
Protein-Mediated Carotenoid Delivery Suppresses the Photoinducible Oxidation of Lipofuscin in Retinal Pigment Epithelial Cells
 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of the Lipofuscin Granules
2.2. ARPE-19 Cell Preparation
2.2.1. ARPE-19 Cell Line Culturing
2.2.2. Enrichment of ARPE-19 Cells with Lipofuscin Granules
2.2.3. Photo-Oxidation Protocol
2.2.4. Cell Viability Assessment Protocol
2.2.5. Production of Recombinant BmCBP Carotenoprotein as a Holoform with Zeaxanthin
2.2.6. Administration of BmCBP Complexed with ZEA to ARPE-19 Cells Fed with Lipofuscin Granules
2.3. Fluorescence Microscopy Measurements
2.3.1. Laser Confocal Microscopy
2.3.2. Fluorescence Lifetime Imaging Microscopy
2.3.3. FLIM Data Analysis
3. Results
3.1. Characteristics of Lipofuscin Granules-Fed RPE Cell Samples
3.2. Intracellular Localization of Lipofuscin Granules in ARPE-19 Cells
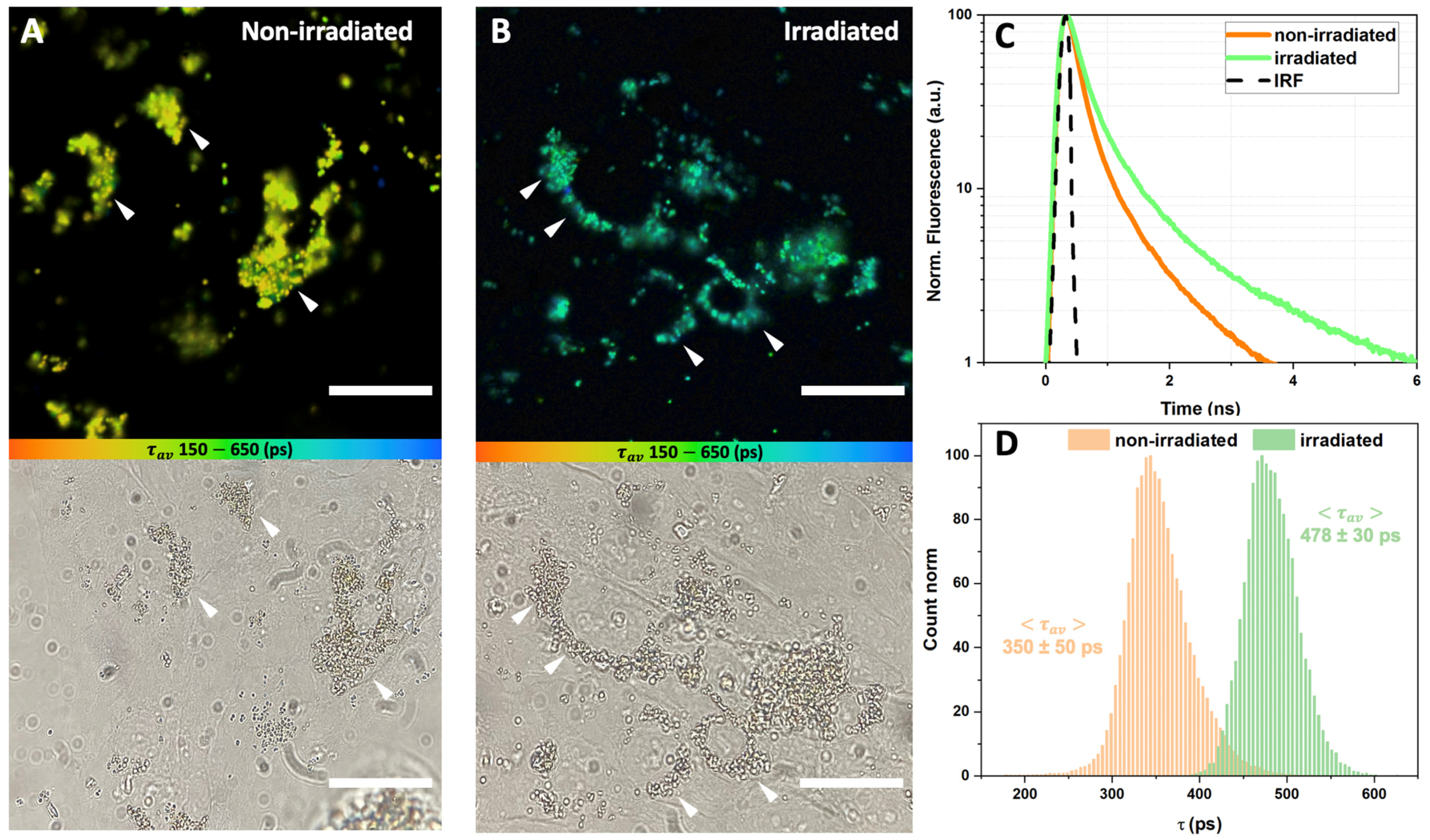
3.3. Fluorescence Lifetime Imaging (FLIM) of Lipofuscin Granules-Fed RPE Cells
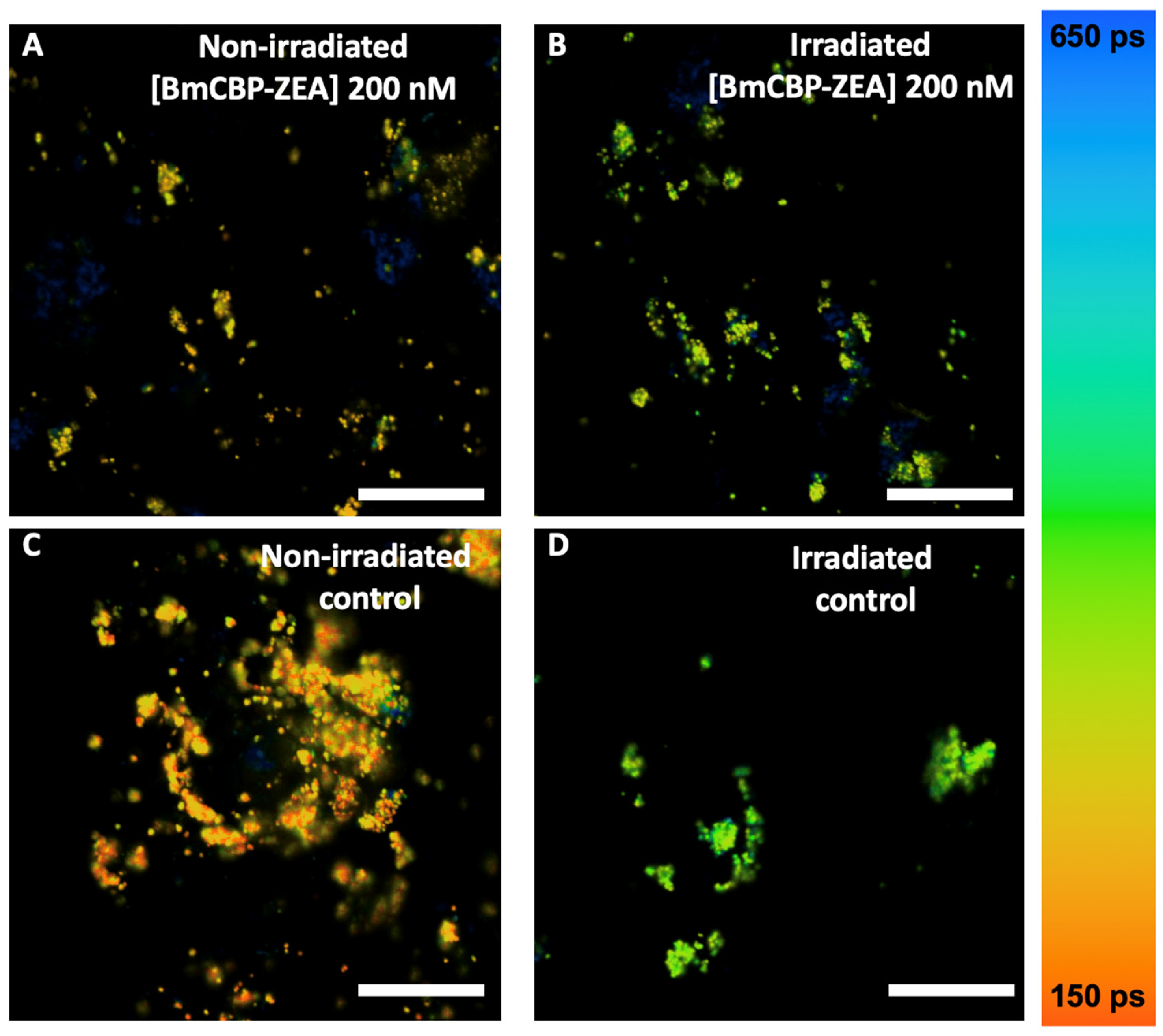
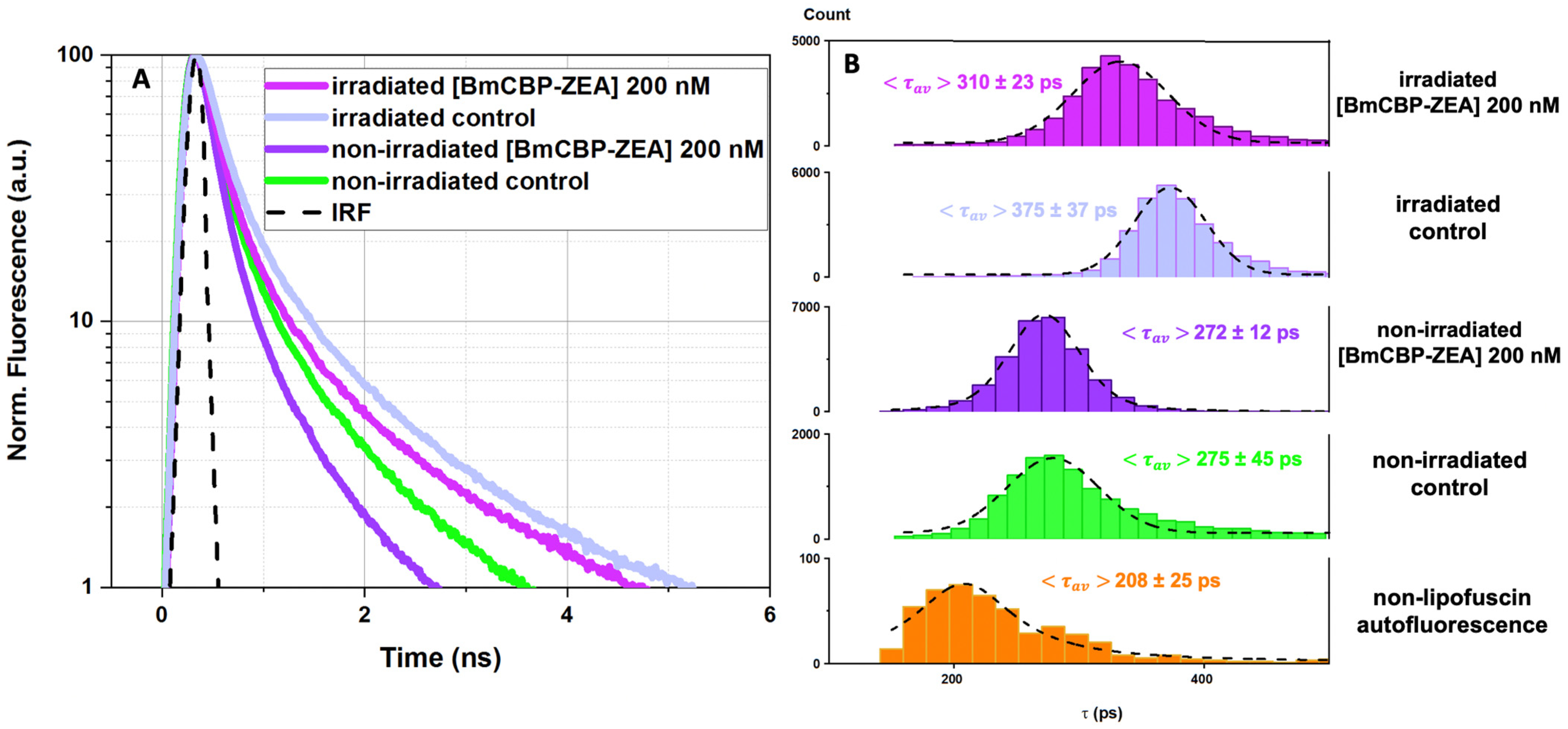
3.4. Effect of the Carotenoprotein-Mediated Carotenoid Delivery on the Photo-Oxidation of Lipofuscin in ARPE-19 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geng, L.; Wihlmark, U.; Algvere, P.V. Lipofuscin Accumulation in Iris Pigment Epithelial Cells Exposed to Photoreceptor Outer Segments. Exp. Eye Res. 1999, 69, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Boulton, M.; Dontsov, A.; Jarvis-Evans, J.; Ostrovsky, M.; Svistunenko, D. Lipofuscin Is a Photoinducible Free Radical Generator. J. Photochem. Photobiol. B Biol 1993, 19, 201–204. [Google Scholar]
- Yakovleva, M.; Dontsov, A.; Trofimova, N.; Sakina, N.; Kononikhin, A.; Aybush, A.; Gulin, A.; Feldman, T.; Ostrovsky, M. Lipofuscin Granule Bisretinoid Oxidation in the Human Retinal Pigment Epithelium Forms Cytotoxic Carbonyls. Int. J. Mol. Sci. 2022, 23, 222. [Google Scholar] [CrossRef]
- Pan, C.; Banerjee, K.; Lehmann, G.L.; Almeida, D.; Hajjar, K.A.; Benedicto, I.; Jiang, Z.; Radu, R.A.; Thompson, D.H.; Rodriguez-Boulan, E.; et al. Lipofuscin Causes Atypical Necroptosis through Lysosomal Membrane Permeabilization. Proc. Natl. Acad. Sci. USA 2021, 118, e2100122118. [Google Scholar] [CrossRef] [PubMed]
- Davies, S.; Elliott, M.H.; Floor, E.; George Truscott, T.; Zareba, M.; Sarna, T.; Shamsi, F.A.; Boulton, M.E. Photocytotoxicity of Lipofuscin in Human Retinal Pigment Epithelial Cells. Free Radic. Biol. Med. 2001, 31, 256–265. [Google Scholar] [PubMed]
- Sparrow, J.R.; Duncker, T. Fundus Autofluorescence and RPE Lipofuscin in Are-Related Macular Degeneration. J. Clin. Med. 2014, 3, 1302–1321. [Google Scholar] [CrossRef]
- Kathleen Dorey, C.; Wu, G.; Ebensrein, D.; Garsd, A. Cell Loss in the Aging Retina Relationship to Lipofuscin Accumulation and Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 1989, 30, 1691–1699. [Google Scholar]
