
The Pleiotropic Regulator AdpA Regulates the Removal of Excessive Sulfane Sulfur in Streptomyces coelicolor

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Growth Conditions
2.2. S. coelicolor ΔadpA, ΔadpA::adpA, and Trx-Overexpression Strains
2.3. Quantitative Determination of H2S and RSS Content
2.4. RNA Preparation and RT-qPCR
2.5. AdpA Expression and Purification
2.6. Electrophoretic Mobility Shift Assays (EMSA)
3. Results
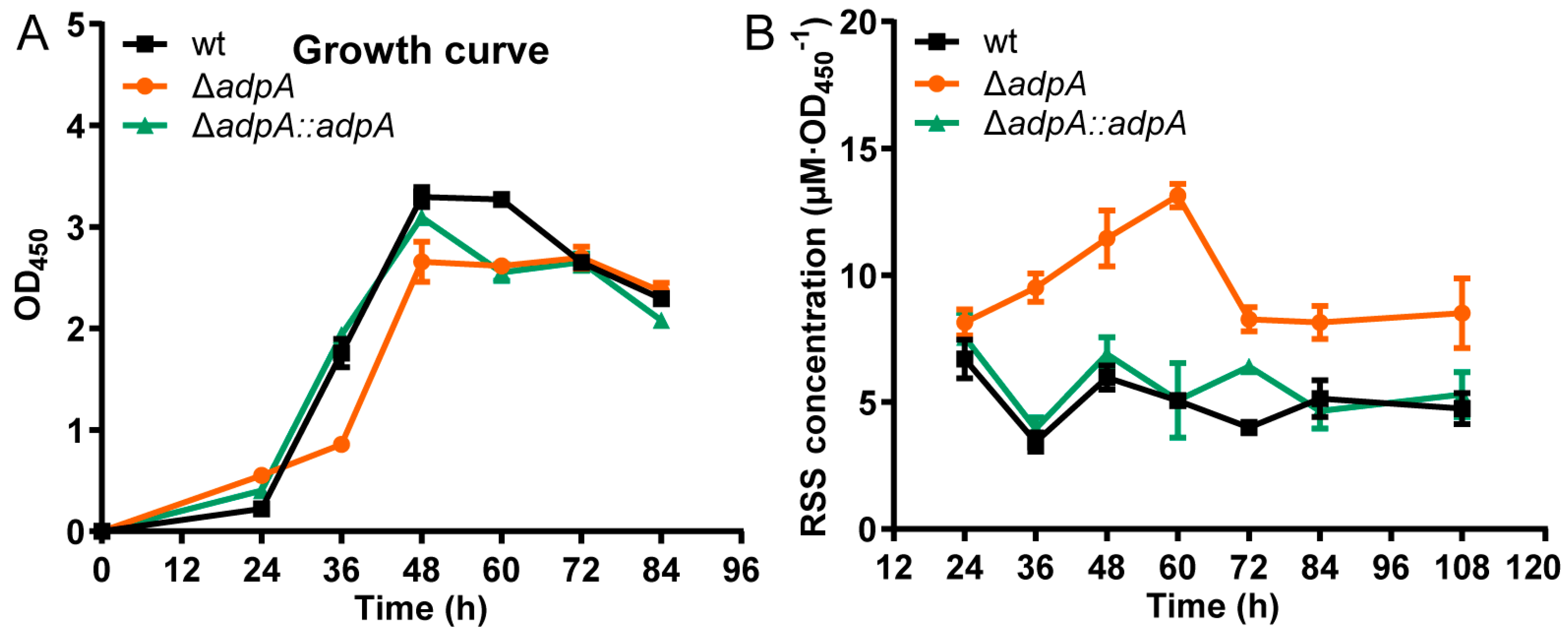
3.1. Deleting AdpA Increased the Sensitivity of S. coelicolor M145 to RSS
3.2. Deleting AdpA Led to RSS Accumulation in S. coelicolor M145 Cells
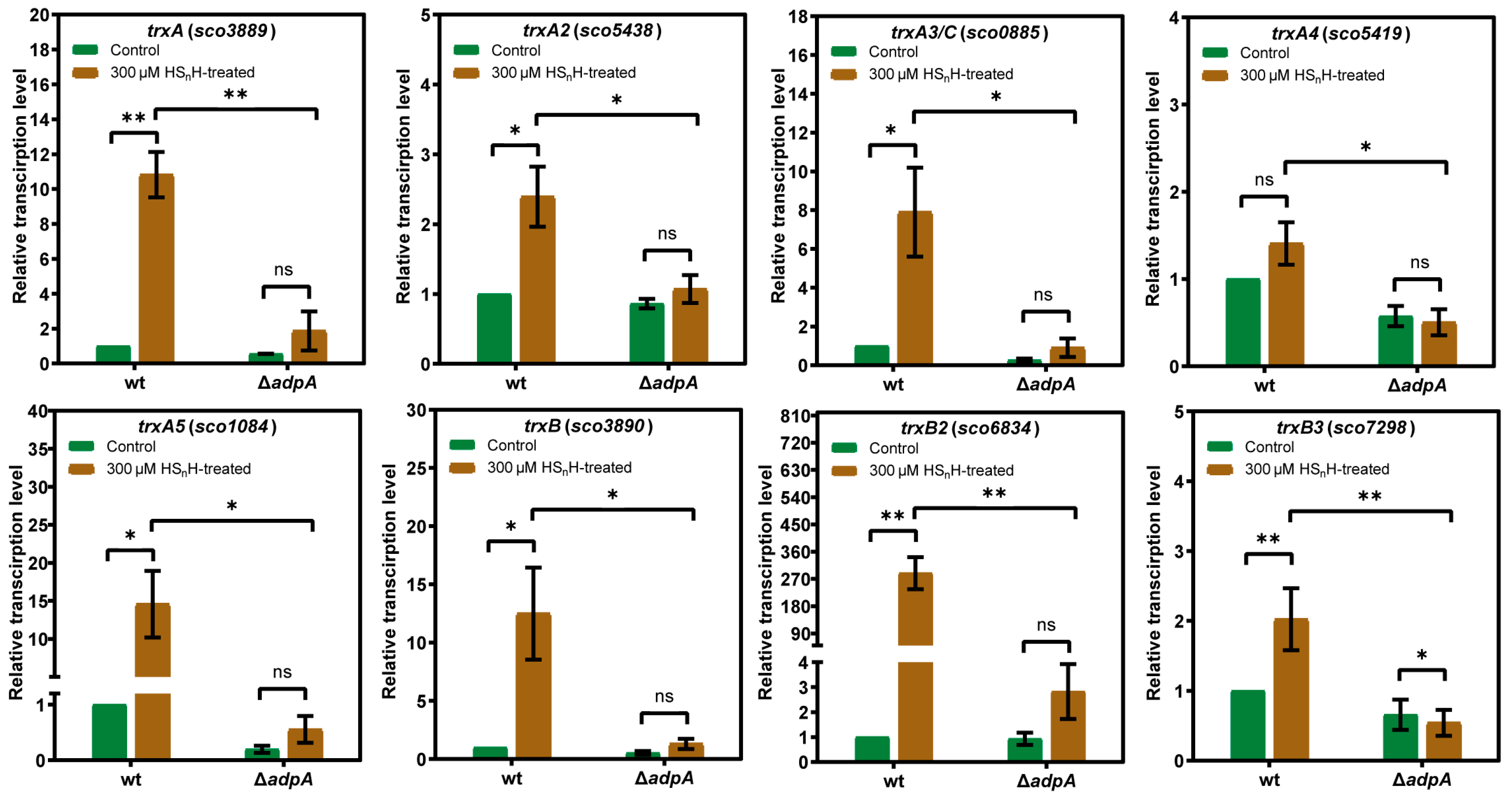
3.3. AdpA-Dependent Expression of Thioredoxins Was Induced by RSS
3.4. Overexpressing Thioredoxins Promoted H2S Release
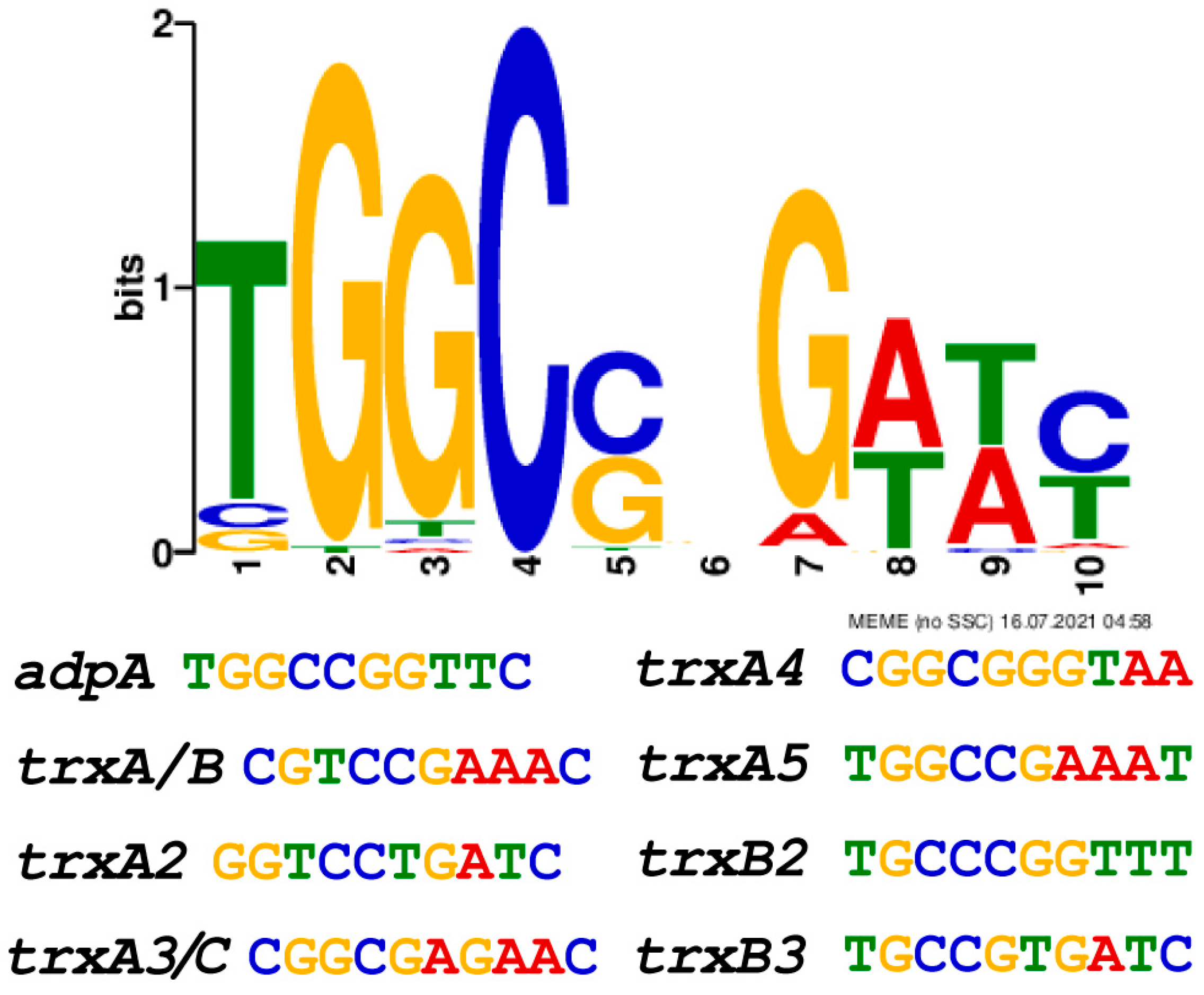
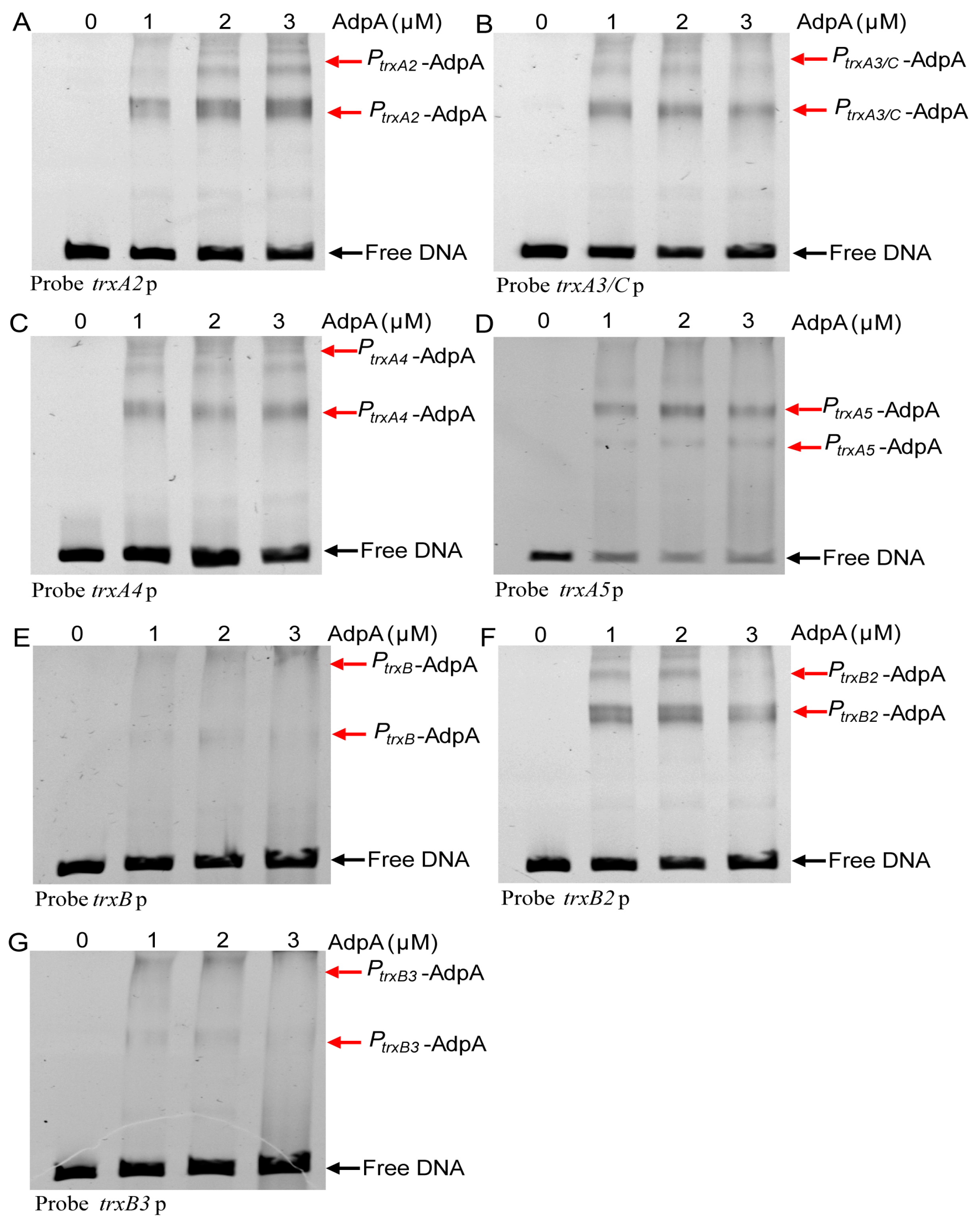
3.5. AdpA Directly Regulated the Transcription of Trx Genes
4. Discussion
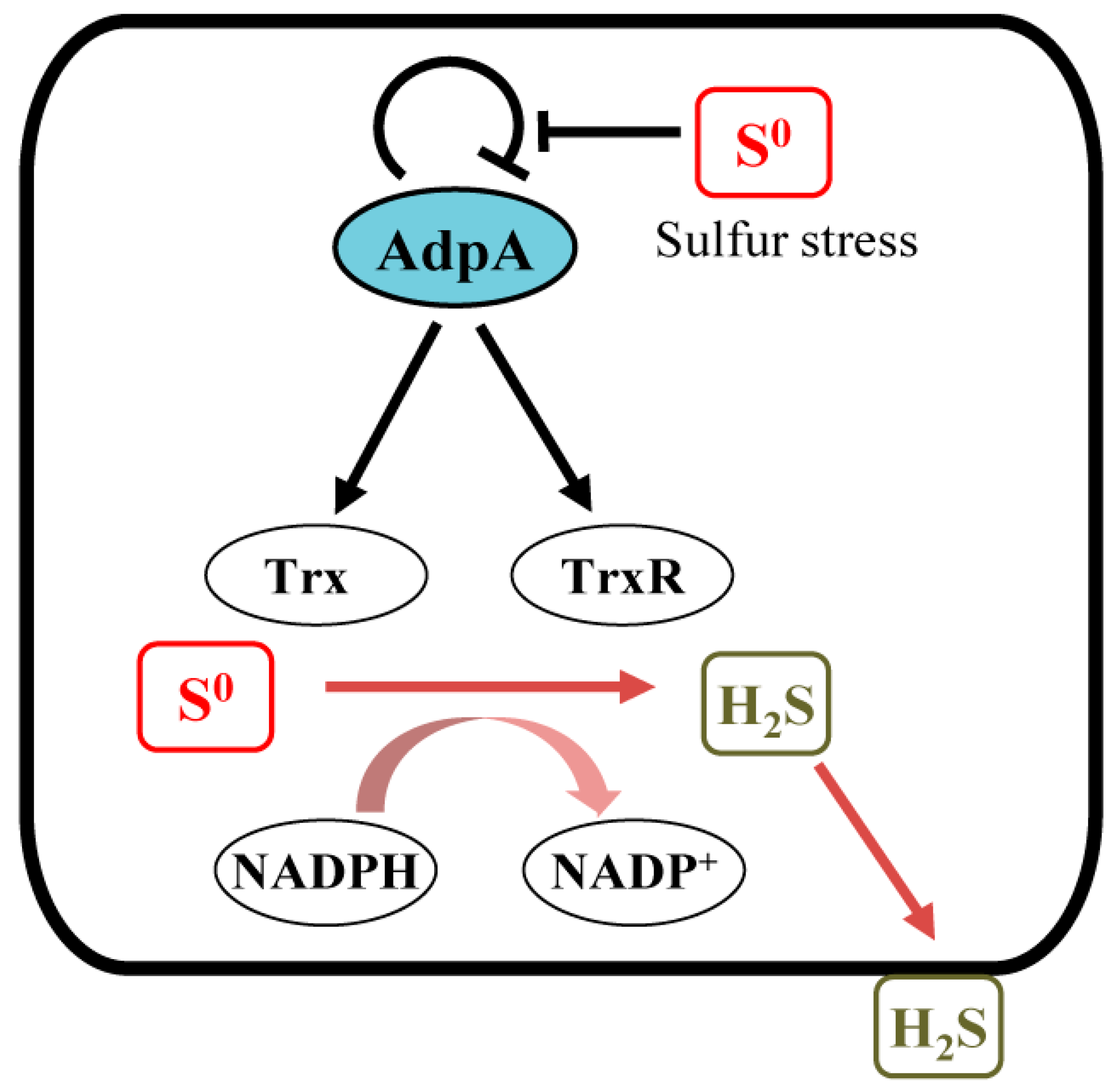
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [Green Version]
