
m6A Modification—Association with Oxidative Stress and Implications on Eye Diseases

Abstract
:1. Introduction
2. m6A Modification
2.1. Writers
2.2. Erasers
2.3. Readers
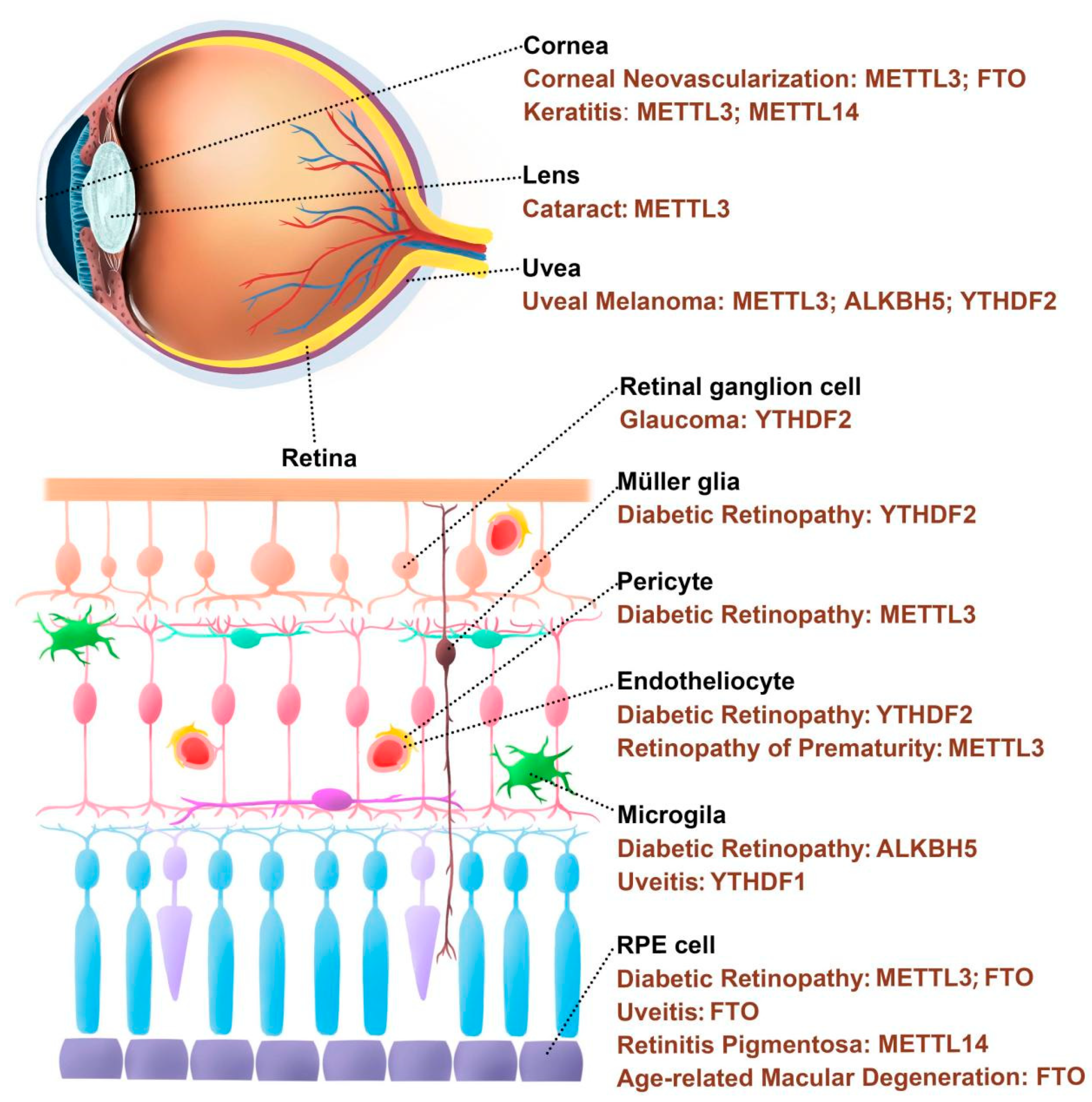
3. m6A Modification and Oxidative Stress-Related Eye Diseases
3.1. Corneal Disease
3.2. Cataract
3.3. Glaucoma
3.4. Uveitis
3.5. Retinopathy
3.5.1. Diabetic Retinopathy (DR)
3.5.2. Retinopathy of Prematurity (ROP)
3.5.3. Retinitis Pigmentosa (RP)
3.5.4. Age-Related Macular Degeneration (AMD)
3.6. Traumatic Optic Neuropathy (TON)
3.7. Ocular Tumor

{kind=link}
| Eye Diseases | Total m6A Level | Enzymes | Functions |
|---|---|---|---|
| Corneal Neovascularization | N/A | METTL3 | Targets stem cell regulatory factors Ahnak and Ddit4 to regulate limbal stem cell proliferation and migration [41]. |
| Down | FTO | Increases the expression of FAK to promote vascular endothelial cell function and angiogenesis [39]. | |
| Cataract | Up | METTL3 | Regulates the lens epithelial cells proliferation and apoptosis by targeting the 3’UTR of ICAM-1 to stabilize mRNA stability [54]. |
| Glaucoma | N/A | YTHDF2 | Targets Hspa12a and Islr2 to affect the tolerance of RGC to acute intraocular hypertension [67]. |
| Uveitis | Down | YTHDF1 | Increases Sirt1 expression thus restraining M1 polarization and migration in microglia [74]. |
| N/A | FTO | Promotes ATF4 expression via both m6A-dependent manner and PERK pathway, thereby reducing the inflammatory response [75]. | |
| Diabetic Retinopathy | N/A | METTL3 | Targets miR-25-3p/PTEN/AKT signaling cascade to enhance RPE cell viability [93]. |
| Up | METTL3 | Reduces PKC-η, FAT4 and PDGFRA mRNA expression thus aggravating pericyte dysfunction [90]. | |
| N/A | FTO | Promotes the mRNA stability of NLRP3 to activate pyroptosis in RPE cells [94]. | |
| N/A | ALKBH5 | Protects A20 mRNA from degradation to reduce M1 microglia polarization [92]. | |
| N/A | YTHDF2 | Promotes the instability of IGTB1 mRNA which is a positive regulator of the FAK/PI3K/AKT signaling pathway [91]. | |
| Retinopathy of Prematurity | Up | METTL3 | Regulates Wnt signaling activation by targeting LRP6 and DVL1 [40]. |
| Retinitis Pigmentosa | Down | METTL14 | Inhibits expression of MAP2 which interacts with NEUROD1 to induce pathologic changes in RPE cells [104]. |
| Age-related Macular Degeneration | N/A | FTO | Rescues RPE from degeneration by inhibiting PKA/CREB signaling [109]. |
| Ocular Melanoma | Down | METTL3, ALKBH5 | Regulates the translation of the tumor suppressor gene HINT2 to regulate ocular melanoma tumorigenesis [117]. |
| N/A | ALKBH5 | Increases FOXM1 expression to promote invasion, migration, and epithelial–mesenchymal transition in UM cells [118]. | |
| Up | METTL3 | Enhances UM cell proliferation, migration, and invasion by promoting c-Met translation [119]. | |
| N/A | METTL3 | Increases RNA and protein level of BACE2, which accelerates tumorigenesis via TMEM38B/Ca2+ pathway [120]. | |
| N/A | YTHDF2 | Promotes degradation of PER1 and TP53 mRNA, thereby accelerating tumorigenesis of ocular melanoma [121]. | |
| Retinoblastoma | N/A | METTL3 | Promotes RB progression in vivo and in vitro through PI3K/AKT/mTOR pathway [126]. |
4. Treatment
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviation
| AAV | adeno-associated virus |
| AGEs | advanced glycation end products |
| AH | aqueous humor |
| ALKBH5 | alkB homolog 5 |
| AMD | age-related macular degeneration |
| AML | acute myeloid leukemia |
| AOPP | advanced oxidation protein product |
| ARCC | age-related cortical cataract |
| BACE2 | beta-secretase 2 |
| CAT | catalases |
| CM | conjunctival melanoma |
| DDIT4 | DNA damage inducible transcript 4 |
| DR | diabetic retinopathy |
| DVL1 | dishevelled segment polarity protein 1 |
| eIF3 | eukaryotic initiation factor 3 |
| EPO | erythropoietin |
| f6A | N6-formyladenosine |
| FAK | focal adhesion kinase |
| FAT4 | FAT atypical cadherin 4 |
| FOXM1 | forkhead box M1 |
| FTO | fat mass and obesity-associated protein |
| GPXs | glutathione peroxidases |
| GSCs | glioma stem cells |
| GSH | glutathione |
| GSTs | glutathione S-transferases |
| HINT2 | histidine triad nucleotide binding protein 2 |
| hm5C | 5-hydroxymethylcytosine |
| HNRNPC | heterogeneous nuclear ribonucleoprotein C |
| Hspa12a | heat shock protein family A member 12A |
| HUVECs | human umbilical vein endothelial cells |
| ICAM1 | intercellular adhesion molecule 1 |
| IGF2BP1 | Insulin-like growth factor 2 mRNA binding protein 1 |
| IOP | intraocular pressure |
| Islr2 | immunoglobulin superfamily containing leucine rich repeat 2 |
| ITGB1 | integrin subunit beta 1 |
| KAT1 | lysine acetyltransferase 1 |
| LCA10 | leber congenital amaurosis 10 |
| lncRNA | long noncoding RNA |
| LRP6 | LDL receptor related protein 6 |
| m1A | N1-methyladenosine |
| m5C | 5-methylcytosine |
| m6A | N6-methyladenosine |
| MAP2 | microtubule associated protein 2 |
| MDA | malondialdehyde |
| MEF | mouse embryonic fibroblast |
| METTL3 | methyltransferase-like 3 |
| miRNA | microRNA |
| mRNA | messenger RNA |
| mtDNA | mitochondrial DNA |
| NEUROD1 | neuronal differentiation 1 |