- Lu, L.J.; Liu, J.; Adelman, R.A. Novel Therapeutics for Stargardt Disease. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 1057–1062. [Google Scholar] [CrossRef]
- Kennedy, C.J.; Rakoczy, P.E.; Constable, I.J. Lipofuscin of the Retinal Pigment Epithelium: A Review. Eye 1995, 9, 763–771. [Google Scholar]
- Schutt, F.; Ueberle, B.; Schnölzer, M.; Holz, F.G.; Kopitz, J. Proteome Analysis of Lipofuscin in Human Retinal Pigment Epithelial Cells. FEBS Lett. 2002, 528, 217–221. [Google Scholar] [CrossRef]
- Ng, K.P.; Gugiu, B.; Renganathan, K.; Davies, M.W.; Gu, X.; Crabb, J.S.; Kim, S.R.; Rózanowska, M.B.; Bonilha, V.L.; Rayborn, M.E.; et al. Retinal Pigment Epithelium Lipofuscin Proteomics. Mol. Cell. Proteom. 2008, 7, 1397–1405. [Google Scholar] [CrossRef]
- Sparrow, J.R.; Wu, Y.; Kim, C.Y.; Zhou, J. Phospholipid Meets All-Trans-Retinal: The Making of RPE Bisretinoids. J. Lipid Res. 2010, 51, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Sakai, N.; Decatur, J.; Nakanishi, K.; Eldred, G.E. Ocular Age Pigment “A2E”: An Unprecedented Pyridinium Bisretinoid. J. Am. Chem. Soc. 1996, 118, 1559–1560. [Google Scholar] [CrossRef]
- Ben-Shabat, S.; Parish, C.A.; Vollmer, H.R.; Itagaki, Y.; Fishkin, N.; Nakanishi, K.; Sparrow, J.R. Biosynthetic Studies of A2E, a Major Fluorophore of Retinal Pigment Epithelial Lipofucin. J. Biol. Chem. 2002, 277, 7183–7190. [Google Scholar] [CrossRef]
- Sparrow, J.R.; Fishkin, N.; Zhou, J.; Cai, B.; Jang, Y.P.; Krane, S.; Itagaki, Y.; Nakanishi, K. A2E, a Byproduct of the Visual Cycle. Vis. Res. 2003, 43, 2983–2990. [Google Scholar] [CrossRef]
- Wielgus, A.R.; Chignell, C.F.; Ceger, P.; Roberts, J.E. Comparison of A2E Cytotoxicity and Phototoxicity with All-Trans-Retinal in Human Retinal Pigment Epithelial Cells. Photochem. Photobiol. 2010, 86, 781–791. [Google Scholar] [CrossRef]
- Różanowska, M.; Wessels, J.; Boulton, M.; Burke, J.M.; Rodgers, M.A.J.; George Truscott, T.; Sarna, T. Blue Light-Induced Singlet Oxygen Generation by Retinal Lipofuscin in Non-Polar Media. Free Radic. Biol. Med. 1998, 24, 1107–1112. [Google Scholar]
- Dontsov, A.E.; Glickman, R.D.; Ostrovsky, M.A. Retinal Pigment Epithelium Pigment Granules Stimulate the Photo-Oxidation of Unsaturated Fatty Acids. Free Radic. Biol. Med. 1999, 26, 1436–1446. [Google Scholar]
- Ruan, Y.; Jiang, S.; Gericke, A. Age-Related Macular Degeneration: Role of Oxidative Stress and Blood Vessels. Int. J. Mol. Sci. 2021, 22, 1296. [Google Scholar] [CrossRef] [PubMed]
- Wihlmark, U.; Wrigstad, A.; Roberg, K.; Erik Nilsson, S.G.; Brunk, U.T. Lipofuscin Accumulation in Cultered Retinal Pigment Epithelial Cells Causes Enhanced Sensitivity to Blue Light Irradiation. Free Radic. Biol. Med. 1997, 22, 1229–1234. [Google Scholar]
