

Bacterial Volatiles (mVOC) Emitted by the Phytopathogen Erwinia amylovora Promote Arabidopsis thaliana Growth and Oxidative Stress
 ,
,  , and
, and 
Abstract
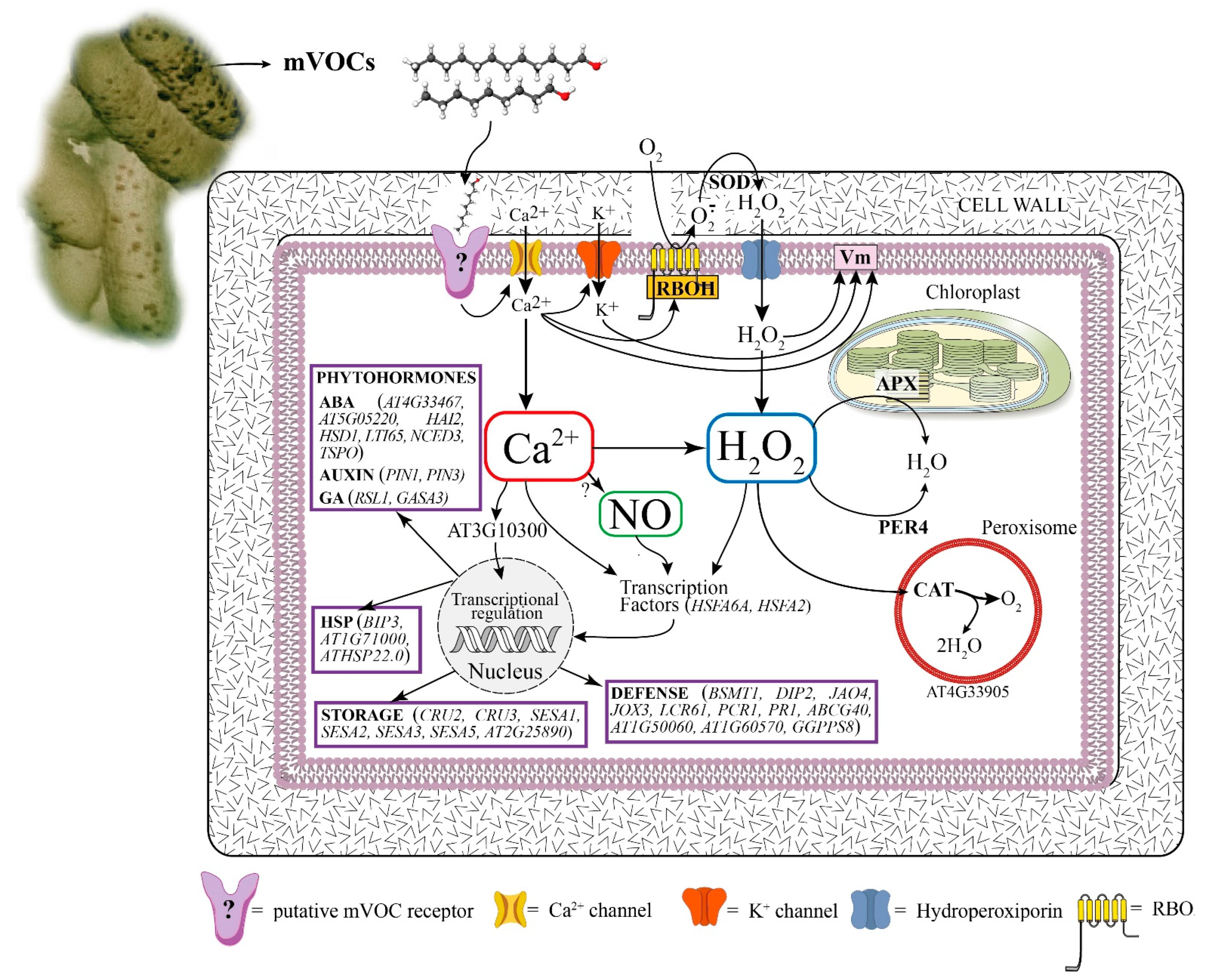
:1. Introduction
2. Materials and Methods
2.1. Plant and Bacterial Co-Cultivation
2.2. Head Space Analysis of E. amylovora mVOCs
2.3. Membrane Potential Determination
2.4. Evaluation of Intracellular Calcium Variations by Confocal Laser Scanning Microscopy Usign Calcium Orange
2.5. Localization of Voltage-Gated K+ Channels (VGKC) and Ligand-Gated or Resting Inward Rectifier K+ Channels (LG/RIRKC) Using FluxORTM
2.6. Localization of H2O2 by CLSM Using Amplex Red
2.7. Determination of Nitric Oxide with 4,5-diaminofluorescein Diacetate (DAF-FM DA) Using CLSM
2.8. RNA Extraction from Arabidopsis Shoots and Roots upon Exposure to E. amylovora mVOCs
2.9. cDNA Synthesis and Gene Microarray Analyses (Including MIAME)
2.10. Validation of Microarray Data by Real-Time PCR
2.11. Expression Analysis of PIN1 and PIN3
2.12. Exposure of A. thaliana to Synthetic 1-nonanol or 1-dodecanol
2.13. Gene Expression of RBOHH, SOD1, CAT1, PER4 and APX1 of A. thaliana Exposed to Synthetic mVOCs
2.14. Statistical Analysis
3. Results and Discussion
3.1. Erwnia Amylovora mVOCs Promote Arabidopsis Shoot and Root Growth
3.2. E. amylovora Emits a Complex Blend of mVOCs
3.3. E. amlylovora mVOCs Induce Early Signaling Events in Arabidopsis thaliana
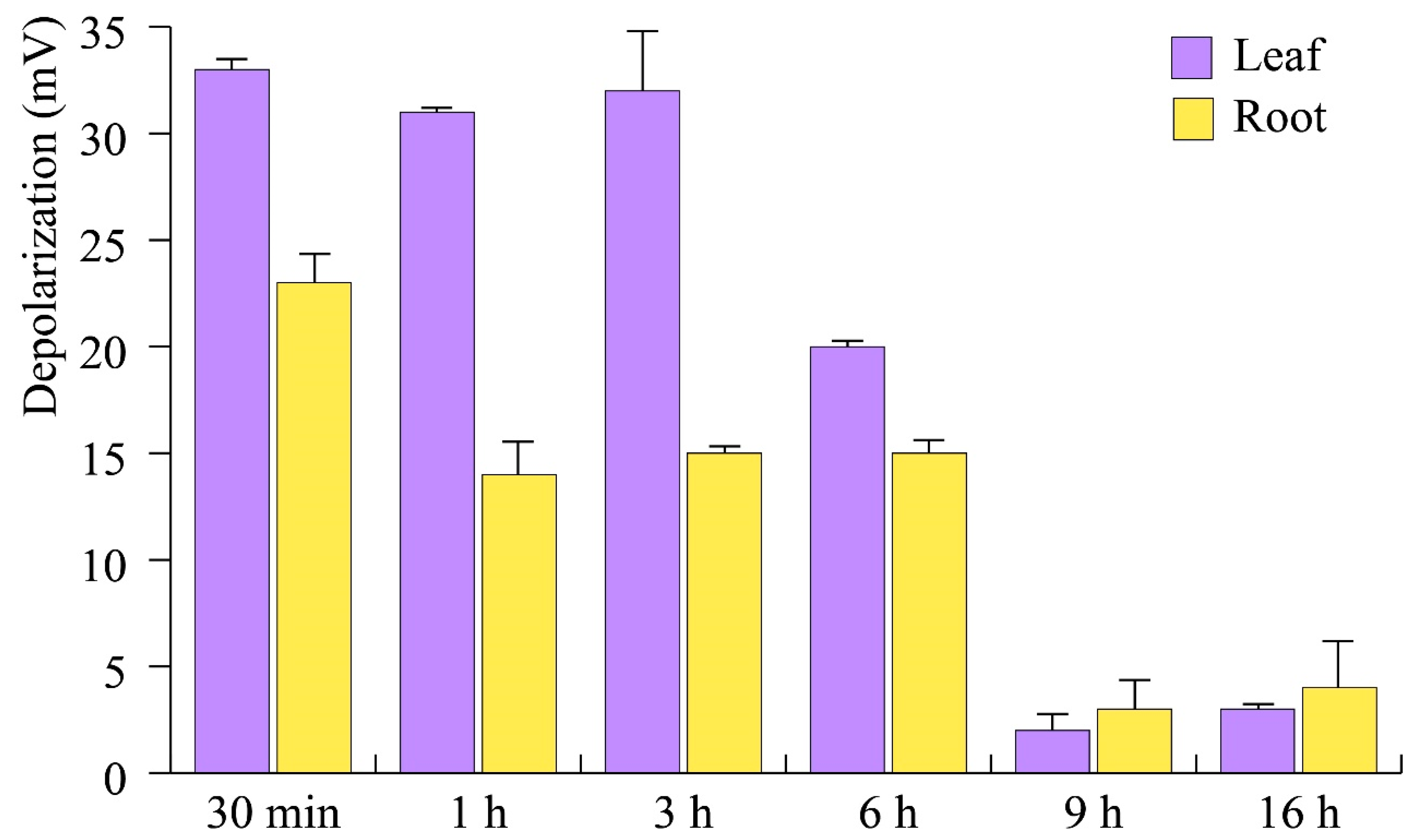
3.3.1. E. amylovora mVOCs Depolarize A. thaliana Plasma Membrane Potential (Vm)
3.3.2. E. amylovora mVOCs Induce Calcium Efflux and Modulate K+ Channels of A. thaliana
3.3.3. E. amylovora mVOCs Induce a Strong Burst of ROS and NO in A. thaliana Seedlings
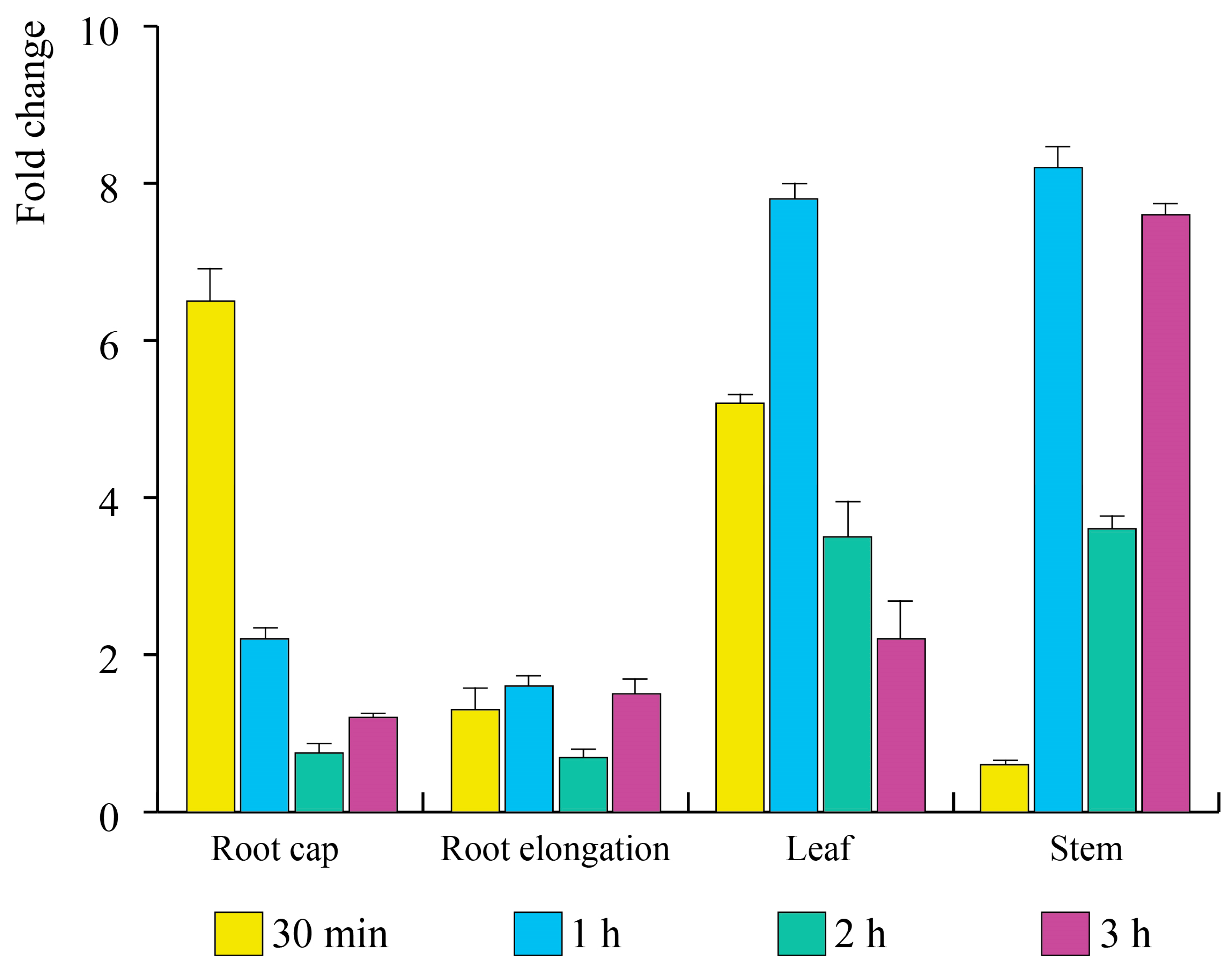
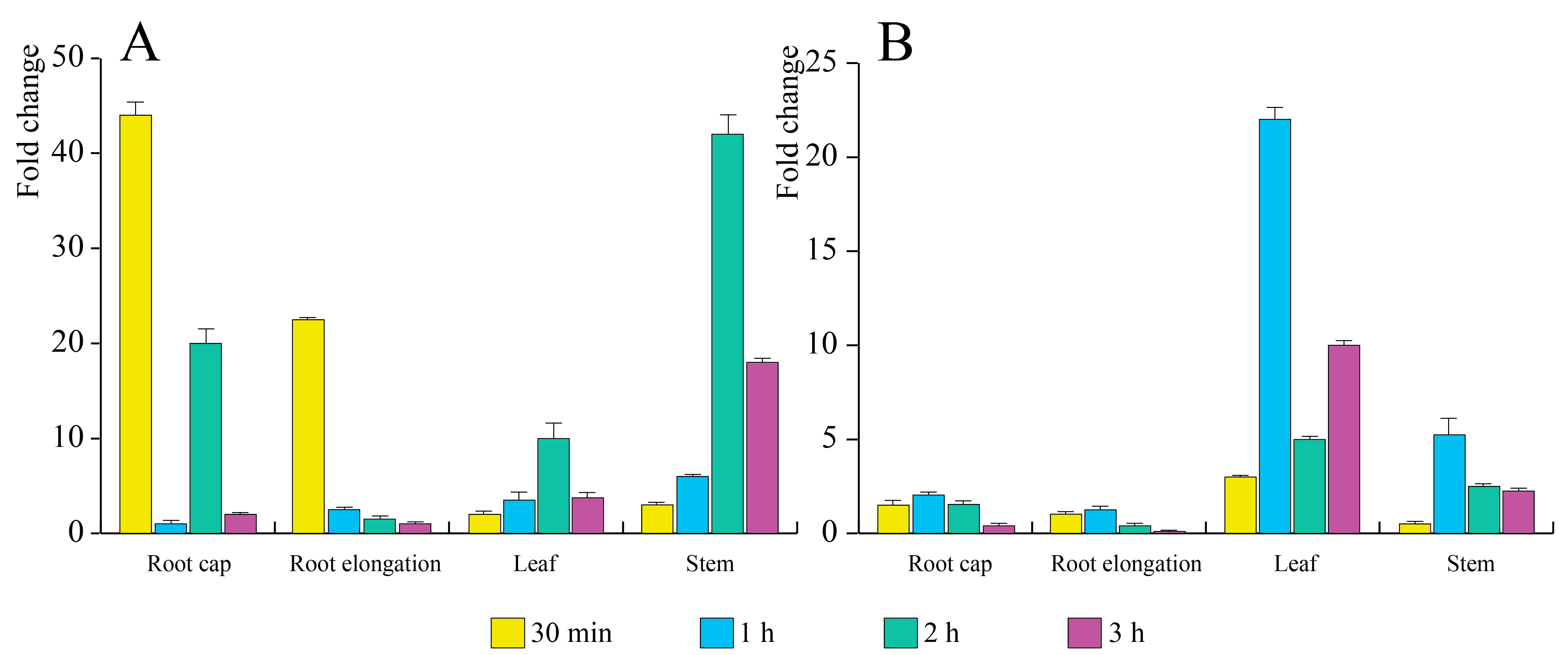
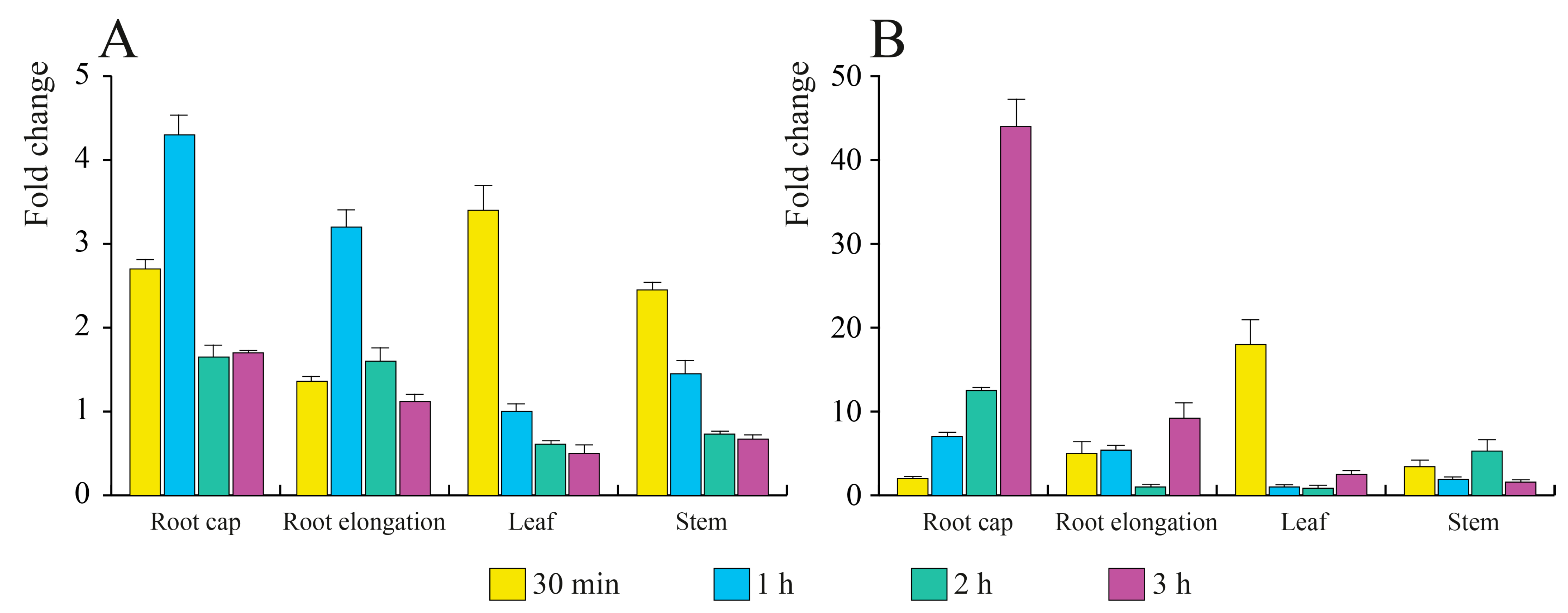
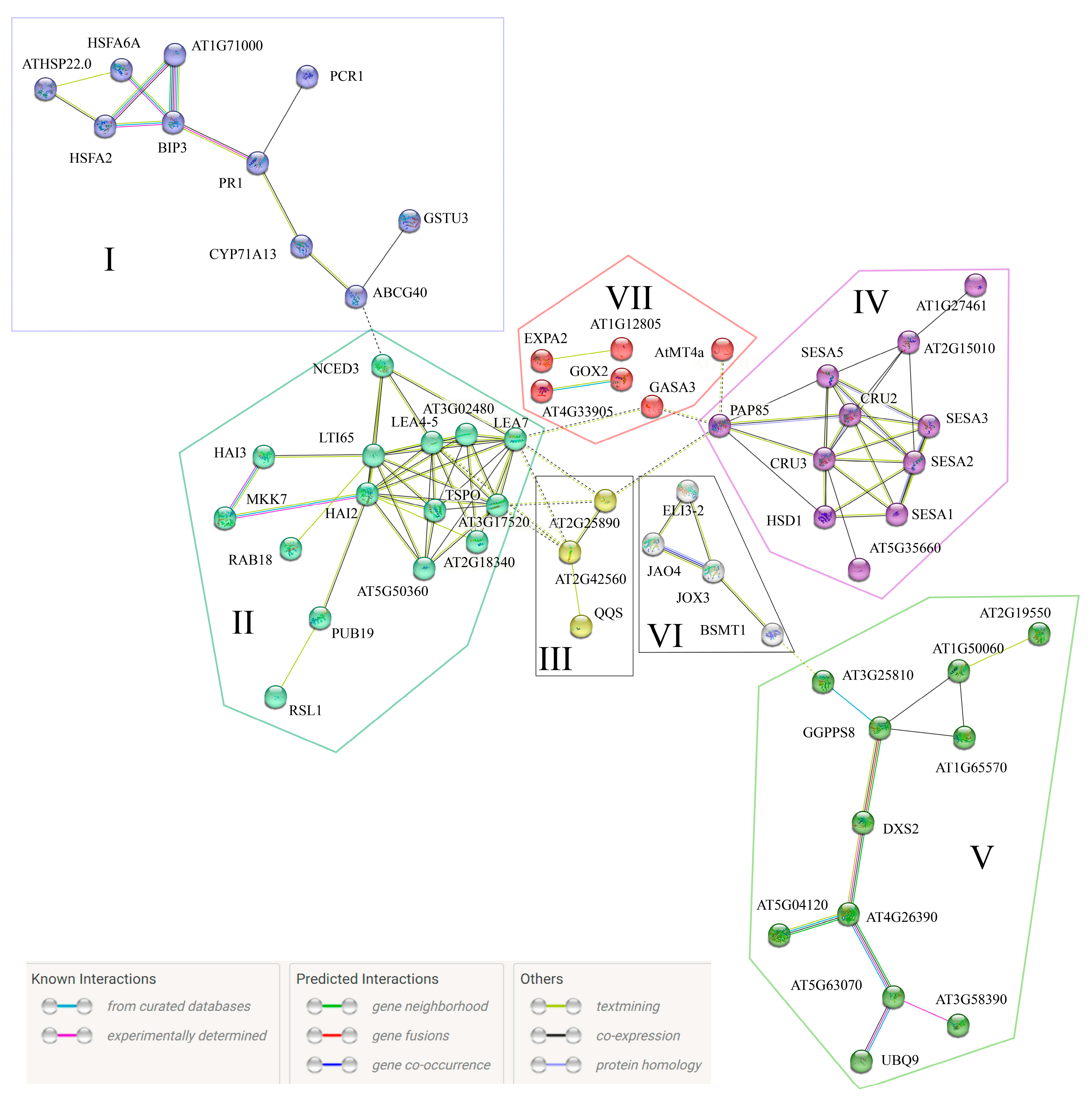
3.4. E. amylovora mVOCs Regulate the Gene Expression of A. thaliana Seedlings
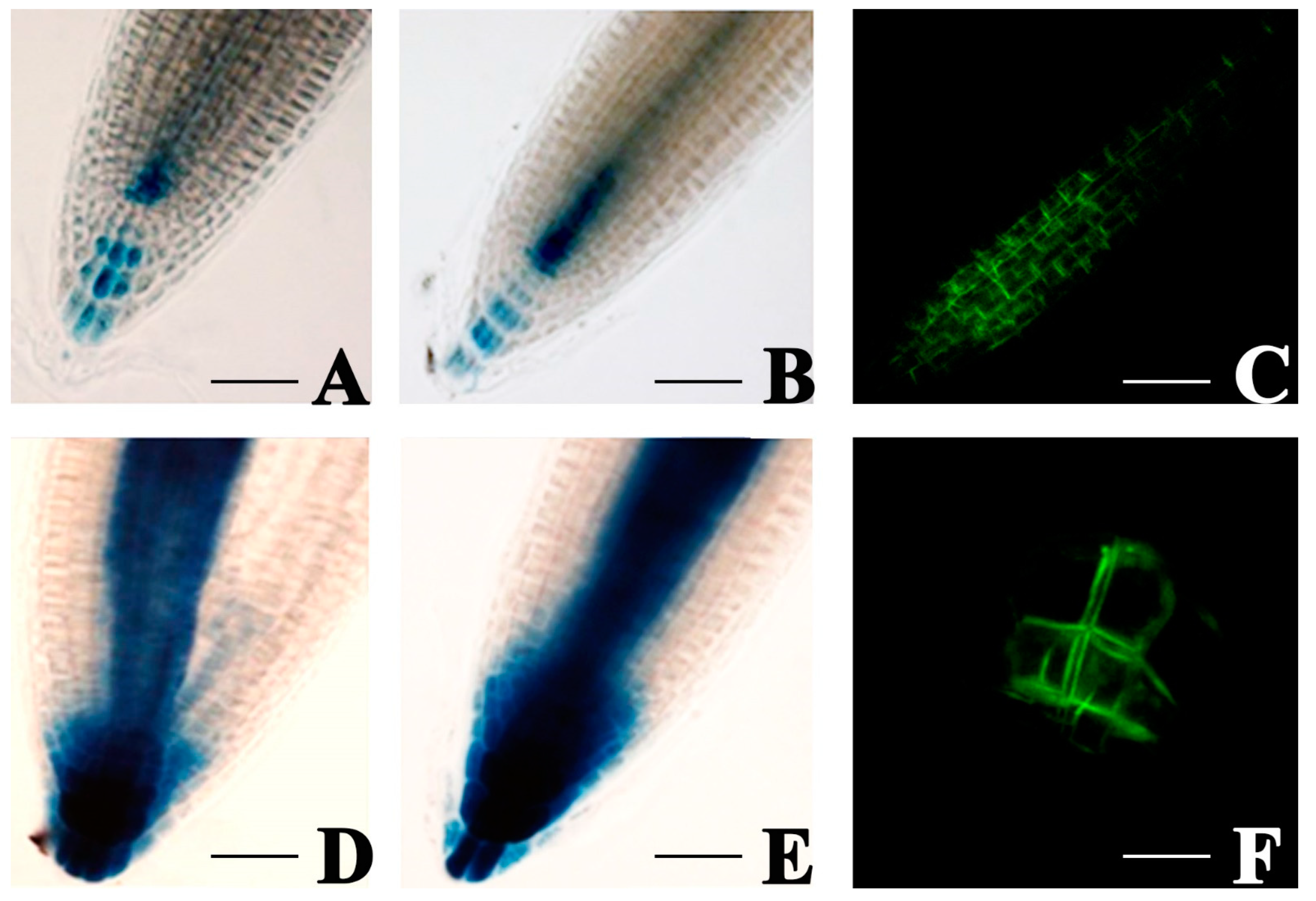
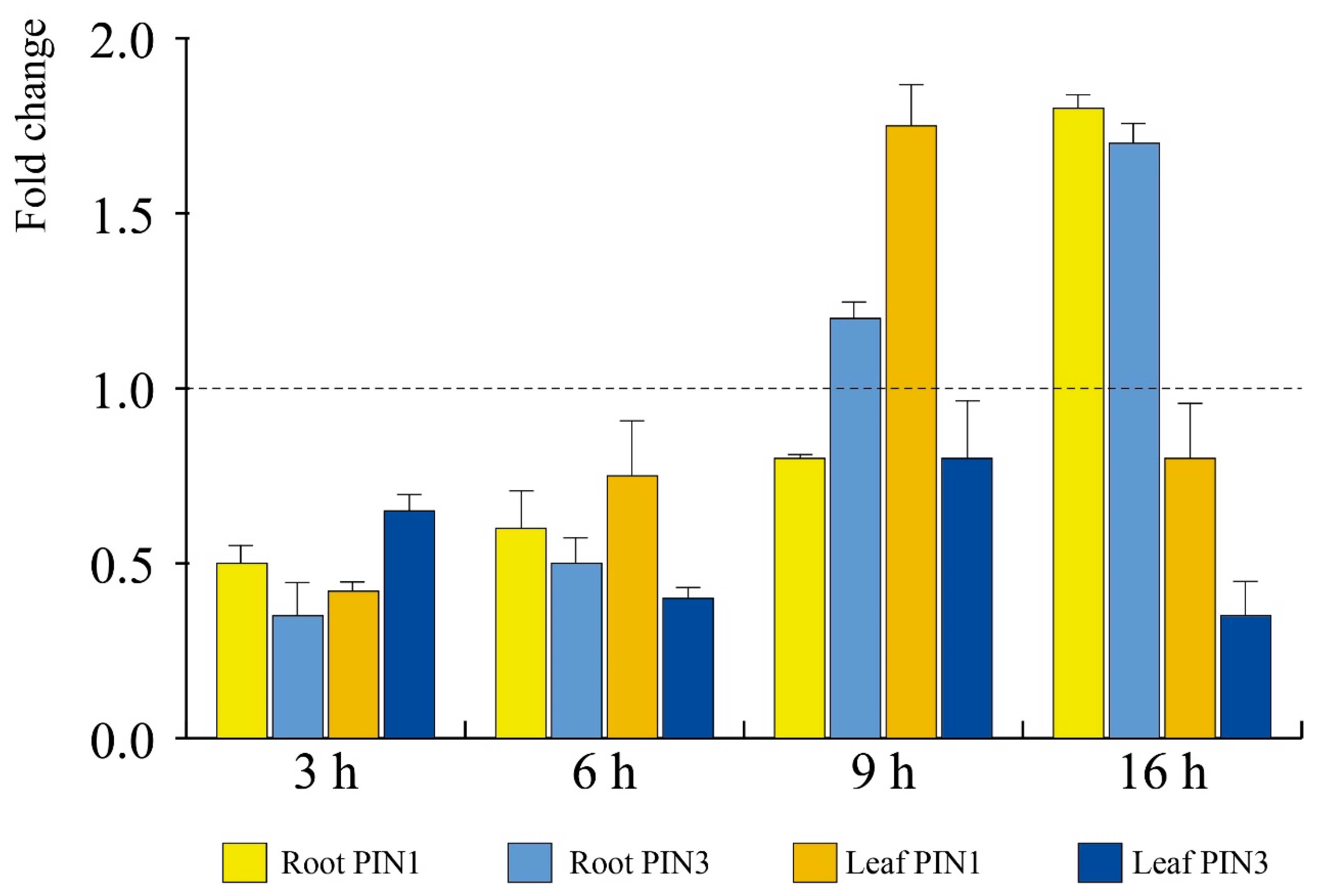
3.5. E. amylovora mVOCs Modulate Auxin Efflux Carriers PIN1 and PIN3 of A. thaliana Roots
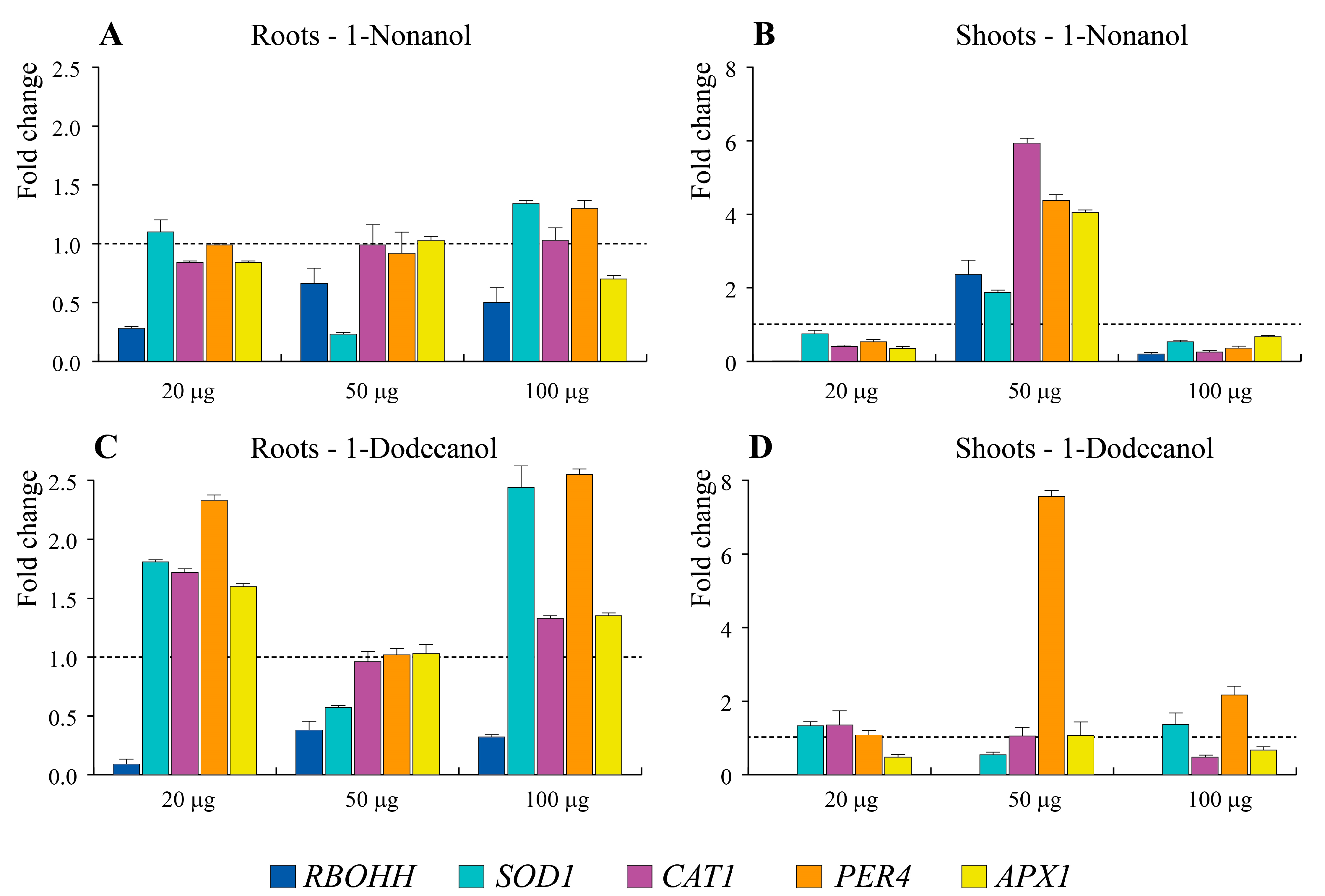
3.6. E. amylovora Synthetic mVOCs 1-nonanol and 1-dodecanol Alter Morphology and Induce Oxidative Stress in A. thaliana Seedlings
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malnoy, M.; Martens, S.; Norelli, J.L.; Barny, M.A.; Sundin, G.W.; Smits, T.H.; Duffy, B. Fire blight: Applied genomic insights of the pathogen and host. Annu. Rev. Phytopathol. 2012, 50, 475–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Hulin, M.T.; Sundin, G.W. Effectors, chaperones, and harpins of the Type III secretion system in the fire blight pathogen Erwinia amylovora: A review. J. Plant Pathol. 2021, 103, 25–39. [Google Scholar] [CrossRef]
- Pineda, A.; Soler, R.; Weldegergis, B.T.; Shimwela, M.M.; JJ, V.A.N.L.; Dicke, M. Non-pathogenic rhizobacteria interfere with the attraction of parasitoids to aphid-induced plant volatiles via jasmonic acid signalling. Plant Cell Environ. 2013, 36, 393–404. [Google Scholar] [CrossRef]
- Park, H.B.; Lee, B.; Kloepper, J.W.; Ryu, C.M. One shot-two pathogens blocked: Exposure of Arabidopsis to hexadecane, a long chain volatile organic compound, confers induced resistance against both Pectobacterium carotovorum and Pseudomonas syringae. Plant Signal. Behav. 2013, 8, e24619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, A.; Weisskopf, L. The modulating effect of bacterial volatiles on plant growth: Current knowledge and future challenges. Plant Signal. Behav. 2012, 7, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Al-Ani, L.K.T.; Franzino, T.; Aguilar-Marcelino, L.; Haichar, F.E.Z.; Furtado, E.L.; Raza, W.; Jatoi, G.H.; Raza, M. Chapter 14—The role of microbial signals in plant growth and development: Current status and future prospects. In New and Future Developments in Microbial Biotechnology and Bioengineering; Rastegari, A.A., Yadav, A.N., Yadav, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 225–242. [Google Scholar]
- Bais, H.P.; Park, S.-W.; Weir, T.L.; Callaway, R.M.; Vivanco, J.M. How plants communicate using the underground information superhighway. Trends Plant Sci. 2004, 9, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Bakker, P.A.; Pieterse, C.M. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maffei, M.E.; Arimura, G.I.; Mithofer, A. Natural elicitors, effectors and modulators of plant responses. Nat. Prod. Rep. 2012, 29, 1288–1303. [Google Scholar] [CrossRef]
- Gallucci, S.; Maffei, M.E. DNA Sensing across the Tree of Life. Trends Immunol. 2017, 38, 719–732. [Google Scholar] [CrossRef]
