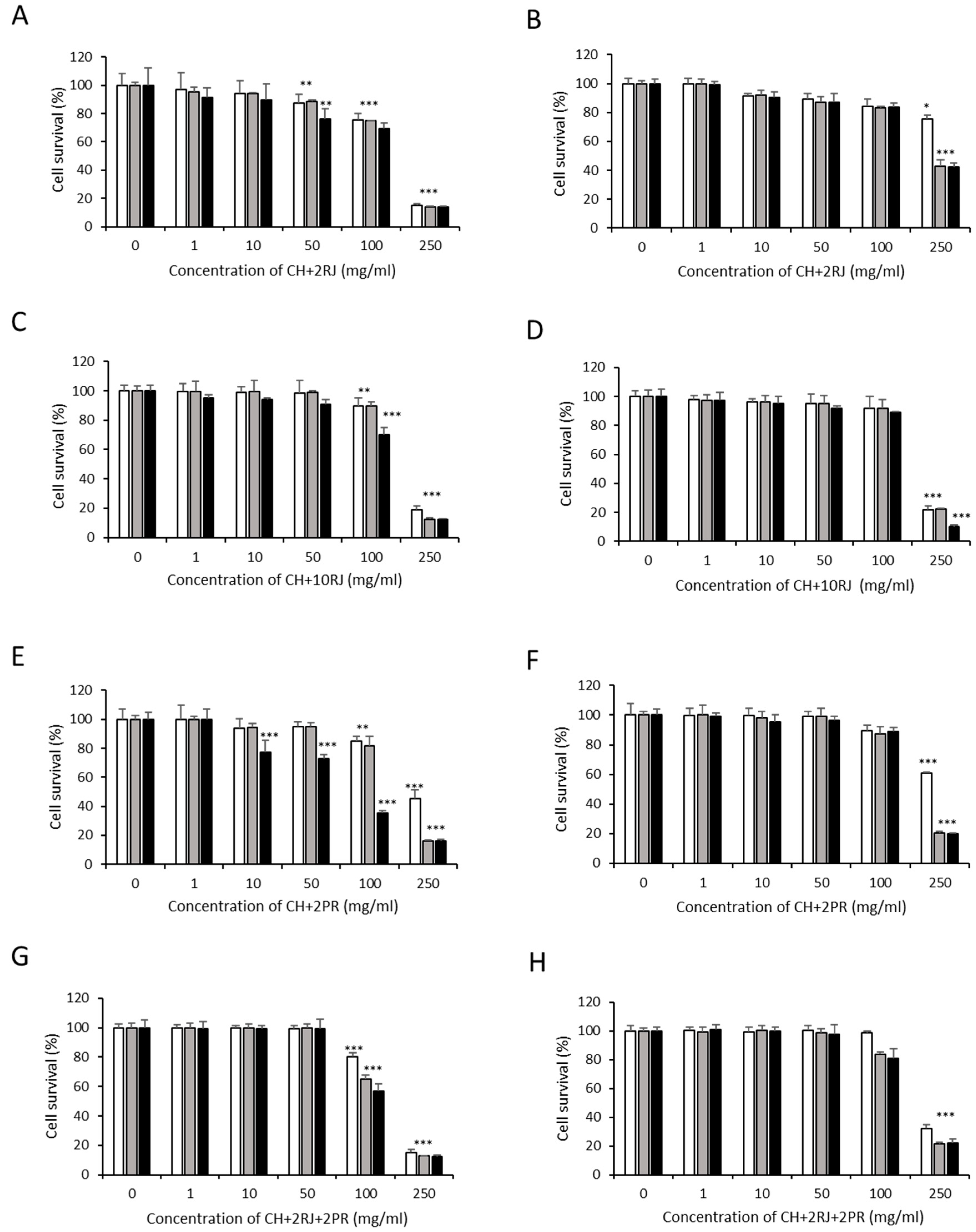
3.1. Cytotoxic Effects of Bee Product Mixtures on Human Liver Cells
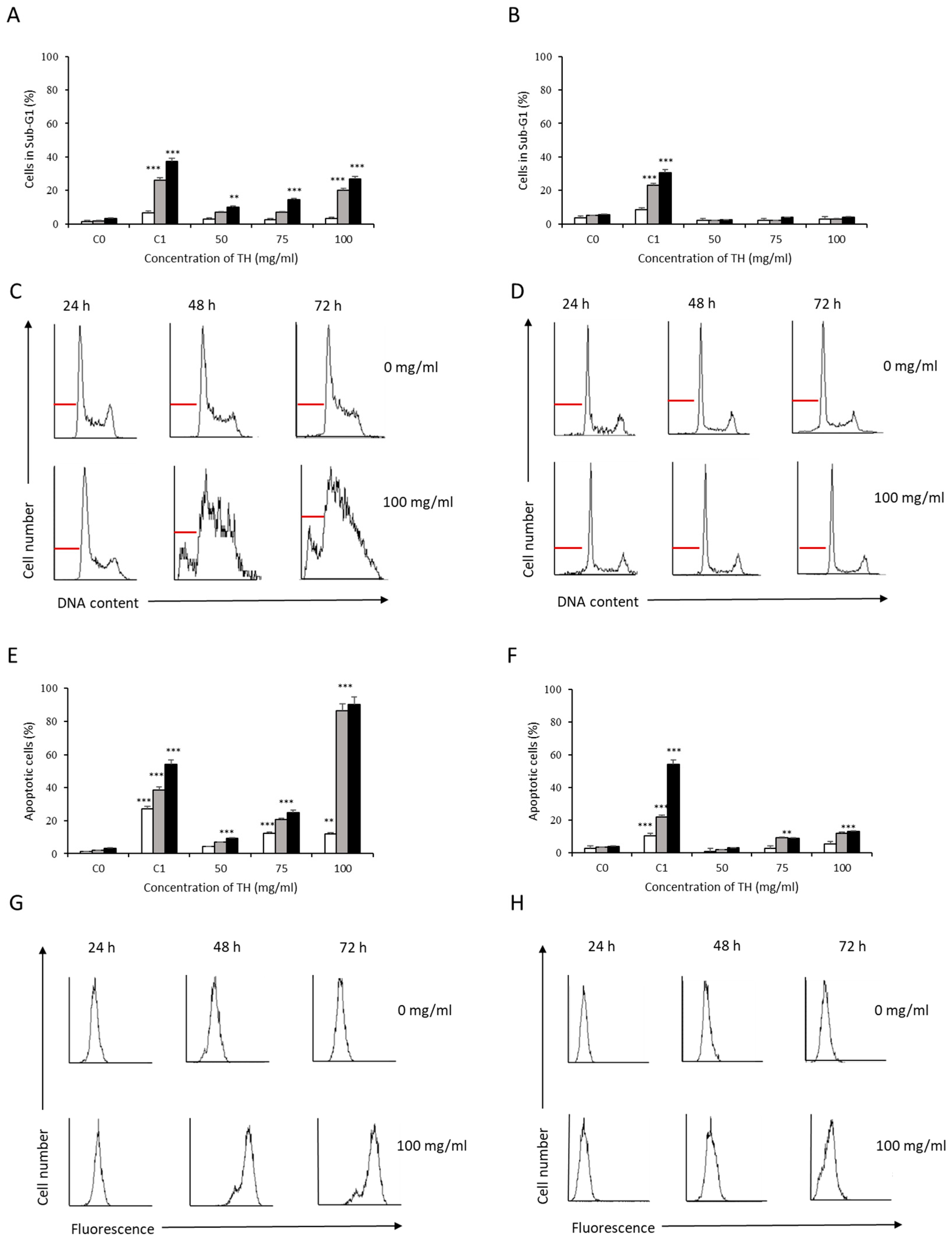
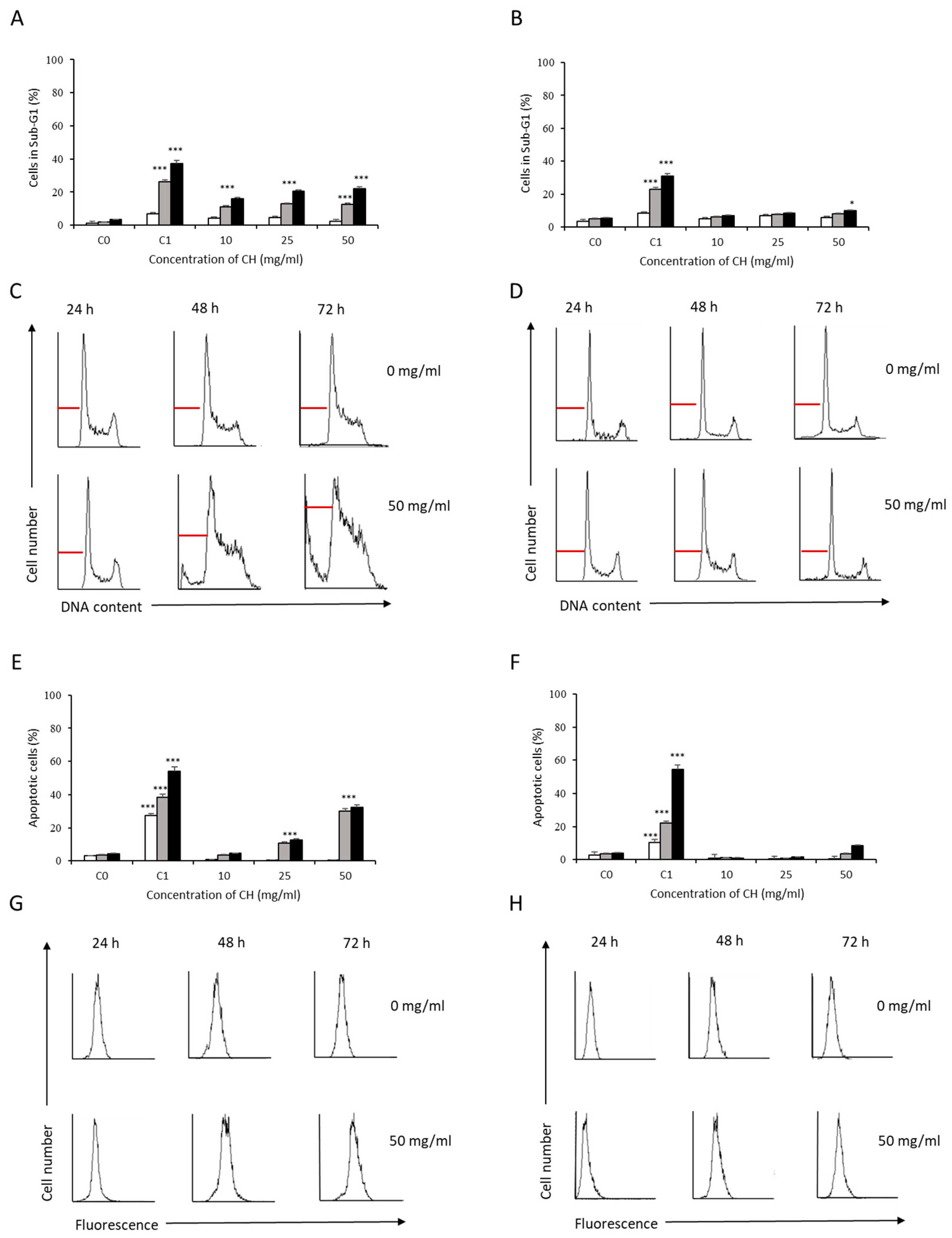