- Sparrow, J.R.; Nakanishi, K.; Parish, C.A. The Lipofuscin Fluorophore A2E Mediates Blue Light-Induced Damage to Retinal Pigmented Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1981–1989. [Google Scholar]
- Sundelin, S.P.; Nilsson, S.E.G. Lipofuscin-Formation in Retinal Pigment Epithelial Cells Is Reduced by Antioxidants. Free Radic. Biol. Med. 2001, 31, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.P.; Zhou, J.; Nakanishi, K.; Sparrow, J.R. Anthocyanins Protect Against A2E Photooxidation and Membrane Permeabilization in Retinal Pigment Epithelial Cells. Photochem. Photobiol. 2005, 81, 529–536. [Google Scholar] [PubMed]
- Khoo, H.E.; Ng, H.S.; Yap, W.S.; Goh, H.J.H.; Yim, H.S. Nutrients for Prevention of Macular Degeneration and Eye-Related Diseases. Antioxidants 2019, 8, 85. [Google Scholar] [CrossRef]
- Sparrow, J.R.; Vollmer-Snarr, H.R.; Zhou, J.; Jang, Y.P.; Jockusch, S.; Itagaki, Y.; Nakanishi, K. A2E-Epoxides Damage DNA in Retinal Pigment Epithelial Cells. Vitamin E and Other Antioxidants Inhibit A2E-Epoxide Formation. J. Biol. Chem. 2003, 278, 18207–18213. [Google Scholar] [CrossRef] [PubMed]
- Alaimo, A.; di Santo, M.C.; Domínguez Rubio, A.P.; Chaufan, G.; García Liñares, G.; Pérez, O.E. Toxic Effects of A2E in Human ARPE-19 Cells Were Prevented by Resveratrol: A Potential Nutritional Bioactive for Age-Related Macular Degeneration Treatment. Arch. Toxicol. 2020, 94, 553–572. [Google Scholar] [CrossRef]
- Mrowicka, M.; Mrowicki, J.; Kucharska, E.; Majsterek, I. Lutein and Zeaxanthin and Their Roles in Age-Related Macular Degeneration—Neurodegenerative Disease. Nutrients 2022, 14, 827. [Google Scholar]
- Rózanowski, B.; Cuenco, J.; Davies, S.; Shamsi, F.A.; Zadło, A.; Dayhaw-Barker, P.; Rózanowska, M.; Sarna, T.; Boulton, M.E. The Phototoxicity of Aged Human Retinal Melanosomes. Photochem. Photobiol. 2008, 84, 650–657. [Google Scholar] [CrossRef]
- Li, B.; Ahmed, F.; Bernstein, P.S. Studies on the Singlet Oxygen Scavenging Mechanism of Human Macular Pigment. Arch. Biochem. Biophys. 2010, 504, 56–60. [Google Scholar] [CrossRef]
- Kim, S.R.; Nakanishi, K.; Itagaki, Y.; Sparrow, J.R. Photooxidation of A2-PE, a Photoreceptor Outer Segment Fluorophore, and Protection by Lutein and Zeaxanthin. Exp. Eye Res. 2006, 82, 828–839. [Google Scholar] [CrossRef]
- Roberts, J.E.; Dennison, J. The Photobiology of Lutein and Zeaxanthin in the Eye. J. Ophthalmol. 2015, 2015, 687173. [Google Scholar] [CrossRef] [PubMed]