| NPs | nanoparticles |
| NSCLC | non-small cell lung cancer |
| OIR | oxygen-induced retinopathy |
| OS | oxidative stress |
| PDGFRA | platelet derived growth factor receptor alpha |
| PKC | protein kinase C |
| PRDXs | peroxiredoxins |
| p-STAT3 | phospho-signal transducer and activator of transcription 3 |
| RB | retinoblastoma |
| RBM15 | RNA-binding motif protein 15 |
| RGCs | retinal ganglion cells |
| ROP | retinopathy of prematurity |
| ROS | reactive oxygen species |
| RP | retinitis pigmentosa |
| RPE | retinal pigment epithelium |
| rRNA | ribosomal RNA |
| SAM | S-adenosylmethionine |
| snoRNA | small nucleolar RNA |
| snRNA | small nuclear RNA |
| SODs | superoxide dismutases |
| TM | trabecular meshwork |
| tmRNA | transfer-messenger RNA |
| TON | traumatic optic neuropathy |
| tRNA | transfer RNA |
| TRXPs | thioredoxin peroxidases |
| TRXs | thioredoxins |
| UM | uveal melanoma |
| VEGF | vascular endothelial growth factor |
| VIRMA | vir-like m6A methyltransferase-associated protein |
| WTAP | Wilms tumor 1 associated protein |
| YTHDC1 | YTH domain containing 1 |
| YTHDF1 | YTH N6-methyladenosine RNA binding protein 1 |
| ZC3H13 | zinc finger CCCH-type containing 13 |
| 2′-O-Me | ribosemethylation |
| 3-NP | 3-nitropropionic acid |
| 4-HNE | 4-hydroxy-2-nonenal |
| 8-OHdG | 8-OH 2-deoxyguanosine |
References
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting Free Radicals in Oxidative Stress-Related Human Diseases. Trends Pharmacol. Sci. 2017, 38, 592–607. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.C. Mitochondria and cancer. Nat. Rev. Cancer 2012, 12, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [Green Version]
- Wiener, D.; Schwartz, S. The epitranscriptome beyond m6A. Nat. Rev. Genet. 2020, 22, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Shi, X.; Dai, C.; Shen, F.; Rocco, G.; Chen, J.; Huang, Z.; Chen, C.; He, C.; Huang, T.; et al. RNA m6A Modification in Cancers: Molecular Mechanisms and Potential Clinical Applications. Innovation 2020, 1, 100066. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chai, P.; Jia, R.; Fan, X. Novel insight into the regulatory roles of diverse RNA modifications: Re-defining the bridge between transcription and translation. Mol. Cancer 2020, 19, 78. [Google Scholar] [CrossRef]
- Desrosiers, R.; Friderici, K.; Rottman, F. Identification of Methylated Nucleosides in Messenger RNA from Novikoff Hepatoma Cells. Proc. Natl. Acad. Sci. USA 1974, 71, 3971–3975. [Google Scholar] [CrossRef] [Green Version]
- Jonkhout, N.; Tran, J.; Smith, M.A.; Schonrock, N.; Mattick, J.S.; Novoa, E.M. The RNA modification landscape in human disease. RNA 2017, 23, 1754–1769. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; He, C. Site-specific m6A editing. Nat. Chem. Biol. 2019, 15, 848–849. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Liu, J.; He, C. RNA N6-methyladenosine methylation in post-transcriptional gene expression regulation. Genes Dev. 2015, 29, 1343–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA Modifications in Gene Expression Regulation. Cell 2017, 169, 1187–1200. [Google Scholar] [CrossRef] [Green Version]
- Garbo, S.; Zwergel, C.; Battistelli, C. m6A RNA methylation and beyond—The epigenetic machinery and potential treatment options. Drug Discov. Today 2021, 26, 2559–2574. [Google Scholar] [CrossRef]
- Wang, X.; Feng, J.; Xue, Y.; Guan, Z.; Zhang, D.; Liu, Z.; Gong, Z.; Huang, J.; Tang, C.; Zou, T.; et al. Structural basis of N6-adenosine methylation by the METTL3–METTL14 complex. Nature 2016, 534, 575–578. [Google Scholar] [CrossRef]
- Wang, P.; Doxtader, K.A.; Nam, Y. Structural Basis for Cooperative Function of Mettl3 and Mettl14 Methyltransferases. Mol. Cell 2016, 63, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3–METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2013, 10, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Ping, X.-L.; Sun, B.-F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.-J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.-S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Hao, Y.; Zhang, X.; Xu, S.; Da Pang, D. Targeting RNA N6-methyladenosine modification: A precise weapon in overcoming tumor immune escape. Mol. Cancer 2022, 21, 176. [Google Scholar] [CrossRef]
- Yang, Z.; Yu, G.-L.; Zhu, X.; Peng, T.-H.; Lv, Y.-C. Critical roles of FTO-mediated mRNA m6A demethylation in regulating adipogenesis and lipid metabolism: Implications in lipid metabolic disorders. Genes Dis. 2021, 9, 51–61. [Google Scholar] [CrossRef]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fu, Y.; Klungland, A.; Yang, Y.-G.; He, C. Sprouts of RNA epigenetics: The discovery of mammalian RNA demethylases. RNA Biol. 2013, 10, 915–918. [Google Scholar] [CrossRef] [Green Version]
- Roundtree, I.A.; Luo, G.-Z.; Zhang, Z.; Wang, X.; Zhou, T.; Cui, Y.; Sha, J.; Huang, X.; Guerrero, L.; Xie, P.; et al. YTHDC1 mediates nuclear export of N6-methyladenosine methylated mRNAs. eLife 2017, 6, e31311. [Google Scholar] [CrossRef]