- Hine, C.; Harputlugil, E.; Zhang, Y.; Ruckenstuhl, C.; Lee, B.C.; Brace, L.; Longchamp, A.; Trevino-Villarreal, J.H.; Mejia, P.; Ozaki, C.K.; et al. Endogenous hydrogen sulfide production is essential for dietary restriction benefits. Cell 2015, 160, 132–144. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.H.; Krokowski, D.; Guan, B.J.; Bederman, I.; Majumder, M.; Parisien, M.; Diatchenko, L.; Kabil, O.; Willard, B.; Banerjee, R.; et al. Quantitative H2S-mediated protein sulfhydration reveals metabolic reprogramming during the integrated stress response. eLife 2015, 4, e10067. [Google Scholar] [CrossRef]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.W.; Camacho, M.I.; Chen, Y.; Bhat, A.H.; Chang, C.; Peluso, E.A.; Wu, C.; Das, A.; Ton-That, H. Genetic determinants of hydrogen sulfide biosynthesis in fusobacterium nucleatum are required for bacterial fitness, antibiotic sensitivity, and virulence. mBio 2022, 13, e0193622. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.R.X.; Chen, Y.; Ng, L.F.; Gruber, J.; Gan, Y.H. Glutathione catabolism by Enterobacteriaceae species to hydrogen sulphide adversely affects the viability of host systems in the presence of 5’fluorodeoxyuridine. Mol. Microbiol. 2022, 117, 1089–1103. [Google Scholar] [CrossRef]
