
Increased Levels of Circulating Iron-Albumin Complexes in Peripheral Arterial Disease Patients
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Design
2.2. Serum Sample Collection
2.3. Gel Filtration and Purification of Iron Complexes
2.4. Iron Quantitation
2.5. Analysis of Protein Content
2.6. Immuno-Isolation of Albumin from Serum Samples
2.7. Statistical Analysis
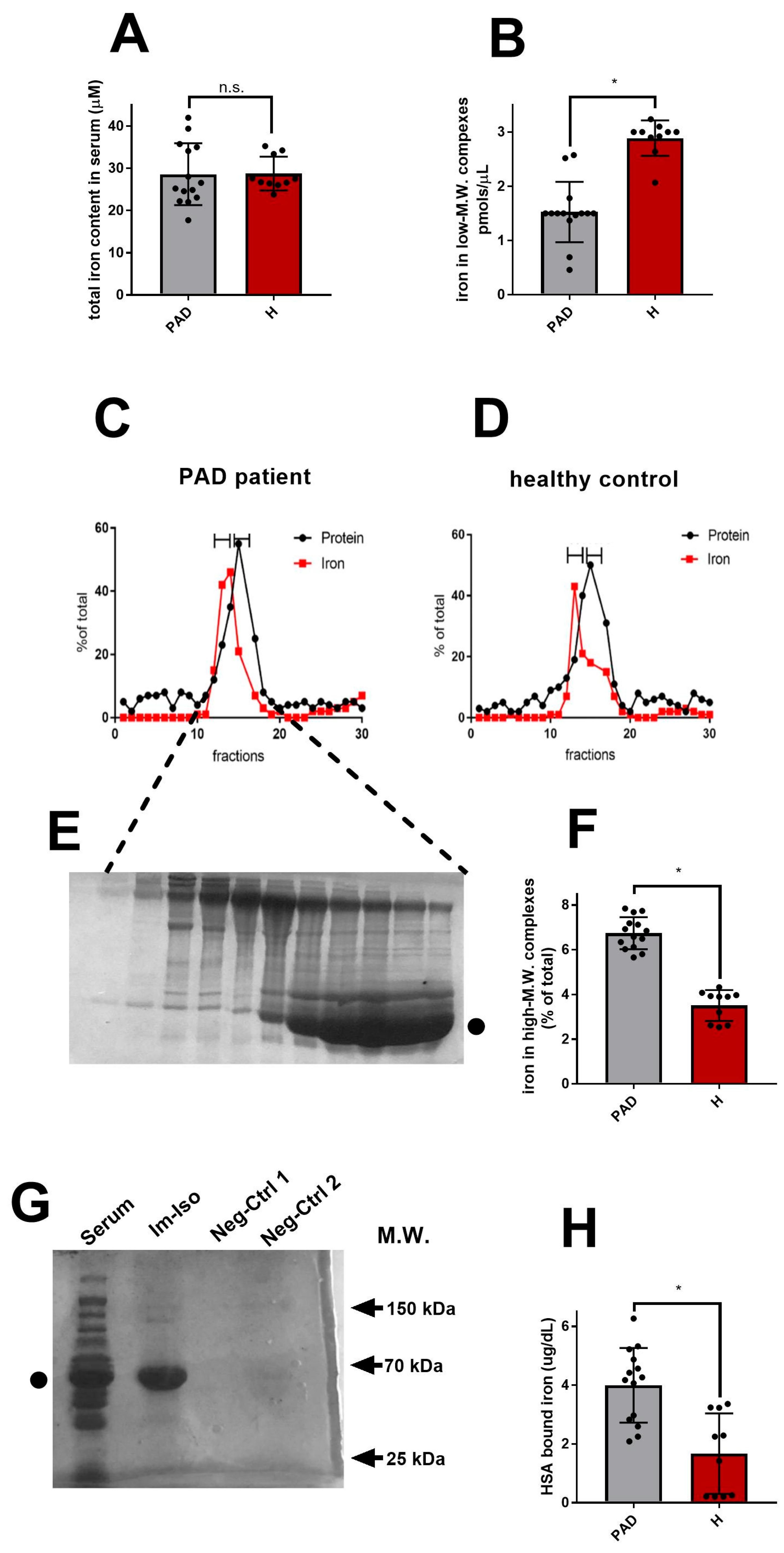
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crichton, R. (Ed.) The Importance of Iron for Biological Systems. In Iron Metabolism: From Molecular Mechanisms to Clinical Consequences, 3rd ed.; Wiley Online Books: Hoboken, NJ, USA, 2009; Volume 1, pp. 17–58. [Google Scholar]
- Sánchez, M.; Sabio, L.; Gálvez, N.; Capdevila, M.; Dominguez-Vera, J.M. Iron chemistry at the service of life. IUBMB Life 2017, 69, 382–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.J. Non-transferrin-bound iron and cellular toxicity: Editorial. J. Gastroenterol. Hepatol. 1999, 14, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Batey, R.G.; Fong, P.L.C.; Shamir, S.; Sherlock, S. A non-transferrin-bound serum iron in idiopathic hemochromatosis. Dig. Dis. Sci. 1980, 25, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.W.; Rafique, R.; Zarea, A.; Rapisarda, C.; Cammack, R.; Evans, P.J.; Porter, J.B.; Hider, R.C. Nature of non-transferrin-bound iron: Studies on iron citrate complexes and thalassemic sera. J. Biol. Inorg. Chem. 2008, 13, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Breuer, W.; Ghoti, H.; Shattat, A.; Goldfarb, A.; Koren, A.; Levin, C.; Rachmilewitz, E.; Cabantchik, Z.I. Non-transferrin bound iron in Thalassemia: Differential detection of redox active forms in children and older patients. Am. J. Hematol. 2012, 87, 55–61. [Google Scholar] [CrossRef]
- Wood, J.C. Guidelines for quantifying iron overload. Hematol. Am. Soc. Hematol. Educ. Program 2014, 2014, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.M.N.; Rangel, M. The (Bio)Chemistry of Non-Transferrin-Bound Iron. Molecules 2022, 27, 1784. [Google Scholar] [CrossRef]
- Patel, M.; Ramavataram, D.V.S.S. Non transferrin bound iron: Nature, manifestations and analytical approaches for estimation. Indian J. Clin. Biochem. 2012, 27, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Izzo, M.; Gasbarro, V.; Coscia, V. The role of free iron in cardiovascular diseases—Part I. J. Theor. Appl. Vasc. Res. 2017, 2, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Lane, D.J.R.; Merlot, A.M.; Huang, M.L.H.; Bae, D.H.; Jansson, P.J.; Sahni, S.; Kalinowski, D.S.; Richardson, D.R. Cellular iron uptake, trafficking and metabolism: Key molecules and mechanisms and their roles in disease. Biochim. Biophys. Acta—Mol. Cell Res. 2015, 1853, 1130–1144. [Google Scholar] [CrossRef] [Green Version]
