
Comparative Study of Different H2S Donors as Vasodilators and Attenuators of Superoxide-Induced Endothelial Damage
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis
2.2. Stability and H2S Release
2.2.1. Stability of the Compounds at pH 7.4 in Phosphate Buffer (PBS) in the Absence and Presence of L-Cysteine
2.2.2. Stability of the Compounds in Human Serum
2.2.3. RP-HPLC Analysis of Stability Assays
2.2.4. Determination of H2S Release in Human Serum (Dansilazide Method)
2.3. Functional Experiments
2.3.1. Vasodilating Activity
2.3.2. Vasoprotection in Rat Aorta with Endothelium Impairment Induced by Pyrogallol
2.3.3. Data Analysis
3. Results
3.1. Stability and H2S Release
3.2. Functional Studies
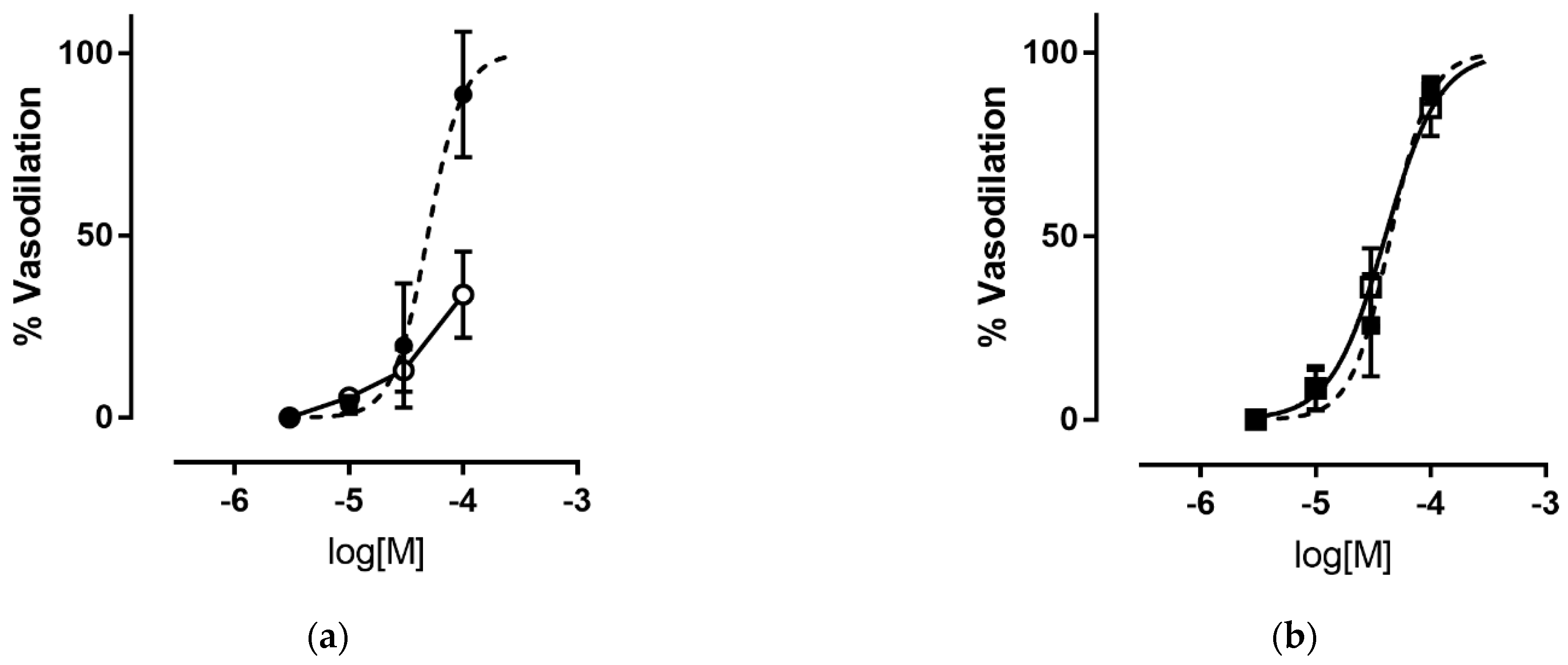
3.2.1. Vasodilating Activity
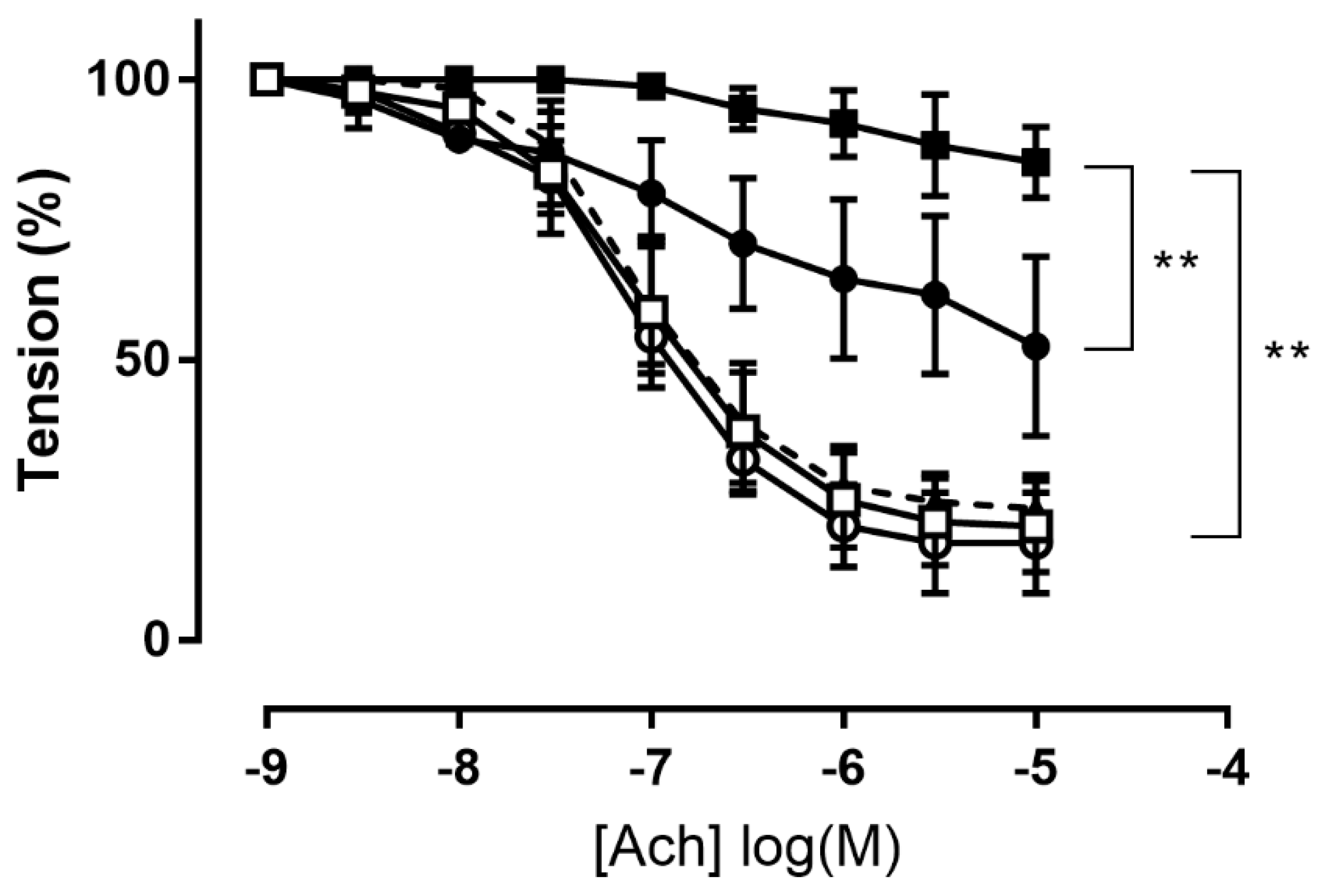
3.2.2. Effect of H2S Donor on Acetylcholine-Induced Vasodilation in Aorta Incubated with Pyrogallol
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, R. Hydrogen sulfide: The third gasotransmitter in biology and medicine. Antioxid. Redox Signal. 2010, 12, 1061–1064. [Google Scholar] [CrossRef]
- Wang, R. Two’s company, three’s a crowd: Can H2S be the third endogenous gaseous transmitter. FASEB J. 2002, 16, 1792–1798. [Google Scholar] [CrossRef]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef]
- Szabo, C. Gasotransmitters in cancer: From pathophysiology to experimental therapy. Nat. Rev. Drug Discov. 2016, 15, 185–203. [Google Scholar] [CrossRef]
- Szabo, C. A timeline of hydrogen sulfide (H2S) research: From environmental toxin to biological mediator. Biochem. Pharmacol. 2018, 149, 5–19. [Google Scholar] [CrossRef]
- Kolluru, G.K.; Shen, X.; Bir, S.C.; Kevil, C.G. Hydrogen sulfide chemical biology: Pathophysiological roles and detection. Nitric Oxide 2013, 35, 5–20. [Google Scholar] [CrossRef]
- Kolluru, G.K.; Shackelford, R.E.; Shen, X.; Dominic, P.; Kevil, C.G. Sulfide regulation of cardiovascular function in health and disease. Nat. Rev. Cardiol. 2023, 20, 109–125. [Google Scholar] [CrossRef]
- Liu, Y.H.; Lu, M.; Hu, L.F.; Wong, P.T.; Webb, G.D.; Bian, J.S. Hydrogen sulfide in the mammalian cardiovascular system. Antioxid. Redox Signal. 2012, 17, 141–185. [Google Scholar] [CrossRef]
- Pan, L.-L.; Qin, M.; Liu, X.-H.; Zhu, Y.-Z. The role of hydrogen sulfide on cardiovascular homeostasis: An overview with update on immunomodulation. Front. Pharmacol. 2017, 8, 686. [Google Scholar] [CrossRef]
- Szabo, C. Hydrogen sulfide, an enhancer of vascular nitric oxide signaling: Mechanisms and implications. Am. J. Physiol. Cell Physiol. 2017, 312, C3–C15. [Google Scholar] [CrossRef]