- Xiao, W.; Adhikari, S.; Dahal, U.; Chen, Y.-S.; Hao, Y.-J.; Sun, B.-F.; Sun, H.-Y.; Li, A.; Ping, X.-L.; Lai, W.-Y.; et al. Nuclear m 6 A Reader YTHDC1 Regulates mRNA Splicing. Mol. Cell 2016, 61, 507–519. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.J.; Zhu, Y.; Ma, H.; Guo, Y.; Shi, X.; Liu, Y.; Qi, M.; Lu, Z.; Shi, H.; Wang, J.; et al. Ythdc2 is an N6-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res. 2017, 27, 1115–1127. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. YTHDF2 destabilizes m6A-containing RNA through direct recruitment of the CCR4–NOT deadenylase complex. Nat. Commun. 2016, 7, 12626. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N6-methyladenosine-dependent RNA structural switches regulate RNA–protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef] [Green Version]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.-B.; Jaffrey, S.R. 5′UTR m6A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, Y.-J.; Chen, Y.-N.; Tsao, Y.-T.; Cheng, C.-M.; Wu, W.-C.; Chen, H.-C. The Pathomechanism, Antioxidant Biomarkers, and Treatment of Oxidative Stress-Related Eye Diseases. Int. J. Mol. Sci. 2022, 23, 1255. [Google Scholar] [CrossRef]
- Vallabh, N.A.; Romano, V.; Willoughby, C.E. Mitochondrial dysfunction and oxidative stress in corneal disease. Mitochondrion 2017, 36, 103–113. [Google Scholar] [CrossRef]
- Umapathy, A.; Donaldson, P.; Lim, J. Antioxidant Delivery Pathways in the Anterior Eye. BioMed Res. Int. 2013, 2013, 207250. [Google Scholar] [CrossRef] [Green Version]
- Ung, L.; Pattamatta, U.; Carnt, N.; Wilkinson-Berka, J.L.; Liew, G.; White, A.J. Oxidative stress and reactive oxygen species: A review of their role in ocular disease. Clin. Sci. 2017, 131, 2865–2883. [Google Scholar] [CrossRef]
- Dogru, M.; Kojima, T.; Simsek, C.; Tsubota, K. Potential Role of Oxidative Stress in Ocular Surface Inflammation and Dry Eye Disease. Investig. Ophthalmol. Vis. Sci. 2018, 59, DES163–DES168. [Google Scholar] [CrossRef] [Green Version]
- Uchino, Y.; Kawakita, T.; Miyazawa, M.; Ishii, T.; Onouchi, H.; Yasuda, K.; Ogawa, Y.; Shimmura, S.; Ishii, N.; Tsubota, K. Oxidative stress induced inflammation initiates functional decline of tear production. PLoS ONE 2012, 7, e45805. [Google Scholar] [CrossRef] [Green Version]
- Lakhundi, S.; Siddiqui, R.; Khan, N.A. Pathogenesis of microbial keratitis. Microb. Pathog. 2017, 104, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Lin, Y. Fusarium infection alters the m6A-modified transcript landscape in the cornea. Exp. Eye Res. 2020, 200, 108216. [Google Scholar] [CrossRef]
- Sharif, Z.; Sharif, W. Corneal neovascularization: Updates on pathophysiology, investigations & management. Rom. J. Ophthalmol. 2019, 63, 15–22. [Google Scholar] [CrossRef]
- Shan, K.; Zhou, R.-M.; Xiang, J.; Sun, Y.-N.; Liu, C.; Lv, M.-W.; Xu, J.-J. FTO regulates ocular angiogenesis via m6A-YTHDF2-dependent mechanism. Exp. Eye Res. 2020, 197, 108107. [Google Scholar] [CrossRef]
- Yao, M.-D.; Jiang, Q.; Ma, Y.; Liu, C.; Zhu, C.-Y.; Sun, Y.-N.; Shan, K.; Ge, H.-M.; Zhang, Q.-Y.; Zhang, H.-Y.; et al. Role of METTL3-Dependent N6-Methyladenosine mRNA Modification in the Promotion of Angiogenesis. Mol. Ther. 2020, 28, 2191–2202. [Google Scholar] [CrossRef]
- Dai, Y.; Cheng, M.; Zhang, S.; Ling, R.; Wen, J.; Cheng, Y.; Huang, B.; Li, J.; Dai, C.; Mao, S.; et al. METTL3-Mediated m(6)A RNA Modification Regulates Corneal Injury Repair. Stem Cells Int. 2021, 2021, 1–14. [Google Scholar] [CrossRef]
- Liu, Y.C.; Wilkins, M.; Kim, T.; Malyugin, B.; Mehta, J.S. Cataracts. Lancet 2017, 390, 600–612. [Google Scholar] [CrossRef]
- Fan, X.; Monnier, V.M.; Whitson, J. Lens glutathione homeostasis: Discrepancies and gaps in knowledge standing in the way of novel therapeutic approaches. Exp. Eye Res. 2017, 156, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.; Zhang, X.; Rong, S.; Sha, Q.; Liu, P.; Han, T.; Pan, H. Serum Antioxidative Enzymes Levels and Oxidative Stress Products in Age-Related Cataract Patients. Oxidative Med. Cell Longev. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kaur, J. The oxidative stress in cataract patients. J. Clin. Diagn. Research. 2012, 6, 1629–1632. [Google Scholar] [CrossRef]
- Kandarakis, S.A.; Piperi, C.; Topouzis, F.; Papavassiliou, A.G. Emerging role of advanced glycation-end products (AGEs) in the pathobiology of eye diseases. Prog. Retin. Eye Res. 2014, 42, 85–102. [Google Scholar] [CrossRef]
- Braakhuis, A.J.; I Donaldson, C.; Lim, J.C.; Donaldson, P.J. Nutritional Strategies to Prevent Lens Cataract: Current Status and Future Strategies. Nutrients 2019, 11, 1186. [Google Scholar] [CrossRef] [Green Version]
- Kessel, L.; Lundeman, J.H.; Herbst, K.; Andersen, T.V.; Larsen, M. Age-related changes in the transmission properties of the human lens and their relevance to circadian entrainment. J. Cataract. Refract. Surg. 2010, 36, 308–312. [Google Scholar] [CrossRef]
- Downie, L.E.; Busija, L.; Keller, P.R. Blue-light filtering intraocular lenses (IOLs) for protecting macular health. Cochrane Database Syst. Rev. 2018, 5, CD011977. [Google Scholar] [CrossRef]