- Brissot, P.; Ropert, M.; Le Lan, C.; Loréal, O. Non-transferrin bound iron: A key role in iron overload and iron toxicity. Biochim. Biophys. Acta—Gen. Subj. 2012, 1820, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.L. Iron and the genetics of cardiovascular disease. Circulation 1999, 100, 1260–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kell, D.B. Iron behaving badly: Inappropriate iron chelation as a major contributor to the aetiology of vascular and other progressive inflammatory and degenerative diseases. BMC Med. Genomics 2009, 2, 2. [Google Scholar] [CrossRef]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Bishop, G.M.; Dang, T.N.; Dringen, R.; Robinson, S.R. Accumulation of non-transferrin-bound iron by neurons, astrocytes, and microglia. Neurotox. Res. 2011, 19, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Garton, T.; Keep, R.F.; Hua, Y.; Xi, G. Brain iron overload following intracranial haemorrahe. Stroke Vasc. Neurol. 2016, 1, 172–184. [Google Scholar] [CrossRef]
- Radner, S.; Celie, P.H.N.; Fuchs, K.; Sieghart, W.; Sixma, T.K.; Stornaiuolo, M. Transient transfection coupled to baculovirus infection for rapid protein expression screening in insect cells. J. Struct. Biol. 2012, 179, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Oliviero, G.; Stornaiuolo, M.; D’Atri, V.; Nici, F.; Yousif, A.M.; D’Errico, S.; Piccialli, G.; Mayol, L.; Novellino, E.; Marinelli, L.; et al. Screening Platform toward New Anti-HIV Aptamers Set on Molecular Docking and Fluorescence Quenching Techniques. Anal. Chem. 2016, 88, 2327–2334. [Google Scholar] [CrossRef]
- Criqui, M.H.; Aboyans, V. Epidemiology of Peripheral Artery Disease. Circ. Res. 2015, 116, 1509–1526. [Google Scholar] [CrossRef] [Green Version]
- Horváth, L.; Németh, N.; Fehér, G.; Kívés, Z.; Endrei, D.; Boncz, I. Epidemiology of Peripheral Artery Disease: Narrative Review. Life 2022, 12, 1041. [Google Scholar] [CrossRef]
- Nehler, M.R.; Duval, S.; Diao, L.; Annex, B.H.; Hiatt, W.R.; Rogers, K.; Zakharyan, A.; Hirsch, A.T. Epidemiology of peripheral arterial disease and critical limb ischemia in an insured national population. J. Vasc. Surg. 2014, 60, 686–695.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izzo, M.; Gasbarro, V.; Coscia, V. The role of free iron in cardiovascular diseases. Part II. J. Theor. Appl. Vasc. Res. 2017, 2, 83–95. [Google Scholar] [CrossRef]
- Gutteridge, J.M.C. Iron promoters of the Fenton reaction and lipid peroxidation can be released from haemoglobin by peroxides. FEBS Lett. 1986, 201, 291–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, J.L. Iron and the Sex Difference in Heart Disease Risk. Lancet 1981, 317, 1293–1294. [Google Scholar] [CrossRef]
- Ramakrishnan, U.; Kuklina, E.; Stein, A.D. Iron stores and cardiovascular disease risk factors in women of reproductive age in the United States. Am. J. Clin. Nutr. 2002, 76, 1256–1260. [Google Scholar] [CrossRef] [Green Version]
- Danesh, J.; Appleby, P. Coronary heart disease and iron status: Meta-Analyses of Prospective Studies. Circulation. 1999, 99, 852–854. [Google Scholar] [CrossRef] [Green Version]
