
Recent Advances in Cellular Signaling Interplay between Redox Metabolism and Autophagy Modulation in Cancer: An Overview of Molecular Mechanisms and Therapeutic Interventions

 , ,
, ,  and
and
Abstract
:1. Introduction
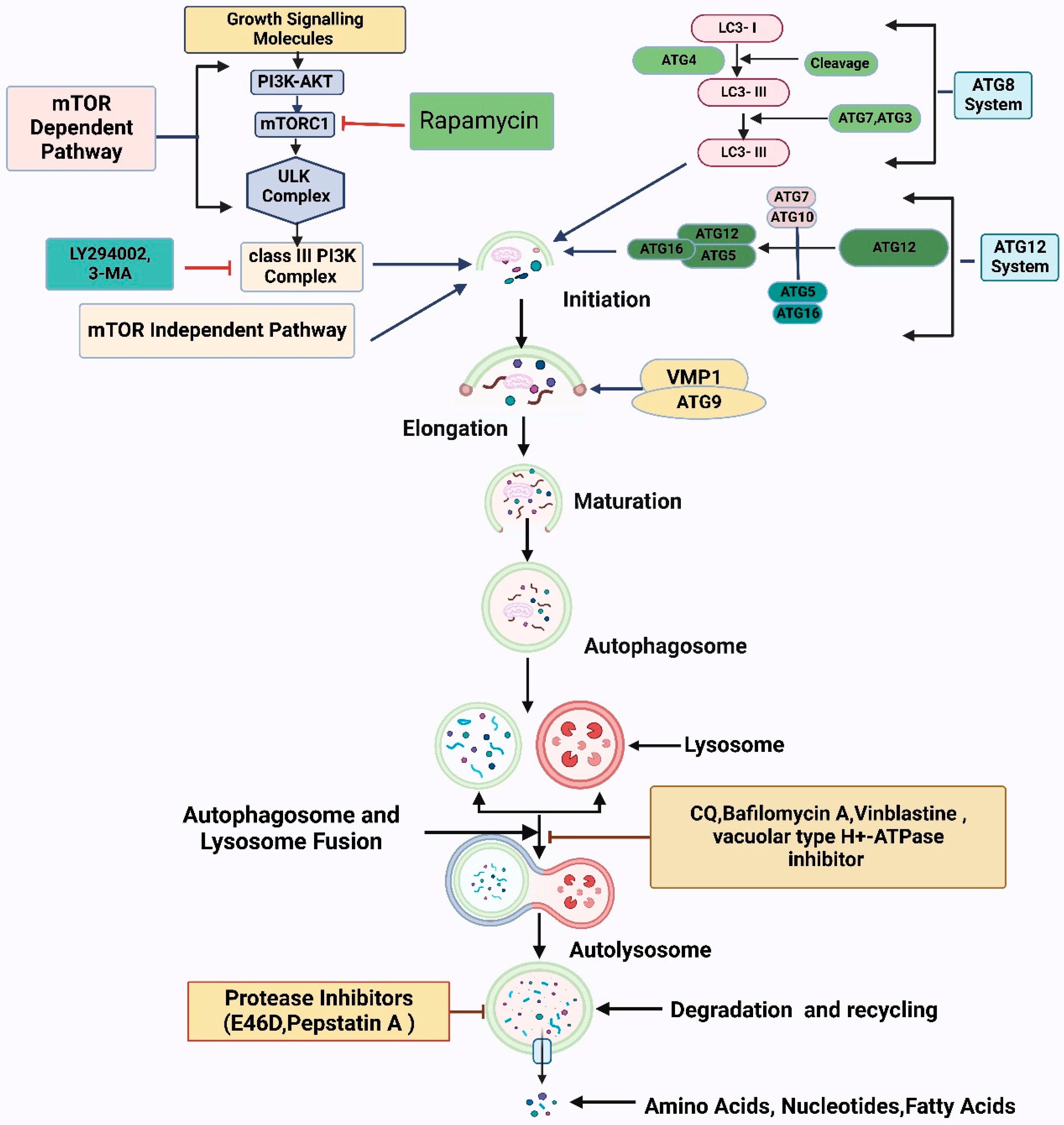
2. Cellular Signaling and Physiological Roles of Redox Metabolism and Autophagy
3. Interplay between Redox Signaling and Autophagy in Cancer
4. Autophagy Controls ROS Production and Antioxidants to Regulate Cellular Redox Homeostasis
5. Recent Update and Therapeutic Application of Redox Homeostasis and Autophagy in Cancer
5.1. Synthetic Drugs Targeting Redox Homeostasis and Autophagy in Cancer
5.2. Natural-Compound-Mediated Targeting of Redox Metabolism and Autophagy in Cancer
5.3. Nanoparticle-Mediated Targeting of Redox Metabolism and Autophagy in Cancer
5.4. MicroRNA-Mediated Targeting of Redox Metabolism and Autophagy in Cancer
6. Limitations and Future Perspectives on Redox Metabolism and Autophagy in Cancer
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rangel, M.; Kong, J.; Bhatt, V.; Khayati, K.; Guo, J.Y. Autophagy and tumorigenesis. FEBS J. 2022, 289, 7177–7198. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, L.D.; Narita, M. Autophagy at the intersection of aging, senescence, and cancer. Mol. Oncol. 2022, 16, 3259–3275. [Google Scholar] [CrossRef]
- Gubas, A.; Dikic, I. A guide to the regulation of selective autophagy receptors. FEBS J. 2022, 289, 75–89. [Google Scholar] [CrossRef]
- Rahman, M.A.; Bishayee, K.; Sadra, A.; Huh, S.O. Oxyresveratrol activates parallel apoptotic and autophagic cell death pathways in neuroblastoma cells. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Rhim, H. Therapeutic implication of autophagy in neurodegenerative diseases. BMB Rep. 2017, 50, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Ahmed, K.R.; Rahman, M.H.; Park, M.N.; Kim, B. Potential therapeutic action of autophagy in gastric cancer managements: Novel treatment strategies and pharmacological interventions. Front. Pharmacol. 2021, 12, 3894. [Google Scholar]
- Reyes-Castellanos, G.; Abdel Hadi, N.; Carrier, A. Autophagy contributes to metabolic reprogramming and therapeutic resistance in pancreatic tumors. Cells 2022, 11, 426. [Google Scholar] [CrossRef]
- Qin, S.; Li, B.; Ming, H.; Nice, E.C.; Zou, B.; Huang, C. Harnessing redox signaling to overcome therapeutic-resistant cancer dormancy. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2022, 1877, 188749. [Google Scholar] [CrossRef]
- Castaneda, M.; den Hollander, P.; Kuburich, N.A.; Rosen, J.M.; Mani, S.A. Mechanisms of Cancer Metastasis. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Worsley, C.M.; Veale, R.B.; Mayne, E.S. The acidic tumour microenvironment: Manipulating the immune response to elicit escape. Hum. Immunol. 2022, 83, 399–408. [Google Scholar]
- Taucher, E.; Mykoliuk, I.; Fediuk, M.; Smolle-Juettner, F.-M. Autophagy, Oxidative Stress and Cancer Development. Cancers 2022, 14, 1637. [Google Scholar]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef]
- Clarke, T.L.; Mostoslavsky, R. DNA repair as a shared hallmark in cancer and ageing. Mol. Oncol. 2022, 16, 3352–3379. [Google Scholar] [PubMed]
- Zhang, Y.; Guo, R.; Wang, S.-S.; Jiang, X.-Y.; Cui, H.-Y.; Guo, Y.; Song, X.-Y.; Guo, Q.-Q.; Cao, L. Autophagy-related Proteins in Genome Stability: Autophagy-Dependent and Independent Actions. Int. J. Biol. Sci. 2022, 18, 5329–5344. [Google Scholar] [PubMed]
- Bayır, H.; Maguire, J.J.; Cadenas, E. Redox Pioneer: Professor Valerian Kagan. Antioxid. Redox Signal. 2022, 36, 813–823. [Google Scholar]
- Rahman, M.A.; Ahmed, K.R.; Rahman, M.H.; Parvez, M.A.K.; Lee, I.-S.; Kim, B. Therapeutic Aspects and Molecular Targets of Autophagy to Control Pancreatic Cancer Management. Biomedicines 2022, 10, 1459. [Google Scholar]
- Rahman, M.A.; Rahman, M.H.; Mamun-Or-Rashid, A.; Hwang, H.; Chung, S.; Kim, B.; Rhim, H. Autophagy Modulation in Aggresome Formation: Emerging Implications and Treatments of Alzheimer’s Disease. Biomedicines 2022, 10, 1027. [Google Scholar] [PubMed]
- Uddin, M.S.; Rahman, M.A.; Kabir, M.T.; Behl, T.; Mathew, B.; Perveen, A.; Barreto, G.E.; Bin-Jumah, M.N.; Abdel-Daim, M.M.; Ashraf, G.M. Multifarious roles of mTOR signaling in cognitive aging and cerebrovascular dysfunction of Alzheimer’s disease. IUBMB Life 2020, 72, 1843–1855. [Google Scholar] [CrossRef]
- Rahman, M.A.; Cho, Y.; Nam, G.; Rhim, H. Antioxidant compound, oxyresveratrol, inhibits APP production through the AMPK/ULK1/mTOR-mediated autophagy pathway in mouse cortical astrocytes. Antioxidants 2021, 10, 408. [Google Scholar] [CrossRef]