- Downie, L.E.; Wormald, R.; Evans, J.; Virgili, G.; Keller, P.R.; Lawrenson, J.G.; Li, T. Analysis of a Systematic Review About Blue Light–Filtering Intraocular Lenses for Retinal Protection. JAMA Ophthalmol. 2019, 137, 694–697. [Google Scholar] [CrossRef]
- Achiron, A.; Elbaz, U.; Hecht, I.; Spierer, O.; Einan-Lifshitz, A.; Karesvuo, P.; Laine, I.; Tuuminen, R. The Effect of Blue-Light Filtering Intraocular Lenses on the Development and Progression of Neovascular Age-Related Macular Degeneration. Ophthalmology 2020, 128, 410–416. [Google Scholar] [CrossRef]
- Lee, J.-S.; Li, P.-R.; Hou, C.-H.; Lin, K.-K.; Kuo, C.-F.; See, L.-C. Effect of Blue Light-Filtering Intraocular Lenses on Age-Related Macular Degeneration: A Nationwide Cohort Study With 10-Year Follow-up. Am. J. Ophthalmol. 2021, 234, 138–146. [Google Scholar] [CrossRef]
- Li, P.; Yu, H.; Zhang, G.; Kang, L.; Qin, B.; Cao, Y.; Luo, J.; Chen, X.; Wang, Y.; Qin, M.; et al. Identification and Characterization of N6-Methyladenosine CircRNAs and Methyltransferases in the Lens Epithelium Cells From Age-Related Cataract. Investig. Opthalmol. Vis. Sci. 2020, 61, 13. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, J.; Zhao, S.; Tian, F. N6-Methyladenosine METTL3 Modulates the Proliferation and Apoptosis of Lens Epithelial Cells in Diabetic Cataract. Mol. Ther. Nucleic Acids 2020, 20, 111–116. [Google Scholar] [CrossRef]
- Gaskin, J.F.; Shah, M.; Chan, E. Oxidative Stress and the Role of NADPH Oxidase in Glaucoma. Antioxidants 2021, 10, 238. [Google Scholar] [CrossRef]
- Tham, Y.-C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.-Y. Global Prevalence of Glaucoma and Projections of Glaucoma Burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef]
- Izzotti, A.; Sacca, S.; Longobardi, M.; Cartiglia, C. Sensitivity of Ocular Anterior Chamber Tissues to Oxidative Damage and Its Relevance to the Pathogenesis of Glaucoma. Investig. Opthalmol. Vis. Sci. 2009, 50, 5251–5258. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Li, Y.; Yue, B.Y. Oxidative stress affects cytoskeletal structure and cell-matrix interactions in cells from an ocular tissue: The trabecular meshwork. J. Cell. Physiol. 1999, 180, 182–189. [Google Scholar] [CrossRef]
- Shen, W.; Wang, C.; Huang, B. Oxidative Stress-Induced circHBEGF Promotes Extracellular Matrix Production via Regulating miR-646/EGFR in Human Trabecular Meshwork Cells. Oxidative Med. Cell. Longev. 2020, 2020, 4692034. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Sahay, P.; Chakraborty, M.; Prusty, B.K.; Srinivasan, S.; Jhingan, G.D.; Mishra, P.; Modak, R.; Suar, M. Switch to Autophagy the Key Mechanism for Trabecular Meshwork Death in Severe Glaucoma. Clin. Ophthalmol. 2021, 15, 3027–3039. [Google Scholar] [CrossRef] [PubMed]
- Awai-Kasaoka, N.; Inoue, T.; Kameda, T.; Fujimoto, T.; Inoue-Mochita, M.; Tanihara, H. Oxidative stress response signaling pathways in trabecular meshwork cells and their effects on cell viability. Mol. Vis. 2013, 19, 1332–1340. [Google Scholar]
- Izzotti, A. Mitochondrial Damage in the Trabecular Meshwork of Patients with Glaucoma. Arch. Ophthalmol. 2010, 128, 724–730. [Google Scholar] [CrossRef] [Green Version]
- Stamer, W.D.; Clark, A.F. The many faces of the trabecular meshwork cell. Exp. Eye Res. 2016, 158, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Ju, W.-K.; Crowston, J.G.; Xie, F.; Perry, G.; Smith, M.A.; Lindsey, J.D.; Weinreb, R.N. Oxidative Stress Is an Early Event in Hydrostatic Pressure–Induced Retinal Ganglion Cell Damage. Investig. Opthalmol. Vis. Sci. 2007, 48, 4580–4589. [Google Scholar] [CrossRef] [Green Version]
- Naguib, S.; Backstrom, J.R.; Gil, M.; Calkins, D.J.; Rex, T.S. Retinal oxidative stress activates the NRF2/ARE pathway: An early endogenous protective response to ocular hypertension. Redox Biol. 2021, 42, 101883. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Huang, Y.-S.; Chen, J.-T.; Tai, M.-C.; Chen, C.-L.; Liang, C.-M. Protective Effects of Glucosamine on Oxidative-Stress and Ischemia/Reperfusion-Induced Retinal Injury. Investig. Opthalmol. Vis. Sci. 2015, 56, 1506–1516. [Google Scholar] [CrossRef]
- Niu, F.; Han, P.; Zhang, J.; She, Y.; Yang, L.; Yu, J.; Zhuang, M.; Tang, K.; Shi, Y.; Yang, B.; et al. The m6A reader YTHDF2 is a negative regulator for dendrite development and maintenance of retinal ganglion cells. eLife 2022, 11, e75827. [Google Scholar] [CrossRef]
- Selmi, C. Diagnosis and classification of autoimmune uveitis. Autoimmun. Rev. 2014, 13, 591–594. [Google Scholar] [CrossRef]
- Kalariya, N.M.; Ramana, K.V. Emerging Role of Antioxidants in the Protection of Uveitis Complications. Curr. Med. Chem. 2011, 18, 931–942. [Google Scholar] [CrossRef]
- Ohkawa, E.; Hiraoka, M.; Abe, A.; Murata, M.; Ohguro, H. Fluctuation of lysosomal phospholipase A2 in experimental autoimmune uveitis in rats. Exp. Eye Res. 2016, 149, 66–74. [Google Scholar] [CrossRef]
- Khurana, R.; Parikh, J.G.; Saraswathy, S.; Rao, N.A.; Wu, G.-S. Mitochondrial Oxidative DNA Damage in Experimental Autoimmune Uveitis. Investig. Opthalmol. Vis. Sci. 2008, 49, 3299–3304. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.-M.; Yang, C.-H.; Teng, Y.-T.; Tsai, H.-Y.; Lin, C.-Y.; Lin, C.-J.; Shieh, C.-C.; Chen, S.-H. Suppression of the Reactive Oxygen Response Alleviates Experimental Autoimmune Uveitis in Mice. Int. J. Mol. Sci. 2020, 21, 3261. [Google Scholar] [CrossRef]