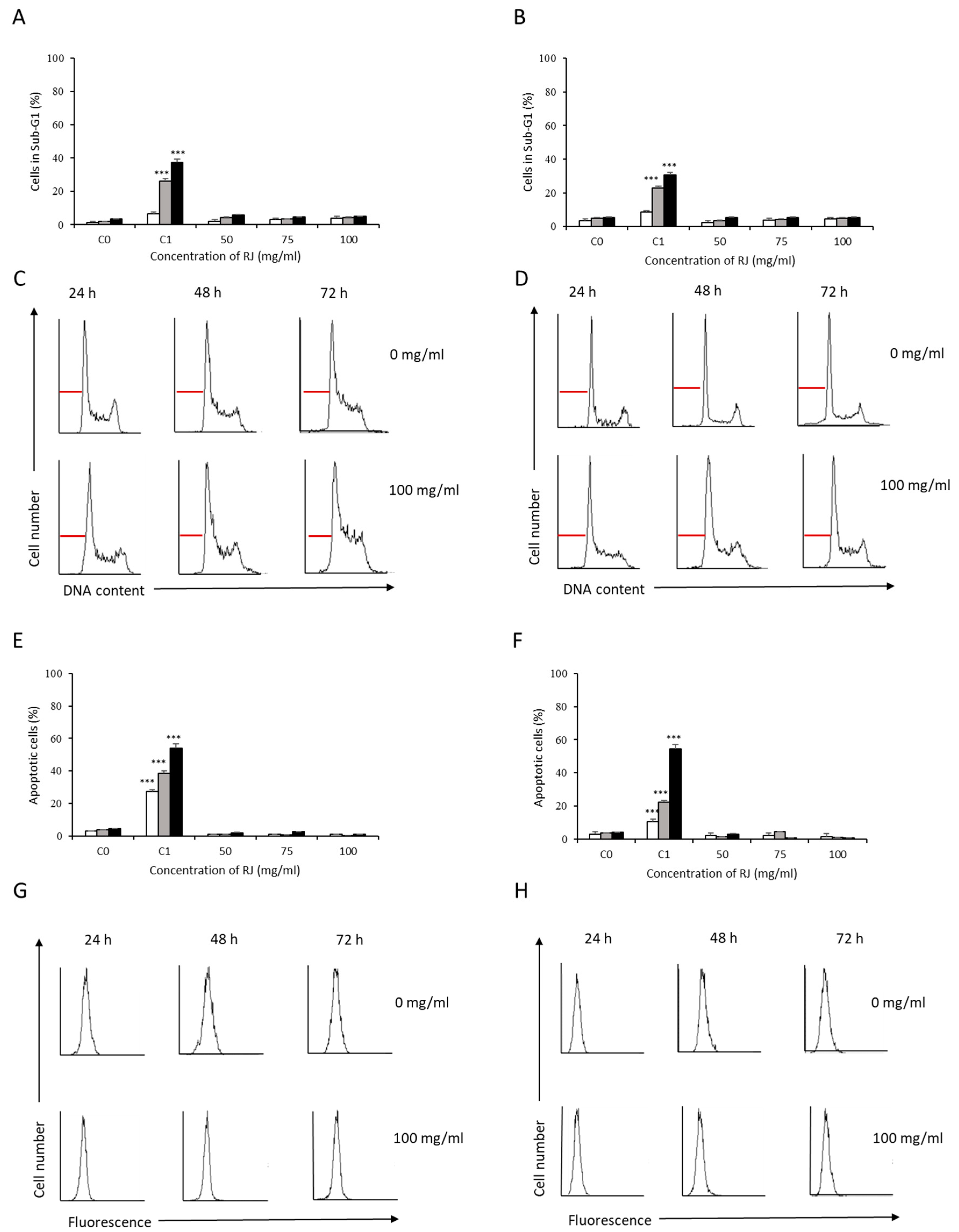
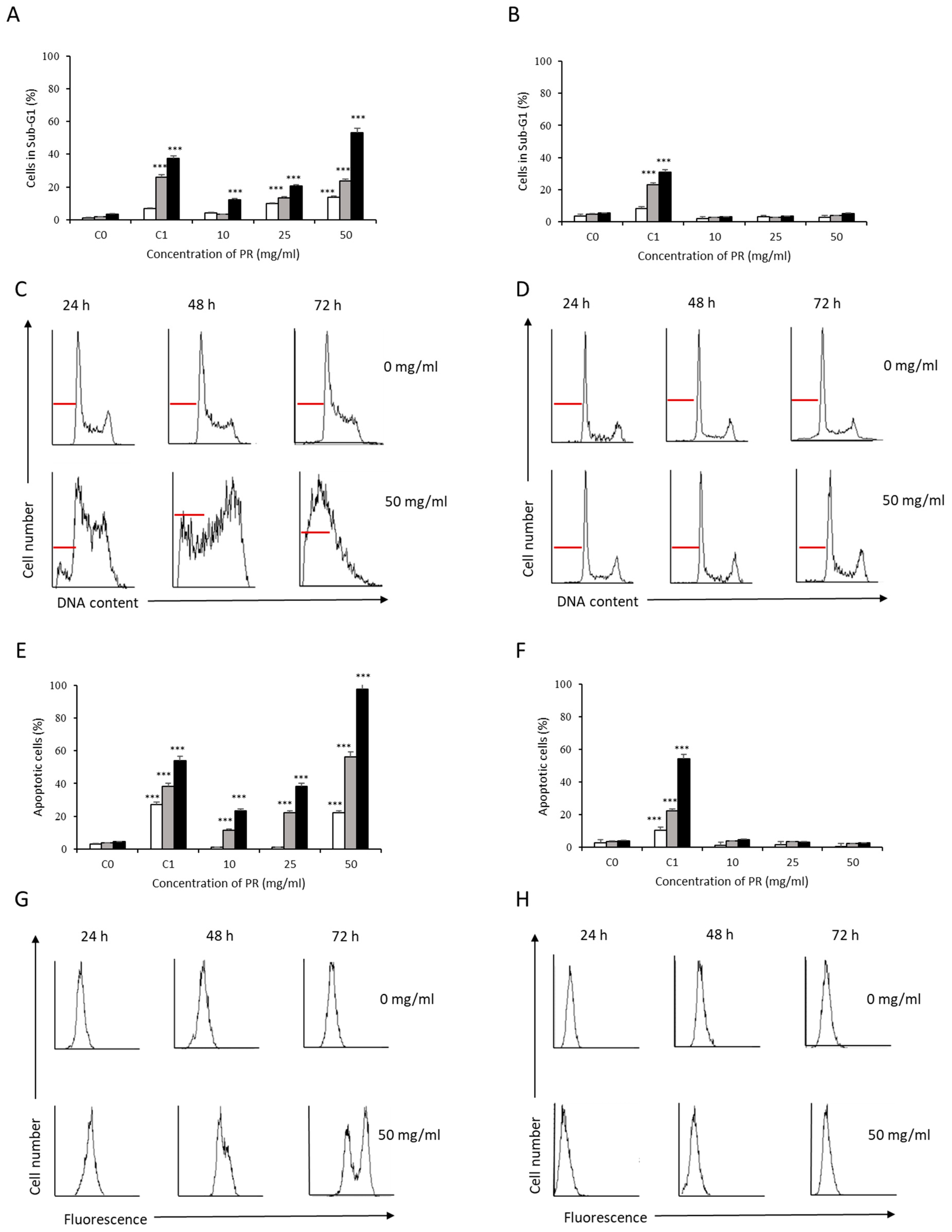
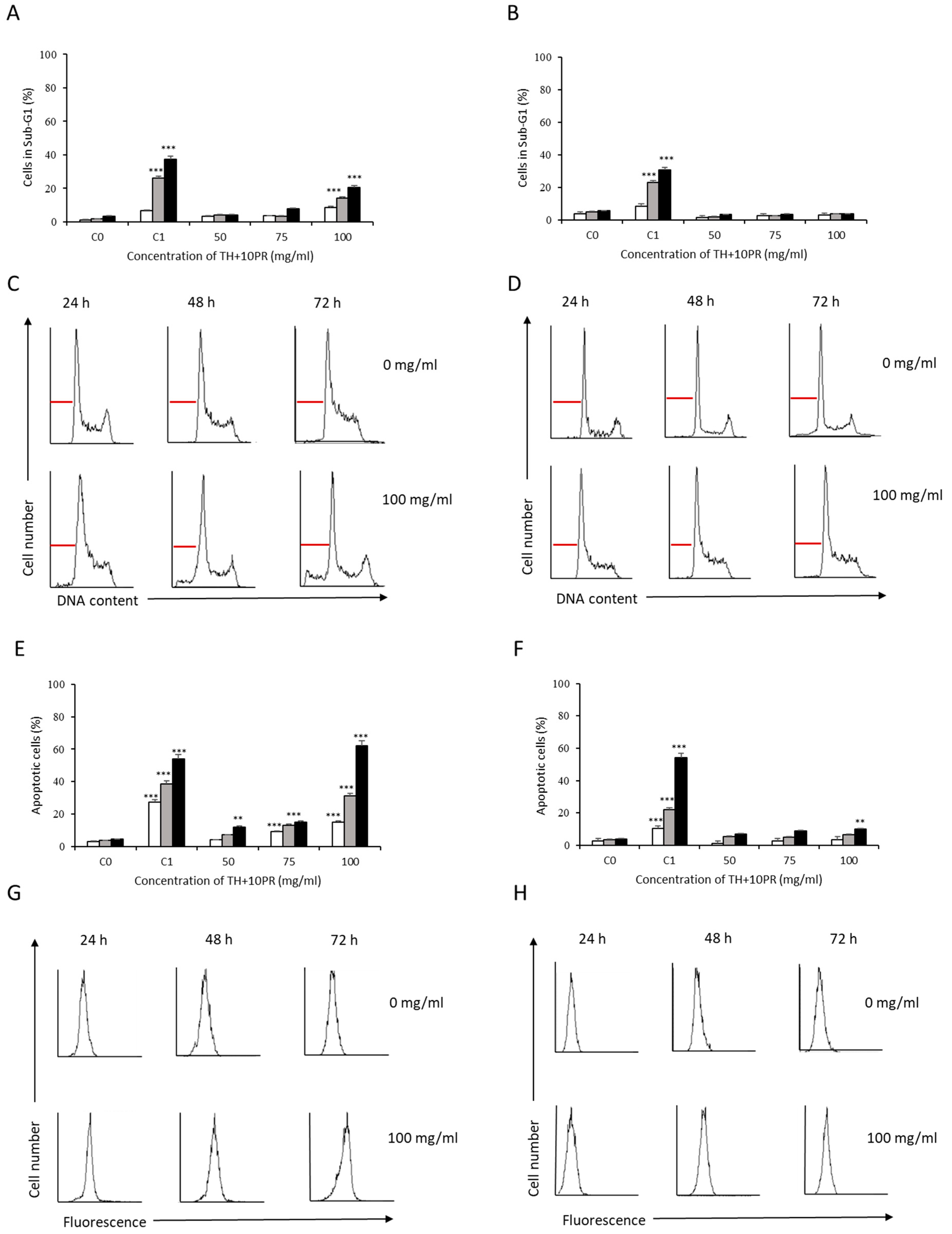
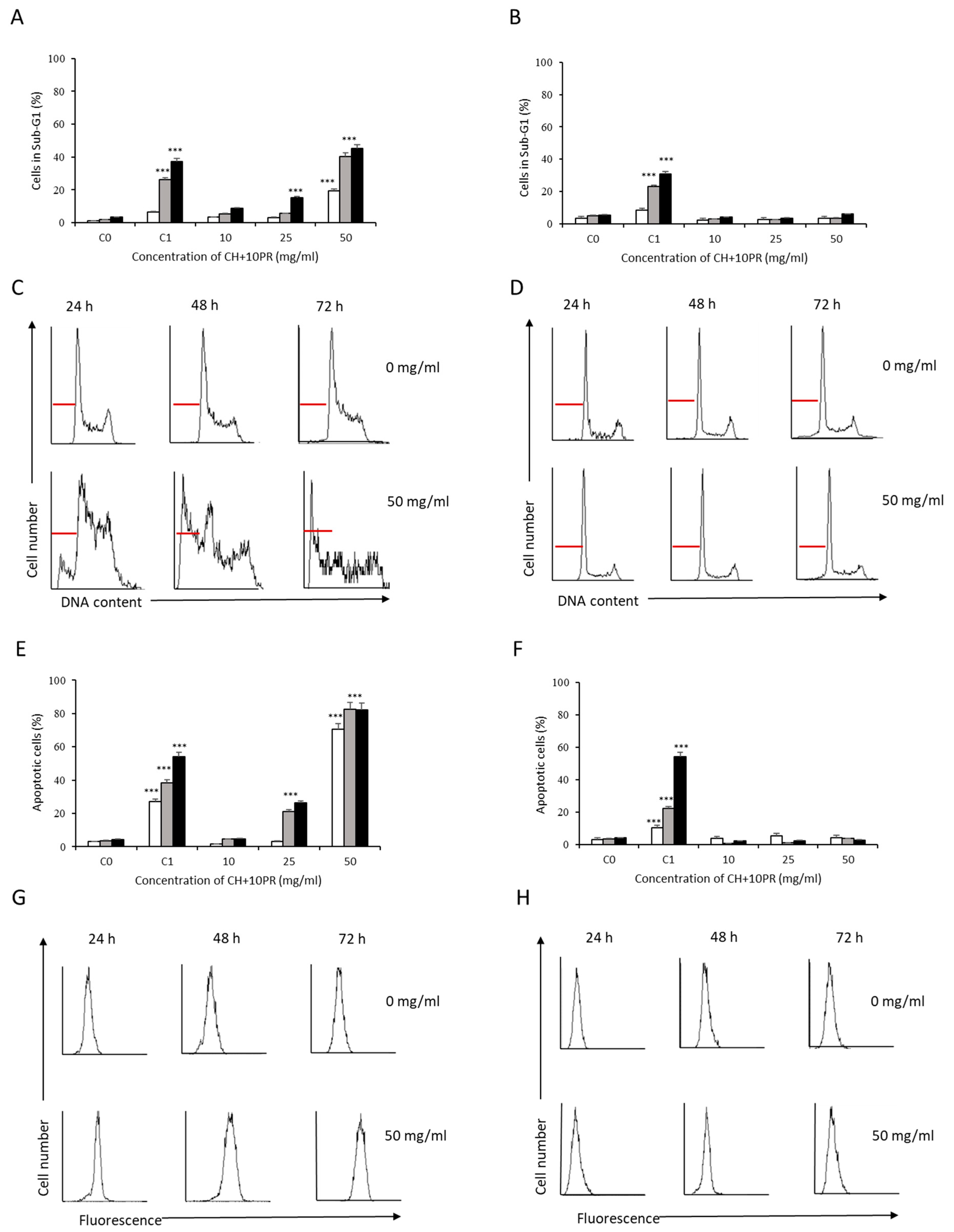
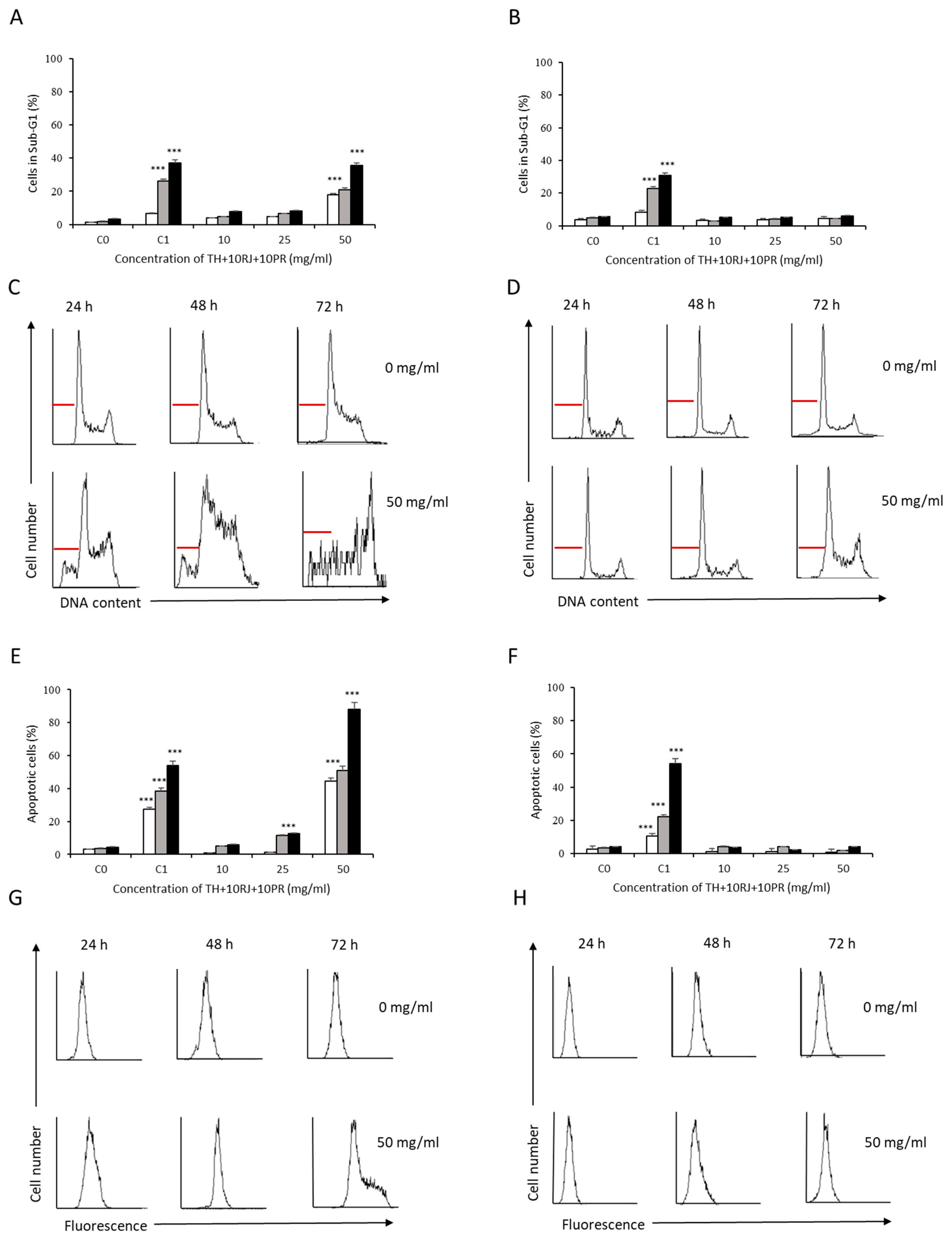