- Millet, Y.A.; Danna, C.H.; Clay, N.K.; Songnuan, W.; Simon, M.D.; Werck-Reichhart, D.; Ausubel, F.M. Innate Immune Responses Activated in Arabidopsis Roots by Microbe-Associated Molecular Patterns. Plant Cell 2010, 22, 973–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanchiswamy, C.N.; Mainoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Bioprospecting bacterial and fungal volatiles for sustainable agriculture. Trends Plant Sci. 2015, 20, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, M.; Paramasivan, M.; Sahayarayan, J.J. Microbial Volatile Organic Compounds: An Alternative for Chemical Fertilizers in Sustainable Agriculture Development. Microorganisms 2022, 11, 42. [Google Scholar] [CrossRef]
- Ravelo-Ortega, G.; Raya-González, J.; López-Bucio, J. Compounds from rhizosphere microbes that promote plant growth. Curr. Opin. Plant Biol. 2023, 73, 102336. [Google Scholar] [CrossRef]
- Álvarez-García, S.; Manga-Robles, A.; Encina, A.; Gutiérrez, S.; Casquero, P.A. Novel culture chamber to evaluate in vitro plant-microbe volatile interactions: Effects of Trichoderma harzianum volatiles on wheat plantlets. Plant Sci. Int. J. Exp. Plant Biol. 2022, 320, 111286. [Google Scholar] [CrossRef]
- Girón-Calva, P.S.; Molina-Torres, J.; Heil, M. Volatile dose and exposure time impact perception in neighboring plants. J. Chem. Ecol. 2012, 38, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Bitas, V.; Kim, H.S.; Bennett, J.W.; Kang, S. Sniffing on microbes: Diverse roles of microbial volatile organic compounds in plant health. Mol. Plant-Microbe Interact. MPMI 2013, 26, 835–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Splivallo, R.; Bossi, S.; Maffei, M.; Bonfante, P. Discrimination of truffle fruiting body versus mycelial aromas by stir bar sorptive extraction. Phytochemistry 2007, 68, 2584–2598. [Google Scholar] [CrossRef] [PubMed]
- Barbero, F.; Guglielmotto, M.; Islam, M.; Maffei, M.E. Extracellular Fragmented Self-DNA Is Involved in Plant Responses to Biotic Stress. Front. Plant Sci. 2021, 12, 17. [Google Scholar] [CrossRef]
- Mithöfer, A.; Mazars, C.; Maffei, M.E. Probing Spatio-temporal Intracellular Calcium Variations in Plants. Methods Mol. Biol. (Clifton N.J.) 2009, 479, 79–92. [Google Scholar]
- Bricchi, I.; Occhipinti, A.; Bertea, C.M.; Zebelo, S.A.; Brillada, C.; Verrillo, F.; De Castro, C.; Molinaro, A.; Faulkner, C.; Maule, A.J.; et al. Separation of early and late responses to herbivory in Arabidopsis by changing plasmodesmal function. Plant J. 2013, 73, 14–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zebelo, S.A.; Maffei, M.E. The ventral eversible gland (VEG) of Spodoptera littoralis triggers early responses to herbivory in Arabidopsis thaliana. Arthropod-Plant Interact. 2012, 6, 543–551. [Google Scholar] [CrossRef]
- Paponov, I.A.; Fliegmann, J.; Narayana, R.; Maffei, M.E. Differential root and shoot magnetoresponses in Arabidopsis thaliana. Sci. Rep. 2021, 11, 14. [Google Scholar] [CrossRef]
- Brazma, A.; Hingamp, P.; Quackenbush, J.; Sherlock, G.; Spellman, P.; Stoeckert, C.; Aach, J.; Ansorge, W.; Ball, C.A.; Causton, H.C.; et al. Minimum information about a microarray experiment (MIAME)—Toward standards for microarray data. Nat. Genet. 2001, 29, 365–371. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Bawa, G.; Liu, Z.; Wu, R.; Zhou, Y.; Liu, H.; Sun, S.; Liu, Y.; Qin, A.; Yu, X.; Zhao, Z.; et al. PIN1 regulates epidermal cells development under drought and salt stress using single-cell analysis. Front. Plant Sci. 2022, 13, 1043204. [Google Scholar] [CrossRef]
- Gentleman, R.; Carey, V.J.; Huber, W.; Irizarry, R.A.; Dudoit, S. Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Springer: New York, NY, USA, 2006. [Google Scholar]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Gamboa-Becerra, R.; Desgarennes, D.; Molina-Torres, J.; Ramírez-Chávez, E.; Kiel-Martínez, A.L.; Carrión, G.; Ortiz-Castro, R. Plant growth-promoting and non-promoting rhizobacteria from avocado trees differentially emit volatiles that influence growth of Arabidopsis thaliana. Protoplasma 2022, 259, 835–854. [Google Scholar] [CrossRef] [PubMed]
- Phoka, N.; Suwannarach, N.; Lumyong, S.; Ito, S.I.; Matsui, K.; Arikit, S.; Sunpapao, A. Role of Volatiles from the Endophytic Fungus Trichoderma asperelloides PSU-P1 in Biocontrol Potential and in Promoting the Plant Growth of Arabidopsis thaliana. J. Fungi (Basel Switz.) 2020, 6, 341. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Paré, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [Green Version]