- Bir, S.C.; Kolluru, G.K.; McCarthy, P.; Shen, X.; Pardue, S.; Pattillo, C.B.; Kevil, C.G. Hydrogen sulfide stimulates ischemic vascular remodeling through nitric oxide synthase and nitrite reduction activity regulating hypoxiainducible factor-1α and vascular endothelial growth factor-dependent angiogenesis. J. Am. Heart Assoc. 2012, 1, e004093. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Polhemus, D.J.; Lefer, D.J. Evolution of hydrogen sulfide therapeutics to treat cardiovascular disease. Circ. Res. 2018, 123, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Rose, P.; Moore, P.K. Hydrogen sulfide and cell signaling. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 169–187. [Google Scholar] [CrossRef]
- Polhemus, D.J.; Lefer, D.J. Emergence of hydrogen sulfide as an endogenous gaseous signaling molecule in cardiovascular disease. Circ. Res. 2014, 114, 730–737. [Google Scholar] [CrossRef]
- Huang, S.; Li, H.; Ge, J. A Cardioprotective insight of the cystathionine-lyase/hydrogen sulfide pathway. IJC Heart Vasc. 2015, 7, 51–57. [Google Scholar] [CrossRef]
- Meng, G.; Ma, Y.; Xie, L.; Ferro, A.; Ji, Y. Emerging role of hydrogen sulfide in hypertension and related cardiovascular diseases. Br. J. Pharmacol. 2015, 172, 5501–5511. [Google Scholar] [CrossRef]
- Wang, X.-H.; Wang, F.; You, S.-J.; Cao, Y.-J.; Cao, L.-D.; Han, Q.; Liu, C.-F.; Hu, L.-F. Dysregulation of cystathionine-lyase (CSE)/hydrogen sulfide pathway contributes to ox-LDL-induced inflammation in macrophage. Cell Signal. 2013, 25, 2255–2262. [Google Scholar] [CrossRef]
- Whiteman, M.; Moore, P.K. Hydrogen sulfide and the vasculature: A novel vasculoprotective entity and regulator of nitric oxide bioavailability? J. Cell. Mol. Med. 2009, 13, 488–507. [Google Scholar] [CrossRef]
- Xu, Y.; Du, H.-P.; Li, J.; Xu, R.; Wang, Y.-L.; You, S.-J.; Liu, H.; Wang, F.; Cao, Y.-J.; Liu, C.-F.; et al. Statins upregulate cystathionine-lyase transcription and H2S generation via activating Akt signaling in macrophage. Pharmacol. Res. 2014, 87, 18–25. [Google Scholar] [CrossRef]
- Yu, X.-H.; Cui, L.-B.; Wu, K.; Zheng, X.-L.; Cayabyab, F.S.; Chen, Z.-W.; Tang, C.-K. Hydrogen sulfide as a potent cardiovascular protective agent. Clin. Chim. Acta 2014, 437, 78–87. [Google Scholar] [CrossRef]
- Zheng, Y.; Ji, X.; Ji, K.; Wang, B. Hydrogen sulfide prodrugs-A review. Acta Pharm. Sin. B 2015, 5, 367–377. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Li, Y.; Li, L.; Xu, S.; Feng, X.; Liu, S. Hydrogen sulfide (H2S)-releasing cCompounds: Therapeutic potential in cardiovascular diseases. Front. Pharmacol. 2018, 9, 1066. [Google Scholar] [CrossRef] [PubMed]