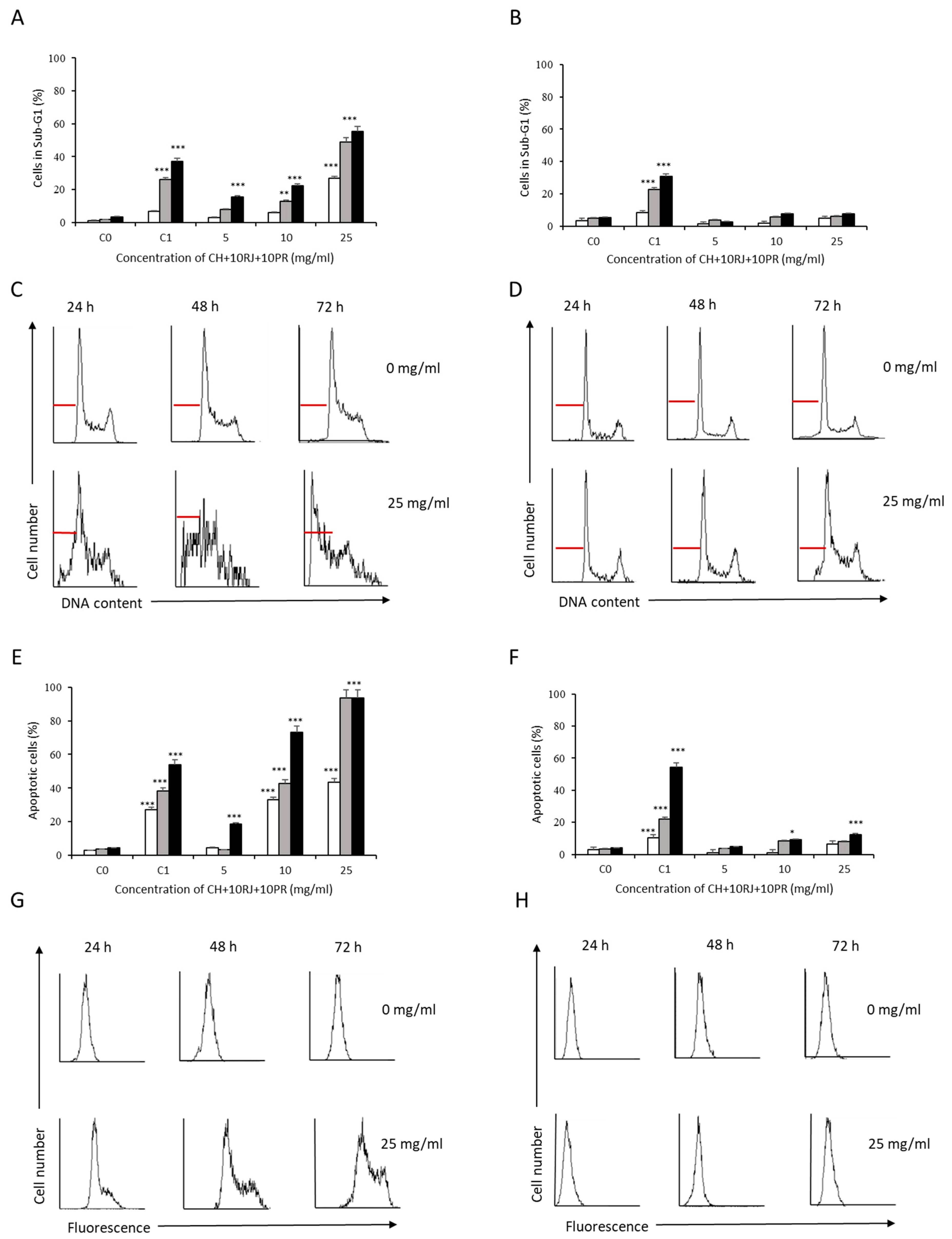
3.2. Analysis of Apoptosis Induction by Bee Product Mixtures on Human Liver Cells
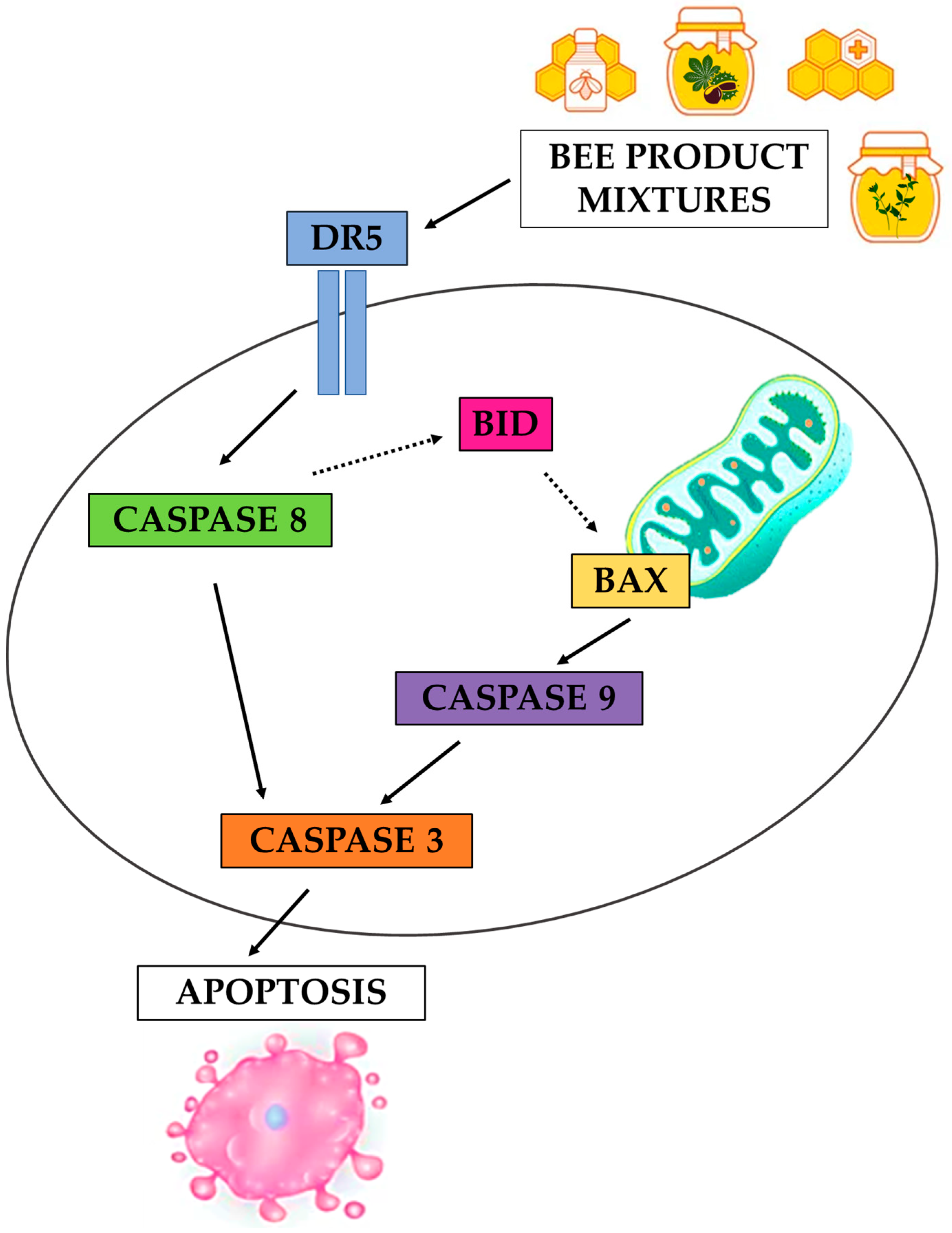
3.3. Evaluation of the Apoptotic Mechanisms of Selected Samples on HepG2 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations Development Programme. Buzzing with Life. Available online: https://bees.undp.org/ (accessed on 11 January 2023).