- Amavizca, E.; Bashan, Y.; Ryu, C.-M.; Farag, M.A.; Bebout, B.M.; de-Bashan, L.E. Enhanced performance of the microalga Chlorella sorokiniana remotely induced by the plant growth-promoting bacteria Azospirillum brasilense and Bacillus pumilus. Sci. Rep. 2017, 7, 41310. [Google Scholar] [CrossRef]
- Buśko, M.; Kulik, T.; Ostrowska, A.; Góral, T.; Perkowski, J. Quantitative volatile compound profiles in fungal cultures of three different Fusarium graminearum chemotypes. FEMS Microbiol. Lett. 2014, 359, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Blom, D.; Fabbri, C.; Connor, E.C.; Schiestl, F.P.; Klauser, D.R.; Boller, T.; Eberl, L.; Weisskopf, L. Production of plant growth modulating volatiles is widespread among rhizosphere bacteria and strongly depends on culture conditions. Environ. Microbiol. 2011, 13, 3047–3058. [Google Scholar] [CrossRef]
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef] [PubMed]
- Hunziker, L.; Bönisch, D.; Groenhagen, U.; Bailly, A.; Schulz, S.; Weisskopf, L. Pseudomonas Strains Naturally Associated with Potato Plants Produce Volatiles with High Potential for Inhibition of Phytophthora infestans. Appl. Environ. Microbiol. 2015, 81, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Luna, F.M.; López-Bucio, J.; Altamirano-Hernández, J.; Valencia-Cantero, E.; de la Cruz, H.R.; Macías-Rodríguez, L. Plant growth-promoting rhizobacteria modulate root-system architecture in Arabidopsis thaliana through volatile organic compound emission. Symbiosis 2010, 51, 75–83. [Google Scholar] [CrossRef]
- Tait, E.; Perry, J.D.; Stanforth, S.P.; Dean, J.R. Identification of volatile organic compounds produced by bacteria using HS-SPME-GC-MS. J. Chromatogr. Sci. 2014, 52, 363–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ercolini, D.; Russo, F.; Nasi, A.; Ferranti, P.; Villani, F. Mesophilic and psychrotrophic bacteria from meat and their spoilage potential in vitro and in beef. Appl Env. Microbiol 2009, 75, 1990–2001. [Google Scholar] [CrossRef] [Green Version]
- Mauriello, G.; Marino, R.; D’Auria, M.; Cerone, G.; Rana, G.L. Determination of Volatile Organic Compounds from Truffles via SPME-GC-MS. J. Chromatogr. Sci. 2004, 42, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, N.; Valiya Nadakkakath, A.; Munjal, V.; Kundu, A.; Subaharan, K.; Venugopal, V.; Rajamma, S.; Eapen, S.J.; Kumar, A. Genetic analysis of plant endophytic Pseudomonas putida BP25 and chemo-profiling of its antimicrobial volatile organic compounds. Microbiol. Res. 2015, 173, 66–78. [Google Scholar] [CrossRef]
- Brock, N.L.; Tudzynski, B.; Dickschat, J.S. Biosynthesis of sesqui- and diterpenes by the gibberellin producer Fusarium fujikuroi. Chembiochem A Eur. J. Chem. Biol. 2011, 12, 2667–2676. [Google Scholar] [CrossRef] [PubMed]
- Nakano, C.; Kim, H.-K.; Ohnishi, Y. Identification of the First Bacterial Monoterpene Cyclase, a 1,8-Cineole Synthase, that Catalyzes the Direct Conversion of Geranyl Diphosphate. Chembiochem A Eur. J. Chem. Biol. 2011, 12, 1988–1991. [Google Scholar] [CrossRef]
- Piechulla, B.; Schnitzler, J.P. Circumvent CO2 Effects in Volatile-Based Microbe-Plant Interactions. Trends Plant Sci. 2016, 21, 541–543. [Google Scholar] [CrossRef]
- Boller, T. Chemoperception of Microbial Signals in Plant Cells. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 189–214. [Google Scholar] [CrossRef]
- Baker, B.; Zambryski, P.; Staskawicz, B.; Dinesh-Kumar, S.P. Signaling in plant-microbe interactions. Science 1997, 276, 726–733. [Google Scholar] [CrossRef]
- Felle, H.H.; Kondorosi, É.; Kondorosi, Á.; Schultze, M. How Alfalfa Root Hairs Discriminate between Nod Factors and Oligochitin Elicitors. Plant Physiol. 2000, 124, 1373–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mithöfer, A.; Ebel, J.; Felle, H.H. Cation Fluxes Cause Plasma Membrane Depolarization Involved in β-Glucan Elicitor-Signaling in Soybean Roots. Mol. Plant-Microbe Interact. 2005, 18, 983–990. [Google Scholar] [CrossRef] [Green Version]
- THAIN, J.F.; GUBB, I.R.; WILDON, D.C. Depolarization of tomato leaf cells by oligogalacturonide elicitors. Plant Cell Environ. 1995, 18, 211–214. [Google Scholar] [CrossRef]
- Maffei, M.; Camusso, W.; Sacco, S. Effect of Mentha x piperita essential oil and monoterpenes on cucumber root membrane potential. Phytochemistry 2001, 58, 703–707. [Google Scholar] [CrossRef]
- Maffei, M.E.; Gertsch, J.; Appendino, G. Plant volatiles: Production, function and pharmacology. Nat. Prod. Rep. 2011, 28, 1359–1380. [Google Scholar] [CrossRef] [PubMed]
- Lecourieux, D.; Ranjeva, R.; Pugin, A. Calcium in plant defence-signalling pathways. New Phytol. 2006, 171, 249–269. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, S.; Sentenac, H. Plant ion channels: From molecular structures to physiological functions. Curr. Opin. Plant Biol. 1999, 2, 477–482. [Google Scholar] [CrossRef]