- Karlstetter, M.; Scholz, R.; Rutar, M.; Wong, W.T.; Provis, J.M.; Langmann, T. Retinal microglia: Just bystander or target for therapy? Prog. Retin. Eye Res. 2015, 45, 30–57. [Google Scholar] [CrossRef]
- Zhou, H.; Xu, Z.; Liao, X.; Tang, S.; Li, N.; Hou, S. Low Expression of YTH Domain-Containing 1 Promotes Microglial M1 Polarization by Reducing the Stability of Sirtuin 1 mRNA. Front. Cell Neurosci. 2021, 15, 774305. [Google Scholar] [CrossRef]
- Tang, S.; Meng, J.; Tan, J.; Liu, X.; Zhou, H.; Li, N.; Hou, S. N6-methyladenosine demethylase FTO regulates inflammatory cytokine secretion and tight junctions in retinal pigment epithelium cells. Clin. Immunol. 2022, 241, 109080. [Google Scholar] [CrossRef]
- Tao, J.-X.; Zhou, W.-C.; Zhu, X.-G. Mitochondria as Potential Targets and Initiators of the Blue Light Hazard to the Retina. Oxidative Med. Cell Longev. 2019, 2019, 6435364. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, Y. Oxidative stress in the light-exposed retina and its implication in age-related macular degeneration. Redox Biol. 2020, 37, 101779. [Google Scholar] [CrossRef]
- Nebbioso, M.; Franzone, F.; Lambiase, A.; Bonfiglio, V.; Limoli, P.G.; Artico, M.; Taurone, S.; Vingolo, E.M.; Greco, A.; Polimeni, A. Oxidative Stress Implication in Retinal Diseases—A Review. Antioxidants 2022, 11, 1790. [Google Scholar] [CrossRef]
- Koranda, J.L.; Dore, L.; Shi, H.; Patel, M.; Vaasjo, L.O.; Rao, M.N.; Chen, K.; Lu, Z.; Yi, Y.; Chi, W.; et al. Mettl14 Is Essential for Epitranscriptomic Regulation of Striatal Function and Learning. Neuron 2018, 99, 283–292.e285. [Google Scholar] [CrossRef] [Green Version]
- Bucolo, C.; Leggio, G.M.; Drago, F.; Salomone, S. Dopamine outside the brain: The eye, cardiovascular system and endocrine pancreas. Pharmacol. Ther. 2019, 203, 107392. [Google Scholar] [CrossRef]
- Stitt, A.W.; Curtis, T.M.; Chen, M.; Medina, R.J.; McKay, G.J.; Jenkins, A.; Gardiner, T.A.; Lyons, T.J.; Hammes, H.-P.; Simó, R.; et al. The progress in understanding and treatment of diabetic retinopathy. Prog. Retin. Eye Res. 2016, 51, 156–186. [Google Scholar] [CrossRef]
- Semeraro, F.; Morescalchi, F.; Cancarini, A.; Russo, A.; Rezzola, S.; Costagliola, C. Diabetic retinopathy, a vascular and inflammatory disease: Therapeutic implications. Diabetes Metab. 2019, 45, 517–527. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Chan, P.-S. Oxidative Stress and Diabetic Retinopathy. Exp. Diabetes Res. 2007, 2007, 43603. [Google Scholar] [CrossRef] [Green Version]
- Kang, Q.; Yang, C. Oxidative stress and diabetic retinopathy: Molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biol. 2020, 37, 101799. [Google Scholar] [CrossRef]
- Hombrebueno, J.R.; Cairns, L.; Dutton, L.R.; Lyons, T.J.; Brazil, D.P.; Moynagh, P.; Curtis, T.M.; Xu, H. Uncoupled turnover disrupts mitochondrial quality control in diabetic retinopathy. J. Clin. Investig. 2019, 4, e129760. [Google Scholar] [CrossRef]
- Cheng, Y.; Yu, X.; Zhang, J.; Chang, Y.; Xue, M.; Li, X.; Lu, Y.; Li, T.; Meng, Z.; Su, L.; et al. Pancreatic kallikrein protects against diabetic retinopathy in KK Cg-Ay/J and high-fat diet/streptozotocin-induced mouse models of type 2 diabetes. Diabetologia 2019, 62, 1074–1086. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yu, J.; Ye, M.; Zhao, H. Upregulation of CKIP-1 inhibits high-glucose induced inflammation and oxidative stress in HRECs and attenuates diabetic retinopathy by modulating Nrf2/ARE signaling pathway: An in vitro study. Cell Biosci. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Torres-Cuevas, I.; Millán, I.; Asensi, M.; Vento, M.; Oger, C.; Galano, J.M.; Durand, T.; Ortega, Á.L. Analysis of Lipid Peroxidation by UPLC-MS/MS and Retinoprotective Effects of the Natural Polyphenol Pterostilbene. Antioxidants 2021, 10, 168. [Google Scholar] [CrossRef]
- Catalani, E.; Fanelli, G.; Silvestri, F.; Cherubini, A.; Del Quondam, S.; Bongiorni, S.; Taddei, A.R.; Ceci, M.; De Palma, C.; Perrotta, C.; et al. Nutraceutical Strategy to Counteract Eye Neurodegeneration and Oxidative Stress in Drosophila melanogaster Fed with High-Sugar Diet. Antioxidants 2021, 10, 1197. [Google Scholar] [CrossRef]
- Bucolo, C.; Ward, K.W.; Mazzon, E.; Cuzzocrea, S.; Drago, F. Protective Effects of a Coumarin Derivative in Diabetic Rats. Investig. Opthalmol. Vis. Sci. 2009, 50, 3846–3852. [Google Scholar] [CrossRef] [Green Version]
- Suo, L.; Liu, C.; Zhang, Q.-Y.; Yao, M.-D.; Ma, Y.; Yao, J.; Jiang, Q.; Yan, B. METTL3-mediated N6-methyladenosine modification governs pericyte dysfunction during diabetes-induced retinal vascular complication. Theranostics 2022, 12, 277–289. [Google Scholar] [CrossRef]
- Qi, Y.; Yao, R.; Zhang, W.; Cui, Q. KAT1 triggers YTHDF2-mediated ITGB1 mRNA instability to alleviate the progression of diabetic retinopathy. Pharmacol. Res. 2021, 170, 105713. [Google Scholar] [CrossRef]
