
Fermented Glutinous Rice Extract Mitigates DSS-Induced Ulcerative Colitis by Alleviating Intestinal Barrier Function and Improving Gut Microbiota and Inflammation
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. FGR Preparation
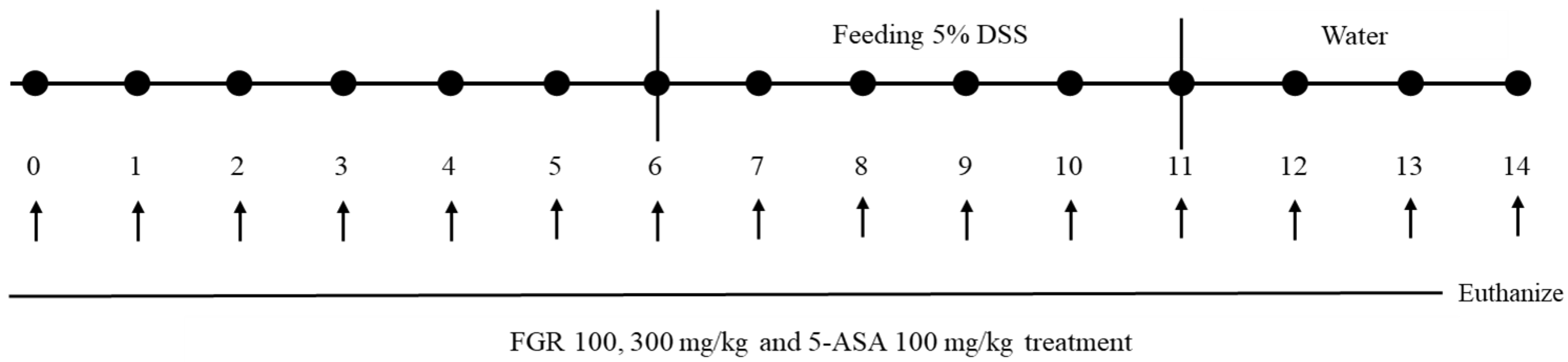
2.3. Animal Studies
2.4. Epithelial Paracellular Permeability
2.5. Large Intestine Endoscopy and Histological Analysis
2.6. ELISA for Myeloperoxidase (MPO) Activity and Levels of IL-6, TNF-α and IL-1β
2.7. ELISA for Malondialdehyde (MDA) Content, Catalase Activity, Superoxide Dismutase (SOD) Activity, and Glutathione (GSH) Contents
2.8. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL) Assay
2.9. Real-Time PCR for Colon Tissue
2.10. Western Blot Analysis
2.11. Biochemical Assays
2.12. Intestinal Microbiome Analysis (Metagenomics)
2.13. qRT-PCR Analysis of Beneficial and Harmful Bacteria
2.14. Statistical Analysis
3. Results
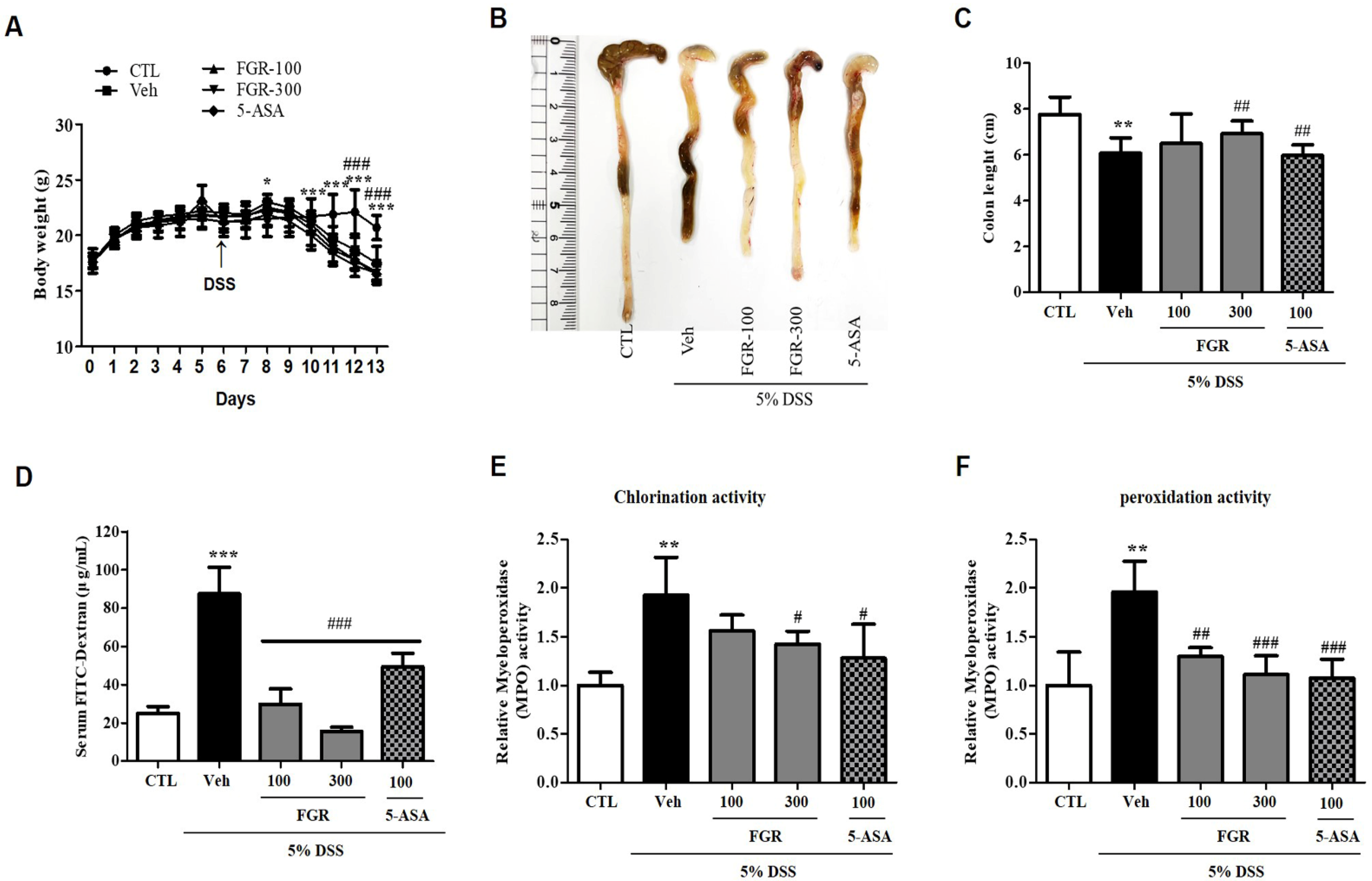
3.1. Effects of FGR on DSS-Induced Body Weight Loss, Colon Length, Permeability, and MPO Activity
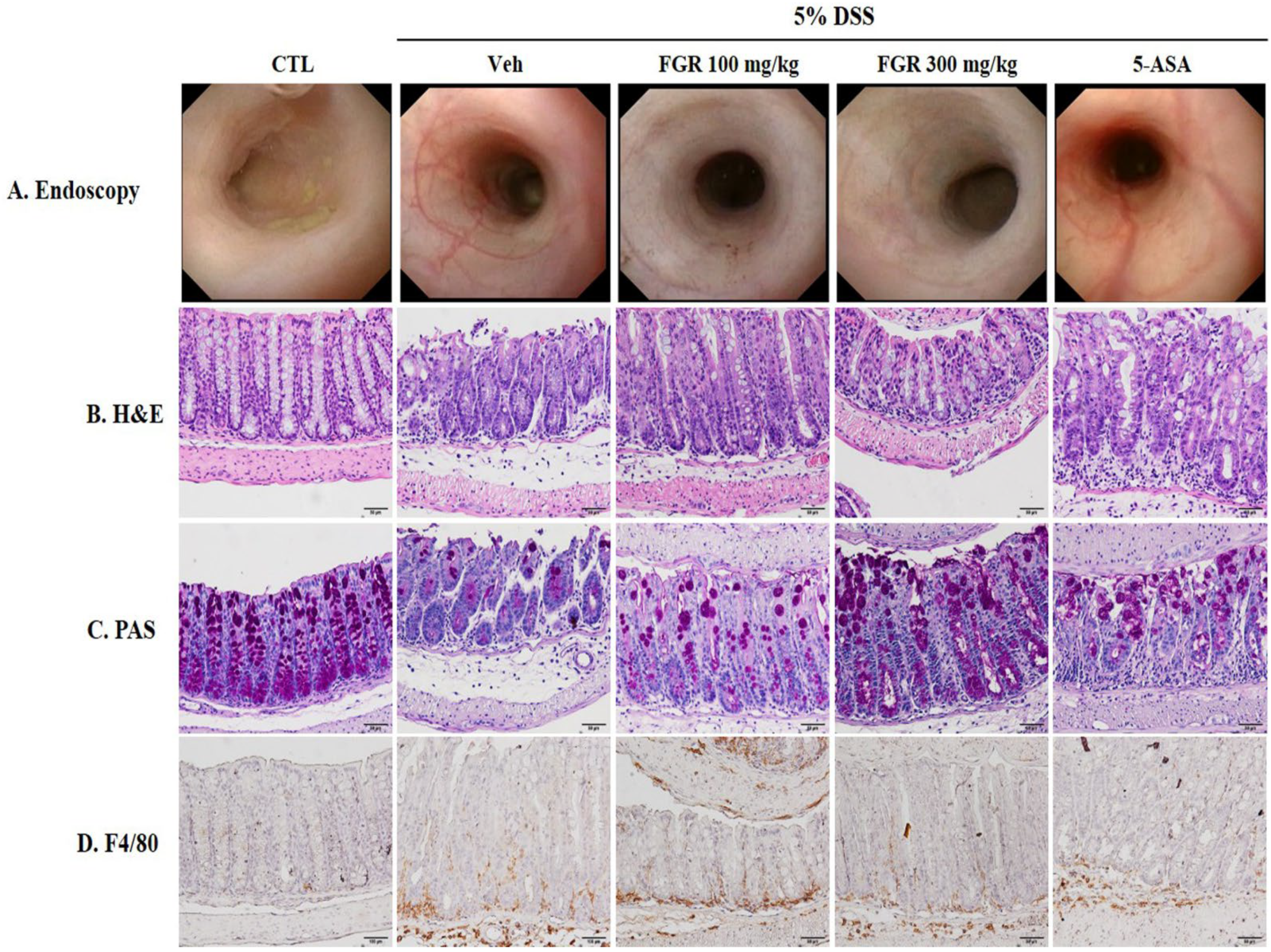
3.2. Effects of FGR on DSS-Induced Histological and Morphological Changes of the Large Intestine
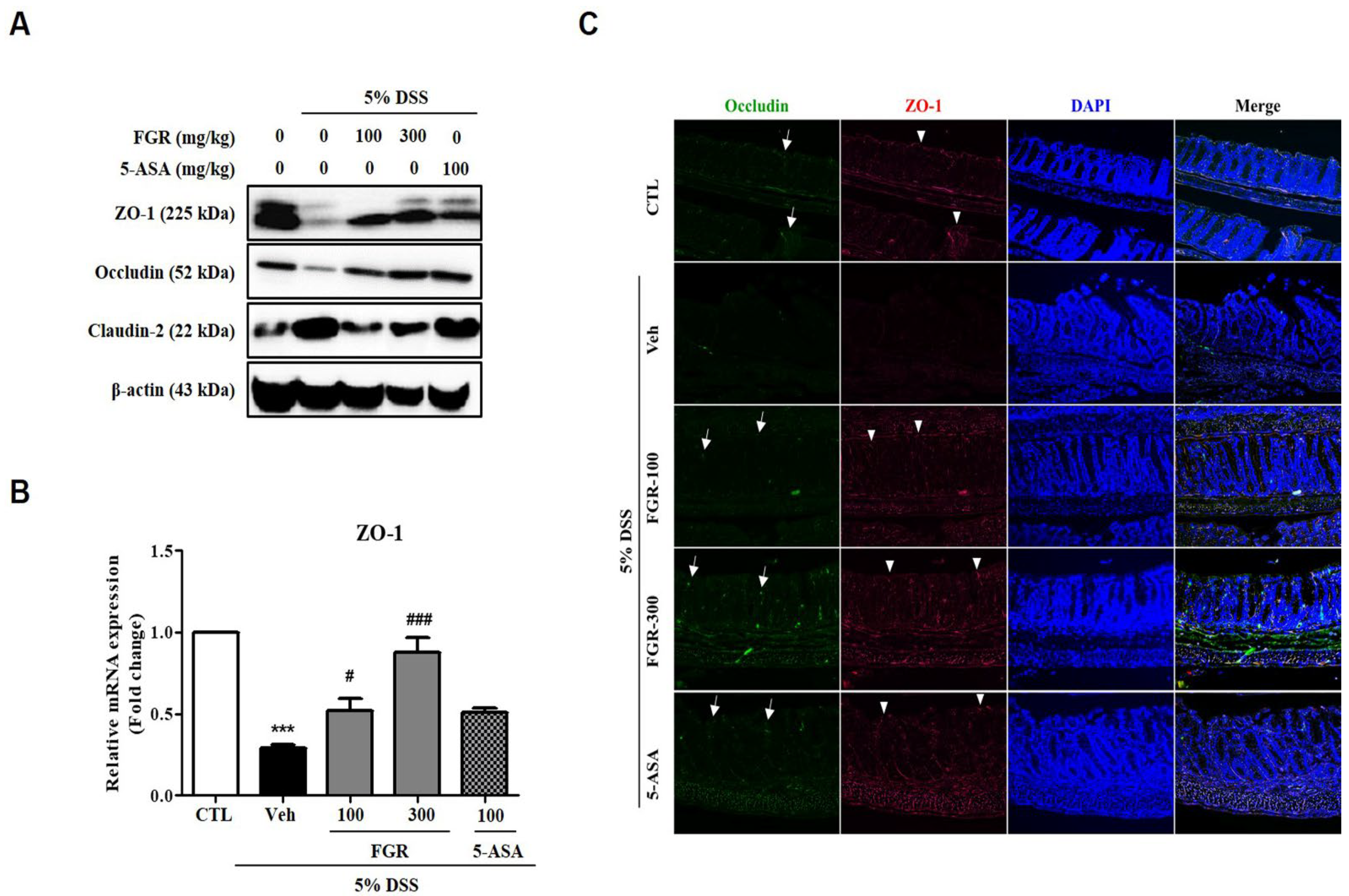
3.3. Effects of FGR on Intestinal Barrier Function in Mice with DSS-Induced Colitis
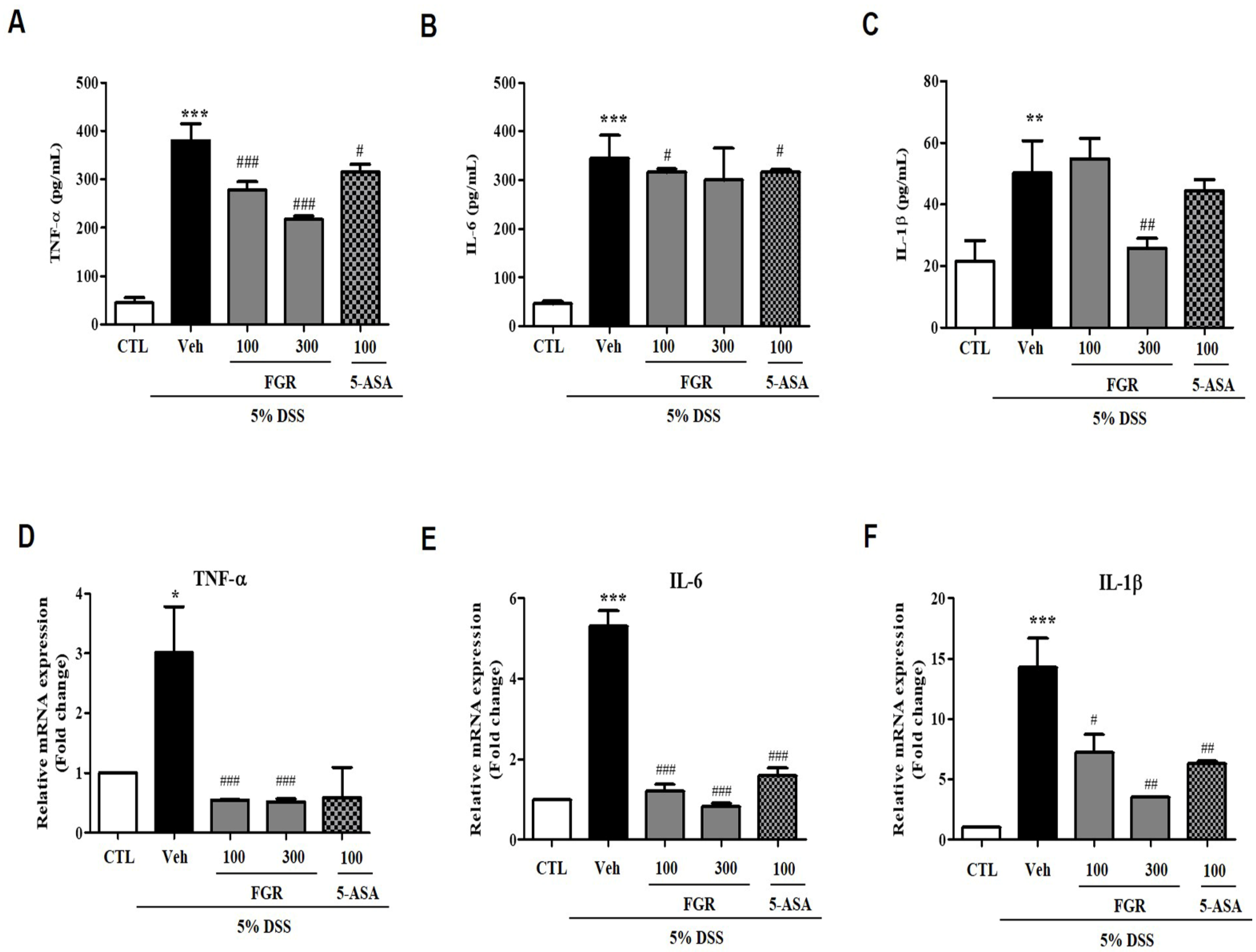