- Ivashikina, N.; Becker, D.; Ache, P.; Meyerhoff, O.; Felle, H.H.; Hedrich, R. K(+) channel profile and electrical properties of Arabidopsis root hairs. FEBS Lett. 2001, 508, 463–469. [Google Scholar] [CrossRef] [Green Version]
- El-Maarouf, H.; Barny, M.A.; Rona, J.P.; Bouteau, F. Harpin, a hypersensitive response elicitor from Erwinia amylovora, regulates ion channel activities in Arabidopsis thaliana suspension cells. FEBS Lett. 2001, 497, 82–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadassery, J.; Ranf, S.; Drzewiecki, C.; Mithofer, A.; Mazars, C.; Scheel, D.; Lee, J.; Oelmuller, R. A cell wall extract from the endophytic fungus Piriformospora indica promotes growth of Arabidopsis seedlings and induces intracellular calcium elevation in roots. Plant J. 2009, 59, 193–206. [Google Scholar] [CrossRef]
- Zebelo, S.A.; Matsui, K.; Ozawa, R.; Maffei, M.E. Plasma membrane potential depolarization and cytosolic calcium flux are early events involved in tomato (Solanum lycopersicon) plant-to-plant communication. Plant Sci. 2012, 196, 93–100. [Google Scholar] [CrossRef]
- Zebelo, S.A.; Maffei, M.E. Role of early signalling events in plant-insect interactions. J. Exp. Bot. 2015, 66, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Lamb, C.; Dixon, R.A. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 251–275. [Google Scholar] [CrossRef]
- Vandelle, E.; Ling, T.F.; Imanifard, Z.; Liu, R.T.; Delledonne, M.; Bellin, D. Nitric Oxide Signaling during the Hypersensitive Disease Resistance Response. In Nitric Oxide and Signaling in Plants; Wendehenne, D., Ed.; Academic Press Ltd.-Elsevier Science Ltd.: London, UK, 2016; Volume 77, pp. 219–243. [Google Scholar]
- Glazener, J.A.; Orlandi, E.W.; Baker, C.J. The Active Oxygen Response of Cell Suspensions to Incompatible Bacteria Is Not Sufficient to Cause Hypersensitive Cell Death. Plant Physiol 1996, 110, 759–763. [Google Scholar] [CrossRef] [Green Version]
- Venisse, J.-S.p.; Gullner, G.; Brisset, M.-N.l. Evidence for the Involvement of an Oxidative Stress in the Initiation of Infection of Pear by Erwinia amylovora 1. Plant Physiol. 2001, 125, 2164–2172. [Google Scholar] [CrossRef] [Green Version]
- Launay, A.; Patrit, O.; Wénès, E.; Fagard, M. DspA/E Contributes to Apoplastic Accumulation of ROS in Non-host A. thaliana. Front. Plant Sci. 2016, 7, 545. [Google Scholar] [CrossRef] [Green Version]
- Bellin, D.; Asai, S.; Delledonne, M.; Yoshioka, H. Nitric Oxide as a Mediator for Defense Responses. Mol. Plant-Microbe Interact. 2013, 26, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Liu, G.; Chang, Y.; He, C.; Shi, H. Heat shock transcription factor 3 regulates plant immune response through modulation of salicylic acid accumulation and signalling in cassava. Mol. Plant Pathol. 2018, 19, 2209–2220. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Mittler, R. Could heat shock transcription factors function as hydrogen peroxide sensors in plants? Ann. Bot. 2006, 98, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Berka, M.; Kopecka, R.; Berkova, V.; Brzobohaty, B.; Cerny, M. Regulation of heat shock proteins 70 and their role in plant immunity. J. Exp. Bot. 2022, 73, 1894–1909. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R. Protein Quality Control in the Endoplasmic Reticulum of Plants. Annu. Rev. Plant Biol. 2018, 69, 147–172. [Google Scholar] [CrossRef]
- Park, C.J.; Song, M.Y.; Kim, C.Y.; Jeon, J.S.; Ronald, P.C. Rice BiP3 regulates immunity mediated by the PRRs XA3 and XA21 but not immunity mediated by the NB-LRR protein, Pi5. Biochem. Biophys. Res. Commun. 2014, 448, 70–75. [Google Scholar] [CrossRef]
- Li, C.H.; Cao, S.F.; Wang, K.T.; Lei, C.Y.; Ji, N.N.; Xu, F.; Jiang, Y.B.; Qiu, L.L.; Zheng, Y.H. Heat Shock Protein HSP24 Is Involved in the BABA-Induced Resistance to Fungal Pathogen in Postharvest Grapes Underlying an NPR1-Dependent Manner. Front. Plant Sci. 2021, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, L.; Xing, X.; Sun, L.P.; Pan, J.W.; Kong, X.P.; Zhang, M.Y.; Li, D.Q. ZmLEA3, a Multifunctional Group 3 LEA Protein from Maize (Zea mays L.), is Involved in Biotic and Abiotic Stresses. Plant Cell Physiol. 2013, 54, 944–959. [Google Scholar] [CrossRef] [Green Version]