- Chen, T.; Zhu, W.; Wang, C.; Dong, X.; Yu, F.; Su, Y.; Huang, J.; Huo, L.; Wan, P. ALKBH5-Mediated m6A Modification of A20 Regulates Microglia Polarization in Diabetic Retinopathy. Front. Immunol. 2022, 13, 813979. [Google Scholar] [CrossRef]
- Zha, X.; Xi, X.; Fan, X.; Ma, M.; Zhang, Y.; Yang, Y. Overexpression of METTL3 attenuates high-glucose induced RPE cell pyroptosis by regulating miR-25-3p/PTEN/Akt signaling cascade through DGCR8. Aging 2020, 12, 8137. [Google Scholar] [CrossRef]
- Gu, C.; Zhang, H.; Li, Q.; Zhao, S.; Gao, Y. MiR-192 attenuates high glucose-induced pyroptosis in retinal pigment epithelial cells via inflammasome modulation. Bioengineered 2022, 13, 10362–10372. [Google Scholar] [CrossRef]
- Huang, C.; Qi, P.; Cui, H.; Lu, Q.; Gao, X. CircFAT1 regulates retinal pigment epithelial cell pyroptosis and autophagy via mediating m6A reader protein YTHDF2 expression in diabetic retinopathy. Exp. Eye Res. 2022, 222, 109152. [Google Scholar] [CrossRef]
- Hellström, A.; Smith, L.E.; Dammann, O. Retinopathy of prematurity. Lancet 2013, 382, 1445–1457. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, D.C.; Brüne, B. Mitochondrial Composition and Function Under the Control of Hypoxia. Redox Biol. 2017, 12, 208–215. [Google Scholar] [CrossRef]
- Stone, W.L. Retinopathy of prematurity an oxidative stress neonatal disease. Front. Biosci. 2016, 21, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Lutty, G.A. Retinal vascular development and oxygen-induced retinopathy: A role for adenosine. Prog. Retin. Eye Res. 2003, 22, 95–111. [Google Scholar] [CrossRef]
- Wang, S.; Ji, L.; Li, L.; Li, J. Oxidative stress, autophagy and pyroptosis in the neovascularization of oxygen-induced retinopathy in mice. Mol. Med. Rep. 2018, 19, 927–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, M.F.; Joo, K.; Kemp, J.A.; Fialho, S.; Cunha, A.D.S.; Woo, S.J.; Kwon, Y.J. Molecular genetics and emerging therapies for retinitis pigmentosa: Basic research and clinical perspectives. Prog. Retin. Eye Res. 2018, 63, 107–131. [Google Scholar] [CrossRef] [PubMed]
- Hamel, C. Retinitis pigmentosa. Orphanet J. Rare Dis. 2006, 1, 40. [Google Scholar] [CrossRef] [PubMed]
- Campochiaro, P.A.; Mir, T.A. The mechanism of cone cell death in Retinitis Pigmentosa. Prog. Retin. Eye Res. 2018, 62, 24–37. [Google Scholar] [CrossRef]
- Yin, L.; Ma, C.; Hou, S.; Ma, X. Methyltransferase-like (METTL)14-mediated N6-methyladenosine modification modulates retinal pigment epithelial (RPE) activity by regulating the methylation of microtubule-associated protein (MAP)2. Bioengineered 2022, 13, 4773–4785. [Google Scholar] [CrossRef]
- Fleckenstein, M.; Keenan, T.D.L.; Guymer, R.H.; Chakravarthy, U.; Schmitz-Valckenberg, S.; Klaver, C.C.; Wong, W.T.; Chew, E.Y. Age-related macular degeneration. Nat. Rev. Dis. Prim. 2021, 7, 31. [Google Scholar] [CrossRef]
- Kaarniranta, K.; Uusitalo, H.; Blasiak, J.; Felszeghy, S.; Kannan, R.; Kauppinen, A.; Salminen, A.; Sinha, D.; Ferrington, D. Mechanisms of mitochondrial dysfunction and their impact on age-related macular degeneration. Prog. Retin. Eye Res. 2020, 79, 100858. [Google Scholar] [CrossRef]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Wang, J.-N.; Cao, Q.-C.; Sun, R.-X.; Zhu, H.-J.; Zhang, Y.-R.; Ji, J.-D.; Liu, Q.-H. m6A modification of circSPECC1 suppresses RPE oxidative damage and maintains retinal homeostasis. Cell Rep. 2022, 41, 111671. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, J.; Wang, Y.; Sun, J.; Huang, P.; Feng, J.; Liu, T.; Sun, X. Fat mass and obesity-associated protein alleviates Aβ 1–40 induced retinal pigment epithelial cells degeneration via PKA/CREB signaling pathway. Cell Biol. Int. 2022, 47, 584–597. [Google Scholar] [CrossRef]
- Steinsapir, K.D.; Goldberg, R.A. Traumatic Optic Neuropathy: An Evolving Understanding. Am. J. Ophthalmol. 2011, 151, 928–933.e922. [Google Scholar] [CrossRef]
- Kang, E.Y.-C.; Liu, P.-K.; Wen, Y.-T.; Quinn, P.M.J.; Levi, S.R.; Wang, N.-K.; Tsai, R.-K. Role of Oxidative Stress in Ocular Diseases Associated with Retinal Ganglion Cells Degeneration. Antioxidants 2021, 10, 1948. [Google Scholar] [CrossRef]
- Qu, X.; Zhu, K.; Li, Z.; Zhang, D.; Hou, L. The Alteration of M6A-Tagged Transcript Profiles in the Retina of Rats After Traumatic Optic Neuropathy. Front. Genet. 2021, 12, 628841. [Google Scholar] [CrossRef]
- Van Poppelen, N.M.; De Bruyn, D.P.; Bicer, T.; Verdijk, R.; Naus, N.; Mensink, H.; Paridaens, D.; De Klein, A.; Brosens, E.; Kiliҫ, E. Genetics of Ocular Melanoma: Insights into Genetics, Inheritance and Testing. Int. J. Mol. Sci. 2020, 22, 336. [Google Scholar] [CrossRef]
- Laver, N.V.; McLaughlin, M.E.; Duker, J.S. Ocular Melanoma. Arch. Pathol. Lab. Med. 2010, 134, 1778–1784. [Google Scholar] [CrossRef]
- Pizzimenti, S.; Ribero, S.; Cucci, M.A.; Grattarola, M.; Monge, C.; Dianzani, C.; Barrera, G.; Muzio, G. Oxidative Stress-Related Mechanisms in Melanoma and in the Acquired Resistance to Targeted Therapies. Antioxidants 2021, 10, 1942. [Google Scholar] [CrossRef]
- Simon, J.D.; Peles, D.; Wakamatsu, K.; Ito, S. Current challenges in understanding melanogenesis: Bridging chemistry, biological control, morphology, and function. Pigment. Cell Melanoma Res. 2009, 22, 563–579. [Google Scholar] [CrossRef]