- Mishra, J.; Bhatti, G.K.; Sehrawat, A.; Singh, C.; Singh, A.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Modulating autophagy and mitophagy as a promising therapeutic approach in neurodegenerative disorders. Life Sci. 2022, 311, 121153. [Google Scholar] [CrossRef]
- Hu, Y.; Reggiori, F. Molecular regulation of autophagosome formation. Biochem. Soc. Trans. 2022, 50, 55–69. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Ding, J.-L.; Peng, Y.-J.; Feng, M.-G.; Ying, S.-H. Proteomic and Phosphoryproteomic Investigations Reveal that Autophagy-Related Protein 1, a Protein Kinase for Autophagy Initiation, Synchronously Deploys Phosphoregulation on the Ubiquitin-Like Conjugation System in the Mycopathogen Beauveria bassiana. Msystems 2022, 7, e01463-21. [Google Scholar] [CrossRef] [PubMed]
- Vitto, V.A.M.; Bianchin, S.; Zolondick, A.A.; Pellielo, G.; Rimessi, A.; Chianese, D.; Yang, H.; Carbone, M.; Pinton, P.; Giorgi, C. Molecular Mechanisms of Autophagy in Cancer Development, Progression, and Therapy. Biomedicines 2022, 10, 1596. [Google Scholar] [CrossRef] [PubMed]
- González-Rodríguez, P.; Delorme-Axford, E.; Bernard, A.; Keane, L.; Stratoulias, V.; Grabert, K.; Engskog-Vlachos, P.; Füllgrabe, J.; Klionsky, D.J.; Joseph, B. SETD2 transcriptional control of ATG14L/S isoforms regulates autophagosome–lysosome fusion. Cell Death Dis. 2022, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Chakrabarti, P.; Sen, S.; Barui, A. Reactive oxygen species in modulating intestinal stem cell dynamics and function. Stem Cell Rev. Rep. 2022, 18, 2328–2350. [Google Scholar] [PubMed]
- Olajide, P.A.; Omowumi, O.S.; Odine, G.O. Pathogenesis of Reactive Oxygen Species: A Review. World News Nat. Sci. 2022, 44, 150–164. [Google Scholar]
- Hemagirri, M.; Sasidharan, S. Biology of aging: Oxidative stress and RNA oxidation. Mol. Biol. Rep. 2022, 49, 5089–5105. [Google Scholar]
- Chakraborty, S.; Sircar, E.; Bhattacharyya, C.; Choudhuri, A.; Mishra, A.; Dutta, S.; Bhatta, S.; Sachin, K.; Sengupta, R. S-Denitrosylation: A Crosstalk between Glutathione and Redoxin Systems. Antioxidants 2022, 11, 1921. [Google Scholar]
- Anik, M.I.; Mahmud, N.; Masud, A.A.; Khan, M.I.; Islam, M.N.; Uddin, S.; Hossain, M.K. Role of Reactive Oxygen Species in Aging and Age-Related Diseases: A Review. ACS Appl. Biol. Mater. 2022, 5, 4028–4054. [Google Scholar]
- Shilovsky, G.A. Lability of the Nrf2/Keap/ARE Cell Defense System in Different Models of Cell Aging and Age-Related Pathologies. Biochemistry 2022, 87, 70–85. [Google Scholar]
- Tang, Z.; Hu, B.; Zang, F.; Wang, J.; Zhang, X.; Chen, H. Nrf2 drives oxidative stress-induced autophagy in nucleus pulposus cells via a Keap1/Nrf2/p62 feedback loop to protect intervertebral disc from degeneration. Cell Death Dis. 2019, 10, 510. [Google Scholar]
- Shah, S.Z.A.; Zhao, D.; Hussain, T.; Sabir, N.; Mangi, M.H.; Yang, L. p62-Keap1-NRF2-ARE pathway: A contentious player for selective targeting of autophagy, oxidative stress and mitochondrial dysfunction in prion diseases. Front. Mol. Neurosci. 2018, 11, 310. [Google Scholar] [PubMed] [Green Version]
- Zuo, J.; Zhang, Z.; Luo, M.; Zhou, L.; Nice, E.C.; Zhang, W.; Wang, C.; Huang, C. Redox signaling at the crossroads of human health and disease. MedComm 2022, 3, e127. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Gao, M.; Liu, B.; Qin, Y.; Liu, H.; Wu, H.; Gong, G. Mitochondrial autophagy: Molecular mechanisms and implications for cardiovascular disease. Cell Death Dis. 2022, 13, 1–15. [Google Scholar]
- Bhattacharjee, S.; Paul, S.; RayBarman, C. Redox Signaling: Hallmarks of Cancer Progression and Resistance to Treatment. In Handbook of Oxidative Stress in Cancer: Therapeutic Aspects; Springer: Singapore, 2022; pp. 1–21. [Google Scholar]
- Su, L.; Zhang, J.; Gomez, H.; Kellum, J.A.; Peng, Z. Mitochondria ROS and mitophagy in acute kidney injury. Autophagy 2022, 19, 401–414. [Google Scholar] [CrossRef]
- Han, R.; Liu, Y.; Li, S.; Li, X.-J.; Yang, W. PINK1-PRKN mediated mitophagy: Differences between in vitro and in vivo models. Autophagy 2022. [Google Scholar] [CrossRef]
- Jiang, Y.; Krantz, S.; Qin, X.; Li, S.; Gunasekara, H.; Kim, Y.-M.; Zimnicka, A.; Bae, M.; Ma, K.; Toth, P.T. Caveolin-1 controls mitochondrial damage and ROS production by regulating fission-fusion dynamics and mitophagy. Redox Biol. 2022, 52, 102304. [Google Scholar] [CrossRef] [PubMed]
- Alan, P.; Vandevoorde, K.R.; Joshi, B.; Cardoen, B.; Gao, G.; Mohammadzadeh, Y.; Hamarneh, G.; Nabi, I.R. Basal Gp78-dependent mitophagy promotes mitochondrial health and limits mitochondrial ROS. Cell. Mol. Life Sci. 2022, 79, 1–20. [Google Scholar] [CrossRef]