- Salleh, F.M.; Evans, K.; Goodall, B.; Machin, H.; Mowla, S.B.; Mur, L.A.J.; Runions, J.; Theodoulou, F.L.; Foyer, C.H.; Rogers, H.J. A novel function for a redox-related LEA protein (SAG21/AtLEA5) in root development and biotic stress responses. Plant Cell Environ. 2012, 35, 418–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundertmark, M.; Hincha, D.K. LEA (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genom. 2008, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Lee, Y.; Park, J.; Lee, N.; Choi, G. HONSU, a Protein Phosphatase 2C, Regulates Seed Dormancy by Inhibiting ABA Signaling in Arabidopsis. Plant Cell Physiol. 2013, 54, 555–572. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.W.; Kim, J.-H.; Baek, W.; Kim, B.S.; Lee, S.C. Functional roles of the protein phosphatase 2C, AtAIP1, in abscisic acid signaling and sugar tolerance in Arabidopsis. Plant Sci. 2012, 187, 83–88. [Google Scholar] [CrossRef]
- Yoshida, T.; Nishimura, N.; Kitahata, N.; Kuromori, T.; Ito, T.; Asami, T.; Shinozaki, K.; Hirayama, T. ABA-Hypersensitive Germination3 Encodes a Protein Phosphatase 2C (AtPP2CA) That Strongly Regulates Abscisic Acid Signaling during Germination among Arabidopsis Protein Phosphatase 2Cs. Plant Physiol. 2005, 140, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Guillaumot, D.; Guillon, S.; Déplanque, T.; Vanhee, C.; Gumy, C.; Masquelier, D.; Morsomme, P.; Batoko, H. The Arabidopsis TSPO-related protein is a stress and abscisic acid-regulated, endoplasmic reticulum–Golgi-localized membrane protein. Plant J. 2009, 60, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, M.T.; Akita, M.; Frank, W.; Reski, R.; Valkonen, J.P.T. Involvement of a Class III Peroxidase and the Mitochondrial Protein TSPO in Oxidative Burst Upon Treatment of Moss Plants with a Fungal Elicitor. Mol. Plant-Microbe Interact. 2012, 25, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Trainotti, L.; Li, M.; Varotto, C. Overexpression of Isoprene Synthase Affects ABA- and Drought-Related Gene Expression and Enhances Tolerance to Abiotic Stress. Int. J. Mol. Sci. 2020, 21, 4276. [Google Scholar] [CrossRef]
- Zhang, X.; Dai, Y.; Xiong, Y.; DeFraia, C.; Li, J.; Dong, X.; Mou, Z. Overexpression of Arabidopsis MAP kinase kinase 7 leads to activation of plant basal and systemic acquired resistance. Plant J. Cell Mol. Biol. 2007, 52, 1066–1079. [Google Scholar] [CrossRef]
- Dai, Y.; Wang, H.; Li, B.; Huang, J.; Liu, X.; Zhou, Y.; Mou, Z.; Li, J. Increased expression of MAP KINASE KINASE7 causes deficiency in polar auxin transport and leads to plant architectural abnormality in Arabidopsis. Plant Cell 2006, 18, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Marmagne, A.; Ferro, M.; Meinnel, T.; Bruley, C.; Kuhn, L.; Garin, J.; Barbier-Brygoo, H.; Ephritikhine, G. A High Content in Lipid-modified Peripheral Proteins and Integral Receptor Kinases Features in the <em>Arabidopsis</em> Plasma Membrane Proteome *. Mol. Cell. Proteom. 2007, 6, 1980–1996. [Google Scholar]
- Pei, L.; Peng, L.; Wan, X.; Xiong, J.; Liu, Z.; Li, X.; Yang, Y.; Wang, J. Expression Pattern and Function Analysis of AtPPRT1, a Novel Negative Regulator in ABA and Drought Stress Responses in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 394. [Google Scholar] [CrossRef] [Green Version]
- Bueso, E.; Ibañez, C.; Sayas, E.; Muñoz-Bertomeu, J.; Gonzalez-Guzmán, M.; Rodriguez, P.L.; Serrano, R. A forward genetic approach in Arabidopsis thaliana identifies a RING-type ubiquitin ligase as a novel determinant of seed longevity. Plant Sci. 2014, 215-216, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Bueso, E.; Rodriguez, L.; Lorenzo-Orts, L.; Gonzalez-Guzman, M.; Sayas, E.; Muñoz-Bertomeu, J.; Ibañez, C.; Serrano, R.; Rodriguez, P.L. The single-subunit RING-type E3 ubiquitin ligase RSL1 targets PYL4 and PYR1 ABA receptors in plasma membrane to modulate abscisic acid signaling. Plant J. Cell Mol. Biol. 2014, 80, 1057–1071. [Google Scholar] [CrossRef]
- Krebbers, E.; Herdies, L.; De Clercq, A.; Seurinck, J.; Leemans, J.; Van Damme, J.; Segura, M.; Gheysen, G.; Van Montagu, M.; Vandekerckhove, J. Determination of the Processing Sites of an Arabidopsis 2S Albumin and Characterization of the Complete Gene Family. Plant Physiol 1988, 87, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, B.C.; Xu, Y.; Zhu, Y.X. Systematic studies of 12S seed storage protein accumulation and degradation patterns during Arabidopsis seed maturation and early seedling germination stages. J. Biochem. Mol. Biol. 2007, 40, 373–381. [Google Scholar] [CrossRef]
- McCarty, D.R. Genetic Control and Integration of Maturation and Germination Pathways in Seed Development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 71–93. [Google Scholar] [CrossRef]
- Kagaya, Y.; Okuda, R.; Ban, A.; Toyoshima, R.; Tsutsumida, K.; Usui, H.; Yamamoto, A.; Hattori, T. Indirect ABA-dependent regulation of seed storage protein genes by FUSCA3 transcription factor in Arabidopsis. Plant Cell Physiol. 2005, 46, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Fujita, Y.; Katsura, K.; Maruyama, K.; Narusaka, Y.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional Regulation of ABI3- and ABA-responsive Genes Including RD29B and RD29A in Seeds, Germinating Embryos, and Seedlings of Arabidopsis. Plant Mol. Biol. 2006, 60, 51–68. [Google Scholar] [CrossRef]