- Calvert, J.W.; Elston, M.; Nicholson, C.K.; Gundewar, S.; Jha, S.; Elrod, J.W.; Ramachandran, A.; Lefer, D.J. Genetic and pharmacologic hydrogen sulfide therapy attenuates ischemia-induced heart failure in mice. Circulation 2010, 122, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a Physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, X.; Jin, H.; Wei, H.; Li, W.; Bu, D.; Tang, X.; Ren, Y.; Tang, C.; Du, J. Role of hydrogen sulfide in the development of atherosclerotic lesions in apolipoprotein E knockout mice. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 173–179. [Google Scholar] [CrossRef]
- Van Den Born, J.C.; Mencke, R.; Conroy, S.; Zeebregts, C.J.; Van Goor, H.; Hillebrands, J.L. Cystathionine gamma-lyase is expressed in human atherosclerotic plaque microvessels and is involved in micro-angiogenesis. Sci. Rep. 2016, 6, 34608. [Google Scholar] [PubMed]
- Zhao, W.; Zhang, J.; Lu, Y.; Wang, R. The vasorelaxant effect of H(2)S as a novel endogenous gaseous K(ATP) channel opener. EMBO J. 2001, 20, 6008–6016. [Google Scholar] [CrossRef]
- Dawe, G.S.; Han, S.P.; Bian, J.S.; Moore, P.K. Hydrogen sulphide in the hypothalamus causes an ATP-sensitive K+ channel-dependent decrease in blood pressure in freely moving rats. Neuroscience 2008, 152, 169–177. [Google Scholar] [CrossRef]
- Liu, W.Q.; Chai, C.; Li, X.Y.; Yuan, W.J.; Wang, W.Z.; Lu, Y. The cardiovascular effects of central hydrogen sulfide are related to K(ATP) channels activation. Physiol. Res. 2011, 60, 729–738. [Google Scholar] [CrossRef]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Liu, Z.; Liu, P. Targeting hydrogen sulfide as a promising tTherapeutic strategy for atherosclerosis. Int. J. Cardiol. 2014, 172, 313–317. [Google Scholar] [CrossRef]
- Wen, Y.D.; Wang, H.; Kho, S.H.; Rinkiko, S.; Sheng, X.; Shen, H.M.; Zhu, Y.Z. Hydrogen sulfide protects HUVECs against hydrogen peroxide induced mitochondrial dysfunction and oxidative stress. PLoS ONE 2013, 8, e53147. [Google Scholar] [CrossRef]
- Corvino, A.; Frecentese, F.; Magli, E.; Perissutti, E.; Santagada, V.; Scognamiglio, A.; Caliendo, G.; Fiorino, F.; Severino, B. Trends in H2S-donors chemistry and their effects in cardiovascular diseases. Antioxidants 2021, 10, 429. [Google Scholar] [CrossRef]
- Hughes, M.N.; Centelles, M.N.; Moore, K.P. Making and working with hydrogen sulfide. Free Radic. Biol. Med. 2009, 47, 1346–1353. [Google Scholar] [CrossRef]
- Lazzarato, L.; Chegaev, K.; Marini, E.; Rolando, B.; Borretto, E.; Guglielmo, S.; Joseph, S.; Di Stilo, A.; Fruttero, R.; Gasco, A. New nitric oxide or hydrogen sulfide releasing aspirins. J. Med. Chem. 2011, 54, 5478–5484. [Google Scholar] [CrossRef]
- Brusotti, G.; Papetti, A.; Serra, M.; Temporini, C.; Marini, E.; Orlandini, S.; Sanda, A.K.; Watcho, P.; Kamtchouing, P. Allanblackia floribunda Oliv.: An aphrodisiac plant with vasorelaxant properties. J. Ethnopharmacol. 2016, 192, 480–485. [Google Scholar] [CrossRef]