- Olchawa, M.M.; Furso, J.A.; Szewczyk, G.M.; Sarna, T.J. Lipofuscin-Mediated Photic Stress Inhibits Phagocytic Activity of ARPE-19 Cells; Effect of Donors’ Age and Antioxidants. Free Radic. Res. 2017, 51, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, R.; Gorusupudi, A.; Li, B.; Blount, J.D.; Nwagbo, U.; Kim, H.J.; Sparrow, J.R.; Bernstein, P.S. Lutein and Zeaxanthin Reduce A2E and Iso-A2E Levels and Improve Visual Performance in Abca4−/−/Bco2−/− Double Knockout Mice. Exp. Eye Res. 2021, 209, 108680. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Gencoglu, H.; Akdemir, F.; Orhan, C.; Tuzcu, M.; Sahin, N.; Yilmaz, I.; Juturu, V. Lutein and Zeaxanthin Isomers May Attenuate Photo-Oxidative Retinal Damage via Modulation of G Protein-Coupled Receptors and Growth Factors in Rats. Biochem. Biophys. Res. Commun. 2019, 516, 163–170. [Google Scholar] [CrossRef]
- José, R.; Torres, A.; Correa, C.R. The Role of Non-Enzymatic Antioxidants on Age-Related Macular Degeneration. Front. Drug CHemistry Clin. Res. 2022, 5, 1–19. [Google Scholar] [CrossRef]
- Lornejad-Schäfer, M.R.; Lambert, C.; Breithaupt, D.E.; Biesalski, H.K.; Frank, J. Solubility, Uptake and Biocompatibility of Lutein and Zeaxanthin Delivered to Cultured Human Retinal Pigment Epithelial Cells in Tween40 Micelles. Eur. J. Nutr. 2007, 46, 79–86. [Google Scholar] [CrossRef]
- Ibrahim, A.E.; Shafaa, M.W.; Khedr, M.H.; Rashed, R.F. Comparative Study between Lutein and Its Liposomal Form on Cisplatin-Induced Retinal Injury in Rabbits. Cutan. Ocul. Toxicol. 2019, 38, 279–285. [Google Scholar] [CrossRef]
- Algan, A.H.; Gungor-Ak, A.; Karatas, A. Nanoscale Delivery Systems of Lutein: An Updated Review from a Pharmaceutical Perspective. Pharmaceutics 2022, 14, 1852. [Google Scholar] [CrossRef]
- Ma, Y.; You, T.; Wang, J.; Jiang, Y.; Niu, J. Research Progress on Construction of Lutein-Loaded Nano Delivery System and Their Improvements on the Bioactivity. Coatings 2022, 12, 1449. [Google Scholar]
- Tan, C.; Xue, J.; Lou, X.; Abbas, S.; Guan, Y.; Feng, B.; Zhang, X.; Xia, S. Liposomes as Delivery Systems for Carotenoids: Comparative Studies of Loading Ability, Storage Stability and in Vitro Release. Food Funct. 2014, 5, 1232–1240. [Google Scholar] [CrossRef]
- Shafaa, M.W.I.; Diehl, H.A.; Socaciu, C. The Solubilisation Pattern of Lutein, Zeaxanthin, Canthaxanthin and β-Carotene Differ Characteristically in Liposomes, Liver Microsomes and Retinal Epithelial Cells. Biophys. Chem. 2007, 129, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Petyaev, I.M.; Zigangirova, N.A.; Tsibezov, V.V.; Morgunova, E.Y.; Bondareva, N.E.; Kyle, N.H.; Bashmakov, Y.K. Association with Monoclonal Antibody Promotes Intracellular Delivery of Lycopene. Monoclon. Antib. Immunodiagn. Immunother. 2018, 37, 147–152. [Google Scholar] [CrossRef]
- Borel, P.; Desmarchelier, C. Bioavailability of Fat-Soluble Vitamins and Phytochemicals in Humans: Effects of Genetic Variation. Annu. Rev. Nutr. 2018, 38, 69–96. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Vachali, P.; Chang, F.Y.; Gorusupudi, A.; Arunkumar, R.; Shi, L.; Rognon, G.T.; Frederick, J.M.; Bernstein, P.S. HDL Is the Primary Transporter for Carotenoids from Liver to Retinal Pigment Epithelium in Transgenic ApoA-I−/−/Bco2−/− Mice. Arch. Biochem. Biophys. 2022, 716, 109111. [Google Scholar] [CrossRef]