- Jia, R.; Chai, P.; Wang, S.; Sun, B.; Xu, Y.; Yang, Y.; Ge, S.; Jia, R.; Yang, Y.-G.; Fan, X. m6A modification suppresses ocular melanoma through modulating HINT2 mRNA translation. Mol. Cancer 2019, 18, 161. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.; Yin, J.; Yang, H.; Li, C.; Zhu, L.; Liu, L.; Zhong, J. ALKBH5-mediated m6A demethylation of FOXM1 mRNA promotes progression of uveal melanoma. Aging 2021, 13, 4045–4062. [Google Scholar] [CrossRef]
- Luo, G.; Xu, W.; Zhao, Y.; Jin, S.; Wang, S.; Liu, Q.; Chen, X.; Wang, J.; Dong, F.; Hu, D.; et al. RNA m 6 A methylation regulates uveal melanoma cell proliferation, migration, and invasion by targeting c-Met. J. Cell. Physiol. 2020, 235, 7107–7119. [Google Scholar] [CrossRef]
- He, F.; Yu, J.; Yang, J.; Wang, S.; Zhuang, A.; Shi, H.; Gu, X.; Xu, X.; Chai, P.; Jia, R. m6A RNA hypermethylation-induced BACE2 boosts intracellular calcium release and accelerates tumorigenesis of ocular melanoma. Mol. Ther. 2021, 29, 2121–2133. [Google Scholar] [CrossRef]
- Yu, J.; Chai, P.; Xie, M.; Ge, S.; Ruan, J.; Fan, X.; Jia, R. Histone lactylation drives oncogenesis by facilitating m6A reader protein YTHDF2 expression in ocular melanoma. Genome Biol. 2021, 22, 85. [Google Scholar] [CrossRef]
- Dyson, N.J. RB1: A prototype tumor suppressor and an enigma. Genes Dev. 2016, 30, 1492–1502. [Google Scholar] [CrossRef] [Green Version]
- Ke, F.; Yu, J.; Chen, W.; Si, X.; Li, X.; Yang, F.; Liao, Y.; Zuo, Z. The anti-malarial atovaquone selectively increases chemosensitivity in retinoblastoma via mitochondrial dysfunction-dependent oxidative damage and Akt/AMPK/mTOR inhibition. Biochem. Biophys. Res. Commun. 2018, 504, 374–379. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, W.; Huang, Q.; Wang, J.; Wang, Y.; Li, H.; Fu, X. Tigecycline as a dual inhibitor of retinoblastoma and angiogenesis via inducing mitochondrial dysfunctions and oxidative damage. Sci. Rep. 2018, 8, 11747. [Google Scholar] [CrossRef]
- Li, J.; Min, Y. Pre-clinical evidence that salinomycin is active against retinoblastoma via inducing mitochondrial dysfunction, oxidative damage and AMPK activation. J. Bioenerg. Biomembr. 2021, 53, 513–523. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, P.; Long, C.; Ma, X.; Huang, H.; Kuang, X.; Du, H.; Tang, H.; Ling, X.; Ning, J.; et al. m6A methyltransferase METTL3 promotes retinoblastoma progression via PI3K/AKT/mTOR pathway. J. Cell. Mol. Med. 2020, 24, 12368–12378. [Google Scholar] [CrossRef]
- Trapani, I.; Auricchio, A. Has retinal gene therapy come of age? From bench to bedside and back to bench. Hum. Mol. Genet. 2019, 28, R108–R118. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.S.; Bishop, E.S.; Zhang, R.; Yu, X.; Farina, E.M.; Yan, S.; Zhao, C.; Zeng, Z.; Shu, Y.; Wu, X.; et al. Adenovirus-mediated gene delivery: Potential applications for gene and cell-based therapies in the new era of personalized medicine. Genes Dis. 2017, 4, 43–63. [Google Scholar] [CrossRef]
- Kavanagh, E.W.; Green, J.J. Toward Gene Transfer Nanoparticles as Therapeutics. Adv. Heal. Mater. 2022, 11, e2102145. [Google Scholar] [CrossRef]
- Meza-Rios, A.; Navarro-Partida, J.; Armendariz-Borunda, J.; Santos, A. Therapies Based on Nanoparticles for Eye Drug Delivery. Ophthalmol. Ther. 2020, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B. CRISPR/Cas gene therapy. J. Cell. Physiol. 2020, 236, 2459–2481. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-M.; Zhou, J.; Mao, Y.; Ji, Q.; Qian, S.-B. Programmable RNA N6-methyladenosine editing by CRISPR-Cas9 conjugates. Nat. Chem. Biol. 2019, 15, 865–871. [Google Scholar] [CrossRef]
- Xia, Z.; Tang, M.; Ma, J.; Zhang, H.; Gimple, R.C.; Prager, B.C.; Tang, H.; Sun, C.; Liu, F.; Lin, P.; et al. Epitranscriptomic editing of the RNA N6-methyladenosine modification by dCasRx conjugated methyltransferase and demethylase. Nucleic Acids Res. 2021, 49, 7361–7374. [Google Scholar] [CrossRef]
- Li, J.; Chen, Z.; Chen, F.; Xie, G.; Ling, Y.; Peng, Y.; Lin, Y.; Luo, N.; Chiang, C.-M.; Wang, H. Targeted mRNA demethylation using an engineered dCas13b-ALKBH5 fusion protein. Nucleic Acids Res. 2020, 48, 5684–5694. [Google Scholar] [CrossRef]
- Rauch, S.; He, C.; Dickinson, B.C. Targeted m6A Reader Proteins To Study Epitranscriptomic Regulation of Single RNAs. J. Am. Chem. Soc. 2018, 140, 11974–11981. [Google Scholar] [CrossRef]
- Paul, D.; Sanap, G.; Shenoy, S.; Kalyane, D.; Kalia, K.; Tekade, R.K. Artificial intelligence in drug discovery and development. Drug Discov. Today 2020, 26, 80–93. [Google Scholar] [CrossRef]
- Platania, C.B.M.; Leggio, G.M.; Drago, F.; Salomone, S.; Bucolo, C. Computational systems biology approach to identify novel pharmacological targets for diabetic retinopathy. Biochem. Pharmacol. 2018, 158, 13–26. [Google Scholar] [CrossRef]