- Derstine, J.L.; Murray-Kolb, L.E.; Yu-Poth, S.; Hargrove, R.L.; Kris-Etherton, P.M.; Beard, J.L. Iron status in association with cardiovascular disease risk in 3 controlled feeding studies. Am. J. Clin. Nutr. 2003, 77, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Hershko, C.; Graham, G.; Bates, G.W.; Rachmilewitz, E.A. Non-Specific Serum Iron in Thalassaemia: An Abnormal Serum Iron Fraction of Potential Toxicity. Br. J. Haematol. 1978, 40, 255–263. [Google Scholar] [CrossRef]
- Batey, R.G.; Shamir, S.; Wilms, J. Properties and hepatic metabolism of non-transferrin-bound iron. Dig. Dis. Sci. 1981, 26, 1084–1088. [Google Scholar] [CrossRef]
- Steen, D.L.; Cannon, C.P.; Lele, S.S.; Rajapurkar, M.M.; Mukhopadhyay, B.; Scirica, B.M.; Murphy, S.A.; Morrow, D.A. Prognostic evaluation of catalytic iron in patients with acute coronary syndromes. Clin. Cardiol. 2013, 36, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Basuli, D.; Stevens, R.G.; Torti, F.M.; Torti, S.V. Epidemiological associations between iron and cardiovascular disease and diabetes. Front. Pharmacol. 2014, 5, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lele, S.; Shah, S.; McCullough, P.A.; Rajapurkar, M. Serum catalytic iron as a novel biomarker of vascular injury in acute coronary syndromes. EuroIntervention 2009, 5, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Ding, Y.L.; Jacobs, D.R.; Shin, H.R.; Song, K.; Lee, I.K.; Kim, B.; Hider, R.C. Common presence of non-transferrin-bound iron among patients with type 2 diabetes. Diabetes Care 2006, 29, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Qian, M.; Liu, M.; Eaton, J.W. Transition metals bind to glycated proteins forming redox active “Glycochelates”: Implications for the pathogenesis of certain diabetic complications. Biochem. Biophys. Res. Commun. 1998, 250, 385–389. [Google Scholar] [CrossRef]
- Matta, M.K.; Beekman, C.R.; Gandhi, A.; Narayanasamy, S.; Thomas, C.D.; Mohammad, A.; Stewart, S.; Xu, L.; Chockalingam, A.; Shea, K.; et al. Determination of non-transferrin bound iron, transferrin bound iron, drug bound iron and total iron in serum in a rats after IV administration of sodium ferric gluconate complex by simple ultrafiltration inductively coupled plasma mass spectrometric detec. Nanomaterials 2018, 8, 101. [Google Scholar] [CrossRef] [Green Version]
- Belinskaia, D.A.; Voronina, P.A.; Shmurak, V.I.; Vovk, M.A.; Batalova, A.A.; Jenkins, R.O.; Goncharov, N.V. The universal soldier: Enzymatic and non-enzymatic antioxidant functions of serum albumin. Antioxidants 2020, 9, 966. [Google Scholar] [CrossRef]
- Tabata, F.; Wada, Y.; Kawakami, S.; Miyaji, K. Serum albumin redox states: More than oxidative stress biomarker. Antioxidants 2021, 10, 504. [Google Scholar] [CrossRef]
- Oettl, K.; Marsche, G. Redox State of Human Serum Albumin in Terms of Cysteine-34 in Health and Disease, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2010; Volume 474, ISBN 9780123810038. [Google Scholar]