- European Food Safety Authority (EFSA). Bee Health. Available online: https://www.efsa.europa.eu/en/topics/topic/bee-health (accessed on 11 January 2023).
- Mărgăoan, R.; Topal, E.; Balkanska, R.; Yücel, B.; Oravecz, T.; Cornea-Cipcigan, M.; Vodnar, D.C. Monofloral honeys as a potential source of natural antioxidants, minerals and medicine. Antioxidants 2021, 10, 1023. [Google Scholar] [CrossRef]
- Vercelli, M.; Novelli, S.; Ferrazzi, P.; Lenthini, G.; Ferracini, C. A Qualitative Analysis of Beekeepers’ Perceptions and Farm Honey Bees. Insects 2021, 12, 228. [Google Scholar] [CrossRef]
- The Bee Conservancy. 10 Ways to Save the Bees. Available online: https://thebeeconservancy.org/10-ways-to-save-the-bees/ (accessed on 11 January 2023).
- Martinello, M.; Mutinelli, F. Antioxidant activity in bee products: A review. Antioxidants 2021, 10, 71. [Google Scholar] [CrossRef]
- Münstedt, K.; Männle, H. Bee products and their role in cancer prevention and treatment. Complement. Ther. Med. 2020, 51, 102390. [Google Scholar] [CrossRef]
- Tang, J.Y.; Ou-Yang, F.; Hou, M.F.; Huang, H.W.; Wang, H.R.; Li, K.T.; Fayyaz, S.; Shu, C.W.; Chang, H.W. Oxidative stress-modulating drugs have preferential anticancer effects—Involving the regulation of apoptosis, DNA damage, endoplasmic reticulum stress, autophagy, metabolism, and migration. Semin. Cancer Biol. 2019, 58, 109–117. [Google Scholar] [CrossRef]
- Afrin, S.; Giampieri, F.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Varela-López, A.; Quiles, J.L.; Mezzetti, B.; Battino, M. Chemopreventive and Therapeutic Effects of Edible Berries: A Focus on Colon Cancer Prevention and Treatment. Molecules 2016, 21, 169. [Google Scholar] [CrossRef] [Green Version]
- Yaacob, N.S.; Nengsih, A.; Norazmi, M.N. Tualang honey promotes apoptotic cell death induced by tamoxifen in breast cancer cell lines. Evid.-Based Complement. Altern. Med. 2013, 2013, 989841. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Regolo, L.; Alvarez-Suarez, J.M.; Quinzi, D.; Sargenti, A.; Bai, W.; Tian, L.; Giampieri, F.; Battino, M. Manuka honey in combination with 5-Fluorouracil decreases physical parameters of colonspheres enriched with cancer stem-like cells and reduces their resistance to apoptosis. Food Chem. 2022, 374, 131753. [Google Scholar] [CrossRef]
- Fernandez-Cabezudo, M.J.; El-Kharrag, R.; Torab, F.; Bashir, G.; George, J.A.; El-Taji, H.; al-Ramadi, B.K. Intravenous Administration of Manuka Honey Inhibits Tumor Growth and Improves Host Survival When Used in Combination with Chemotherapy in a Melanoma Mouse Model. PLoS ONE 2013, 8, e55993. [Google Scholar] [CrossRef] [Green Version]
- Erejuwa, O.O.; Sulaiman, S.A.; Ab Wahab, M.S. Effects of honey and its mechanisms of action on the development and progression of cancer. Molecules 2014, 19, 2497–2522. [Google Scholar] [CrossRef] [Green Version]
- Münstedt, K.; Männle, H. Using bee products for the prevention and treatment of oral mucositis induced by cancer treatment. Molecules 2019, 24, 3023. [Google Scholar] [CrossRef] [Green Version]
- Waheed, M.; Hussain, M.B.; Javed, A.; Mushtaq, Z.; Hassan, S.; Shariati, M.A.; Khan, M.U.; Majeed, M.; Nigam, M.; Mishra, A.P.; et al. Honey and cancer: A mechanistic review. Clin. Nutr. 2019, 38, 2499–2503. [Google Scholar] [CrossRef]
- Martinotti, S.; Pellavio, G.; Patrone, M.; Laforenza, U.; Ranzato, E. Manuka Honey Induces Apoptosis of Epithelial Cancer Cells through Aquaporin-3 and Calcium Signaling. Life 2020, 10, 256. [Google Scholar] [CrossRef]
- Kocyigit, A.; Aydogdu, G.; Balkan, E.; Yenigun, V.B.; Guler, E.M.; Bulut, H.; Koktasoglu, F.; Gören, A.C.; Atayoglu, A.T. Quercus pyrenaica Honeydew Honey with High Phenolic Contents Cause DNA Damage, Apoptosis, and Cell Death Through Generation of Reactive Oxygen Species in Gastric Adenocarcinoma Cells. Integr. Cancer Ther. 2019, 18, 1534735419876334. [Google Scholar] [CrossRef] [Green Version]
- Elumalai, P.; Muninathan, N.; Megalatha, S.T.; Suresh, A.; Kumar, K.S.; Jhansi, N.; Kalaivani, K.; Krishnamoorthy, G. An Insight into Anticancer Effect of Propolis and Its Constituents: A Review of Molecular Mechanisms. Evid.-Based Complement. Altern. Med. 2022, 2022, 5901191. [Google Scholar] [CrossRef]
- Salama, S.; Shou, Q.; Abd El-Wahed, A.A.; Elias, N.; Xiao, J.; Swillam, A.; Umair, M.; Guo, Z.; Daglia, M.; Wang, K.; et al. Royal Jelly: Beneficial Properties and Synergistic Effects with Chemotherapeutic Drugs with Particular Emphasis in Anticancer Strategies. Nutrients 2022, 14, 4166. [Google Scholar] [CrossRef]
- Jiang, X.S.; Xie, H.Q.; Li, C.G.; You, M.M.; Zheng, Y.F.; Li, G.Q.; Chen, X.; Zhang, C.P.; Hu, F.L. Chinese Propolis Inhibits the Proliferation of Human Gastric Cancer Cells by Inducing Apoptosis and Cell Cycle Arrest. Evid.-Based Complement. Alternat. Med. 2020, 2020, 2743058. [Google Scholar] [CrossRef]
- Ohta, T.; Kunimasa, K.; Kobayashi, T.; Sakamoto, M.; Kaji, K. Propolis Suppresses Tumor Angiogenesis by Inducing Apoptosis in Tube-Forming Endothelial Cells. Biosci. Biotechnol. Biochem. 2008, 72, 2436–2440. [Google Scholar] [CrossRef]
- Zhang, S.; Shao, Q.; Geng, H.; Su, S. The effect of royal jelly on the growth of breast cancer in mice. Oncol. Lett. 2017, 14, 7615–7621. [Google Scholar] [CrossRef] [Green Version]
- Abu-Serie, M.M.; Habashy, N.H. Suppressing crucial oncogenes of leukemia initiator cells by major royal jelly protein 2 for mediating apoptosis in myeloid and lymphoid leukemia cells. Food Funct. 2022, 13, 8951–8966. [Google Scholar] [CrossRef]
- Long, F.; Yang, H.; Xu, Y.; Hao, H.; Li, P. A strategy for the identification of combinatorial bioactive compounds contributing to the holistic effect of herbal medicines. Sci. Rep. 2015, 5, 12361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, H.; Hao, R.L.; Li, X.F.; Raikos, V.; Li, H.H. Synergistic anticancer and antibacterial activities of cordycepin and selected natural bioactive compounds. Trop. J. Pharm. Res. 2018, 17, 1621–1627. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, A.; Haza, A.I.; Arranz, N.; Delgado, M.E.; Rafter, J.; Morales, P. Organosulfur compounds alone or in combination with vitamin C protect towards N-nitrosopiperidine- and N-nitrosodibutylamine-induced oxidative DNA damage in HepG2 cells. Chem. Biol. Interact. 2008, 173, 9–18. [Google Scholar] [CrossRef]
- Niedzwiecki, A.; Roomi, M.; Kalinovsky, T.; Rath, M. Anticancer Efficacy of Polyphenols and Their Combinations. Nutrients 2016, 8, 552. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Martín, V.; Morales, P.; González-Porto, A.V.; Iriondo-DeHond, A.; López-Parra, M.B.; Del Castillo, M.D.; Hospital, X.F.; Fernández, M.; Hierro, E.; Haza, A.I. Enhancement of the Antioxidant Capacity of Thyme and Chestnut Honey by Addition of Bee Products. Foods 2022, 11, 3118. [Google Scholar] [CrossRef]
- Sánchez-Martín, V.; Haza, A.I.; Iriondo-DeHond, A.; Del Castillo, M.D.; Hospital, X.F.; Fernández, M.; Hierro, E.; Morales, P. Protective Effect of Thyme and Chestnut Honeys Enriched with Bee Products against Benzo(a)pyrene-Induced DNA Damage. Int. J. Environ. Res. Public Health 2022, 19, 16969. [Google Scholar] [CrossRef]