- Bhosale, P.; Larson, A.J.; Frederick, J.M.; Southwick, K.; Thulin, C.D.; Bernstein, P.S. Identification and Characterization of a Pi Isoform of Glutathione S-Transferase (GSTP1) as a Zeaxanthin-Binding Protein in the Macula of the Human Eye. J. Biol. Chem. 2004, 279, 49447–49454. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Vachali, P.; Frederick, J.M.; Bernstein, P.S. Identification of StARD3 as a Lutein-Binding Protein in the Macula of the Primate Retina. Biochemistry 2011, 50, 2541–2549. [Google Scholar] [CrossRef]
- Li, B.; Vachali, P.; Bernstein, P.S. Human Ocular Carotenoid-Binding Proteins. Photochem. Photobiol. Sci. 2010, 9, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Bandara, S.; von Lintig, J. Aster La Vista: Unraveling the Biochemical Basis of Carotenoid Homeostasis in the Human Retina. BioEssays 2022, 44, 2200133. [Google Scholar] [CrossRef]
- Maksimov, E.G.; Moldenhauer, M.; Shirshin, E.A.; Parshina, E.A.; Sluchanko, N.N.; Klementiev, K.E.; Tsoraev, G.V.; Tavraz, N.N.; Willoweit, M.; Schmitt, F.J.; et al. A Comparative Study of Three Signaling Forms of the Orange Carotenoid Protein. Photosynth. Res. 2016, 130, 389–401. [Google Scholar] [CrossRef]
- Hagemann, M.; Eisenhut, M.; Hackenberg, C.; Bauwe, H. Pathway and Importance of Photorespiratory 2-Phosphoglycolate Metabolism in Cyanobacteria; Springer: New York, NY, USA, 2010; Volume 675, ISBN 9781441915276. [Google Scholar]
- Slonimskiy, Y.B.; Zupnik, A.O.; Varfolomeeva, L.A.; Boyko, K.M.; Maksimov, E.G.; Sluchanko, N.N. A Primordial Orange Carotenoid Protein: Structure, Photoswitching Activity and Evolutionary Aspects. Int. J. Biol. Macromol. 2022, 222, 167–180. [Google Scholar] [CrossRef]
- Slonimskiy, Y.B.; Egorkin, N.A.; Ashikhmin, A.A.; Friedrich, T.; Maksimov, E.G.; Sluchanko, N.N. Reconstitution of the Functional Carotenoid-Binding Protein from Silkworm in E. Coli. Int. J. Biol. Macromol. 2022, 214, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Maksimov, E.G.; Zamaraev, A.V.; Parshina, E.Y.; Slonimskiy, Y.B.; Slastnikova, T.A.; Abdrakhmanov, A.A.; Babaev, P.A.; Efimova, S.S.; Ostroumova, O.S.; Stepanov, A.V.; et al. Soluble Cyanobacterial Carotenoprotein as a Robust Antioxidant Nanocarrier and Delivery Module. Antioxidants 2020, 9, 869. [Google Scholar] [CrossRef] [PubMed]
- Sluchanko, N.N.; Slonimskiy, Y.B.; Egorkin, N.A.; Varfolomeeva, L.A.; Faletrov, Y.V.; Moysenovich, A.M.; Parshina, E.Y.; Friedrich, T.; Maksimov, E.G.; Boyko, K.M.; et al. Silkworm Carotenoprotein as an Efficient Carotenoid Extractor, Solubilizer and Transporter. Int. J. Biol. Macromol. 2022, 223, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Feldman, T.B.; Yakovleva, M.A.; Arbukhanova, P.M.; Borzenok, S.A.; Kononikhin, A.S.; Popov, I.A.; Nikolaev, E.N.; Ostrovsky, M.A. Changes in Spectral Properties and Composition of Lipofuscin Fluorophores from Human-Retinal-Pigment Epithelium with Age and Pathology. Anal. Bioanal. Chem. 2015, 407, 1075–1088. [Google Scholar] [CrossRef]
