
Sexual Dimorphism in the Expression of Cardiac and Hippocampal Renin-Angiotensin and Kallikrein–Kinin Systems in Offspring from Mice Exposed to Alcohol during Gestation


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. PAE Protocol
2.3. Biological Samples
2.4. Quantitative PCR
2.4.1. RNA Extraction
2.4.2. Reverse Transcription
2.4.3. Quantitative Real-Time Polymerization Chain Reaction (PCR)—qPCR
2.5. ROS Measurement
2.6. Statistical Analysis
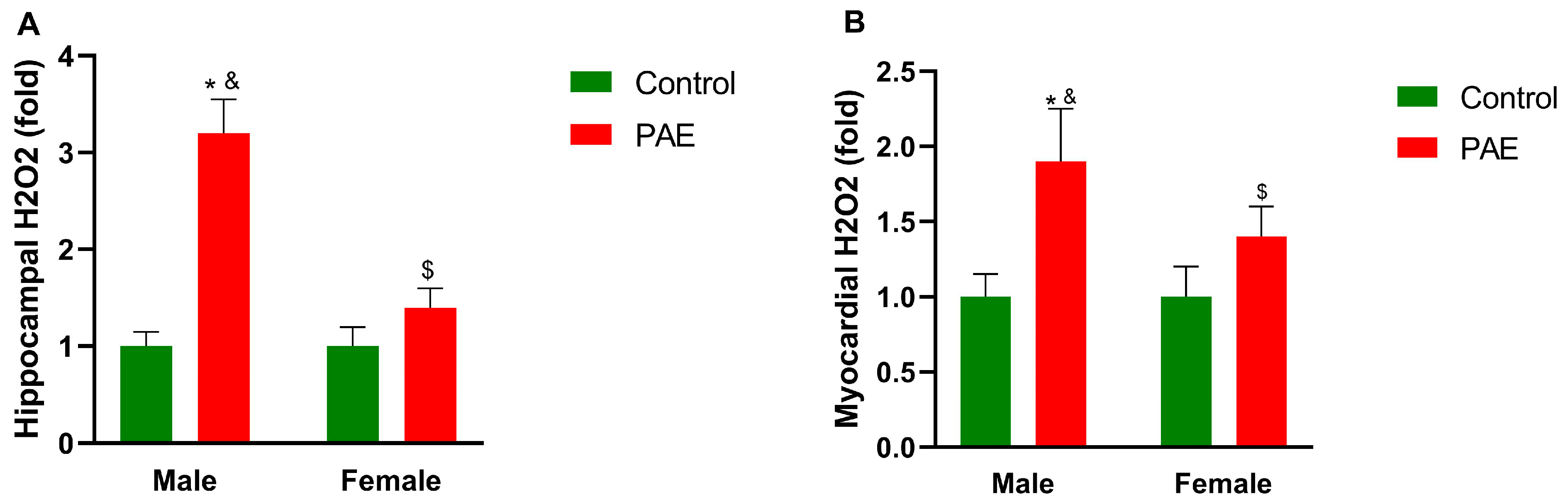
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pinder, R.M.; Sandler, M. Alcohol, wine and mental health: Focus on dementia and stroke. J. Psychopharmacol. 2004, 18, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Panza, F.; Frisardi, V.; Seripa, D.; Logroscino, G.; Santamato, A.; Imbimbo, B.P.; Scafato, E.; Pilotto, A.; Solfrizzi, V. Alcohol consumption in mild cognitive impairment and dementia: Harmful or neuroprotective? Int. J. Geriatr. Psychiatry 2012, 27, 1218–1238. [Google Scholar] [CrossRef] [PubMed]
- Roerecke, M.; Rehm, J. Alcohol consumption, drinking patterns, and ischemic heart disease: A narrative review of meta-analyses and a systematic review and meta-analysis of the impact of heavy drinking occasions on risk for moderate drinkers. BMC Med. 2014, 12, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braillon, A.; Wilson, M. Does moderate alcohol consumption have health benefits? BMJ 2018, 362, k3888. [Google Scholar] [PubMed]
- Day, E.; Rudd, J.H.F. Alcohol use disorders and the heart. Addiction 2019, 114, 1670–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, R.; Sheron, N. No level of alcohol consumption improves health. Lancet 2018, 392, 987–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsiki, N.; Tziomalos, K.; Mikhailidis, D.P. Alcohol and the cardiovascular system: A double-edged sword. Curr. Pharm. Des. 2014, 20, 6276–6288. [Google Scholar] [CrossRef]
- Roerecke, M. Alcohol’s Impact on the Cardiovascular System. Nutrients 2021, 13, 3419. [Google Scholar] [CrossRef]
- WHO. Reproductive, maternal, newborn, child and adolescent health. In Global Status Report on Alcohol and Health 2018; Poznyak, V., Rekve, D., Eds.; World Health Organization: Geneva, Switzerland, 2018; pp. 5–6. [Google Scholar]
- Popova, S.; Lange, S.; Probst, C.; Gmel, G.; Rehm, J. Estimation of national, regional, and global prevalence of alcohol use during pregnancy and fetal alcohol syndrome: A systematic review and meta-analysis. Lancet Global Health 2017, 5, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Larkby, C.; Day, N. The effects of prenatal alcohol exposure. Alcohol Health Res. World. 1997, 21, 192–198. [Google Scholar]
- Popova, S.; Dozet, D.; Shield, K.; Rehm, J.; Burd, L. Alcohol’s Impact on the Fetus. Nutrients 2021, 13, 3452. [Google Scholar] [CrossRef]
- Jones, K.L.; Smith, D.W. Recognition of the fetal alcohol syndrome in early infancy. Lancet 1973, 302, 999–1001. [Google Scholar] [CrossRef] [PubMed]