- Pietri, R.; Roman-Morales, E.; Lopez-Garriga, J. Hydrogen sulfide and hemeproteins: Knowledge and mysteries. Antioxid. Redox Signal. 2011, 15, 393–404. [Google Scholar] [CrossRef] [Green Version]
- Paul, B.D.; Snyder, S.H. H2S signalling through protein sulfhydration and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 499–507. [Google Scholar] [CrossRef]
- Toohey, J.I. Sulfur signaling: Is the agent sulfide or sulfane? Anal. Biochem. 2011, 413, 1–7. [Google Scholar] [CrossRef]
- Walsh, B.J.C.; Giedroc, D.P. H2S and reactive sulfur signaling at the host-bacterial pathogen interface. J. Biol. Chem. 2020, 295, 13150–13168. [Google Scholar] [CrossRef]
- Kimura, H. Hydrogen sulfide and polysulfide signaling. Antioxid. Redox Signal. 2017, 27, 619–621. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Z.; Zhang, X.; Xin, Y.; Xia, Y.; Xun, L.; Liu, H. Rhodanese Rdl2 produces reactive sulfur species to protect mitochondria from reactive oxygen species. Free Radic. Biol. Med. 2021, 177, 287–298. [Google Scholar] [CrossRef]
- Barayeu, U.; Schilling, D.; Eid, M.; Xavier da Silva, T.N.; Schlicker, L.; Mitreska, N.; Zapp, C.; Grater, F.; Miller, A.K.; Kappl, R.; et al. Hydropersulfides inhibit lipid peroxidation and ferroptosis by scavenging radicals. Nat. Chem. Biol. 2023, 19, 28–37. [Google Scholar] [CrossRef]