- Selberg, S.; Blokhina, D.; Aatonen, M.; Koivisto, P.; Siltanen, A.; Mervaala, E.; Kankuri, E.; Karelson, M. Discovery of Small Molecules that Activate RNA Methylation through Cooperative Binding to the METTL3-14-WTAP Complex Active Site. Cell Rep. 2019, 26, 3762–3771.e3765. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Ye, F.; Yu, L.; Jia, G.; Huang, X.; Zhang, X.; Peng, S.; Chen, K.; Wang, M.; Gong, S.; et al. Development of Cell-Active N6-Methyladenosine RNA Demethylase FTO Inhibitor. J. Am. Chem. Soc. 2012, 134, 17963–17971. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yan, J.; Li, Q.; Li, J.; Gong, S.; Zhou, H.; Gan, J.; Jiang, H.; Jia, G.-F.; Luo, C.; et al. Meclofenamic acid selectively inhibits FTO demethylation of m6A over ALKBH5. Nucleic Acids Res. 2014, 43, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Dong, L.; Li, C.; Nachtergaele, S.; Wunderlich, M.; Qing, Y.; Deng, X.; Wang, Y.; Weng, X.; Hu, C.; et al. R-2HG Exhibits Anti-tumor Activity by Targeting FTO/m6A/MYC/CEBPA Signaling. Cell 2017, 172, 90–105.e123. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Su, R.; Sheng, Y.; Dong, L.; Dong, Z.; Xu, H.; Ni, T.; Zhang, Z.S.; Zhang, T.; Li, C.; et al. Small-Molecule Targeting of Oncogenic FTO Demethylase in Acute Myeloid Leukemia. Cancer Cell 2019, 35, 677–691.e610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Hong, T.; Huang, Y.; Su, H.; Wu, F.; Chen, Y.; Wei, L.; Huang, W.; Hua, X.; Xia, Y.; et al. Fluorescein Derivatives as Bifunctional Molecules for the Simultaneous Inhibiting and Labeling of FTO Protein. J. Am. Chem. Soc. 2015, 137, 13736–13739. [Google Scholar] [CrossRef]
- Su, R.; Dong, L.; Li, Y.; Gao, M.; Han, L.; Wunderlich, M.; Deng, X.; Li, H.; Huang, Y.; Gao, L.; et al. Targeting FTO Suppresses Cancer Stem Cell Maintenance and Immune Evasion. Cancer Cell 2020, 38, 79–96.e11. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liang, G.; Xu, H.; Dong, W.; Dong, Z.; Qiu, Z.; Zhang, Z.; Li, F.; Huang, Y.; Li, Y.; et al. Tumors exploit FTO-mediated regulation of glycolytic metabolism to evade immune surveillance. Cell Metab. 2021, 33, 1221–1233.e1211. [Google Scholar] [CrossRef]
- Malacrida, A.; Rivara, M.; Di Domizio, A.; Cislaghi, G.; Miloso, M.; Zuliani, V.; Nicolini, G. 3D proteome-wide scale screening and activity evaluation of a new ALKBH5 inhibitor in U87 glioblastoma cell line. Bioorganic Med. Chem. 2020, 28, 115300. [Google Scholar] [CrossRef]
- Yankova, E.; Blackaby, W.; Albertella, M.; Rak, J.; De Braekeleer, E.; Tsagkogeorga, G.; Pilka, E.S.; Aspris, D.; Leggate, D.; Hendrick, A.G.; et al. Small-molecule inhibition of METTL3 as a strategy against myeloid leukaemia. Nature 2021, 593, 597–601. [Google Scholar] [CrossRef]
- Moroz-Omori, E.V.; Huang, D.; Bedi, R.K.; Cheriyamkunnel, S.J.; Bochenkova, E.; Dolbois, A.; Rzeczkowski, M.D.; Li, Y.; Wiedmer, L.; Caflisch, A. METTL3 Inhibitors for Epitranscriptomic Modulation of Cellular Processes. Chemmedchem 2021, 16, 3035–3043. [Google Scholar] [CrossRef]
- Cully, M. Chemical inhibitors make their RNA epigenetic mark. Nat. Rev. Drug Discov. 2019, 18, 892–894. [Google Scholar] [CrossRef] [PubMed]

| Targets | Inhibitors | Diseases or Cell Lines | Functions |
|---|---|---|---|
| FTO | Rhein | BE(2)-C cells | Reversibly binds FTO enzyme and competitively prevents the recognition of m6A substrates [141]. |
| Meclofenamic Acid | HeLa cells | Inhibits FTO demethylation of an m6A-containing ssDNA or ssRNA [142]. | |
| R-2HG | human leukemia cell lines and leukemic mouse models | Inhibits leukemia cell proliferation/viability and promotes cell-cycle arrest and apoptosis [143]. | |
| FB23/FB23-2 | human acute myeloid leukemia (AML) cell lines and primary blast AML cells | Suppresses proliferation and promotes the differentiation/apoptosis of human AML cell line cells and primary blast AML cells in vivo and in vitro [144]. | |
| Fluorescein | HeLa cells | Inhibits FTO demethylation inside live cells [145]. | |
| CS1/CS2 | AML cell lines, primary AML cells, and leukemic mouse models | Attenuates leukemia stem/initiating cell self-renewal, reprogram immune response, and immune evasion [146]. | |
| Dac51 | B16-OVA, LLC, and MC38 cells, and tumor-bearing mice | Blocks FTO-mediated immune evasion [147]. | |
| ALKBH5 | MV1035 | U87-MG and H460 cells | Reduces U87 glioblastoma cell line migration and invasiveness [148]. |
| METTL3 | STM2457 | AML cell lines, and leukemic mouse models | Reduces AML growth and increases differentiation and apoptosis [149]. |
| UZH1a/UZH1b | MOLM-13, U2OS, and HEK293T cells | Reduces the m6A/A ratio in mRNAs of AML MOLM-13 cells, osteosarcoma U2OS cells, and the embryonic kidney cell line HEK293T [150]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, Y.; Zhang, H.; Chu, L.; Zhao, Y. m6A Modification—Association with Oxidative Stress and Implications on Eye Diseases. Antioxidants 2023, 12, 510. https://doi.org/10.3390/antiox12020510
Ni Y, Zhang H, Chu L, Zhao Y. m6A Modification—Association with Oxidative Stress and Implications on Eye Diseases. Antioxidants. 2023; 12(2):510. https://doi.org/10.3390/antiox12020510
Chicago/Turabian StyleNi, Yueqi, Hong Zhang, Liang Chu, and Yin Zhao. 2023. "m6A Modification—Association with Oxidative Stress and Implications on Eye Diseases" Antioxidants 12, no. 2: 510. https://doi.org/10.3390/antiox12020510