- Doremus-Fitzwater, T.L.; Youngentob, S.L.; Youngentob, L.; Gano, A.; Vore, A.S.; Deak, T. Lingering Effects of Prenatal Alcohol Exposure on Basal and Ethanol-Evoked Expression of Inflammatory-Related Genes in the CNS of Adolescent and Adult Rats. Front. Behav. Neurosci. 2020, 20, 14–82. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.T.; Fritchie, G.E.; Idanpaan-Heikkila, J.E.; McIsaac, W.M. Placental transfer and tissue distribution of ethanol-1-14C; A radioautographic study in monkeys and hamsters. Q. J. Stud. Alcohol. 1972, 33, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Livy, D.J.; Miller, E.K.; Maier, S.E.; West, J.R. Fetal alcohol exposure and temporal vulnerability: Effects of binge-like alcohol exposure on the developing rat hippocampus. Neurotoxicol. Teratol. 2003, 25, 447–458. [Google Scholar] [CrossRef]
- Ikonomidou, C.; Bittigau, P.; Ishimaru, M.J.; Wozniak, D.F.; Koch, C.; Genz, K.; Price, M.T.; Stefovska, V.; Horster, F.; Tenkova, T.; et al. Ethanol-induced apoptotic neurodegeneration and fetal alcohol syndrome. Science 2000, 287, 1056–1060. [Google Scholar] [CrossRef]
- Miki, T.; Harris, S.J.; Wilce, P.A.; Takeuchi, Y.; Bedi, K.S. Effects of age and alcohol exposure during early life on pyramidal cell numbers in the CA1–CA3 region of the rat hippocampus. Hippocampus 2004, 14, 124–134. [Google Scholar] [CrossRef]
- Nixon, K.; Crews, F.T. Binge ethanol exposure decreases neurogenesis in adult rat hippocampus. J. Neurochem. 2002, 83, 1087–1093. [Google Scholar] [CrossRef]
- Parkington, H.C.; Coleman, H.A.; Wintour, E.M.; Tare, M. Prenatal alcohol exposure: Implications for cardiovascular function in the fetus and beyond. Clin. Exp. Pharmacol. Physiol. 2010, 37, 91–98. [Google Scholar] [CrossRef]
- Ren, J.; Wold, L.E.; Natavio, M.; Ren, B.H.; Hannigan, J.H.; Brown, R.A. Influence of prenatal alcohol exposure on myocardial contractile function in adult rat hearts: Role of intracellular calcium and apoptosis. Alcohol Alcohol. 2002, 37, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Atum, A.L.B.; Matos, L.P.; Jesus, B.C.; Nasuk, G.R.; Silva, G.A.; Gomes, C.P.; Pesquero, J.B.; Zamuner, S.R.; Silva, J.A., Jr. Prenatal alcohol exposure induced morphometric changes and accentuated transcriptional changes of genes related to cardiac dysfunction in adult mice. Antioxidants 2023, 12, 256. [Google Scholar] [CrossRef]
- Ferreira, A.J.; Santos, R.A.; Bradford, C.N.; Mecca, A.P.; Sumners, C.; Katovich, M.J.; Raizada, M.K. Therapeutic implications of the vasoprotective axis of the renin-angiotensin system in cardiovascular diseases. Hypertension 2010, 55, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ganten, D.; Lang, R.E.; Archelos, J.; Unger, T. Peptidergic systems: Effects on blood vessels. J. Cardiovasc. Pharmacol. 1984, 6, 598–607. [Google Scholar] [CrossRef]
- Sumners, C.; Horiuchi, M.; Widdop, R.E.; McCarthy, C.; Unger, T.; Steckelings, U.M. Protective arms of the renin-angiotensin-system in neurological disease. Clin. Exp. Pharmacol. Physiol. 2013, 40, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Regoli, D.; Plante, G.E.; Gobeil, F., Jr. Impact of kinins in the treatment of cardiovascular diseases. Pharmacol. Ther. 2012, 135, 94–111. [Google Scholar] [CrossRef] [PubMed]
- Raidoo, D.M.; Bhoola, K.D. Pathophysiology of the kallikrein-kinin system in mammalian nervous tissue. Pharmacol. Ther. 1998, 79, 105–127. [Google Scholar] [CrossRef] [PubMed]
- Díaz, H.S.; Toledo, C.; Andrade, D.C.; Marcus, N.J.; Del Rio, R. Neuroinflammation in heart failure: New insights for an old disease. J. Physiol. 2020, 598, 33–59. [Google Scholar] [CrossRef]