- Jin, B.H.; Qian, L.B.; Chen, S.; Li, J.; Wang, H.P.; Bruce, I.C.; Lin, J.; Xia, Q. Apigenin protects endothelium-dependent relaxation of rat aorta against oxidative stress. Eur. J. Pharmacol. 2009, 616, 200–205. [Google Scholar] [CrossRef]
- Liu, S.; Xing, J.; Zheng, Z.; Song, F.; Liu, Z.; Liu, S. Ultrahigh performance liquid chromatography-triple quadrupole mass spectrometry inhibitors fishing assay: A novel method for simultaneously screening of xanthine oxidase inhibitor and superoxide anion scavenger in a single analysis. Anal. Chim. Acta 2012, 715, 64–70. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. Preventing developmental origins of cardiovascular disease: Hydrogen sulfide as a potential target? Antioxidants 2021, 10, 247. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.; Xian, M. Cysteine-activated hydrogen sulfide (H2S) donors. J. Am. Chem. Soc. 2011, 133, 15–17. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Bhushan, S.; Yang, C.; Otsuka, H.; Stein, J.D.; Pacheco, A.; Peng, B.; Devarie-Baez, N.O.; Aguilar, H.C.; Lefer, D.J.; et al. Controllable hydrogen sulfide donors and their activity against myocardial ischemia-reperfusion injury. ACS Chem. Biol. 2013, 8, 1283–1290. [Google Scholar] [CrossRef]
- Roger, T.; Raynaud, F.; Bouillaud, F.; Ransy, C.; Simonet, S.; Crespo, C.; Bourguignon, M.-P.; Villeneuve, N.; Vilaine, J.-P.; Artaud, I.; et al. New biologically active hydrogen sulfide donors. ChemBioChem 2013, 14, 2268–2271. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Pugliesi, I.; Barresi, E.; Nesi, G.; Rapposelli, S.; Taliani, S.; Da Settimo, F.; et al. Arylthioamides as H2S donors: L-cysteine-activated releasing properties and vascular effects in vitro and in vivo. ACS Med. Chem. Lett. 2013, 4, 904–908. [Google Scholar] [CrossRef]
- Severino, B.; Corvino, A.; Fiorino, F.; Luciano, P.; Frecentese, F.; Magli, E.; Saccone, I.; Di Vaio, P.; Citi, V.; Calderone, V.; et al. 1,2,4-Thiadiazolidin-3,5-diones as novel hydrogen sulfide donors. Eur. J. Med. Chem. 2018, 143, 1677–1686. [Google Scholar] [CrossRef]
- Barresi, E.; Nesi, G.; Citi, V.; Piragine, E.; Piano, I.; Taliani, S.; Da Settimo, F.; Rapposelli, S.; Testai, L.; Breschi, M.C.; et al. Iminothioethers as hydrogen sulfide donors: From the gasotransmitter release to the vascular effects. J. Med. Chem. 2017, 60, 7512–7523. [Google Scholar] [CrossRef]
- Mitidieri, E.; Tramontano, T.; Gurgone, D.; Citi, V.; Calderone, V.; Brancaleone, V.; Katsouda, A.; Nagahara, N.; Papapetropoulos, A.; Cirino, G.; et al. Mercaptopyruvate acts as endogenous vasodilator independently of 3-mercaptopyruvate sulfurtransferase activity. Nitric Oxide 2018, 75, 53–59. [Google Scholar] [CrossRef]