- Zhang, X.; Xin, Y.; Chen, Z.; Xia, Y.; Xun, L.; Liu, H. Sulfide-quinone oxidoreductase is required for cysteine synthesis and indispensable to mitochondrial health. Redox Biol. 2021, 47, 102169. [Google Scholar] [CrossRef]
- Alvarez, L.; Bianco, C.L.; Toscano, J.P.; Lin, J.; Akaike, T.; Fukuto, J.M. Chemical biology of hydropersulfides and related species: Possible roles in cellular protection and redox signaling. Antioxid. Redox Signal. 2017, 27, 622–633. [Google Scholar] [CrossRef]
- Kumagai, Y.; Akiyama, M.; Unoki, T. Adaptive responses to electrophilic stress and reactive sulfur species as their regulator molecules. Toxicol. Res. 2019, 35, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.S.; Cooper, R.M. Elemental sulphur is produced by diverse plant families as a component of defence against fungal and bacterial pathogens. Physiol. Mol. Plant Pathol. 2003, 63, 3–16. [Google Scholar] [CrossRef]
- Leslie, K.S.; Millington, G.W.; Levell, N.J. Sulphur and skin: From Satan to Saddam! J. Cosmet. Dermatol. 2004, 3, 94–98. [Google Scholar] [CrossRef]
- Wang, T.; Yang, Y.; Liu, M.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. Elemental sulfur inhibits yeast growth via producing toxic sulfide and causing disulfide stress. Antioxidants 2022, 11, 576. [Google Scholar] [CrossRef]