- Kamal, H.; Tan, G.C.; Ibrahim, S.F.; Shaikh, M.F.; Mohamed, I.N.; Mohamed, R.M.; Hamid, A.A.; Ugusman, A.; Kumar, J. Alcohol use disorder, neurodegeneration, Alzheimer’s and Parkinson’s disease: Interplay between oxidative stress, Neuroimmune response, and excitotoxicity. Front. Cell. Neurosci. 2020, 14, 282. [Google Scholar] [CrossRef]
- Grupp, L.A. Alcohol satiety, hypertension and the renin-angiotensin system. Med. Hypoth. 1987, 24, 11–19. [Google Scholar] [CrossRef]
- Puddey, I.B.; Vandogen, R.; Beilin, L.G.; Rouse, I.L. Alcohol stimulation of renin release in man: Its relation to hemodynamic, electrolyte and sympathoadrenal response to drinking. J. Clin. Endocrinol. Metab. 1985, 61, 37–42. [Google Scholar] [CrossRef]
- Grupp, L.A.; Perlanski, E.; Stewart, R.B. Regulation of alcohol consumption by the renin-angiotensin system: A review of recent findings and a possible mechanism of action. Neurosci. Biobehav. Rev. 1991, 15, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Maschke, J.; Roetner, J.; Bösl, S.; Plank, A.C.; Rohleder, N.; Goecke, T.W.; Fasching, P.A.; Beckmann, M.W.; Kratz, O.; Moll, G.H.; et al. Association of Prenatal Alcohol Exposure and Prenatal Maternal Depression with Offspring Low-Grade Inflammation in Early Adolescence. Int. J. Environ. Res. Public Health 2021, 18, 7920. [Google Scholar] [CrossRef] [PubMed]
- Akers, K.G.; Kushner, S.A.; Leslie, A.T.; Clarke, L.; Van der Kooy, E.; Lerch, J.P.; Frankland, P.W. Fetal alcohol exposure leads to abnormal olfactory bulb development and impaired odor discrimination in adult mice. Mol. Brain 2011, 4, 2–15. [Google Scholar] [CrossRef] [Green Version]
- Allan, A.M.; Chynoweth, J.; Tyler, L.A.; Caldwell, K.K. A mouse model of prenatal ethanol exposure using a voluntary drinking paradigm. Alcohol. Clin. Exp. Res. 2003, 27, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitts, D.A. Forebrain circumventricular organs mediate captopril-enhanced ethanol intake in rats. Pharmacol. Biochem. Behav. 1993, 45, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zuo, N.; Li, B.; Xiong, Y.; Chen, H.; He, H.; Sun, Z.; Hu, S.; Cheng, H.; Ao, Y.; et al. The expressional disorder of the renal RAS mediates nephrotic syndrome of male rat offspring induced by prenatal ethanol exposure. Toxicology 2018, 400–401, 9–19. [Google Scholar] [CrossRef]
- Gouveia, T.L.; Frangiotti, M.I.; de Brito, J.M.; de Castro Neto, E.F.; Sakata, M.M.; Febba, A.C.; Casarini, D.E.; Amado, D.; Cavalheiro, E.A.; Almeida, S.S.; et al. The levels of renin-angiotensin related components are modified in the hippocampus of rats submitted to pilocarpine model of epilepsy. Neurochem. Int. 2012, 61, 54–62. [Google Scholar] [CrossRef]
- Liu, J.; Feener, E.P. Plasma kallikrein-kinin system and diabetic retinopathy. Biol. Chem. 2013, 394, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.J.; Lorell, B.H.; Ingelfinger, J.R.; Weinberg, E.O.; Schunkert, H.; Diamant, D.; Tang, S.S. Distribution and function of cardiac angiotensin AT1- and AT2-receptor subtypes in hypertrophied rat hearts. Am. J. Physiol. 1994, 267, 844–852. [Google Scholar] [CrossRef]