- Ministerio de la Presidencia. Spanish Royal Decree-Law (RD) 1049/2003; BOE No. 186; BOE: Madrid, Spain, 2003. [Google Scholar]
- Huang, X.; Dorota Halicka, H.; Traganos, F.; Tanaka, T.; Kurose, A.; Darzynkiewicz, Z. Cytometric assessment of DNA damage in relation to cell cycle phase and apoptosis. Cell Prolif. 2005, 38, 223–243. [Google Scholar] [CrossRef]
- Sánchez-Martín, V.; Jiménez-García, L.; Herranz, S.; Luque, A.; Acebo, P.; Amesty, Á.; Estévez-Braun, A.; De Las Heras, B.; Hortelano, S. α-hispanolol induces apoptosis and suppresses migration and invasion of glioblastoma cells likely via downregulation of MMP-2/9 expression and p38MAPK attenuation. Front. Pharmacol. 2019, 10, 935. [Google Scholar] [CrossRef]
- Kyrylkova, K.; Kyryachenko, S.; Leid, M.; Kioussi, C. Detection of apoptosis by TUNEL assay. Methods Mol. Biol. 2012, 887, 41–47. [Google Scholar] [CrossRef]
- Jubri, Z.; Narayanan, N.; Abdul Karim, N.; Wan Ngah, W. Antiproliferative activity and apoptosis induction by gelam honey on liver cancer cell line. Int. J. Appl. Sci. Technol. 2012, 2, 135–141. [Google Scholar]
- Hassan, M.I.; Mabrouk, G.M.; Shehata, H.H.; Aboelhussein, M.M. Antineoplastic Effects of Bee Honey and Nigella sativa on Hepatocellular Carcinoma Cells. Integr. Cancer Ther. 2012, 11, 354–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.; Zahran, F.; Shehata, H.; Abdelhamid, M. The inhibitory, antioxidant and anti-inflammatory effects of bee honey on hepatocellular carcinoma cell line HepG2. Int. J. Biol. Pharm. Res. 2013, 4, 618–626. [Google Scholar]
- Abu, M.N.; Salleh, M.A.; Radzman, N.H.; Ismail, W.I.; Yusoff, R.M.; Hasan, H. Insulin Sensitivity Enhancement of the Mixture of Tinospora Crispa and Gelam (Melaleuca Cajuputi) Honey and Its Antiproliferative Activity on Hepatocellular carcinoma, HepG2: A Preliminary Study. J. Med. Res. Dev. 2013, 2, 48–54. [Google Scholar]
- Al Refaey, H.R.; Newairy, A.S.A.; Wahby, M.M.; Albanese, C.; Elkewedi, M.; Choudhry, M.U.; Sultan, A.S. Manuka honey enhanced sensitivity of HepG2, hepatocellular carcinoma cells, for Doxorubicin and induced apoptosis through inhibition of Wnt/β-catenin and ERK1/2. Biol. Res. 2021, 54, 16. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.; Zhao, H.; Chen, S.; He, Q.; Cao, W. Jujube honey induces apoptosis in human hepatocellular carcinoma HepG2 cell via DNA damage, p53 expression, and caspase activation. J. Food Biochem. 2019, 43, 12998. [Google Scholar] [CrossRef]
- Wang, W.; Upur, H.; Chang, J. Alhagi-honey and its compound abnormal savda munziq (ASMq) inhibit proliferation and enhance apoptosis of human hepatoma BEL-7402 cells. Int. J. Clin. Exp. Med. 2016, 9, 15502–15509. [Google Scholar]
- Morales, P.; Haza, A.I. Antiproliferative and apoptotic effects of spanish honeys. Pharmacogn. Mag. 2013, 9, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Fauzi, A.N.; Yaacob, N.S. Cell cycle and apoptosis pathway modulation by Tualang honey in ER-dependent and -independent breast cancer cell lines. J. Apic. Res. 2016, 55, 366–374. [Google Scholar] [CrossRef]
- Aliyu, M.; Odunola, O.A.; Farooq, A.D.; Rasheed, H.; Mesaik, A.M.; Choudhary, M.I.; Channa, I.S.; Khan, S.A.; Erukainure, O.L. Molecular Mechanism of Antiproliferation Potential of Acacia Honey on NCI-H460 Cell Line. Nutr. Cancer 2013, 65, 296–304. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Afrin, S.; Gasparrini, M.; Reboredo-Rodriguez, P.; Manna, P.P.; Zhang, J.; Lamas, L.B.; Flórez, S.M.; Toyos, P.A.; et al. Phenolic compounds in honey and their associated health benefits: A review. Molecules 2018, 23, 2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, Y.; Meenu, M.; Yu, X.; Xu, B.; Cheung, Y.; Meenu, M. Phenolic acids and flavonoids profiles of commercial honey from different floral sources and geographic sources. Int. J. Food Prop. 2019, 22, 290–308. [Google Scholar] [CrossRef] [Green Version]
- Anantharaju, P.G.; Gowda, P.C.; Vimalambike, M.G.; Madhunapantula, S.V. An overview on the role of dietary phenolics for the treatment of cancers. Nutr. J. 2016, 15, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raina, R.; Hussain, A.; Sharma, R. Molecular insight into apoptosis mediated by flavones in cancer (Review). World Acad. Sci. J. 2020, 2, 6. [Google Scholar] [CrossRef]
- Kassi, E.; Chinou, I.; Spilioti, E.; Tsiapara, A.; Graikou, K.; Karabournioti, S.; Manoussakis, M.; Moutsatsou, P. A monoterpene, unique component of thyme honeys, induces apoptosis in prostate cancer cells via inhibition of NF-κB activity and IL-6 secretion. Phytomedicine 2014, 21, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Kıvrak, Ş.; Kıvrak, İ. Assessment of phenolic profile of Turkish honeys. Int. J. Food Prop. 2017, 20, 864–876. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Hao, Y.; Chen, S.; Jia, G.; Guo, Y.; Zhang, G.; Wang, C.; Cheng, R.; Hu, T.; Zhang, X.; et al. Rutin induces apoptosis via P53 up-regulation in human glioma CHME cells. Transl. Cancer Res. 2019, 8, 2005–2013. [Google Scholar] [CrossRef]
- Zhu, B.; Shang, B.; Li, Y.; Zhen, Y. Inhibition of histone deacetylases by trans-cinnamic acid and its antitumor effect against colon cancer xenografts in athymic mice. Mol. Med. Rep. 2016, 13, 4159–4166. [Google Scholar] [CrossRef] [Green Version]
- Hamadou, W.S.; Bouali, N.; Badraoui, R.; Lajimi, R.H.; Hamdi, A.; Alreshidi, M.; Patel, M.; Adnan, M.; Siddiqui, A.J.; Noumi, E.; et al. Chemical Composition and the Anticancer, Antimicrobial, and Antioxidant Properties of Acacia Honey from the Hail Region: The in vitro and in silico Investigation. Evid.-Based Complement. Alternat. Med. 2022, 2022, 1518511. [Google Scholar] [CrossRef]
- Combarros-Fuertes, P.; Estevinho, L.M.; Dias, L.G.; Castro, J.M.; Tomás-Barberán, F.A.; Tornadijo, M.E.; Fresno-Baro, J.M. Bioactive Components and Antioxidant and Antibacterial Activities of Different Varieties of Honey: A Screening Prior to Clinical Application. J. Agric. Food Chem. 2019, 67, 688–698. [Google Scholar] [CrossRef] [Green Version]
- Sethi, G.; Rath, P.; Chauhan, A.; Ranjan, A.; Choudhary, R.; Ramniwas, S.; Sak, K.; Aggarwal, D.; Rani, I.; Tuli, H.S. Apoptotic Mechanisms of Quercetin in Liver Cancer: Recent Trends and Advancements. Pharmaceutics 2023, 15, 712. [Google Scholar] [CrossRef]
- Sharma, N.; Biswas, S.; Al-Dayan, N.; Alhegaili, A.S.; Sarwat, M. Antioxidant Role of Kaempferol in Prevention of Hepatocellular Carcinoma. Antioxidants 2021, 10, 1419. [Google Scholar] [CrossRef] [PubMed]
- Gong, H. Pinocembrin suppresses proliferation and enhances apoptosis in lung cancer cells in vitro by restraining autophagy. Bioengineered 2021, 12, 6035–6044. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Chen, A.Y.; Ye, X.; Guan, R.; Rankin, G.O.; Chen, Y.C. Galangin, a Flavonoid from Lesser Galangal, Induced Apoptosis via p53-Dependent Pathway in Ovarian Cancer Cells. Molecules 2020, 25, 1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alnomasy, S.; Al Shehri, Z. Anti-cancer and cell toxicity effects of royal jelly and its cellular mechanisms against human hepatoma cells. Pharmacogn. Mag. 2022, 18, 635–640. [Google Scholar]