3.4. Effects of FGR on Serum and mRNA Levels of Proinflammatory Cytokines in Mice with DSS-Induced Colitis
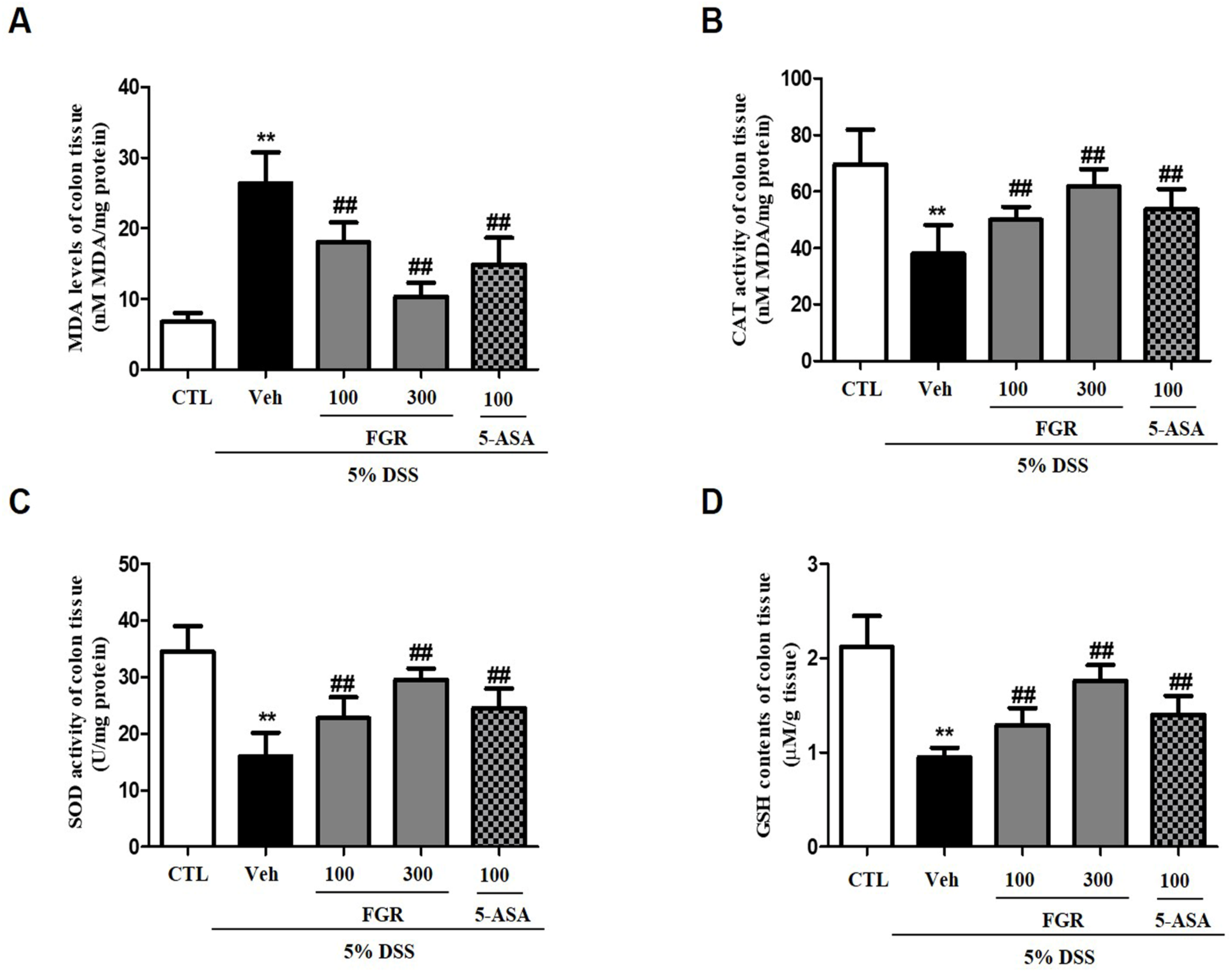
3.5. Effects of FGR on Oxidative Stress in Mice with DSS-Induced Colitis
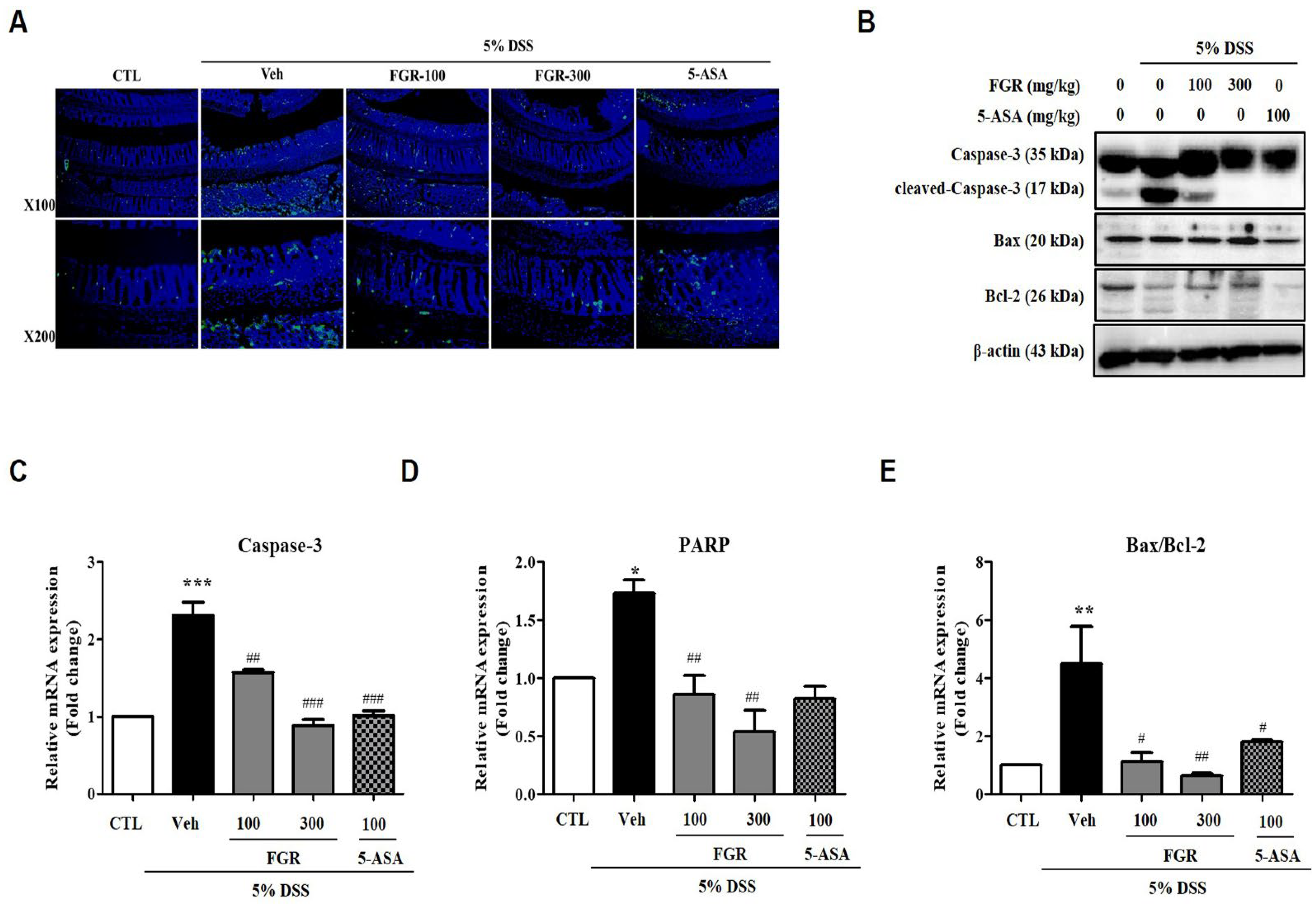
3.6. Effect of FGR on Apoptosis in Mice with DSS-Induced Colitis
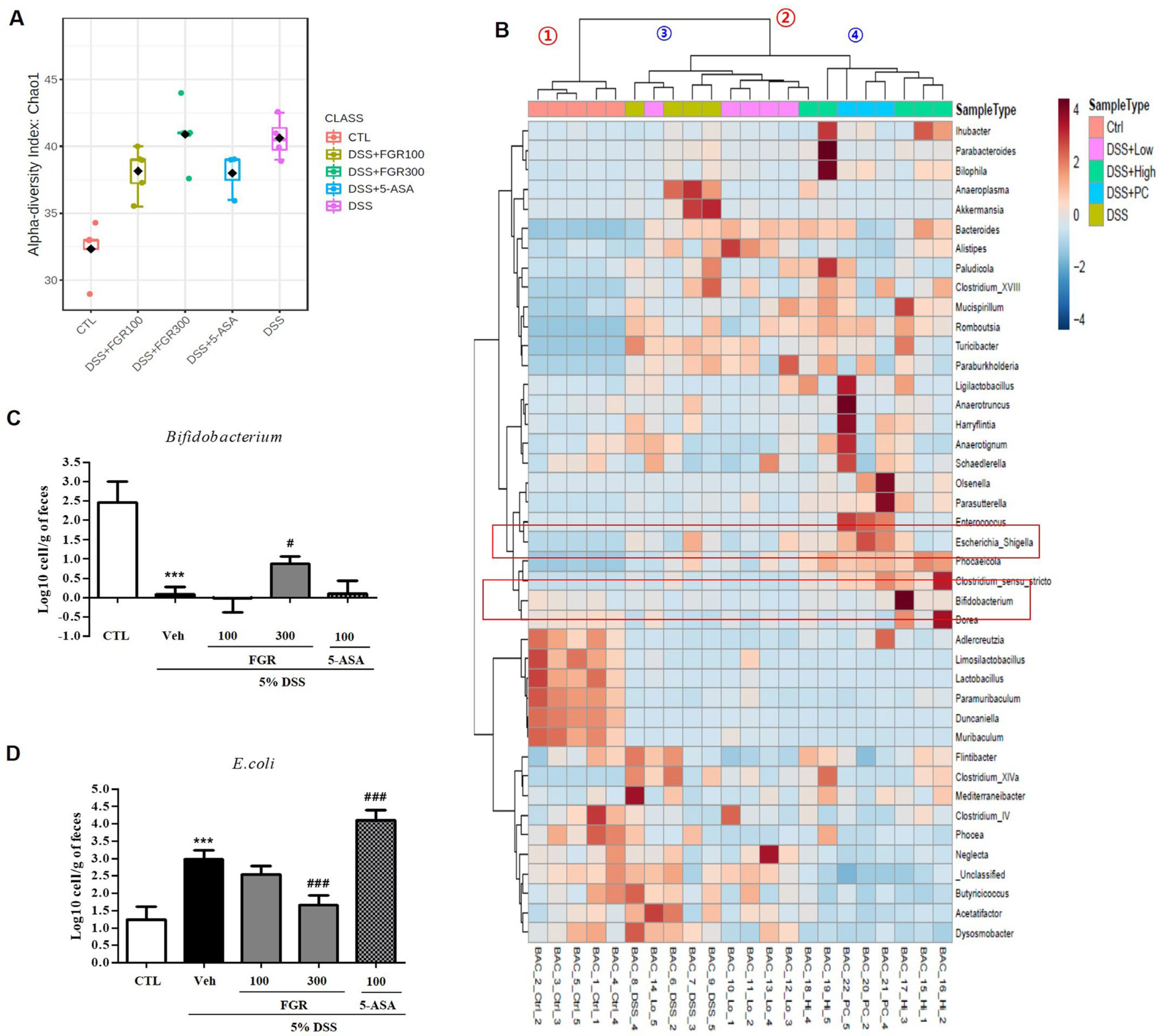
3.7. Effect of FGR on Intestinal Microbial Composition in Mice with DSS-Induced Colitis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morrison, G.; Headon, B.; Gibson, P. Update in inflammatory bowel disease. Aust. Fam. Physician 2009, 38, 956–961. [Google Scholar] [PubMed]
- Rubin, D.C.; Shaker, A.; Levin, M.S. Chronic intestinal inflammation: Inflammatory bowel disease and colitis-associated colon cancer. Front. Immunol. 2012, 3, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, S.; Shivaji, U.N.; Kasturi, R.; Sigamani, A.; Ghosh, S.; Iacucci, M. Inflammatory bowel disease-related colorectal cancer: Past, present and future perspectives. World J. Gastrointest. Oncol. 2022, 14, 547–567. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Strober, W.; Fuss, I.; Mannon, P. The fundamental basis of inflammatory bowel disease. J. Clin. Investig. 2007, 117, 514–521. [Google Scholar] [CrossRef] [Green Version]