- Meggs, L.G.; Coupet, J.; Huang, H.; Cheng, W.; Li, P.; Capasso, J.M.; Homcy, C.J.; Anversa, P. Regulation of angiotensin II receptors on ventricular myocytes after myocardial infarction in rats. Circ. Res. 1993, 72, 1149–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.L.; Zhao, L.X.; Yang, J.; Tong, N.; An, L.; Liu, Q.T.; Xie, M.R.; Li, C.S. Association between ACE2/ACE balance and pneumocyte apoptosis in a porcine model of acute pulmonary thromboembolism with cardiac arrest. Mol. Med. Rep. 2018, 17, 4221–4228. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, S.; Tavasoli, M.; Jamali, F. Inflammation Alters Angiotensin Converting Enzymes (ACE and ACE-2) Balance in Rat Heart. Inflammation 2010, 34, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Cao, X.; Qiao, S.; Yu, S.; Li, L.; Yu, Y.; Fu, C.; Jiang, F.; Dong, B.; Su, Q. (Pro)renin Receptor is Involved in Myocardial Damage in Alcoholic Cardiomyopathy. Alcohol. Clin. Exp. Res. 2019, 43, 2344–2353. [Google Scholar] [CrossRef] [PubMed]
- Medina, D.; Mehay, D.; Arnold, A.C. Sex differences in cardiovascular actions of the renin-angiotensin system. Clin. Auton. Res. 2020, 30, 393–408. [Google Scholar] [CrossRef]
- Regitz-Zagrosek, V.; Kararigas, G. Mechanistic Pathways of Sex Differences in Cardiovascular Disease. Physiol. Rev. 2017, 97, 1–37. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, P.E.; Li, P.; Lenhart, J.R.; Chappell, M.C.; Brosnihan, K.B. Estrogen regulation of angiotensin-converting enzyme mRNA. Hypertension 1999, 33, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Bukowska, A.; Spiller, L.; Wolke, C.; Lendeckel, U.; Weinert, S.; Hoffmann, J.; Bornfleth, P.; Kutschka, I.; Gardemann, A.; Isermann, B.; et al. Protective regulation of the ACE2/ACE gene expression by estrogen in human atrial tissue from elderly men. Exp. Biol. Med. 2017, 242, 1412–1423. [Google Scholar] [CrossRef] [Green Version]
- Komukai, K.; Mochizuki, S.; Yoshimura, M. Gender and the renin-angiotensin-aldosterone system. Fundam. Clin. Pharmacol. 2010, 24, 687–698. [Google Scholar] [CrossRef]
- Fischer, M.; Baessler, A.; Schunkert, H. Renin angiotensin system and gender differences in the cardiovascular system. Cardiovasc. Res. 2002, 53, 672–677. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, M.A.; Sullivan, J.C. Hypertension: What’s sex got to do with it? Physiology 2013, 28, 234–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilliard, L.M.; Sampson, A.K.; Brown, R.D.; Denton, K.M. The “his and hers” of the renin-angiotensin system. Curr. Hypertens Rep. 2013, 15, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Sampson, A.K.; Moritz, K.M.; Denton, K.M. Postnatal ontogeny of angiotensin receptors and ACE2 in male and female rats. Gend. Med. 2012, 9, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, X.; Qu, H.Y.; Jiang, S.; Zhang, J.; Fu, L.; Buggs, J.; Pang, B.; Wei, J.; Liu, R. Role of Kidneys in Sex Differences in Angiotensin II-Induced Hypertension. Hypertension 2017, 70, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, A.; Blacklock, A.; Svojanovsky, S.; Smith, P.G. Estrogen elicits dorsal root ganglion axon sprouting via a renin-angiotensin system. Endocrinology 2008, 149, 3452–3460. [Google Scholar] [CrossRef] [PubMed]