- Parcy, F.; Valon, C.; Raynal, M.; Gaubier-Comella, P.; Delseny, M.; Giraudat, J. Regulation of gene expression programs during Arabidopsis seed development: Roles of the ABI3 locus and of endogenous abscisic acid. Plant Cell 1994, 6, 1567–1582. [Google Scholar] [PubMed] [Green Version]
- Furihata, T.; Maruyama, K.; Fujita, Y.; Umezawa, T.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Abscisic acid-dependent multisite phosphorylation regulates the activity of a transcription activator AREB1. Proc. Natl. Acad. Sci. USA 2006, 103, 1988–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sels, J.; Mathys, J.; De Coninck, B.M.; Cammue, B.P.; De Bolle, M.F. Plant pathogenesis-related (PR) proteins: A focus on PR peptides. Plant Physiol. Biochem. PPB 2008, 46, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Otani, M.; Kanno, Y.; Seo, M.; Yoshitake, Y.; Yoshimoto, K.; Sugimoto, K.; Kawakami, N. Seed dormancy 4 like1 of Arabidopsis is a key regulator of phase transition from embryo to vegetative development. Plant J. Cell Mol. Biol. 2022, 112, 460–475. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; D’Auria, J.C.; Farooq, A.; Pichersky, E.; Gershenzon, J. Biosynthesis and emission of terpenoid volatiles from Arabidopsis flowers. Plant Cell 2003, 15, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Beck, G.; Coman, D.; Herren, E.; Ruiz-Sola, M.A.; Rodríguez-Concepción, M.; Gruissem, W.; Vranová, E. Characterization of the GGPP synthase gene family in Arabidopsis thaliana. Plant Mol. Biol. 2013, 82, 393–416. [Google Scholar] [CrossRef] [Green Version]
- Szádeczky-Kardoss, I.; Gál, L.; Auber, A.; Taller, J.; Silhavy, D. The No-go decay system degrades plant mRNAs that contain a long A-stretch in the coding region. Plant Sci. Int. J. Exp. Plant Biol. 2018, 275, 19–27. [Google Scholar] [CrossRef]
- Chiba, Y.; Oshima, K.; Arai, H.; Ishii, M.; Igarashi, Y. Discovery and Analysis of Cofactor-dependent Phosphoglycerate Mutase Homologs as Novel Phosphoserine Phosphatases in <em>Hydrogenobacter thermophilus</em>*. J. Biol. Chem. 2012, 287, 11934–11941. [Google Scholar]
- Carretero-Paulet, L.; Cairó, A.; Talavera, D.; Saura, A.; Imperial, S.; Rodríguez-Concepción, M.; Campos, N.; Boronat, A. Functional and evolutionary analysis of DXL1, a non-essential gene encoding a 1-deoxy-D-xylulose 5-phosphate synthase like protein in Arabidopsis thaliana. Gene 2013, 524, 40–53. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; D’Auria, J.C.; Tholl, D.; Ross, J.R.; Gershenzon, J.; Noel, J.P.; Pichersky, E. An Arabidopsis thaliana gene for methylsalicylate biosynthesis, identified by a biochemical genomics approach, has a role in defense. Plant J. Cell Mol. Biol. 2003, 36, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.P.; Yang, Y.; Pichersky, E.; Klessig, D.F. Altering expression of benzoic acid/salicylic acid carboxyl methyltransferase 1 compromises systemic acquired resistance and PAMP-triggered immunity in arabidopsis. Mol. Plant-Microbe Interact. MPMI 2010, 23, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Kim, K.W.; Cho, M.H.; Franceschi, V.R.; Davin, L.B.; Lewis, N.G. Expression of cinnamyl alcohol dehydrogenases and their putative homologues during Arabidopsis thaliana growth and development: Lessons for database annotations? Phytochemistry 2007, 68, 1957–1974. [Google Scholar] [CrossRef] [PubMed]
- Somssich, I.E.; Wernert, P.; Kiedrowski, S.; Hahlbrock, K. Arabidopsis thaliana defense-related protein ELI3 is an aromatic alcohol:NADP+ oxidoreductase. Proc. Natl. Acad. Sci. United States Am. 1996, 93, 14199–14203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnova, E.; Marquis, V.; Poirier, L.; Aubert, Y.; Zumsteg, J.; Ménard, R.; Miesch, L.; Heitz, T. Jasmonic Acid Oxidase 2 Hydroxylates Jasmonic Acid and Represses Basal Defense and Resistance Responses against <em>Botrytis cinerea</em> Infection. Mol. Plant 2017, 10, 1159–1173. [Google Scholar] [PubMed] [Green Version]
- Caarls, L.; Elberse, J.; Awwanah, M.; Ludwig, N.R.; de Vries, M.; Zeilmaker, T.; Van Wees, S.C.M.; Schuurink, R.C.; Van den Ackerveken, G. Arabidopsis JASMONATE-INDUCED OXYGENASES down-regulate plant immunity by hydroxylation and inactivation of the hormone jasmonic acid. Proc. Natl. Acad. Sci. USA 2017, 114, 6388–6393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, A.; Wu, M.; Yan, L.; Hu, R.; Ali, I.; Gan, Y. AtEXP2 is involved in seed germination and abiotic stress response in Arabidopsis. PLoS ONE 2014, 9, e85208. [Google Scholar] [CrossRef]