- Kazemi, V.; Eskafi, M.; Saeedi, M.; Manayi, A.; Hadjiakhoondi, A. Physicochemical Properties of Royal Jelly and Comparison of Commercial with Raw Specimens. Jundishapur J. Nat. Pharm. Prod. 2019, 14, 64920. [Google Scholar] [CrossRef]
- Ayna, A.; Tunç, A.; Özbolat, S.; Bengü, A.Ş.; Aykutoğlu, G.; Canli, D.; Polat, R.; Çiftci, M.; Darendelioğlu, E. Anticancer, and antioxidant activities of royal jelly on HT-29 colon cancer cells and melissopalynological analysis. Turk. J. Botany 2021, 45, 808–819. [Google Scholar] [CrossRef]
- Fazili, N.; Soheila Soheili, Z.; Malekzadeh-Shafaroudi, S.; Samiei, S.; Alipoor, S.D.; Moshtaghi, N.; Bagheri, A. Royal Jelly Decreases MMP-9 Expression and Induces Apoptosis in Human 5637 Bladder Cancer Cells. J. Cell Mol. Res. 2021, 13, 36–43. [Google Scholar] [CrossRef]
- Liu, H.; Li, J.; Yuan, W.; Hao, S.; Wang, M.; Wang, F.; Xuan, H. Bioactive components and mechanisms of poplar propolis in inhibiting proliferation of human hepatocellular carcinoma HepG2 cells. Biomed. Pharmacother. 2021, 144, 112364. [Google Scholar] [CrossRef]
- Misir, S.; Aliyazicioglu, Y.; Demir, S.; Turan, I.; Hepokur, C. Effect of Turkish Propolis on miRNA Expression, Cell Cycle, and Apoptosis in Human Breast Cancer (MCF-7) Cells. Nutr. Cancer 2019, 72, 133–145. [Google Scholar] [CrossRef]
- Zingue, S.; Maxeiner, S.; Rutz, J.; Ndinteh, D.T.; Chun, F.K.H.; Fohouo, F.N.T.; Njamen, D.; Blaheta, R.A. Ethanol-extracted Cameroonian propolis: Antiproliferative effects and potential mechanism of action in prostate cancer. Andrologia 2020, 52, e13698. [Google Scholar] [CrossRef] [PubMed]
- Elnakady, Y.A.; Rushdi, A.I.; Franke, R.; Abutaha, N.; Ebaid, H.; Baabbad, M.; Omar, M.O.M.; Al Ghamdi, A.A. Characteristics, chemical compositions and biological activities of propolis from Al-Bahah, Saudi Arabia. Sci. Rep. 2017, 7, 41453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drago, E.; Bordonaro, M.; Lee, S.; Atamna, W.; Lazarova, D.L. Propolis Augments Apoptosis Induced by Butyrate via Targeting Cell Survival Pathways. PLoS ONE 2013, 8, e73151. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, W.A.S.; Ismail, N.Z.; Muhamad, M.; Omar, E.A.; Abdul Samad, N.; Ooi, J.P.; Mohamad, S. Q-TOF LC-MS compounds evaluation of propolis extract derived from Malaysian stingless bees, Tetrigona apicalis, and their bioactivities in breast cancer cell, MCF7. Saudi J. Biol. Sci. 2022, 29, 103403. [Google Scholar] [CrossRef] [PubMed]
- Abutaha, N.; AL-Zharani, M.; Alotaibi, A.; Cordero, M.A.W.; Bepari, A.; Alarifi, S. In vitro and in vivo investigation of polypharmacology of propolis extract as anticancer, antibacterial, anti-inflammatory, and chemical properties. Open Chem. 2021, 19, 864–874. [Google Scholar] [CrossRef]
- de Mendonça, I.C.; Porto, I.C.; do Nascimento, T.G.; de Souza, N.S.; Oliveira, J.M.; Arruda, R.E.; Mousinho, K.C.; dos Santos, A.F.; Basílio-Júnior, I.D.; Parolia, A.; et al. Brazilian red propolis: Phytochemical screening, antioxidant activity and effect against cancer cells. BMC Complement. Altern. Med. 2015, 15, 357. [Google Scholar] [CrossRef] [Green Version]
- Pelinson, L.P.; Assmann, C.E.; Palma, T.V.; da Cruz, I.B.M.; Pillat, M.M.; Mânica, A.; Stefanello, N.; Weis, G.C.C.; de Oliveira Alves, A.; de Andrade, C.M.; et al. Antiproliferative and apoptotic effects of caffeic acid on SK-Mel-28 human melanoma cancer cells. Mol. Biol. Rep. 2019, 46, 2085–2092. [Google Scholar] [CrossRef]
- Sanaei, M.; Kavoosi, F.; Valiani, A.; Ghobadifar, M.A. Effect of Genistein on Apoptosis and Proliferation of Hepatocellular Carcinoma Hepa1-6 Cell Line. Int. J. Prev. Med. 2018, 9, 12. [Google Scholar] [CrossRef]
- Alday, E.; Valencia, D.; Carreño, A.L.; Picerno, P.; Piccinelli, A.L.; Rastrelli, L.; Robles-Zepeda, R.; Hernandez, J.; Velazquez, C. Apoptotic induction by pinobanksin and some of its ester derivatives from Sonoran propolis in a B-cell lymphoma cell line. Chem. Biol. Interact. 2015, 242, 35–44. [Google Scholar] [CrossRef]
- El-Guendouz, S.; Lyoussi, B.; Miguel, M.G. Insight on Propolis from Mediterranean Countries: Chemical Composition, Biological Activities and Application Fields. Chem. Biodivers. 2019, 16, e1900094. [Google Scholar] [CrossRef]
- Hu, X.; Yang, Z.; Liu, W.; Pan, Z.; Zhang, X.; Li, M.; Liu, X.; Zheng, Q.; Li, D. The Anti-tumor Effects of p-Coumaric Acid on Melanoma A375 and B16 Cells. Front. Oncol. 2020, 10, 558414. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Singh, A.K.; Loka, M.; Pandey, A.K.; Bishayee, A. Ferulic acid-mediated modulation of apoptotic signaling pathways in cancer. Adv. Protein Chem. Struct. Biol. 2021, 125, 215–257. [Google Scholar] [CrossRef] [PubMed]
- Hormozi, M.; Ghoreishi, S.; Baharvand, P. Astaxanthin induces apoptosis and increases activity of antioxidant enzymes in LS-180 cells. Artif. Cells, Nanomed. Biotechnol. 2019, 47, 891–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.H.; Lee, M.J.; Kim, J.H. Antioxidant and apoptosis-inducing activities of ellagic acid. Anticancer Res. 2006, 26, 3601–3606. [Google Scholar]
- Fattahi, S.; Ardekani, A.M.; Zabihi, E.; Abedian, Z.; Mostafazadeh, A.; Pourbagher, R.; Akhavan-Niaki, H. Antioxidant and apoptotic effects of an aqueous extract of Urtica dioica on the MCF-7 human breast cancer cell line. Asian Pac. J. Cancer Prev. 2013, 14, 5317–5323. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, L.; Shi, Z.; Zhao, S.; Wang, F.T.; Zhou, T.T.; Liu, B.; Bao, J.K. Programmed cell death pathways in cancer: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2012, 45, 487–498. [Google Scholar] [CrossRef]
- Kim, M.O.; Moon, D.O.; Kang, C.H.; Kwon, T.K.; Choi, Y.H.; Kim, G.Y. β-Ionone enhances TRAIL-induced apoptosis in hepatocellular carcinoma cells through Sp1-dependent upregulation of DR5 and downregulation of NF-κB activity. Mol. Cancer Ther. 2010, 9, 833–843. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.C.; Parajuli, K.R.; Han, S.I. Role of death receptors-associated lipid rafts in oxaliplatin-induced death mode regulation of HepG2 cells. Anticancer Res. 2020, 40, 2573–2582. [Google Scholar] [CrossRef]
- Charette, N.; De Saeger, C.; Horsmans, Y.; Leclercq, I.; Stärkel, P. Salirasib sensitizes hepatocarcinoma cells to TRAIL-induced apoptosis through DR5 and survivin-dependent mechanisms. Cell Death Dis. 2013, 4, e471. [Google Scholar] [CrossRef]
- Lawag, I.L.; Lim, L.Y.; Joshi, R.; Hammer, K.A.; Locher, C. A Comprehensive Survey of Phenolic Constituents Reported in Monofloral Honeys around the Globe. Foods 2022, 11, 1152. [Google Scholar] [CrossRef]
- Raina, R.; Afroze, N.; Sundaram, M.K.; Haque, S.; Bajbouj, K.; Hamad, M.; Hussain, A. Chrysin inhibits propagation of HeLa cells by attenuating cell survival and inducing apoptotic pathways. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2206–2220. [Google Scholar] [CrossRef]
- Li, X.; Huang, J.M.; Wang, J.N.; Xiong, X.K.; Yang, X.F.; Zou, F. Combination of chrysin and cisplatin promotes the apoptosis of Hep G2 cells by up-regulating p53. Chem. Biol. Interact. 2015, 232, 12–20. [Google Scholar] [CrossRef]
- Kumazawa, S.; Bonvehí, J.S.; Torres, C.; Mok-Ryeon, A.; Bermejo, F.J.O. Chemical and Functional Characterisation of Propolis Collected from East Andalusia (Southern Spain). Phytochem. Anal. 2013, 24, 608–615. [Google Scholar] [CrossRef]