- Landazuri, P.; Granobles, C.; Loango, N. Gender differences in serum angiotensin-converting enzyme activity and blood pressure in children: An observational study. Arq. Bras. Cardiol. 2008, 91, 352–357. [Google Scholar] [PubMed] [Green Version]
- Zapater, P.; Novalbos, J.; Gallego-Sandín, S.; Hernández, F.T.; Abad-Santos, F. Gender differences in angiotensin-converting enzyme (ACE) activity and inhibition by enalaprilat in healthy volunteers. J. Cardiovasc. Pharmacol. 2004, 43, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Gembardt, F.; Sterner-Kock, A.; Imboden, H.; Spalteholz, M.; Reibitz, F.; Schultheiss, H.P.; Siems, W.E.; Walther, T. Organ-specific distribution of ACE2 mRNA and correlating peptidase activity in rodents. Peptides 2005, 26, 1270–1277. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, P.; Liang, T.; Chen, Y.; Liu, D.; Yu, H. Relationship between circulating levels of angiotensin-converting enzyme 2-angiotensin-(1-7)-MAS axis and coronary heart disease. Heart Vessel. 2020, 35, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Gupte, M.; Thatcher, S.E.; Boustany-Kari, C.M.; Shoemaker, R.; Yiannikouris, F.; Zhang, X.; Karounos, M.; Cassis, L.A. Angiotensin converting enzyme 2 contributes to sex differences in the development of obesity hypertension in C57BL/6 mice. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1392–1399. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, G.; Delarue, F.; Burcklé, C.; Bouzhir, L.; Giller, T.; Sraer, J.D. Pivotal role of the renin/prorenin receptor in angiotensin II production and cellular responses to renin. J. Clin. Investig. 2002, 109, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Contrepas, A.; Walker, J.; Koulakoff, A.; Franek, K.J.; Qadri, F.; Giaume, C.; Corvol, P.; Schwartz, C.E.; Nguyen, G. A role of the (pro)renin receptor in neuronal cell differentiation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Schunkert, H.; Danser, A.H.; Hense, H.W.; Derkx, F.H.; Kurzinger, S.; Riegger, G.A. Effects of estrogen replacement therapy on the renin–angiotensin system in post-menopausal women. Circulation 1997, 95, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Danser, A.H.; Derkx, F.H.; Schalekamp, M.A.; Hence, H.W.; Riegger, G.A.; Schunkert, H. Determinants of interindividual variation of renin and prorenin concentrations: Evidence for a sexual dimorphism of (pro)renin levels in humans. J. Hypertens. 1998, 16, 853–862. [Google Scholar] [CrossRef]
- Pendergrass, K.D.; Pirro, N.T.; Westwood, B.M.; Ferrario, C.M.; Brosnihan, K.B.; Chappell, M.C. Sex differences in circulating and renal angiotensins of hypertensive mRen(2).Lewis but not normotensive Lewis rats. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H10–H20. [Google Scholar] [CrossRef] [Green Version]
- Pratt, R.E.; Dzau, V.J.; Ouellette, A.J. Influence of androgen on translatable renin mRNA in the mouse submandibular gland. Hypertension 1984, 6, 605–613. [Google Scholar] [CrossRef] [Green Version]
- Madeddu, P.; Pinna Parpaglia, P.; Anania, V.; Glorioso, N.; Chao, C.; Wang, C.; Chao, J. Sexual dimorphism of cardiovascular responses to early blockade of bradykinin receptors. Hypertension 1996, 27, 746–751. [Google Scholar] [CrossRef]