- Khan, S.U.; Fatima, K.; Aisha, S.; Hamza, B.; Malik, F. Redox balance and autophagy regulation in cancer progression and their therapeutic perspective. Med. Oncol. 2023, 40, 1–21. [Google Scholar] [CrossRef]
- Kumar, P.; Jagtap, Y.A.; Patwa, S.M.; Kinger, S.; Dubey, A.R.; Prajapati, V.K.; Dhiman, R.; Poluri, K.M.; Mishra, A. Autophagy based cellular physiological strategies target oncogenic progression. J. Cell. Physiol. 2022, 237, 258–277. [Google Scholar]
- Xi, H.; Wang, S.; Wang, B.; Hong, X.; Liu, X.; Li, M.; Shen, R.; Dong, Q. The role of interaction between autophagy and apoptosis in tumorigenesis. Oncol. Rep. 2022, 48, 1–16. [Google Scholar]
- Gao, W.; Wang, X.; Zhou, Y.; Wang, X.; Yu, Y. Autophagy, ferroptosis, pyroptosis, and necroptosis in tumor immunotherapy. Signal Transduct. Target. Ther. 2022, 7, 1–26. [Google Scholar]
- George, S.; Abrahamse, H. ROS, Cancer, Stem Cells. In Handbook of Oxidative Stress in Cancer: Mechanistic Aspects; Springer: Singapore, 2022; pp. 2147–2163. [Google Scholar]
- Sahoo, B.M.; Banik, B.K.; Borah, P.; Jain, A. Reactive oxygen species (ROS): Key components in cancer therapies. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem. Anti-Cancer Agents) 2022, 22, 215–222. [Google Scholar]
- Luo, Z.; Tian, M.; Yang, G.; Tan, Q.; Chen, Y.; Li, G.; Zhang, Q.; Li, Y.; Wan, P.; Wu, J. Hypoxia signaling in human health and diseases: Implications and prospects for therapeutics. Signal Transduct. Target. Ther. 2022, 7, 1–30. [Google Scholar]
- Hasan, A.; Rizvi, S.F.; Parveen, S.; Pathak, N.; Nazir, A.; Mir, S.S. Crosstalk between ROS and autophagy in tumorigenesis: Understanding the multifaceted paradox. Front. Oncol. 2022, 12, 852424. [Google Scholar] [PubMed]
- Hussain, I. Role of the mTOR/AMPK Pathways in Metabolic and Inflammatory Hepatic Dysfunction. Ph.D. Thesis, Friedrich Schiller University, Jena, Germany, 2022. [Google Scholar]
- Gray, N.E.; Farina, M.; Tucci, P.; Saso, L. The Role of the NRF2 Pathway in Maintaining and Improving Cognitive Function. Biomedicines 2022, 10, 2043. [Google Scholar] [CrossRef]
- Gureev, A.P.; Shaforostova, E.A.; Popov, V.N. Regulation of mitochondrial biogenesis as a way for active longevity: Interaction between the Nrf2 and PGC-1α signaling pathways. Front. Genet. 2019, 10, 435. [Google Scholar] [PubMed]
- Liu, S.; Pi, J.; Zhang, Q. Signal amplification in the KEAP1-NRF2-ARE antioxidant response pathway. Redox Biol. 2022, 54, 102389. [Google Scholar] [CrossRef]
- Sun, C.; Han, B.; Zhai, Y.; Zhao, H.; Li, X.; Qian, J.; Hao, X.; Liu, Q.; Shen, J.; Kai, G. Dihydrotanshinone I inhibits ovarian tumor growth by activating oxidative stress through Keap1-mediated Nrf2 ubiquitination degradation. Free Radic. Biol. Med. 2022, 180, 220–235. [Google Scholar]
- Pang, X.; Zhang, X.; Jiang, Y.; Su, Q.; Li, Q.; Li, Z. Autophagy: Mechanisms and therapeutic potential of flavonoids in cancer. Biomolecules 2021, 11, 135. [Google Scholar] [CrossRef]
- Ulasov, A.V.; Rosenkranz, A.A.; Georgiev, G.P.; Sobolev, A.S. Nrf2/Keap1/ARE signaling: Towards specific regulation. Life Sci. 2022, 291, 120111. [Google Scholar] [CrossRef]
- Tossetta, G.; Marzioni, D. Targeting the NRF2/KEAP1 pathway in cervical and endometrial cancers. Eur. J. Pharm. 2023, 941, 175503. [Google Scholar]
- Babu, S.; Manoharan, S.; Ottappilakkil, H.; Perumal, E. Role of oxidative stress-mediated cell death and signaling pathways in experimental fluorosis. Chem.-Biol. Interact. 2022, 365, 110106. [Google Scholar] [PubMed]
- Loftus, L.V.; Amend, S.R.; Pienta, K.J. Interplay between Cell Death and Cell Proliferation Reveals New Strategies for Cancer Therapy. Int. J. Mol. Sci. 2022, 23, 4723. [Google Scholar] [PubMed]
- Yang, Q.; Fang, Y.; Zhang, C.; Liu, X.; Wu, Y.; Zhang, Y.; Yang, J.; Yong, K. Exposure to zinc induces lysosomal-mitochondrial axis-mediated apoptosis in PK-15 cells. Ecotoxicol. Environ. Saf. 2022, 241, 113716. [Google Scholar]
- Zhou, J.; Li, X.-Y.; Liu, Y.-J.; Feng, J.; Wu, Y.; Shen, H.-M.; Lu, G.-D. Full-coverage regulations of autophagy by ROS: From induction to maturation. Autophagy 2022, 18, 1240–1255. [Google Scholar]
- Dey, S.; Murmu, N.; Mondal, T.; Saha, I.; Chatterjee, S.; Manna, R.; Haldar, S.; Dash, S.K.; Sarkar, T.R.; Giri, B. Multifaceted entrancing role of glucose and its analogue, 2-deoxy-D-glucose in cancer cell proliferation, inflammation, and virus infection. Biomed. Pharmacother. 2022, 156, 113801. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Zhang, Z.; Li, M.; Yang, Y.; Zheng, B.; Wang, P.; Huang, C.; Zhou, S. The crosstalk between reactive oxygen species and noncoding RNAs: From cancer code to drug role. Mol. Cancer 2022, 21, 1–17. [Google Scholar] [CrossRef]