- Hanauer, S.B. Inflammatory bowel disease: Epidemiology, pathogenesis, and therapeutic opportunities. Inflamm. Bowel Dis. 2006, 12 (Suppl. 1), S3–S9. [Google Scholar] [CrossRef]
- Clapper, M.L.; Gary, M.A.; Coudry, R.A.; Litwin, S.; Chang, W.C.; Devarajan, K.; Lubet, R.A.; Cooper, H.S. 5-aminosalicylic acid inhibits colitis-associated colorectal dysplasias in the mouse model of azoxymethane/dextran sulfate sodium-induced colitis. Inflamm. Bowel Dis. 2008, 14, 1341–1347. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Madsen, K.; Doyle, J.; Meddings, J. Reducing small intestinal permeability attenuates colitis in the IL10 gene-deficient mouse. Gut 2009, 58, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Edelblum, K.L.; Turner, J.R. The tight junction in inflammatory disease: Communication breakdown. Curr. Opin. Pharmacol. 2009, 9, 715–720. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.Y.; Oh, T.W.; Do, H.J.; Yang, J.H.; Yang, I.J.; Jeon, Y.H.; Go, Y.H.; Ahn, S.C.; Ma, J.Y.; Park, K.I. Acer palmatum thumb. Ethanol Extract Alleviates Interleukin-6-Induced Barrier Dysfunction and Dextran Sodium Sulfate-Induced Colitis by Improving Intestinal Barrier Function and Reducing Inflammation. J. Immunol. Res. 2018, 2018, 5718396. [Google Scholar] [CrossRef]