- Ercolano, G.; De Cicco, P.; Frecentese, F.; Saccone, I.; Corvino, A.; Giordano, F.; Magli, E.; Fiorino, F.; Severino, B.; Calderone, V.; et al. Anti-metastatic properties of naproxen-HBTA in a murine model of cutaneous melanoma. Front. Pharmacol. 2019, 10, 66. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Bellagambi, F.G.; Ghimenti, S.; Breschi, M.C.; Calderone, V. Pharmacological characterization of the vascular effects of aryl isothiocyanates: Is hydrogen sulfide the real player? Vasc. Pharmacol. 2014, 60, 32–41. [Google Scholar] [CrossRef]
- Citi, V.; Corvino, A.; Fiorino, F.; Frecentese, F.; Magli, E.; Perissutti, E.; Santagada, V.; Brogi, S.; Flori, L.; Gorica, E.; et al. Structure-activity relationships study of isothiocyanates for H2S releasing properties: 3-pyridyl-isothiocyanate as a new promising cardioprotective agent. J. Adv. Res. 2021, 27, 41–53. [Google Scholar] [CrossRef]
- Zhao, Y.; Steiger, A.K.; Pluth, M.D. Cyclic sulfenyl thiocarbamates release carbonyl culfide and hydrogen sulfide independently in thiol-promoted pathways. J. Am. Chem. Soc. 2019, 141, 13610–13618. [Google Scholar] [CrossRef]
- Kang, M.; Hashimoto, A.; Gade, A.; Akbarali, H.I. Interaction between hydrogen sulfide-induced sulfhydration and tyrosine nitration in the KATP channel complex. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G532–G539. [Google Scholar] [CrossRef]
- Bucci, M.; Papapetropoulos, A.; Vellecco, V.; Zhou, Z.; Pyriochou, A.; Roussos, C.; Roviezzo, F.; Brancaleone, V.; Cirino, G. Hydrogen sulfide is an endogenous inhibitor of phosphodiesterase activity. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1998–2004. [Google Scholar] [CrossRef]
- Kiss, L.; Deitch, E.A.; Szabó, C. Hydrogen sulfide decreases adenosine triphosphate levels in aortic rings and leads to vasorelaxation via metabolic inhibition. Life Sci. 2008, 83, 589–594. [Google Scholar] [CrossRef]
- Lee, S.W.; Cheng, Y.; Moore, P.K.; Bian, J.S. Hydrogen sulphide regulates intracellular pH in vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 2007, 358, 1142–1147. [Google Scholar] [CrossRef]
- Liu, Y.H.; Bian, J.S. Bicarbonate-dependent effect of hydrogen sulfide on vascular contractility in rat aortic rings. Am. J. Physiol. Cell Physiol. 2010, 299, C866–C872. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Breschi, M.C.; Lawson, K.; McKay, N.G.; Miceli, F.; Taglialatela, M.; Calderone, V. Vasorelaxation by hydrogen sulphide involves activation of Kv7 potassium channels. Pharmacol. Res. 2013, 70, 27–34. [Google Scholar] [CrossRef]
- Bucci, M.; Papapetropoulos, A.; Vellecco, V.; Zhou, Z.; Zaid, A.; Giannogonas, P.; Cantalupo, A.; Dhayade, S.; Karalis, K.P.; Wang, R.; et al. cGMP-dependent protein kinase contributes to hydrogen sulfide-stimulated vasorelaxation. PLoS ONE 2012, 7, e53319. [Google Scholar] [CrossRef]