- Shen, J.C.; Keithly, M.E.; Armstrong, R.N.; Higgins, K.A.; Edmonds, K.A.; Giedroc, D.P. Staphylococcus aureus CstB is a novel multidomain persulfide dioxygenase-sulfurtransferase involved in hydrogen sulfide detoxification. Biochemistry 2015, 54, 4542–4554. [Google Scholar] [CrossRef]
- Giedroc, D.P. A new player in bacterial sulfide-inducible transcriptional regulation. Mol. Microbiol. 2017, 105, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.P.; Clement, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekurova, O.N.; Zehl, M.; Predl, M.; Hunyadi, P.; Rattei, T.; Zotchev, S.B. Targeted metabolomics and high-throughput RNA sequencing-based transcriptomics reveal massive changes in the Streptomyces venezuelae NRRL B-65442 metabolism caused by ethanol shock. Microbiol. Spectr. 2022, 10, e0367222. [Google Scholar] [CrossRef]
- van der Meij, A.; Worsley, S.F.; Hutchings, M.I.; van Wezel, G.P. Chemical ecology of antibiotic production by actinomycetes. FEMS Microbiol. Rev. 2017, 41, 392–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherdson, E.M.F.; Elliot, M.A. Cryptic specialized metabolites drive Streptomyces exploration and provide a competitive advantage during growth with other microbes. Proc. Natl. Acad. Sci. USA 2022, 119, e2211052119. [Google Scholar] [CrossRef] [PubMed]
- Rabyk, M.; Yushchuk, O.; Rokytskyy, I.; Anisimova, M.; Ostash, B. Genomic insights into evolution of AdpA family master regulators of morphological differentiation and secondary metabolism in Streptomyces. J. Mol. Evol. 2018, 86, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Tistechok, S.; Stierhof, M.; Kachor, A.; Myronovskyi, M.; Gromyko, O.; Luzhetskyy, A. Flavacol and its novel derivative 3-beta-hydroxy flavacol from Streptomyces sp. Pv 4-95 after the expression of heterologous AdpA. Microorganisms 2022, 10, 2335. [Google Scholar] [CrossRef]
- Huang, R.; Liu, H.; Zhao, W.; Wang, S.; Wang, S.; Cai, J.; Yang, C. AdpA, a developmental regulator, promotes epsilon-poly-L-lysine biosynthesis in Streptomyces albulus. Microb. Cell Fact. 2022, 21, 60. [Google Scholar] [CrossRef] [PubMed]
- Plachetka, M.; Krawiec, M.; Zakrzewska-Czerwinska, J.; Wolanski, M. AdpA positively regulates morphological differentiation and chloramphenicol biosynthesis in Streptomyces venezuelae. Microbiol. Spectr. 2021, 9, e0198121. [Google Scholar] [CrossRef]