- Hamazaki, Y.; Itoh, M.; Sasaki, H.; Furuse, M.; Tsukita, S. Multi-PDZ domain protein 1 (MUPP1) is concentrated at tight junctions through its possible interaction with claudin-1 and junctional adhesion molecule. J. Biol. Chem. 2002, 277, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Furuse, M. Molecular basis of the core structure of tight junctions. Cold Spring Harb. Perspect. Biol. 2010, 2, a002907. [Google Scholar] [CrossRef] [Green Version]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Daniel, F.; Seksik, P.; Cacheux, W.; Jian, R.; Marteau, P. Tolerance of 4-aminosalicylic acid enemas in patients with inflammatory bowel disease and 5-aminosalicylic-induced acute pancreatitis. Inflamm. Bowel Dis. 2004, 10, 258–260. [Google Scholar] [CrossRef]
- Yan, Y.; Kolachala, V.; Dalmasso, G.; Nguyen, H.; Laroui, H.; Sitaraman, S.V.; Merlin, D. Temporal and spatial analysis of clinical and molecular parameters in dextran sodium sulfate induced colitis. PLoS ONE 2009, 4, e6073. [Google Scholar] [CrossRef]
- Egger, B.; Bajaj-Elliott, M.; MacDonald, T.T.; Inglin, R.; Eysselein, V.E.; Buchler, M.W. Characterisation of acute murine dextran sodium sulphate colitis: Cytokine profile and dose dependency. Digestion 2000, 62, 240–248. [Google Scholar] [CrossRef]
- Garside, P. Cytokines in experimental colitis. Clin. Exp. Immunol. 1999, 118, 337–339. [Google Scholar] [CrossRef]