- Moldenhauer, M.; Sluchanko, N.N.; Buhrke, D.; Zlenko, D.V.; Tavraz, N.N.; Schmitt, F.J.; Hildebrandt, P.; Maksimov, E.G.; Friedrich, T. Assembly of Photoactive Orange Carotenoid Protein from Its Domains Unravels a Carotenoid Shuttle Mechanism. Photosynth. Res. 2017, 133, 327–341. [Google Scholar] [CrossRef]
- Slonimskiy, Y.B.; Egorkin, N.A.; Friedrich, T.; Maksimov, E.G.; Sluchanko, N.N. Microalgal Protein AstaP Is a Potent Carotenoid Solubilizer and Delivery Module with a Broad Carotenoid Binding Repertoire. FEBS J. 2022, 289, 999–1022. [Google Scholar] [CrossRef]
- Sluchanko, N.N.; Slonimskiy, Y.B.; Egorkin, N.A.; Varfolomeeva, L.A.; Kleymenov, S.Y.; Minyaev, M.E.; Faletrov, Y.V.; Moysenovich, A.M.; Parshina, E.Y.; Friedrich, T.; et al. Structural Basis for the Carotenoid Binding and Transport Function of a START Domain. Structure 2022, 30, 1647–1659. [Google Scholar] [CrossRef]
- Sparrow, J.R.; Wu, Y.; Nagasaki, T.; Yoon, K.D.; Yamamoto, K.; Zhou, J. Fundus Autofluorescence and the Bisretinoids of Retina. Photochem. Photobiol. Sci. 2010, 9, 1480–1489. [Google Scholar] [CrossRef]
- Feng, J.; Chen, X.; Sun, X.; Wang, F.; Sun, X. Expression of Endoplasmic Reticulum Stress Markers GRP78 and CHOP Induced by Oxidative Stress in Blue Light-Mediated Damage of A2E-Containing Retinal Pigment Epithelium Cells. Ophthalmic Res. 2014, 52, 224–233. [Google Scholar] [CrossRef]
- Nordgaard, C.L.; Karunadharma, P.P.; Feng, X.; Olsen, T.W.; Ferrington, D.A. Mitochondrial Proteomics of the Retinal Pigment Epithelium at Progressive Stages of Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 2008, 49, 2848–2855. [Google Scholar] [CrossRef]
- Feldman, T.; Ostrovskiy, D.; Yakovleva, M.; Dontsov, A.; Borzenok, S.; Ostrovsky, M. Lipofuscin-Mediated Photic Stress Induces a Dark Toxic Effect on ARPE-19 Cells. Int. J. Mol. Sci. 2022, 23, 12234. [Google Scholar] [CrossRef] [PubMed]
- Hütter, E.; Skovbro, M.; Lener, B.; Prats, C.; Rabøl, R.; Dela, F.; Jansen-Dürr, P. Oxidative Stress and Mitochondrial Impairment Can Be Separated from Lipofuscin Accumulation in Aged Human Skeletal Muscle. Aging Cell 2007, 6, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shabat, S.; Itagaki, Y.; Jockusch, S.; Sparrow, J.R.; Turro, N.J.; Nakanishi, K. Formation of a Nonaoxirane from A2E, a Lipofuscin Fluorophore Related to Macular Degeneration, and Evidence of Singlet Oxygen Involvement. Angew. Chem. Int. Ed. 2002, 41, 814–817. [Google Scholar]