- Couée, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Bohnert, H.J. Integration of Arabidopsis thaliana stress-related transcript profiles, promoter structures, and cell-specific expression. Genome Biol. 2007, 8, R49. [Google Scholar] [CrossRef] [Green Version]
- Herzog, M.; Dorne, A.M.; Grellet, F. GASA, a gibberellin-regulated gene family from Arabidopsis thaliana related to the tomato GAST1 gene. Plant Mol. Biol. 1995, 27, 743–752. [Google Scholar] [CrossRef]
- Ren, Y.; Liu, Y.; Chen, H.; Li, G.; Zhang, X.; Zhao, J. Type 4 metallothionein genes are involved in regulating Zn ion accumulation in late embryo and in controlling early seedling growth in Arabidopsis. Plant Cell Environ. 2012, 35, 770–789. [Google Scholar] [CrossRef]
- Takahashi, T.; Mori, T.; Ueda, K.; Yamada, L.; Nagahara, S.; Higashiyama, T.; Sawada, H.; Igawa, T. The male gamete membrane protein DMP9/DAU2 is required for double fertilization in flowering plants. Dev. (Camb. Engl.) 2018, 145, dev170076. [Google Scholar] [CrossRef] [Green Version]
- Mur, L.A.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death. Plant Physiol. 2006, 140, 249–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inzé, A.; Vanderauwera, S.; Hoeberichts, F.A.; Vandorpe, M.; Van Gaever, T.; Van Breusegem, F. A subcellular localization compendium of hydrogen peroxide-induced proteins. Plant Cell Environ. 2012, 35, 308–320. [Google Scholar] [CrossRef]
- Fischer-Kilbienski, I.; Miao, Y.; Roitsch, T.; Zschiesche, W.; Humbeck, K.; Krupinska, K. Nuclear targeted AtS40 modulates senescence associated gene expression in Arabidopsis thaliana during natural development and in darkness. Plant Mol. Biol. 2010, 73, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Castelló, M.J.; Carrasco, J.L.; Navarrete-Gómez, M.; Daniel, J.; Granot, D.; Vera, P. A plant small polypeptide is a novel component of DNA-binding protein phosphatase 1-mediated resistance to plum pox virus in Arabidopsis. Plant Physiol. 2011, 157, 2206–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, C.; Jahrmann, T.; Puigdomenech, P.; Vicient, C.M. Ankyrin repeat-containing proteins in Arabidopsis: Characterization of a novel and abundant group of genes coding ankyrin-transmembrane proteins. Gene 2004, 340, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.; Teale, W.; Fleck, C.; Volpers, M.; Ruperti, B.; Palme, K. Modelling polar auxin transport in developmental patterning. Plant Biol. 2010, 12, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Friml, J.; Benkova, E.; Blilou, I.; Wisniewska, J.; Hamann, T.; Ljung, K.; Woody, S.; Sandberg, G.; Scheres, B.; Jurgens, G.; et al. AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 2002, 108, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Friml, J.; Wiśniewska, J.; Benková, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 415, 806–809. [Google Scholar] [CrossRef] [Green Version]
- Grunewald, W.; Cannoot, B.; Friml, J.; Gheysen, G. Parasitic Nematodes Modulate PIN-Mediated Auxin Transport to Facilitate Infection. PLoS Pathog. 2009, 5, e1000266. [Google Scholar] [CrossRef] [Green Version]
- Kidd, B.N.; Kadoo, N.Y.; Dombrecht, B.; Tekeoglu, M.; Gardiner, D.M.; Thatcher, L.F.; Aitken, E.A.B.; Schenk, P.M.; Manners, J.M.; Kazan, K. Auxin Signaling and Transport Promote Susceptibility to the Root-Infecting Fungal Pathogen Fusarium oxysporum in Arabidopsis. Mol. Plant-Microbe Interact. 2011, 24, 733–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meents, A.K.; Furch, A.C.U.; Almeida-Trapp, M.; Ozyurek, S.; Scholz, S.S.; Kirbis, A.; Lenser, T.; Theissen, G.; Grabe, V.; Hansson, B.; et al. Beneficial and Pathogenic Arabidopsis Root-Interacting Fungi Differently Affect Auxin Levels and Responsive Genes During Early Infection. Front. Microbiol. 2019, 10, 14. [Google Scholar] [CrossRef]
- Hwang, H.-H.; Chien, P.-R.; Huang, F.-C.; Yeh, P.-H.; Hung, S.-H.W.; Deng, W.-L.; Huang, C.-C. A Plant Endophytic Bacterium Priestia megaterium StrainBP-R2 Isolated from the Halophyte Bolboschoenus planiculmis Enhances Plant Growth under Salt and Drought Stresses. Microorganisms 2022, 10, 2047. [Google Scholar] [CrossRef]
- Pasternak, T.; Palme, K.; Pérez-Pérez, J.M. Role of reactive oxygen species in the modulation of auxin flux and root development in Arabidopsis thaliana. Plant J. 2023. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.M.; Muhlemann, J.K.; Gayornba, S.R.; Muday, G.K. RBOH-Dependent ROS Synthesis and ROS Scavenging by Plant Specialized Metabolites To Modulate Plant Development and Stress Responses. Chem. Res. Toxicol. 2019, 32, 370–396. [Google Scholar] [CrossRef]