- Tschöpe, C.; Heringer-Walther, S.; Koch, M.; Spillmann, F.; Wendorf, M.; Hauke, D.; Bader, M.; Schultheiss, H.P.; Walther, T. Myocardial bradykinin B2-receptor expression at different time points after induction of myocardial infarction. J. Hypertens. 2000, 18, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Perosa, S.R.; Argañaraz, G.A.; Goto, E.M.; Costa, L.G.; Konno, A.C.; Varella, P.P.; Santiago, J.F.; Pesquero, J.B.; Canzian, M.; Amado, D.; et al. Kinin B1 and B2 receptors are overexpressed in the hippocampus of humans with temporal lobe epilepsy. Hippocampus 2007, 17, 26–33. [Google Scholar] [CrossRef]
- Argañaraz, G.A.; Silva, J.A., Jr.; Perosa, S.R.; Pessoa, L.G.; Carvalho, F.F.; Bascands, J.L.; Bader, M.; da Silva Trindade, E.; Amado, D.; Cavalheiro, E.A.; et al. The synthesis and distribution of the kinin B1 and B2 receptors are modified in the hippocampus of rats submitted to pilocarpine model of epilepsy. Brain Res. 2004, 1006, 114–125. [Google Scholar] [CrossRef]
- Pesquero, J.B.; Araujo, R.C.; Heppenstall, P.A.; Stucky, C.L.; Silva, J.A.; Jr Walther, T.; Oliveira, S.M.; Pesquero, J.L.; Paiva, A.C.; Calixto, J.B.; et al. Hypoalgesia and altered inflammatory responses in mice lacking kinin B1 receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 8140–8145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marceau, F.; Hess, J.F.; Bachvarov, D.R. The B1 receptors for kinins. Pharmacol. Rev. 1998, 50, 357–386. [Google Scholar] [PubMed]
- Tschöpe, C.; Spillmann, F.; Altmann, C.; Koch, M.; Westermann, D.; Dhayat, N.; Dhayat, S.; Bascands, J.L.; Gera, L.; Hoffmann, S.; et al. The bradykinin B1 receptor contributes to the cardioprotective effects of AT1 blockade after experimental myocardial infarction. Cardiovasc. Res. 2004, 61, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Flores-Monroy, J.; Lezama-Martínez, D.; Fonseca-Coronado, S.; Martínez-Aguilar, L. Differences in the expression of the renin angiotensin system and the kallikrein-kinin system during the course of myocardial infarction in male and female Wistar rats. J. Renin Angiotensin Aldosterone Syst. 2020, 21, 1470320319900038. [Google Scholar] [CrossRef] [PubMed]
- Theobald, D.; Sriramula, S. Kinin B1 Receptor Mediates Bidirectional Interaction between Neuroinflammation and Oxidative Stress. Antioxidants 2023, 12, 150. [Google Scholar] [CrossRef]
- Iwadate, H.; Kudo, M.; Kizuki, K. Expression of tissue kallikrein mRNA in developing rat brains. Can. J. Physiol. Pharmacol. 2002, 80, 245–248. [Google Scholar] [CrossRef]
- Yao, Y.Y.; Yin, H.; Shen, B.; Chao, L.; Chao, J. Tissue kallikrein infusion prevents cardiomyocyte apoptosis, inflammation and ventricular remodeling after myocardial infarction. Regul. Pept. 2007, 140, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Iwadate, H.; Kawamata, K.; Kudo, M.; Kizuki, K. Biochemical and immunohistochemical demonstration of tissue kallikrein in the neuronal nuclei of the developing rat brains. Brain Res. 2000, 863, 87–93. [Google Scholar] [CrossRef]
- Simões, P.S.; Perosa, S.R.; Arganãraz, G.A.; Yacubian, E.M.; Carrete, H., Jr.; Centeno, R.S.; Varella, P.P.; Santiago, J.F.; Canzian, M.; Silva, J.A., Jr.; et al. Kallikrein 1 is overexpressed by astrocytes in the hippocampus of patients with refractory temporal lobe epilepsy, associated with hippocampal sclerosis. Neurochem. Int. 2011, 58, 477–482. [Google Scholar] [CrossRef]