- Strober, W.; Fuss, I.J. Proinflammatory Cytokines in the Pathogenesis of Inflammatory Bowel Diseases. Gastroenterology 2011, 140, 1756–1767. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.R.; Noh, E.M.; Lee, S.H.; Lee, S.; Kim, D.H.; Lee, N.H.; Kim, S.Y.; Park, M.H. Momordica charantia extracts obtained by ultrasound-assisted extraction inhibit the inflammatory pathways. Mol. Cell. Toxicol. 2022. [Google Scholar] [CrossRef]
- Nishitani, Y.; Zhang, L.; Yoshida, M.; Azuma, T.; Kanazawa, K.; Hashimoto, T.; Mizuno, M. Intestinal Anti-Inflammatory Activity of Lentinan: Influence on IL-8 and TNFR1 Expression in Intestinal Epithelial Cells. PLoS ONE 2013, 8, e62441. [Google Scholar] [CrossRef]
- Katsanos, K.H.; Papadakis, K.A. Inflammatory Bowel Disease: Updates on Molecular Targets for Biologics. Gut Liver 2017, 11, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, H.; Neurath, M.F.; Atreya, R. Role of the IL23/IL17 Pathway in Crohn’s Disease. Front. Immunol. 2021, 12, 622934. [Google Scholar] [CrossRef]
- Head, K.A.; Jurenka, J.S. Inflammatory bowel disease Part 1: Ulcerative colitis--pathophysiology and conventional and alternative treatment options. Altern. Med. Rev. 2003, 8, 247–283. [Google Scholar]