- Shcheslavskiy, V.I.; Shirmanova, M.V.; Dudenkova, V.V.; Lukyanov, K.A.; Gavrina, A.I.; Shumilova, A.V.; Zagaynova, E.; Becker, W. Fluorescence Time-Resolved Macroimaging. Opt. Lett. 2018, 43, 3152. [Google Scholar] [CrossRef] [PubMed]
- Pospíšil, P.; Prasad, A.; Rác, M. Mechanism of the Formation of Electronically Excited Species by Oxidative Metabolic Processes: Role of Reactive Oxygen Species. Biomolecules 2019, 9, 258. [Google Scholar] [CrossRef] [PubMed]
- Semenov, A.N.; Yakimov, B.P.; Rubekina, A.A.; Gorin, D.A.; Drachev, V.P.; Zarubin, M.P.; Velikanov, A.N.; Lademann, J.; Fadeev, V.V.; Priezzhev, A.V.; et al. The Oxidation-Induced Autofluorescence Hypothesis: Red Edge Excitation and Implications for Metabolic Imaging. Molecules 2020, 25, 258. [Google Scholar] [CrossRef]
- Hashimoto, H.; Uragami, C.; Yukihira, N.; Gardiner, A.T.; Cogdell, R.J. Understanding/Unravelling Carotenoid Excited Singlet States. J. R. Soc. Interface 2018, 15, 20180026. [Google Scholar]
- Murillo, A.G.; Hu, S.; Fernandez, M.L. Zeaxanthin: Metabolism, Properties, and Antioxidant Protection of Eyes, Heart, Liver, and Skin. Antioxidants 2019, 8, 390. [Google Scholar] [CrossRef]
- Holz, F.G.; Fleckenstein, M.; Schmitz-Valckenberg, S.; Bird, A.C. Evaluation of Fundus Autofluorescence Images. In Atlas of Fundus Autofluorescence Imaging; Holz, F.G., Schmitz-Valckenberg, S., Spaide, R.F., Bird, A.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 71–76. [Google Scholar]
- Pole, C.; Ameri, H. Fundus Autofluorescence and Clinical Applications. J. Ophthalmic. Vis. Res. 2021, 16, 432–461. [Google Scholar] [CrossRef]
- Wu, Y.; Yanase, E.; Feng, X.; Siegel, M.M.; Sparrow, J.R. Structural Characterization of Bisretinoid A2E Photocleavage Products and Implications for Age-Related Macular Degeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 7275–7280. [Google Scholar] [CrossRef]
- Delori, F.C.; Goger, D.G.; Dorey, C.K. Age-Related Accumulation and Spatial Distribution of Lipofuscin in RPE of Normal Subjects. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1855–1866. [Google Scholar] [PubMed]
- Feldman, T.B.; Yakovleva, M.A.; Larichev, A.V.; Arbukhanova, P.M.; Radchenko, A.S.; Borzenok, S.A.; Kuzmin, V.A.; Ostrovsky, M.A. Spectral Analysis of Fundus Autofluorescence Pattern as a Tool to Detect Early Stages of Degeneration in the Retina and Retinal Pigment Epithelium. Eye 2018, 32, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Sauer, L.; Gensure, R.H.; Andersen, K.M.; Kreilkamp, L.; Hageman, G.S.; Hammer, M.; Bernstein, P.S. Patterns of Fundus Autofluorescence Lifetimes in Eyes of Individuals with Nonexudative Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 2018, 59, AMD65–AMD77. [Google Scholar] [CrossRef]
- Dysli, C.; Schürch, K.; Pascal, E.; Wolf, S.; Zinkernagel, M.S. Fundus Autofluorescence Lifetime Patterns in Retinitis Pigmentosa. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1769–1778. [Google Scholar] [CrossRef]