- Kim, M.-J.; Jeon, J.-H. Recent advances in understanding Nrf2 agonism and its potential clinical application to metabolic and inflammatory diseases. Int. J. Mol. Sci. 2022, 23, 2846. [Google Scholar]
- Kryszczuk, M.; Kowalczuk, O. Significance of NRF2 in physiological and pathological conditions an comprehensive review. Arch. Biochem. Biophys. 2022, 730, 109417. [Google Scholar]
- Navarro-Yepes, J.; Burns, M.; Anandhan, A.; Khalimonchuk, O.; Del Razo, L.M.; Quintanilla-Vega, B.; Pappa, A.; Panayiotidis, M.I.; Franco, R. Oxidative stress, redox signaling, and autophagy: Cell death versus survival. Antioxid. Redox Signal. 2014, 21, 66–85. [Google Scholar]
- He, J.; Dong, L.; Luo, L.; Wang, K. Redox Regulation of Autophagy in Cancer: Mechanism, Prevention and Therapy. Life 2022, 13, 98. [Google Scholar] [PubMed]
- Montégut, L.; de Cabo, R.; Zitvogel, L.; Kroemer, G. Science-driven nutritional interventions for the prevention and treatment of cancer. Cancer Discov. 2022, 12, 2258–2279. [Google Scholar] [PubMed]
- Shabani Dargah, M.; Hadjizadeh, A. Improvement of ascorbic acid delivery into human skin via hyaluronic acid-coated niosomes. J. Microencapsul. 2022, 39, 552–562. [Google Scholar] [CrossRef]
- Bieńkowska-Tokarczyk, A.; Manda-Handzlik, A.; Gawrychowski, K.; Demkow, U.; Małecki, M. The Activity of Vitamin C Against Ovarian Cancer Cells Is Enhanced by Hyperthermia. Anticancer Res. 2022, 42, 5365–5383. [Google Scholar] [CrossRef]
- Lee, C.T.; Galloway, T.J. Pathogenesis and amelioration of radiation-induced oral mucositis. Curr. Treat. Options Oncol. 2022, 23, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Park, M.; Kim, N.; Lee, Y.; Jung, E.; Lee, D. Tumor-targeting oxidative stress nanoamplifiers as anticancer nanomedicine with immunostimulating activity. Biomater. Sci. 2022, 10, 6160–6171. [Google Scholar]
- Shen, Y.; Zhang, H.; Yao, S.; Su, F.; Wang, H.; Yin, J.; Fang, Y.; Tan, L.; Zhang, K.; Fan, X. Methionine oxidation of CLK4 promotes the metabolic switch and redox homeostasis in esophageal carcinoma via inhibiting MITF selective autophagy. Clin. Transl. Med. 2022, 12, e719. [Google Scholar] [CrossRef]
- Kim, J.; Park, A.; Hwang, J.; Zhao, X.; Kwak, J.; Kim, H.W.; Ku, M.; Yang, J.; Kim, T.I.; Jeong, K.-S. KS10076, a chelator for redox-active metal ions, induces ROS-mediated STAT3 degradation in autophagic cell death and eliminates ALDH1+ stem cells. Cell Rep. 2022, 40, 111077. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, D.K.; Nayel, O.A.; Abdulmalek, S.; Abdelbary, A.A.; Ismail, C.A. Modulation of autophagy, apoptosis and oxidative stress: A clue for repurposing metformin in photoaging. Inflammopharmacology 2022, 30, 2521–2535. [Google Scholar]
- Parisotto, M.; Vuong-Robillard, N.; Kalegari, P.; Meharwade, T.; Joumier, L.; Igelmann, S.; Bourdeau, V.; Rowell, M.-C.; Pollak, M.; Malleshaiah, M. The NAMPT Inhibitor FK866 Increases Metformin Sensitivity in Pancreatic Cancer Cells. Cancers 2022, 14, 5597. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, W.; Lu, S.-Y.; Cai, X.; Chen, C.; Zhang, Q.; Duan, Y.; Xie, D.; Zhang, Q.; Ran, H. Biodegradable doxorubicin-loaded ferric phosphate nanosheets for specific tumor elimination through autophagy inhibition-enhanced apoptosis/ferroptosis pathway. Chem. Eng. J. 2022, 454, 140455. [Google Scholar] [CrossRef]
- Jung, M.; Lee, K.M.; Im, Y.; Seok, S.H.; Chung, H.; Kim, D.Y.; Han, D.; Lee, C.H.; Hwang, E.H.; Park, S.Y. Nicotinamide (niacin) supplement increases lipid metabolism and ROS-induced energy disruption in triple-negative breast cancer: Potential for drug repositioning as an anti-tumor agent. Mol. Oncol. 2022, 16, 1795–1815. [Google Scholar] [CrossRef] [PubMed]
- Lae Lae Phoo, N.; Sukhamwang, A.; Dejkriengkraikul, P.; Yodkeeree, S. Diclofenac Sensitizes Signet Ring Cell Gastric Carcinoma Cells to Cisplatin by Activating Autophagy and Inhibition of Survival Signal Pathways. Int. J. Mol. Sci. 2022, 23, 12066. [Google Scholar] [PubMed]
- Gao, F.; Li, R.; Wei, P.-F.; Ou, L.; Li, M.; Bai, Y.; Luo, W.-J.; Fan, Z. Synergistic anticancer effects of everolimus (RAD001) and Rhein on gastric cancer cells via phosphoinositide-3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR) pathway. Bioengineered 2022, 13, 6332–6342. [Google Scholar] [PubMed]