- Sun, H.J.; Wu, Z.Y.; Nie, X.W.; Bian, J.S. Role of endothelial dysfunction in cardiovascular diseases: The link between inflammation and hydrogen sulfide. Front. Pharmacol. 2020, 10, 1568. [Google Scholar] [CrossRef]
- Xie, L.; Feng, H.; Li, S.; Meng, G.; Liu, S.; Tang, X.; Ma, Y.; Han, Y.; Xiao, Y.; Gu, Y.; et al. SIRT3 mediates the antioxidant effect of hydrogen sulfide in endothelial cells. Antioxid. Redox Signal. 2016, 24, 329–343. [Google Scholar] [CrossRef]
- Kimura, Y.; Kimura, H. Hydrogen sulfide protects neurons from oxidative stress. FASEB J. 2004, 18, 1165–1167. [Google Scholar] [CrossRef]
- Tyagi, N.; Moshal, K.S.; Sen, U.; Vacek, T.P.; Kumar, M.; Hughes, W.M., Jr.; Kundu, S.; Tyagi, S.C. H2S protects against methionine-induced oxidative stress in brain endothelial cells. Antioxid. Redox Signal. 2009, 11, 25–33. [Google Scholar] [CrossRef]
- Zhang, L.M.; Jiang, C.X.; Liu, D.W. Hydrogen sulfide attenuates neuronal injury induced by vascular dementia via inhibiting apoptosis in rats. Neurochem. Res. 2009, 34, 1984–1992. [Google Scholar] [CrossRef]
- Szabó, G.; Veres, G.; Radovits, T.; Ger, D.; Módis, K.; Miesel-Gröschel, C.; Horkay, F.; Karck, M.; Szabó, C. Cardioprotective effects of hydrogen sulfide. Nitric Oxide 2011, 25, 201–210. [Google Scholar] [CrossRef]
- Mistry, R.K.; Brewer, A.C. Redox regulation of gasotransmission in the vascular system: A focus on angiogenesis. Free Radic. Biol. Med. 2017, 108, 500–516. [Google Scholar] [CrossRef]
- Fukuto, J.M.; Carrington, S.J.; Tantillo, D.J.; Harrison, J.G.; Ignarro, L.J.; Freeman, B.A.; Chen, A.; Wink, D.A. Small molecule signaling agents: The integrated chemistry and biochemistry of nitrogen oxides, oxides of carbon, dioxygen, hydrogen sulfide, and their derived species. Chem. Res. Toxicol. 2012, 25, 769–793. [Google Scholar] [CrossRef]
- Ono, K.; Akaike, T.; Sawa, T.; Kumagai, Y.; Wink, D.A.; Tantillo, D.J.; Hobbs, A.J.; Nagy, P.; Xian, M.; Lin, J.; et al. Redox chemistry and chemical biology of H2S, hydropersulfides, and derived species: Implications of their possible biological activity and utility. Free Radic. Biol. Med. 2014, 77, 82–94. [Google Scholar] [CrossRef]
- Gryglewski, R.J.; Palmer, R.M.; Moncada, S. Superoxide anion is involved in the breakdown of endothelium-derived vascular relaxing factor. Nature 1986, 320, 454–456. [Google Scholar] [CrossRef]
- MacKenzie, A.; Martin, W. Loss of endothelium-derived nitric oxide in rabbit aorta by oxidant stress: Restoration by superoxide dismutase mimetics. Br. J. Pharmacol. 1998, 124, 719–728. [Google Scholar] [CrossRef]