- Dysli, C.; Wolf, S.; Hatz, K.; Zinkernagel, M.S. Fluorescence Lifetime Imaging in Stargardt Disease: Potential Marker for Disease Progression. Investig. Ophthalmol. Vis. Sci. 2016, 57, 832–841. [Google Scholar] [CrossRef]
- Sauer, L.; Gensure, R.H.; Hammer, M.; Bernstein, P.S. Fluorescence Lifetime Imaging Ophthalmoscopy: A Novel Way to Assess Macular Telangiectasia Type 2. Ophthalmol. Retin. 2018, 2, 587–598. [Google Scholar] [CrossRef]
- Schweitzer, D.; Deutsch, L.; Klemm, M.; Jentsch, S.; Hammer, M.; Peters, S.; Haueisen, J.; Müller, U.A.; Dawczynski, J. Fluorescence Lifetime Imaging Ophthalmoscopy in Type 2 Diabetic Patients Who Have No Signs of Diabetic Retinopathy. J. Biomed. Opt. 2015, 20, 061106. [Google Scholar] [CrossRef]
- Sadda, S.V.R.; Borrelli, E.; Fan, W.; Ebraheem, A.; Marion, K.M.; Harrington, M.; Kwon, S. A Pilot Study of Fluorescence Lifetime Imaging Ophthalmoscopy in Preclinical Alzheimer’s Disease. Eye 2019, 33, 1271–1279. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ||||||||
|---|---|---|---|---|---|---|---|---|
| 0 h After Irradiation | ||||||||
| Non-irradiated | 323 ± 23 | 243 ± 15 | 83.8 ± 0.9 | 487 ± 73 | 14.1 ± 0.6 | 2109 ± 87 | 2.1 ± 0.4 | 0.893 |
| Irradiated | 482 ± 10 | 368 ± 51 | 83.2 ± 0.4 | 817 ± 90 | 14.5 ± 0.2 | 2564 ± 95 | 2.3 ± 0.6 | 0.832 |
| 48 h After Irradiation | ||||||||
| Non-irradiated | 311 ± 17 | 224 ± 21 | 87.8 ± 1.0 | 538 ± 37 | 10.7 ± 0.9 | 2348 ± 84 | 1.5 ± 0.1 | 0.880 |
| Irradiated | 439 ± 16 | 253 ± 20 | 80.5 ± 2.1 | 554 ± 81 | 17.1 ± 1.9 | 2355 ± 145 | 3.3 ± 0.5 | 0.861 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semenov, A.N.; Maksimov, E.G.; Moysenovich, A.M.; Yakovleva, M.A.; Tsoraev, G.V.; Ramonova, A.A.; Shirshin, E.A.; Sluchanko, N.N.; Feldman, T.B.; Rubin, A.B.; et al. Protein-Mediated Carotenoid Delivery Suppresses the Photoinducible Oxidation of Lipofuscin in Retinal Pigment Epithelial Cells. Antioxidants 2023, 12, 413. https://doi.org/10.3390/antiox12020413
Semenov AN, Maksimov EG, Moysenovich AM, Yakovleva MA, Tsoraev GV, Ramonova AA, Shirshin EA, Sluchanko NN, Feldman TB, Rubin AB, et al. Protein-Mediated Carotenoid Delivery Suppresses the Photoinducible Oxidation of Lipofuscin in Retinal Pigment Epithelial Cells. Antioxidants. 2023; 12(2):413. https://doi.org/10.3390/antiox12020413
Chicago/Turabian StyleSemenov, Alexey N., Eugene G. Maksimov, Anastasia M. Moysenovich, Marina A. Yakovleva, Georgy V. Tsoraev, Alla A. Ramonova, Evgeny A. Shirshin, Nikolai N. Sluchanko, Tatiana B. Feldman, Andrew B. Rubin, and et al. 2023. "Protein-Mediated Carotenoid Delivery Suppresses the Photoinducible Oxidation of Lipofuscin in Retinal Pigment Epithelial Cells" Antioxidants 12, no. 2: 413. https://doi.org/10.3390/antiox12020413