- Yao, C.; Wang, F. Inhibition of hypoxia-induced HIF-1α-mediated autophagy enhances the in vitro antitumor activity of rhein in pancreatic cancer cells. J. Appl. Toxicol. 2022, 42, 1937–1947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yung, K.K.-L.; Ko, J.K.-S. Therapeutic intervention in cancer by isoliquiritigenin from licorice: A natural antioxidant and redox regulator. Antioxidants 2022, 11, 1349. [Google Scholar]
- Cheng, T.; Wang, C.; Lu, Q.; Cao, Y.; Yu, W.; Li, W.; Liu, B.; Gao, X.; Lü, J.; Pan, X. Metformin inhibits the tumor-promoting effect of low-dose resveratrol, and enhances the anti-tumor activity of high-dose resveratrol by increasing its reducibility in triple negative breast cancer. Free Radic. Biol. Med. 2022, 180, 108–120. [Google Scholar] [CrossRef]
- Amirazodi, M.; Mehrabi, A.; Rajizadeh, M.A.; Bejeshk, M.A.; Esmaeilpour, K.; Daryanoosh, F.; Gaeini, A. The effects of combined resveratrol and high intensity interval training on the hippocampus in aged male rats: An investigation into some signaling pathways related to mitochondria. Iran. J. Basic Med. Sci. 2022, 25, 254. [Google Scholar]
- Chen, Y.; Li, Q.; Ren, S.; Chen, T.; Zhai, B.; Cheng, J.; Shi, X.; Song, L.; Fan, Y.; Guo, D. Investigation and experimental validation of curcumin-related mechanisms against hepatocellular carcinoma based on network pharmacology. J. Zhejiang Univ.-Sci. B 2022, 23, 682–698. [Google Scholar]
- Ashrafizadeh, M.; Tavakol, S.; Ahmadi, Z.; Roomiani, S.; Mohammadinejad, R.; Samarghandian, S. Therapeutic effects of kaempferol affecting autophagy and endoplasmic reticulum stress. Phytother. Res. 2020, 34, 911–923. [Google Scholar]
- Memete, A.R.; Timar, A.V.; Vuscan, A.N.; Miere, F.; Venter, A.C.; Vicas, S.I. Phytochemical composition of different botanical parts of Morus species, health benefits and application in Food Industry. Plants 2022, 11, 152. [Google Scholar] [CrossRef] [PubMed]
- Almatroodi, S.A.; Alsahli, M.A.; Almatroudi, A.; Verma, A.K.; Aloliqi, A.; Allemailem, K.S.; Khan, A.A.; Rahmani, A.H. Potential therapeutic targets of quercetin, a plant flavonol, and its role in the therapy of various types of cancer through the modulation of various cell signaling pathways. Molecules 2021, 26, 1315. [Google Scholar]
- Li, H.; Li, W.; Li, J.; Li, S.; Kuang, L.; Pang, F.; Jiang, H.; Jin, H.; Bian, X. Honokiol Microemulsion Causes Stage-Dependent Toxicity Via Dual Roles in Oxidation-Reduction and Apoptosis through FoxO Signaling Pathway. Cells 2022, 11, 3562. [Google Scholar] [CrossRef]
- Lu, J.; Yang, L.; Zhu, Q.; Gao, P.; Zhang, Z.; Wang, W.; Fu, Y. Ginsenoside induces apoptosis, autophagy and cell cycle arrest in gastric cancer cells by regulation of reactive oxygen species and activation of MAPK pathway. Trop. J. Pharm. Res. 2022, 21, 2071–2076. [Google Scholar] [CrossRef]
- Yang, S.; Li, F.; Lu, S.; Ren, L.; Bian, S.; Liu, M.; Zhao, D.; Wang, S.; Wang, J. Ginseng root extract attenuates inflammation by inhibiting the MAPK/NF-κB signaling pathway and activating autophagy and p62-Nrf2-Keap1 signaling in vitro and in vivo. J. Ethnopharmacol. 2022, 283, 114739. [Google Scholar]
- Moskot, M.; Montefusco, S.; Jakóbkiewicz-Banecka, J.; Mozolewski, P.; Węgrzyn, A.; Di Bernardo, D.; Węgrzyn, G.; Medina, D.L.; Ballabio, A.; Gabig-Cimińska, M. The phytoestrogen genistein modulates lysosomal metabolism and transcription factor EB (TFEB) activation. J. Biol. Chem. 2014, 289, 17054–17069. [Google Scholar]
- Rendón, J.P.; Cañas, A.I.; Correa, E.; Bedoya-Betancur, V.; Osorio, M.; Castro, C.; Naranjo, T.W. Evaluation of the Effects of Genistein In Vitro as a Chemopreventive Agent for Colorectal Cancer—Strategy to Improve Its Efficiency When Administered Orally. Molecules 2022, 27, 7042. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, X.; Zhang, Z.; Yin, M.; Chen, X.; Zhao, S.; Wu, L. Apigenin Induced Apoptosis by Downregulating Sulfiredoxin Expression in Cutaneous Squamous Cell Carcinoma. Oxidative Med. Cell. Longev. 2022, 2022, 8172866. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; He, J.; Luo, L.; Wang, K. Targeting the Interplay of Autophagy and ROS for Cancer Therapy: An Updated Overview on Phytochemicals. Pharmaceuticals 2023, 16, 92. [Google Scholar] [CrossRef]
- Pagano, M.; Mosca, L.; Vitiello, F.; Ilisso, C.P.; Coppola, A.; Borzacchiello, L.; Mele, L.; Caruso, F.P.; Ceccarelli, M.; Caraglia, M. Mi-RNA-888-5p is involved in S-adenosylmethionine antitumor effects in laryngeal squamous cancer cells. Cancers 2020, 12, 3665. [Google Scholar]
- Miao, Y.; Yang, T.; Yang, S.; Yang, M.; Mao, C. Protein nanoparticles directed cancer imaging and therapy. Nano Converg. 2022, 9, 1–21. [Google Scholar]