- McLean, M.H.; Neurath, M.F.; Durum, S.K. Targeting interleukins for the treatment of inflammatory bowel disease-what lies beyond anti-TNF therapy? Inflamm. Bowel Dis. 2014, 20, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.X.; Ren, M.T.; Yang, J.P.; Pan, H.H.; Yu, M.S.; Ji, F. Curcumin Improves Epithelial Barrier Integrity of Caco-2 Monolayers by Inhibiting Endoplasmic Reticulum Stress and Subsequent Apoptosis. Gastroenterol. Res. Pract. 2021, 2021, 5570796. [Google Scholar] [CrossRef]
- Fu, Y.P.; Yuan, H.; Xu, Y.; Liu, R.M.; Luo, Y.; Xiao, J.H. Protective effects of Ligularia fischeri root extracts against ulcerative colitis in mice through activation of Bcl-2/Bax signalings. Phytomedicine 2022, 99, 154006. [Google Scholar] [CrossRef]
- Shawki, A.; McCole, D.F. Mechanisms of Intestinal Epithelial Barrier Dysfunction by Adherent-Invasive Escherichia coli. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.C.H. Microbiota dysbiosis and barrier dysfunction in inflammatory bowel disease and colorectal cancers: Exploring a common ground hypothesis. J. Biomed. Sci. 2018, 25, 79. [Google Scholar] [CrossRef] [Green Version]
- Stevceva, L.; Pavli, P.; Husband, A.J.; Doe, W.F. The inflammatory infiltrate in the acute stage of the dextran sulphate sodium induced colitis: B cell response differs depending on the percentage of DSS used to induce it. BMC Clin. Pathol. 