- Dewson, G.; Kluck, R.M. Mechanisms by which Bak and Bax permeabilise mitochondria during apoptosis. J. Cell Sci. 2009, 122, 2801–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarghandian, S.; Nezhad, M.; Mohammadi, G. Role of Caspases, Bax and Bcl-2 in Chrysin-Induced Apoptosis in the A549 Human Lung Adenocarcinoma Epithelial Cells. Anticancer. Agents Med. Chem. 2014, 14, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Motomura, M.; Kwon, K.M.; Suh, S.J.; Lee, Y.C.; Kim, Y.K.; Lee, I.S.; Kim, M.S.; Kwon, D.Y.; Suzuki, I.; Kim, C.H. Propolis induces cell cycle arrest and apoptosis in human leukemic U937 cells through Bcl-2/Bax regulation. Environ. Toxicol. Pharmacol. 2008, 26, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Frion-Herrera, Y.; Díaz-García, A.; Ruiz-Fuentes, J.; Rodríguez-Sánchez, H.; Sforcin, J.M. Brazilian green propolis induced apoptosis in human lung cancer A549 cells through mitochondrial-mediated pathway. J. Pharm. Pharmacol. 2015, 67, 1448–1456. [Google Scholar] [CrossRef] [PubMed]
- Salavatipour, M.; Kouhbananinejad, S.; Lashkari, M.; Bardsiri, M.; Moghadari, M.; Kashani, B.; Farsinejad, A.; Vahidi, R. Kermanian propolis induces apoptosis through upregulation of Bax/Bcl-2 ratio in acute myeloblastic leukemia cell line (NB4). J. Cancer Res. Ther. 2023. [Google Scholar] [CrossRef]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Ghaffari, T.; Hong, J.H.; Asnaashari, S.; Farajnia, S.; Delazar, A.; Hamishehkar, H.; Kim, K.H. Natural phytochemicals derived from gymnosperms in the prevention and treatment of cancers. Int. J. Mol. Sci. 2021, 22, 6636. [Google Scholar] [CrossRef]
- Amalia, E.; Diantini, A.; Prabandari, E.E.; Waluyo, D.; Subarnas, A. Caffeic Acid Phenethyl Ester as a DHODH Inhibitor and Its Synergistic Anticancer Properties in Combination with 5-Fluorouracil in a Breast Cancer Cell Line. J. Exp. Pharmacol. 2022, 14, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Funk, K.; Czauderna, C.; Klesse, R.; Becker, D.; Hajduk, J.; Oelgeklaus, A.; Reichenbach, F.; Fimm-Todt, F.; Lauterwasser, J.; Galle, P.R.; et al. Bax redistribution induces apoptosis resistance and selective stress sensitivity in human HCC. Cancers 2020, 12, 1437. [Google Scholar] [CrossRef] [PubMed]
- Lawag, I.L.; Islam, K.; Sostaric, T.; Lim, L.Y.; Hammer, K.; Locher, C. Antioxidant Activity and Phenolic Compound Identification and Quantification in Western Australian Honeys. Antioxidants 2023, 12, 189. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, S.; Balaji, A.; Vellayappan, M.; Asokan, M.; Subramanian, A.; John, A.; Supriyanto, E.; Razak, S.; Marvibaigi, M. A Review on Antiproliferative and Apoptotic Activities of Natural Honey. Anticancer. Agents Med. Chem. 2014, 15, 48–56. [Google Scholar] [CrossRef]
- Rouibah, H.; Kebsa, W.; Lahouel, M.; Zihlif, M.; Ahram, M.; Aburmeleih, B.; Mustafa, E.; Al-Ameer, H.J. Algerian Propolis Potentiates Doxorubicin Mediated Anticancer Effect Against Human Pancreatic PANC-1 Cancer Cell Line through Cell Cycle Arrest, Apoptosis Induction and P-Glycoprotein Inhibition. Anticancer. Agents Med. Chem. 2018, 18, 375–387. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Scientific and Common Names | Type | Family | Geographic Region |
|---|---|---|---|---|
| Chestnut honey | Castanea sativa Chestnut | Monofloral (47.19%) | Fagaceae | Spain, Toledo (4°44′48″ W, 40°06′26″ N) |
| Thyme honey | Thymus spp. Thyme | Monofloral (64.98%) | Lamiaceae | Spain, Zamora (5°44′40″ W, 41°30′22″ N) |
| Royal jelly | - | - | - | France |
| Propolis tincture * | - | - | - | Spain, Zamora (5°44′40″ W, 41°30′22″ N) |
| Sample No. | Description | Code |
|---|---|---|
| 1 | Thyme honey | TH |
| 2 | Thyme honey + 2% royal jelly | TH+2RJ |
| 3 | Thyme honey + 10% royal jelly | TH+10RJ |
| 4 | Thyme honey + 2% propolis | TH+2PR |
| 5 | Thyme honey + 10% propolis | TH+10PR |
| 6 | Chestnut honey | CH |
| 7 | Chestnut honey + 2% royal jelly | CH+2RJ |
| 8 | Chestnut honey + 10% royal jelly | CH+10RJ |
| 9 | Chestnut honey + 2% propolis | CH+2PR |
| 10 | Chestnut honey + 10% propolis | CH+10PR |
| 11 | Thyme honey + 2% royal jelly + 2% propolis | TH+2RJ+2PR |
| 12 | Thyme honey + 10% royal jelly + 10% propolis | TH+10RJ+10PR |
| 13 | Chestnut honey + 2% royal jelly + 2% propolis | CH+2RJ+2PR |
| 14 | Chestnut honey + 10% royal jelly + 10% propolis | CH+10RJ+10PR |
| 15 | Royal jelly | RJ |
| 16 | Propolis | PR |
| 17 | Artificial honey | AH |
| Sample | Code | Concentration (mg/mL) | DR5-Positive Cells 48 h (%) | DR5-Positive Cells 72 h (%) | BAX-Positive Cells 48 h (%) | BAX-Positive Cells 72 h (%) |
|---|---|---|---|---|---|---|
| Control | - | - | 4.19 ± 0.68 | 4.87 ± 0.73 | 4.86 ± 0.97 | 5.79 ± 0.52 |
| 1 | TH | 100 | 16.19 ± 0.21 *** | 25.97 ± 2.09 *** | 7.53 ± 1.17 | 20.21 ± 1.49 *** |
| 5 | TH+10PR | 100 | 25.83 ± 3.02 *** | 68.28 ± 2.78 *** | 6.01 ± 1.11 | 19.46 ± 2.26 *** |
| 6 | CH | 50 | 27.53 ± 2.60 | 28.58 ± 3.19 *** | 3.45 ± 0.98 | 17.60 ± 2.03 *** |
| 10 | CH+10PR | 25 | 33.10 ± 2.39 *** | 98.08 ± 1.54 *** | 3.01 ± 1.84 | 99.62 ± 0.16 *** |
| 12 | TH+10RJ+10PR | 50 | 10.93 ± 1.07 *** | 19.18 ± 2.51 *** | 2.12 ± 0.43 | 15.83 ± 1.20 *** |
| 14 | CH+10RJ+10PR | 10 | 84.82 ± 0.68 *** | 89.99 ± 1.32 *** | 32.33 ± 2.12 *** | 72.73 ± 2.05 *** |
| 16 | PR | 25 | 99.97 ± 0.07 *** | 99.77 ± 0.14 *** | 89.78 ± 2.60 *** | 99.99 ± 0.06 *** |
| Etoposide | - | 50 µM | 30.26 ± 1.34 *** | 47.78 ± 2.58 *** | 20.62 ± 2.24 *** | 46.95 ± 1.14 *** |
| Sample | Code | Concentration (mg/mL) | Caspase 8 (%) | Caspase 9 (%) | Caspase 3 (%) |
|---|---|---|---|---|---|
| 1 | TH | 100 | 93.48 ± 4.56 *** | 17.42 ± 2.77 *** | 45.34 ± 2.99 *** |
| 5 | TH+10PR | 100 | 89.75 ± 4.15 *** | 15.87 ± 2.54 *** | 44.28 ± 3.78 *** |
| 6 | CH | 50 | 93.68 ± 2.78 *** | 16.35 ± 2.65 *** | 47.13 ± 3.11 *** |
| 10 | CH+10PR | 25 | 83.75 ± 2.47 *** | 15.44 ± 2.04 *** | 45.07 ± 2.73 *** |
| 12 | TH+10RJ+10PR | 50 | 88.40 ± 3.37 *** | 15.92 ± 2.40 *** | 50.28 ± 3.27 *** |
| 14 | CH+10RJ+10PR | 10 | 86.12 ± 3.54 *** | 19.71 ± 2.39 *** | 48.96 ± 2.76 *** |
| 16 | PR | 25 | 83.73 ± 3.38 *** | 15.90 ± 2.38 *** | 44.06 ± 3.19 *** |
| Etoposide | - | 50 µM | 92.98 ± 3.76 *** | 22.36 ± 2.26 *** | 70.78 ± 3.41 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Martín, V.; Morales, P.; Iriondo-DeHond, A.; Hospital, X.F.; Fernández, M.; Hierro, E.; Haza, A.I. Differential Apoptotic Effects of Bee Product Mixtures on Normal and Cancer Hepatic Cells. Antioxidants 2023, 12, 615. https://doi.org/10.3390/antiox12030615
Sánchez-Martín V, Morales P, Iriondo-DeHond A, Hospital XF, Fernández M, Hierro E, Haza AI. Differential Apoptotic Effects of Bee Product Mixtures on Normal and Cancer Hepatic Cells. Antioxidants. 2023; 12(3):615. https://doi.org/10.3390/antiox12030615
Chicago/Turabian StyleSánchez-Martín, Vanesa, Paloma Morales, Amaia Iriondo-DeHond, Xavier F. Hospital, Manuela Fernández, Eva Hierro, and Ana I. Haza. 2023. "Differential Apoptotic Effects of Bee Product Mixtures on Normal and Cancer Hepatic Cells" Antioxidants 12, no. 3: 615. https://doi.org/10.3390/antiox12030615