- Zhai, X.; Shan, S.; Wan, J.; Tian, H.; Wang, J.; Xin, L. Silver Nanoparticles Induce a Size-dependent Neurotoxicity to SH-SY5Y Neuroblastoma Cells via Ferritinophagy-mediated Oxidative Stress. Neurotox. Res. 2022, 40, 1369–1379. [Google Scholar]
- Wang, J.; Liu, N.; Su, Q.; Lv, Y.; Yang, C.; Zhan, H. Green Synthesis of Gold Nanoparticles and Study of Their Inhibitory Effect on Bulk Cancer Cells and Cancer Stem Cells in Breast Carcinoma. Nanomaterials 2022, 12, 3324. [Google Scholar]
- Subramaniyan, S.; Kamaraj, Y.; Kumaresan, V.; Kannaiyan, M.; David, E.; Ranganathan, B.; Selvaraj, V.; Balupillai, A. Green synthesized zinc oxide nanoparticles induce apoptosis by suppressing PI3K/Akt/mTOR signaling pathway in osteosarcoma MG63 cells. Glob. Transl. Med. 2022, 1, 1–12. [Google Scholar]
- Park, E.-J.; Choi, D.-H.; Kim, Y.; Lee, E.-W.; Song, J.; Cho, M.-H.; Kim, J.-H.; Kim, S.-W. Magnetic iron oxide nanoparticles induce autophagy preceding apoptosis through mitochondrial damage and ER stress in RAW264. 7 cells. Toxicol. Vitr. 2014, 28, 1402–1412. [Google Scholar] [CrossRef]
- Amjadi, S.; Hamishehkar, H.; Ghorbani, M. A novel smart PEGylated gelatin nanoparticle for co-delivery of doxorubicin and betanin: A strategy for enhancing the therapeutic efficacy of chemotherapy. Mater. Sci. Eng. C 2019, 97, 833–841. [Google Scholar]
- Ebrahimi, S.; Hashemy, S.I. MicroRNA-mediated redox regulation modulates therapy resistance in cancer cells: Clinical perspectives. Cell. Oncol. 2019, 42, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, Z.; Xuan, Z.; Xu, P.; Wang, W.; Chen, Z.; Wang, S.; Sun, G.; Xu, J.; Xu, Z. Novel role of miR-133a-3p in repressing gastric cancer growth and metastasis via blocking autophagy-mediated glutaminolysis. J. Exp. Clin. Cancer Res. 2018, 37, 1–22. [Google Scholar]
- Gu, D.-n.; Jiang, M.-j.; Mei, Z.; Dai, J.-j.; Dai, C.-y.; Fang, C.; Huang, Q.; Tian, L. microRNA-7 impairs autophagy-derived pools of glucose to suppress pancreatic cancer progression. Cancer Lett. 2017, 400, 69–78. [Google Scholar] [CrossRef]
- Clotaire, D.Z.J.; Zhang, B.; Wei, N.; Gao, R.; Zhao, F.; Wang, Y.; Lei, M.; Huang, W. MiR-26b inhibits autophagy by targeting ULK2 in prostate cancer cells. Biochem. Biophys. Res. Commun. 2016, 472, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, H.; Wu, F.; Wei, W.; Dang, Y.; Yang, B.; Ma, X.; Han, F.; Li, Y. Glycine decarboxylase induces autophagy and is downregulated by miRNA-30d-5p in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 1–14. [Google Scholar]
- Zhong, G.; Long, H.; Ma, S.; Shunhan, Y.; Li, J.; Yao, J. miRNA-335-5p relieves chondrocyte inflammation by activating autophagy in osteoarthritis. Life Sci. 2019, 226, 164–172. [Google Scholar] [CrossRef]
- Tong, L.; Yu, H.; Huang, X.; Shen, J.; Xiao, G.; Chen, L.; Wang, H.; Xing, L.; Chen, D. Current understanding of osteoarthritis pathogenesis and relevant new approaches. Bone Res. 2022, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ning, S.; Li, Z.; Ji, Z.; Fan, D.; Wang, K.; Wang, Q.; Hua, L.; Zhang, J.; Meng, X.; Yuan, Y. MicroRNA-494 suppresses hypoxia/reoxygenation-induced cardiomyocyte apoptosis and autophagy via the PI3K/AKT/mTOR signaling pathway by targeting SIRT1. Mol. Med. Rep. 2020, 22, 5231–5242. [Google Scholar] [CrossRef] [PubMed]
- Ilisso, C.P.; Delle Cave, D.; Mosca, L.; Pagano, M.; Coppola, A.; Mele, L.; Caraglia, M.; Cacciapuoti, G.; Porcelli, M. S-Adenosylmethionine regulates apoptosis and autophagy in MCF-7 breast cancer cells through the modulation of specific microRNAs. Cancer Cell Int. 2018, 18, 1–13. [Google Scholar] [CrossRef]
- Soni, M.; Patel, Y.; Markoutsa, E.; Jie, C.; Liu, S.; Xu, P.; Chen, H. Autophagy, cell viability, and chemoresistance are regulated by miR-489 in breast cancer. Mol. Cancer Res. 2018, 16, 1348–1360. [Google Scholar] [PubMed]
- Si, L.; Yang, Z.; Ding, L.; Zhang, D. Regulatory effects of lncRNAs and miRNAs on the crosstalk between autophagy and EMT in cancer: A new era for cancer treatment. J. Cancer Res. Clin. Oncol. 2022, 148, 547–564. [Google Scholar] [CrossRef]
- Guo, T.; He, K.; Chen, W.; Xu, M.; Sun, J.; Chen, Y.; Yang, Z. Construction and experimental validation of ferroptosis-related competing endogenous RNA networks in hepatocellular carcinoma based on WGCNA. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, N.; Zhao, N.; Hu, X.; He, X.; Xu, Y.; Chen, J.; Chen, W.; Liu, X.; Zhou, Z. Tumor-Suppressive and Oncogenic Roles of microRNA-149-5p in Human Cancers. Int. J. Mol. Sci. 2022, 23, 10823. [Google Scholar]
- Li, R.; Jin, Y.; Li, Q.; Sun, X.; Zhu, H.; Cui, H. MiR-93-5p targeting PTEN regulates the NMDA-induced autophagy of retinal ganglion cells via AKT/mTOR pathway in glaucoma. Biomed. Pharmacother. 2018, 100, 1–7. [Google Scholar] [CrossRef]