- Silva, J.A., Jr.; Araujo, R.C.; Baltatu, O.; Oliveira, S.M.; Tschöpe, C.; Fink, E.; Hoffmann, S.; Plehm, R.; Chai, K.X.; Chao, L.; et al. Reduced cardiac hypertrophy and altered blood pressure control in transgenic rats with the human tissue kallikrein gene. FASEB J. 2000, 14, 1858–1860. [Google Scholar] [CrossRef] [Green Version]
- Chao, J.; Yin, H.; Gao, L.; Hagiwara, M.; Shen, B.; Yang, Z.R.; Chao, L. Tissue kallikrein elicits cardioprotection by direct kinin b2 receptor activation independent of kinin formation. Hypertension 2008, 52, 715–720. [Google Scholar] [CrossRef]
- Smith, A.M.; Zeve, D.R.; Grisel, J.J.; Chen, W.J. Neonatal alcohol exposure increases malondialdehyde (MDA) and glutathione (GSH) levels in the developing cerebellum. Brain Res. Dev. Brain Res. 2005, 160, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Tong, M.; de la Monte, S.M. Chronic ethanol exposure causes mitochondrial dysfunction and oxidative stress in immature central nervous system neurons. Acta Neuropathol. 2007, 113, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Dennery, P.A. Effects of oxidative stress on embryonic development. Birth Defects Res. C Embryo Today 2007, 81, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Drever, N.; Yin, H.; Kechichian, T.; Costantine, M.; Longo, M.; Saade, G.R.; Bytautiene, E. The expression of antioxidant enzymes in a mouse model of fetal alcohol syndrome. Am. J. Obstet. Gynecol. 2012, 206, 358.e19–358.e22. [Google Scholar] [CrossRef] [Green Version]
- Theofanous, T.; Kourti, M. Abrogating Oxidative Stress as a Therapeutic Strategy Against Parkinson’s Disease: A Mini Review of the Recent Advances on Natural Therapeutic Antioxidant and Neuroprotective Agents. Med. Chem. 2022, 18, 772–783. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.; Li, Y.; Peng, Y. A review of interventions against fetal alcohol spectrum disorder targeting oxidative stress. Int. J. Dev. Neurosci. 2018, 71, 140–145. [Google Scholar] [CrossRef]
- Bergamini, C.M.; Gambetti, S.; Dondi, A.; Cervellati, C. Oxygen, reactive oxygen species and tissue damage. Curr. Pharm. Des. 2004, 10, 1611–1626. [Google Scholar] [CrossRef]
- Brocardo, P.S.; Gil-Mohapel, J.; Christie, B.R. The role of oxidative stress in fetal alcohol spectrum disorders. Brain Res. Rev. 2011, 67, 209–225. [Google Scholar] [CrossRef]
- Khalifa, A.R.; Abdel-Rahman, E.A.; Mahmoud, A.M.; Ali, M.H.; Noureldin, M.; Saber, S.H.; Mohsen, M.; Ali, S.S. Sex-specific differences in mitochondria biogenesis, morphology, respiratory function, and ROS homeostasis in young mouse heart and brain. Physiol. Rep. 2017, 5, 13125. [Google Scholar] [CrossRef]
- Borras, C.; Sastre, J.; Garcia-Sala, D.; Lloret, A.; Pallardo, F.V.; Vina, J. Mitochondria from females exhibit higher antioxidant gene expression and lower oxidative damage than males. Free Radic. Biol. Med. 2003, 34, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Michel, M.C.; Wieland, T.; Tsujimoto, G. How reliable are G-protein-coupled receptor antibodies? Naunyn-Schmiedeberg’s Arch. Pharmacol. 