- Belinskaia, D.A.; Voronina, P.A.; Shmurak, V.I.; Jenkins, R.O.; Goncharov, N.V. Serum albumin in health and disease: Esterase, antioxidant, transporting and signaling properties. Int. J. Mol. Sci. 2021, 22, 10318. [Google Scholar] [CrossRef]
- Sozarukova, M.M.; Proskurnina, E.V.; Vladimirov, Y.A. Serum albumin as a source of and a target for free radicals in pathology. Bull. Russ. State Med. Univ. 2016, 1, 56–61. [Google Scholar] [CrossRef]
- Bhakthavatsala Reddy, C.; Cyriac, C.; Desle, H.B. Role of “ischemia Modified Albumin” (IMA) in acute coronary syndromes. Indian Heart J. 2014, 66, 656–662. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.H.; Morris, D.L.; Fletcher, D.R.; Apple, F.S.; Christenson, R.H.; Painter, P.C. Analysis of the Albumin Cobalt Binding “ACB” test as an adjunct to cardiac troponin I for the early detection of acute myocardial infarction. Cardiovasc. Toxicol. 2001, 1, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Sitar, M.E.; Aydin, S.; Çakatay, U. Human serum albumin and its relation with oxidative stress. Clin. Lab. 2013, 59, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Isik, B.; Ceylan, A.; Isik, R. Oxidative stress in smokers and non-smokers. Inhal. Toxicol. 2007, 19, 767–769. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Demographics | PAD (n = 14) | H (n = 10) | Normal Value Range |
|---|---|---|---|
| Age (years) | 73.2 (47–92) | 60.1 (25–69) | |
| Male sex (No. (%)) | 10 (71%) | 8 (80%) | |
| White ethnicity (No. (%)) | 14 (100%) | 10 (100%) | |
| Clinical Parameters | |||
| Diabetes | 7 (50%) * | 0 (0%) | |
| Hypertension | 13 (93%) * | 0 (0%) | |
| Dyslipidemia | 11 (79%) * | 3 (30%) | |
| Cigarette Smoking | 1 (7%) * | 5 (50%) | |
| Obstructive arteriopathy (No. (%)) | 11 (79%) * | 0 (0%) | |
| CVI (No. (%)) | 3 (21%) * | 0 (0%) | |
| Serum Iron (µg/dL) | 154 (115–176) | 121 (91–158) | (60–170) |
| Serum Transferrin (g/L) | 2.9 (2.6–3.2) | 2.8 (2.6–3.1) | (2.5–3.8) |
| Serum Ferritin (ng/mL) | 75.2 (22–134) | 91.3 (38–156) | (22–322) |
| Total Iron Binding Capacity (µg/dL) | 296.2 (273–350) | 302.4 (260–380) | (240–450) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiano, E.; Cappello, E.; Cecere, D.; Pompeo, F.; Novellino, E.; Stornaiuolo, M.; Izzo, M. Increased Levels of Circulating Iron-Albumin Complexes in Peripheral Arterial Disease Patients. Antioxidants 2023, 12, 503. https://doi.org/10.3390/antiox12020503
Schiano E, Cappello E, Cecere D, Pompeo F, Novellino E, Stornaiuolo M, Izzo M. Increased Levels of Circulating Iron-Albumin Complexes in Peripheral Arterial Disease Patients. Antioxidants. 2023; 12(2):503. https://doi.org/10.3390/antiox12020503
Chicago/Turabian StyleSchiano, Elisabetta, Enrico Cappello, Domenico Cecere, Francesco Pompeo, Ettore Novellino, Mariano Stornaiuolo, and Marcello Izzo. 2023. "Increased Levels of Circulating Iron-Albumin Complexes in Peripheral Arterial Disease Patients" Antioxidants 12, no. 2: 503. https://doi.org/10.3390/antiox12020503