- Sun, D.; Li, C.; Liu, J.; Wang, Z.; Liu, Y.; Luo, C.; Chen, Y.; Wen, S. Expression profile of microRNAs in hypertrophic cardiomyopathy and effects of microRNA-20 in inducing cardiomyocyte hypertrophy through regulating gene MFN2. DNA Cell Biol. 2019, 38, 796–807. [Google Scholar] [CrossRef]
- Babaei, G.; Aziz, S.G.-G.; Jaghi, N.Z.Z. EMT, cancer stem cells and autophagy; The three main axes of metastasis. Biomed. Pharmacother. 2021, 133, 110909. [Google Scholar] [CrossRef]
- Liu, R.; Peng, L.; Zhou, L.; Huang, Z.; Zhou, C.; Huang, C. Oxidative Stress in Cancer Immunotherapy: Molecular Mechanisms and Potential Applications. Antioxidants 2022, 11, 853. [Google Scholar]
- Oršolić, N.; Jazvinšćak Jembrek, M. Molecular and cellular mechanisms of propolis and its polyphenolic compounds against cancer. Int. J. Mol. Sci. 2022, 23, 10479. [Google Scholar] [CrossRef]
- Azlan, A.; Sultana, S.; Huei, C.S.; Razman, M.R. Antioxidant, Anti-Obesity, Nutritional and Other Beneficial Effects of Different Chili Pepper: A Review. Molecules 2022, 27, 898. [Google Scholar] [CrossRef]
- Busta, L.; Dweikat, I.; Sato, S.J.; Qu, H.; Xue, Y.; Zhou, B.; Gan, L.; Yu, B.; Clemente, T.E.; Cahoon, E.B. Chemical and genetic variation in feral Cannabis sativa populations across the Nebraska climate gradient. Phytochemistry 2022, 200, 113206. [Google Scholar] [CrossRef]
- Naseer, F.; Ahmed, M.; Majid, A.; Kamal, W.; Phull, A.R. Green nanoparticles as multifunctional nanomedicines: Insights into anti-inflammatory effects, growth signaling and apoptosis mechanism in cancer. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Kaur, V.; Kumar, M.; Kumar, A.; Kaur, K.; Dhillon, V.S.; Kaur, S. Pharmacotherapeutic potential of phytochemicals: Implications in cancer chemoprevention and future perspectives. Biomed. Pharmacother. 2018, 97, 564–586. [Google Scholar] [CrossRef]
- Anand, K.; Tiloke, C.; Naidoo, P.; Chuturgoon, A. Phytonanotherapy for management of diabetes using green synthesis nanoparticles. J. Photochem. Photobiol. B Biol. 2017, 173, 626–639. [Google Scholar]
- Broekgaarden, M.; Anbil, S.; Bulin, A.-L.; Obaid, G.; Mai, Z.; Baglo, Y.; Rizvi, I.; Hasan, T. Modulation of redox metabolism negates cancer-associated fibroblasts-induced treatment resistance in a heterotypic 3D culture platform of pancreatic cancer. Biomaterials 2019, 222, 119421. [Google Scholar]
- Le, A.; Lane, A.N.; Hamaker, M.; Bose, S.; Gouw, A.; Barbi, J.; Tsukamoto, T.; Rojas, C.J.; Slusher, B.S.; Zhang, H. Glucose-independent glutamine metabolism via TCA cycling for proliferation and survival in B cells. Cell Metab. 2012, 15, 110–121. [Google Scholar] [PubMed]
- Park, H.; Kim, J.; Shin, C.; Lee, S. Intersection between redox homeostasis and autophagy: Valuable insights into neurodegeneration. Antioxidants 2021, 10, 694. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Li, P.; Zhang, D.; Shen, L.; Dong, K.; Wu, M.; Ou, Z.; Shi, D. Redox homeostasis protects mitochondria through accelerating ROS conversion to enhance hypoxia resistance in cancer cells. Sci. Rep. 2016, 6, 22831. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Interactions between reactive oxygen species and autophagy: Special issue: Death mechanisms in cellular homeostasis. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2021, 1868, 119041. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kim, H.S.; Seo, Y.R. Understanding of ROS-inducing strategy in anticancer therapy. Oxidative Med. Cell. Longev. 2019, 2019, 5381692. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Synthetic Drug | Model | Redox Mechanism | Autophagy Modulation | Reference |
|---|---|---|---|---|
| Ammonium chloride (NH4Cl), bafilomycin A (BafA), 3-methyladenine (3-MA), and chloroquine | Esophageal squamous cell carcinoma | Modulating redox status and nucleotide metabolism | Inhibition | [71] |
| KS10076 | Cancer stem cells | Induces ROS-mediated STAT3 degradation | Induction | [72] |
| FK866 | Pancreatic Cancer Cells | Decreasing oxidized (NAD+) to reduced (NADH) nicotinamide adenine dinucleotide ratio | Autophagy-mediated cell death | [74] |
| Doxorubicin | Tumor cells | Apoptosis/ferroptosis pathway | Impaired autophagy | [75] |
| Nicotinamide | Triple-negative breast cancer | Mitochondrial dysfunction and ROS activation | Autophagy modulation | [76] |
| Metformin | Ultraviolet (UVA)-exposed mice | Elevated oxidative stress | Enhanced autophagic flux | [73] |
| Diclofenac (DCF) | Cisplatin-resistant signet ring cell gastric carcinoma cells (KATO/DDP) | Reduction in antioxidant enzyme expression while inhibiting Nrf2 activity | Activation | [77] |