2009, 379, 385–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heusch, G.; Rose, J.; Ehring, T. Cardioprotection by ACE inhibitors in myocardial ischaemia/reperfusion. The importance of bradykinin. Drugs 1997, 54, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Tao, M.X.; Xue, X.; Gao, L.; Lu, J.L.; Zhou, J.S.; Jiang, T.; Zhang, Y.D. Involvement of angiotensin-(1-7) in the neuroprotection of captopril against focal cerebral ischemia. Neurosci. Lett. 2018, 687, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Sahin, B.; Ergul, M. Captopril exhibits protective effects through anti-inflammatory and anti-apoptotic pathways against hydrogen peroxide-induced oxidative stress in C6 glioma cells. Metab. Brain Dis. 2022, 37, 1221–1230. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Control | P.A.E. | |||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| AT1 | 4.72 ± 0.51 | 3.48 ± 0.39 a | 8.78 ± 1.23 aa | 4.28 ± 0.32 b,c |
| AT2 | 0.92 ± 0.21 | 1.57 ± 0.31 a | 1.21 ± 0.16 | 2.44 ± 0.41 b,c |
| REN | 3.43 ± 0.28 | 2.96 ± 0.21 | 5.82 ± 0.86 a | 3.53 ± 0.30 b,c |
| ACE | 3.79 ± 0.81 | 3.99 ± 0.54 | 7.63 ± 0.92 aa | 4.27 ± 0.41 c |
| ACE2 | 1.30 ± 0.33 | 2.49 ± 0.23 a | 3.28 ± 0.34 a | 4.19 ± 0.46 b,c |
| Control | P.A.E. | |||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| AT1 | 3.43 ± 0.41 | 1.92 ± 0.53 | 9.91 ± 1.46 aa | 3.43 ± 0.69 b,c |
| AT2 | 1.01 ± 0.18 | 1.34 ± 0.31 a | 1.30 ± 0.15 a | 2.77 ± 0.46 b,c |
| REN | 2.71 ± 0.72 | 2.19 ± 0.83 | 2.94 ± 0.68 | 2.63 ± 0.33 |
| ACE | 4.12 ± 0.84 | 3.64 ± 0.27 a | 5.68 ± 0.50 a | 4.28 ± 0.23 b,c |
| ACE2 | 3.01 ± 0.17 | 3.38 ± 0.21 a | 2.93 ± 0.57 a | 4.87 ± 0.77 b,c |
| Control | P.A.E. | |||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| B1 | 0.32 ± 0.27 | 0.43 ± 0.40 | 3.67 ± 0.43 a | 1.31 ± 0.11 b,cc |
| B2 | 2.73 ± 0.23 | 3.28 ± 0.32 | 3.31 ± 0.17 | 4.29 ± 0.35 b,c |
| KLK | 2.47 ± 0.37 | 3.65 ± 0.90 | 3.89 ± 0.81 a | 4.98 ± 0.33 b |
| Control | P.A.E. | |||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| B1 | 0.86 ± 0.21 | 0.99 ± 0.40 | 1.12 ± 0.48 | 1.07 ± 0.37 |
| B2 | 2.18 ± 0.69 | 2.29 ± 0.41 | 2.32 ± 0.34 | 5.39 ± 0.64 b,cc |
| KLK | 3.48 ± 0.28 | 4.09 ± 0.37 | 3.90 ± 0.92 a | 4.95 ± 0.36 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, G.A.; Atum, A.L.B.; de Matos, L.P.; Nasuk, G.R.; de Jesus, B.C.; Gouveia, T.L.F.; Baltatu, O.C.; Zamuner, S.R.; Silva Júnior, J.A. Sexual Dimorphism in the Expression of Cardiac and Hippocampal Renin-Angiotensin and Kallikrein–Kinin Systems in Offspring from Mice Exposed to Alcohol during Gestation. Antioxidants 2023, 12, 541. https://doi.org/10.3390/antiox12030541
da Silva GA, Atum ALB, de Matos LP, Nasuk GR, de Jesus BC, Gouveia TLF, Baltatu OC, Zamuner SR, Silva Júnior JA. Sexual Dimorphism in the Expression of Cardiac and Hippocampal Renin-Angiotensin and Kallikrein–Kinin Systems in Offspring from Mice Exposed to Alcohol during Gestation. Antioxidants. 2023; 12(3):541. https://doi.org/10.3390/antiox12030541
Chicago/Turabian Styleda Silva, Gabriel Almeida, Allan Luís Barboza Atum, Leonardo Paroche de Matos, Guilherme Rabelo Nasuk, Bruna Calixto de Jesus, Telma Luciana Furtado Gouveia, Ovidiu Constantin Baltatu, Stella Regina Zamuner, and José Antônio Silva Júnior. 2023. "Sexual Dimorphism in the Expression of Cardiac and Hippocampal Renin-Angiotensin and Kallikrein–Kinin Systems in Offspring from Mice Exposed to Alcohol during Gestation" Antioxidants 12, no. 3: 541. https://doi.org/10.3390/antiox12030541