- Misak, A.; Grman, M.; Bacova, Z.; Rezuchova, I.; Hudecova, S.; Ondriasova, E.; Krizanova, O.; Brezova, V.; Chovanec, M.; Ondrias, K. Polysulfides and products of H2S/S-nitrosoglutathione in comparison to H2S, glutathione and antioxidant Trolox are potent scavengers of superoxide anion radical and produce hydroxyl radical by decomposition of H2O2. Nitric Oxide 2018, 76, 136–151. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, C.; Organ, C.; Li, Z.; Bhushan, S.; Otsuka, H.; Pacheco, A.; Kang, J.; Aguilar, H.C.; Lefer, D.J.; et al. Design, Synthesis, and cardioprotective effects of N-mercapto-based hydrogen sulfide donors. J. Med. Chem. 2015, 58, 7501–7511. [Google Scholar] [CrossRef]
- Dickinson, D.A.; Forman, H.J. Glutathione in defense and signaling: Lessons from a small thiol. Ann. N. Y. Acad. Sci. 2002, 973, 488–504. [Google Scholar] [CrossRef] [PubMed]
- Izigov, N.; Farzam, N.; Savion, N. S-allylmercapto-N-acetylcysteine up-regulates cellular glutathione and protects vascular endothelial cells from oxidative stress. Free Radic. Biol. Med. 2011, 50, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Chegaev, K.; Rolando, B.; Cortese, D.; Gazzano, E.; Buondonno, I.; Lazzarato, L.; Fanelli, M.; Hattinger, C.M.; Serra, M.; Riganti, C.; et al. H2S-Donating doxorubicins may overcome cardiotoxicity and multidrug resistance. J. Med. Chem. 2016, 59, 4881–4889. [Google Scholar] [CrossRef]
- Lee, M.; Tazzari, V.; Giustarini, D.; Rossi, R.; Sparatore, A.; Del Soldato, P.; McGeer, E.; McGeer, P.L. Effects of hydrogen sulfide-releasing L-DOPA derivatives on glial activation. J. Biol. Chem. 2010, 23, 17318–17328. [Google Scholar] [CrossRef]
- Dartigues, B.; Cambar, J.; Trebaul, C.; Brelivet, J.; Guglielmetti, R. Proprietes diuretiques de derives des dithiole-thiones. Recherche d’une relation structure-activite. Eur. J. Med. Chem. 1980, 15, 405–412. [Google Scholar]
- Terauchi, T.; Kobayashi, Y.; Misaki, Y. Synthesis of bis-fused tetrathiafulvalene with mono- and dicarboxylic acids. Tetrahedron Lett. 2012, 53, 3277–3280. [Google Scholar] [CrossRef]




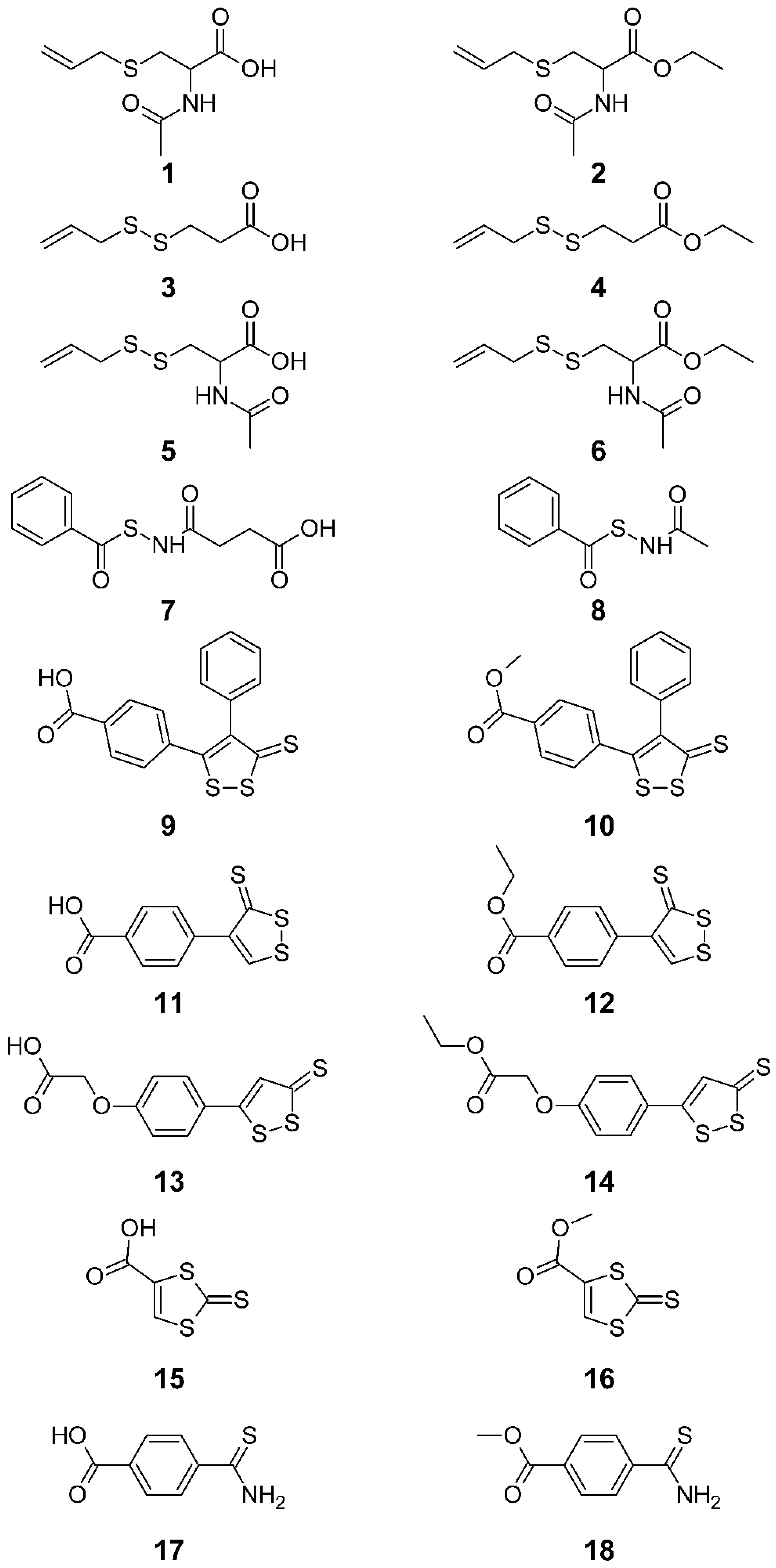{kind=link}
{kind=link}
{kind=link}
| Compd 1 | Stability and H2S Release In Human Serum | Vasodilating Activity | ||||
|---|---|---|---|---|---|---|
| % Compd (at 1, 4, 24 h) ± SE or Half-Life | % H2S mol/mol (at 1, 4, 24 h) Hours ± SE | EC50 (μM) ± SEM [% Vasodilation] | + Glib. 10 μM EC50 (μM) ± SEM [% Vasodilation] | |||
| 1 | 88 ± 1 | 1 h | 0.4 ± 0.1 | 1 h | NA | NA |
| 35 ± 1 | 4 h | 1.5 ± 0.1 | 4 h | |||
| 8.0 ± 0.5 | 24 h | 3.9 ± 0.9 | 24 h | |||
| 2 | t1/2 < 30 min | 0.6 ± 0.3 | 1 h | NA | NA | |
| 1.8 ± 0.3 | 4 h | |||||
| 5 ± 1 | 24 h | |||||
| 3 | 80.0 ± 1.0 | 1 h | 1.5 ± 0.2 | 1 h | NA | NA |
| 62 ± 2 | 4 h | 8.2 ± 0.3 | 4 h | |||
| 44 ± 1 | 24 h | 23 ± 2 | 24 h | |||
| 4 | t1/2 < 30 min | 5.8 ± 0.2 | 1 h | 64 ± 6 | [44 ± 3] 2 | |