2001, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Abdelbaqi, M.; Chidlow, J.H.; Matthews, K.M.; Pavlick, K.P.; Barlow, S.C.; Linscott, A.J.; Grisham, M.B.; Fowler, M.R.; Kevil, C.G. Regulation of dextran sodium sulfate induced colitis by leukocyte beta 2 integrins. Lab. Investig. 2006, 86, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Klimstra, W.B.; Ryman, K.D.; Bernard, K.A.; Nguyen, K.B.; Biron, C.A.; Johnston, R.E. Infection of neonatal mice with sindbis virus results in a systemic inflammatory response syndrome. J. Virol. 1999, 73, 10387–10398. [Google Scholar] [CrossRef]
- Yin, A.; Luo, Y.; Chen, W.; He, M.; Deng, J.H.; Zhao, N.; Cao, L.; Wang, L. FAM96A Protects Mice From Dextran Sulfate Sodium (DSS)-Induced Colitis by Preventing Microbial Dysbiosis. Front. Cell. Infect. Microbiol. 2019, 9, 381. [Google Scholar] [CrossRef] [Green Version]
- Wan, S.; Song, G.; Hu, H.; Xu, Y.; Zeng, P.; Lin, S.; Yang, J.; Jiang, J.; Song, X.; Luo, Y.; et al. Intestine epithelial cell-derived extracellular vesicles alleviate inflammation induced by Clostridioides difficile TcdB through the activity of TGF-beta1. Mol. Cell. Toxicol. 2022, 1–11. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Primer Sequence | Annealing Temperature (°C) | Gene Information | |
|---|---|---|---|---|
| Beneficial bacteria | Lactobacillus acidophilus | Fwd: AGCAGTAGGGAATCTTCCA | 52 | NCBI Genome UID: 1099 |
| Rev: CACCGCTACACATGGAG | ||||