| Rapamycin | MiaPaCa-2 and PANC-1 pancreatic cancer cells | Inhibition of HIF-1α-mediated autophagy | Enhanced autophagy | [79] |
| Everolimus (RAD001) | Human gastric cancer cells (MGC-803) | Phosphorylation of PI3K/Akt/mTOR | Autophagy induction | [78] |
| Natural Product | Model | Redox Mechanism | Autophagy Modulation | Reference |
|---|---|---|---|---|
| Isoliquiritigenin | Pancreatic cancer | ROS-autophagic redox homeostasis | Autophagy modulation | [80] |
| Metformin | Triple-negative breast cancer | Increased catalase activity and NAD(P)H level | Induction | [81] |
| Curcumin | Hepatocellular carcinoma | Promoted apoptosis via the p53 pathway | Autophagy induction | [83] |
| Resveratrol | Albino Wistar rats | Increased NAD+/NADH, SOD2, and AMPK levels | Autophagy modulation | [82] |
| Honokiol | Neuroblastoma cells | Oxidation–reduction | Autophagy induction | [87] |
| Ginsenoside | BGC-823 human gastric cancer cell line | Modulates ROS and MAPK | Activation | [88] |
| Oxyresveratrol | Several cancer models | Increased lysosomal activity | Autophagy induction | [85] |
| Genistein | Colorectal cancer | Increased ROS production | Autophagy modulation | [91] |
| Apigenin | Cutaneous squamous cell carcinoma patients | Maintained redox | Autophagy modulation | [92] |
| Ginseng root extract | RAW264.7 cells | MAPK/NF-kB signaling | Activation | [89] |
| Nanoparticle | Model | Redox Mechanism | Autophagic Condition | Reference |
|---|---|---|---|---|
| Silver | Human neuroblastoma cancer cell line (SH-SY5Y) | Cellular redox homeostasis | Autophagy inhibition | [96] |
| Gold | Bulk cancer cells and cancer stem cells in breast carcinoma | Redox homeostasis | Autophagy modulation | [97] |
| Zinc oxide | MG63 human osteosarcoma cells | Suppressed P13K/AKT/mTOR signaling | Autophagy induction | [98] |
| Magnetic iron oxide | RAW264.7 cells | ER stress homeostasis | Autophagy induction | [99] |
| PEGylated gelatin | MCF-7 cells | Targeting sequestosome 1 (SQSTM1) and cathepsin F | Autophagy induction | [100] |
| MicroRNAs | Model | Redox Mechanism | Autophagy Modulation | Reference |
|---|---|---|---|---|
| microRNA (miRNA) 133a-3p | Patient-derived xenograft model and human gastric cancer organoid model | Glutaminolysis | Autophagy-mediated | [102] |
| miRNA-7 | Pancreatic cancer | Reprogrammed metabolic homeostasis | Autophagy modulation | [103] |
| miRNA-26b | Prostate cancer cells | ULK2 is a direct target of miRNA-26b | Autophagy inhibition | [104] |
| miRNA-30d-5p | Hepatocellular carcinoma | Decreased ROS-mediated ubiquitination of cofilin | Autophagy inhibition | [105] |
| miRNA-335-5p | Human osteoarthritis chondrocytes | Anti-inflammatory | Autophagy induction | [106] |
| miRNA-93 | Retinal ganglion cells | AKT/mTOR | Autophagy induction | [114] |
| miRNA-20a-5p | Human hepatocellular cancer cells | Downregulated hypertrophic cardiomyopathy | Autophagy modulation | [115] |
| miRNA-494 | Acute myocardial infarction | PI3K/AKT/mTOR signaling | Autophagy induction | [108] |
| miRNA-486-5p | MCF-7 breast cancer cells | Modulation of miRNA expression profile | Autophagy induction | [109] |
| miRNA-489 | Breast cancer cells | ROS regulation | Autophagy induction | [110] |
| Long noncoding RNAs | Several cancers | Maintained redox balance | Autophagy modulation | [111] |
| Competing endogenous RNAs | Hepatocellular carcinoma | Ferroptosis-related competing endogenous RNA | Autophagy regulation | [112] |
| miRNA-149-5p networks | Human cancers | Lipogenesis and vascular endothelial cells | Autophagy modulation | [113] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.A.; Ahmed, K.R.; Haque, F.; Park, M.N.; Kim, B. Recent Advances in Cellular Signaling Interplay between Redox Metabolism and Autophagy Modulation in Cancer: An Overview of Molecular Mechanisms and Therapeutic Interventions. Antioxidants 2023, 12, 428. https://doi.org/10.3390/antiox12020428
Rahman MA, Ahmed KR, Haque F, Park MN, Kim B. Recent Advances in Cellular Signaling Interplay between Redox Metabolism and Autophagy Modulation in Cancer: An Overview of Molecular Mechanisms and Therapeutic Interventions. Antioxidants. 2023; 12(2):428. https://doi.org/10.3390/antiox12020428
Chicago/Turabian StyleRahman, Md. Ataur, Kazi Rejvee Ahmed, Farzana Haque, Moon Nyeo Park, and Bonglee Kim. 2023. "Recent Advances in Cellular Signaling Interplay between Redox Metabolism and Autophagy Modulation in Cancer: An Overview of Molecular Mechanisms and Therapeutic Interventions" Antioxidants 12, no. 2: 428. https://doi.org/10.3390/antiox12020428