| 10 ± 1 | 4 h | |||||
| 24.2 ± 0.9 | 24 h | |||||
| 5 | 64 ± 1 | 1 h | 2.7 ± 0.2 | 1 h | [16 ± 4] 3 | |
| 39 ± 1 | 4 h | 11 ± 2 | 4 h | |||
| 16 ± 1 | 24 h | 21 ± 1 | 24 h | |||
| 6 | t1/2 < 30 min | 5.8 ± 0.2 | 1 h | 47 ± 8 | [34 ± 7] 2 | |
| 10 ± 1 | 4 h | |||||
| 24.2 ± 0.8 | 24 h | |||||
| 7 | 19.8 ± 0.7 | 1 h | 0.3 ± 0.2 | 1 h | [12 ± 1] 3 | |
| 0 | 4 h | 1.4 ± 0.2 | 4 h | |||
| 0 | 24 h | 5 ± 1 | 24 h | |||
| 8 | t1/2 < 30 min | 16 ± 1 | 1 h | 40 ± 3 | 40 ± 4 | |
| 25.8 ± 0.4 | 4 h | |||||
| 47 ± 1 | 24 h | |||||
| 9 | 94.2 ± 0.2 | 1 h | 0.4 ± 0.1 | 1 h | 78 ± 7 | 262 ± 62 |
| 90.0 ± 0.8 | 4 h | 1.7 ± 0.6 | 4 h | |||
| 83 ± 1 | 24 h | 4.4 ± 0.9 | 24 h | |||
| 10 | t1/2 = 2.4 h | 1.8 ± 0.3 | 1 h | [29 ± 3] 4 | ||
| 7.5 ± 0.6 | 4 h | |||||
| 12.0 ± 0.9 | 24 h | |||||
| 11 | 96.3 ± 0.2 | 1 h | 0.4 ± 0.4 | 1 h | [42 ± 3] 2 | |
| 90.0 ± 0.9 | 4 h | 4 ± 1 | 4 h | |||
| 53 ± 2 | 24 h | 33 ± 4 | 24 h | |||
| 12 | t1/2 = 2.7 h | 17 ± 8 | 1 h | 20 ± 2 | 20 ± 3 | |
| 63 ± 30 | 4 h | |||||
| 146 ± 34 | 24 h | |||||
| 13 | 99.2 ± 0.1 | 1 h | 0 | 1 h | [39 ± 8] 2 | |
| 96.0 ± 0.8 | 4 h | 0.7 ± 0.2 | 4 h | |||
| 94.3 ± 2 | 24 h | 2.4 ± 0.5 | 24 h | |||
| 14 | t1/2 = 30 min | 0 | 1 h | [37 ± 3] 4 | ||
| 0.5 ± 0.3 | 4 h | |||||
| 1.8 ± 0.6 | 24 h | |||||
| 15 | 99.2 ± 0.1 | 1 h | 0.2 ± 0.4 | 1 h | [17 ± 3] 3 | |
| 95 ± 1 | 4 h | 1.3 ± 0.2 | 4 h | |||
| 94.8 ± 2 | 24 h | 2.8 ± 0.5 | 24 h | |||
| 16 | t1/2 = 1.5 h | 0.4 ± 0.2 | 1 h | [25 ± 5] 2 | ||
| 1.7 ± 0.5 | 4 h | |||||
| 2.5 ± 0.9 | 24 h | |||||
| 17 | 91.6 ± 0.3 | 1 h | 0.2 ± 0.2 | 1 h | [27 ± 6] 3 | |
| 85.0 ± 0.1 | 4 h | 1.0 ± 0.3 | 4 h | |||
| 64.7 ± 0.2 | 24 h | 6 ± 1 | 24 h | |||
| 18 | t1/2 = 3.7 h | 1.4 ± 0.4 | 1 h | [33 ± 6] 4 | ||
| 11.0 ± 0.3 | 4 h | |||||
| 44 ± 2 | 24 h | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marini, E.; Rolando, B.; Sodano, F.; Blua, F.; Concina, G.; Guglielmo, S.; Lazzarato, L.; Chegaev, K. Comparative Study of Different H2S Donors as Vasodilators and Attenuators of Superoxide-Induced Endothelial Damage. Antioxidants 2023, 12, 344. https://doi.org/10.3390/antiox12020344
Marini E, Rolando B, Sodano F, Blua F, Concina G, Guglielmo S, Lazzarato L, Chegaev K. Comparative Study of Different H2S Donors as Vasodilators and Attenuators of Superoxide-Induced Endothelial Damage. Antioxidants. 2023; 12(2):344. https://doi.org/10.3390/antiox12020344
Chicago/Turabian StyleMarini, Elisabetta, Barbara Rolando, Federica Sodano, Federica Blua, Giulia Concina, Stefano Guglielmo, Loretta Lazzarato, and Konstantin Chegaev. 2023. "Comparative Study of Different H2S Donors as Vasodilators and Attenuators of Superoxide-Induced Endothelial Damage" Antioxidants 12, no. 2: 344. https://doi.org/10.3390/antiox12020344