- Valério, L.; De Meyer, M.; Penel, C.; Dunand, C. Expression analysis of the Arabidopsis peroxidase multigenic family. Phytochemistry 2004, 65, 1331–1342. [Google Scholar] [CrossRef]
- Rhee, S.G. H2O2, a Necessary Evil for Cell Signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef]
- Kaya, H.; Nakajima, R.; Iwano, M.; Kanaoka, M.M.; Kimura, S.; Takeda, S.; Kawarazaki, T.; Senzaki, E.; Hamamura, Y.; Higashiyama, T.; et al. Ca2+-Activated Reactive Oxygen Species Production by Arabidopsis RbohH and RbohJ Is Essential for Proper Pollen Tube Tip Growth. Plant Cell 2014, 26, 1069–1080. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.L.; Cao, X.Y.; Wang, X.Y.; Zhang, W.; Li, W.X.; Wang, X.Q.; Liu, S.Q.; Lyu, D.G. RBOH-dependent hydrogen peroxide signaling mediates melatonin-induced anthocyanin biosynthesis in red pear fruit. Plant Sci. 2021, 313, 11. [Google Scholar] [CrossRef]
- Araki, R.; Mermod, M.; Yamasaki, H.; Kamiya, T.; Fujiwara, T.; Shikanai, T. SPL7 locally regulates copper-homeostasis-related genes in Arabidopsis. J. Plant Physiol. 2018, 224-225, 137–143. [Google Scholar] [CrossRef]
- Sunkar, R.; Kapoor, A.; Zhu, J.-K. Posttranscriptional Induction of Two Cu/Zn Superoxide Dismutase Genes in Arabidopsis Is Mediated by Downregulation of miR398 and Important for Oxidative Stress Tolerance. Plant Cell 2006, 18, 2051–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gou, X.; Zhong, C.; Zhang, P.; Mi, L.; Li, Y.; Lu, W.; Zheng, J.; Xu, J.; Meng, Y.; Shan, W. miR398b and AtC2GnT form a negative feedback loop to regulate Arabidopsis thaliana resistance against Phytophthora parasitica. Plant J. 2022, 111, 360–373. [Google Scholar] [CrossRef]
- Rasul, S.; Dubreuil-Maurizi, C.; Lamotte, O.; Koen, E.; Poinssot, B.; Alcaraz, G.; Wendehenne, D.; Jeandroz, S. Nitric oxide production mediates oligogalacturonide-triggered immunity and resistance to Botrytis cinerea in Arabidopsis thaliana. Plant Cell Environ. 2012, 35, 1483–1499. [Google Scholar] [CrossRef]
- Wei, S.-S.; Niu, W.-T.; Zhai, X.-T.; Liang, W.-Q.; Xu, M.; Fan, X.; Lv, T.-T.; Xu, W.-Y.; Bai, J.-T.; Jia, N.; et al. Arabidopsis mtHSC70-1 plays important roles in the establishment of COX-dependent respiration and redox homeostasis. J. Exp. Bot. 2019, 70, 5575–5590. [Google Scholar] [CrossRef] [PubMed]
- Kvaratskhelia, M.; George, S.J.; Thorneley, R.N.F. Salicylic Acid Is a Reducing Substrate and Not an Effective Inhibitor of Ascorbate Peroxidase*. J. Biol. Chem. 1997, 272, 20998–21001. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Kovàts Retention Index | Relative Percentage |
|---|---|---|
| Iso amyl alcohol | 697 | 1.21 |
| Styrene | 883 | 4.04 |
| 6-Methyl-2-heptanone | 888 | 0.93 |
| 4-Octen-3-one | 960 | 2.91 |
| Benzaldehyde | 982 | 1.13 |
| Octanal | 1005 | 1.08 |
| Tetraisobutylene | 1030 | 1.93 |
| (E)-2-Octenal | 1036 | 1.96 |
| 1,8-Cineole | 1058 | 0.31 |
| 1-octanol | 1065 | 2.58 |
| Nonanal | 1104 | 1.32 |
| 1-Nonanol | 1159 | 3.03 |
| Dichloromethyl propyl sulfone | 1172 | 5.54 |
| trans-2-Decenal | 1212 | 2.16 |
| Methylcyclodecane | 1260 | 3.33 |
| trans-2-Decenol | 1266 | 2.36 |
| Undecanal | 1303 | 2.32 |
| Tridecane | 1313 | 1.08 |
| 6,10-Dimethyl-2-undecanone | 1321 | 8.32 |
| 1-Undecanol | 1357 | 2.08 |
| 2-Ethyl-1-decanol | 1393 | 0.39 |
| 1-Tetradecene | 1403 | 0.94 |
| E-2-dodecenal | 1410 | 0.48 |
| Tetradecane | 1413 | 1.21 |
| 1-Dodecanol | 1457 | 10.10 |
| Tridecanal | 1502 | 2.21 |
| Pentadecane | 1512 | 11.73 |
| Tetradecanal | 1601 | 1.38 |
| Tetradecanal | 1601 | 2.12 |
| 2-Pentadecanone | 1648 | 1.23 |
| 1-Tetradecanol | 1656 | 1.94 |
| Pentadecanal | 1701 | 6.19 |
| Heptadecane | 1711 | 2.68 |
| 6,10,14-Trimethyl-2-pentadecanone | 1754 | 6.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parmagnani, A.S.; Kanchiswamy, C.N.; Paponov, I.A.; Bossi, S.; Malnoy, M.; Maffei, M.E. Bacterial Volatiles (mVOC) Emitted by the Phytopathogen Erwinia amylovora Promote Arabidopsis thaliana Growth and Oxidative Stress. Antioxidants 2023, 12, 600. https://doi.org/10.3390/antiox12030600
Parmagnani AS, Kanchiswamy CN, Paponov IA, Bossi S, Malnoy M, Maffei ME. Bacterial Volatiles (mVOC) Emitted by the Phytopathogen Erwinia amylovora Promote Arabidopsis thaliana Growth and Oxidative Stress. Antioxidants. 2023; 12(3):600. https://doi.org/10.3390/antiox12030600
Chicago/Turabian StyleParmagnani, Ambra S., Chidananda Nagamangala Kanchiswamy, Ivan A. Paponov, Simone Bossi, Mickael Malnoy, and Massimo E. Maffei. 2023. "Bacterial Volatiles (mVOC) Emitted by the Phytopathogen Erwinia amylovora Promote Arabidopsis thaliana Growth and Oxidative Stress" Antioxidants 12, no. 3: 600. https://doi.org/10.3390/antiox12030600