| Bifidobacterium breve | Fwd: CTCCTGGAAACGGGTGG | 52 | NCBI Genome UID: 1273 | |
| Rev: GGTGTTCTTCCCGATATCTACA | ||||
| Harmful bacteria | E. coli | Fwd: GTTAATACCTTTGCTCATTGA | 52 | NCBI Genome UID: 167 |
| Rev: ACCAGGGTATCTAATCCTGTT | ||||
| Clostridium perfringens | Fwd: GGGGGTTTCAACACCTCC | 48 | NCBI Genome UID: 158 | |
| Rev: GCAAGGGATGTCAAGTGT | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.-Y.; Son, J.D.; Hwang, S.-J.; Lee, J.K.; Park, J.Y.; Park, K.I.; Oh, T.W. Fermented Glutinous Rice Extract Mitigates DSS-Induced Ulcerative Colitis by Alleviating Intestinal Barrier Function and Improving Gut Microbiota and Inflammation. Antioxidants 2023, 12, 336. https://doi.org/10.3390/antiox12020336
Kim K-Y, Son JD, Hwang S-J, Lee JK, Park JY, Park KI, Oh TW. Fermented Glutinous Rice Extract Mitigates DSS-Induced Ulcerative Colitis by Alleviating Intestinal Barrier Function and Improving Gut Microbiota and Inflammation. Antioxidants. 2023; 12(2):336. https://doi.org/10.3390/antiox12020336
Chicago/Turabian StyleKim, Kwang-Youn, Jae Dong Son, Su-Jin Hwang, Jong Kwang Lee, Jae Young Park, Kwang Il Park, and Tae Woo Oh. 2023. "Fermented Glutinous Rice Extract Mitigates DSS-Induced Ulcerative Colitis by Alleviating Intestinal Barrier Function and Improving Gut Microbiota and Inflammation" Antioxidants 12, no. 2: 336. https://doi.org/10.3390/antiox12020336