- Yushchuk, O.; Ostash, I.; Vlasiuk, I.; Gren, T.; Luzhetskyy, A.; Kalinowski, J.; Fedorenko, V.; Ostash, B. Heterologous AdpA transcription factors enhance landomycin production in Streptomyces cyanogenus S136 under a broad range of growth conditions. Appl. Microbiol. Biotechnol. 2018, 102, 8419–8428. [Google Scholar] [CrossRef]
- Bu, X.L.; Weng, J.Y.; He, B.B.; Xu, M.J.; Xu, J. A novel AdpA homologue negatively regulates morphological differentiation in Streptomyces xiamenensis 318. Appl. Environ. Microbiol. 2019, 85, e03107-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.; Wang, Y.; Hou, B.; Wang, R.; Ye, J.; Zhu, X.; Wu, H.; Zhang, H. AdpAlin, a pleiotropic transcriptional regulator, is involved in the cascade regulation of lincomycin biosynthesis in Streptomyces lincolnensis. Front. Microbiol. 2019, 10, 2428. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Wu, X.; Cao, Q.; Xia, Y.; Xun, L.; Liu, H. Sulfane sulfur posttranslationally modifies the global regulator AdpA to influence actinorhodin production and morphological differentiation of Streptomyces coelicolor. mBio 2022, 13, e0386221. [Google Scholar] [CrossRef] [PubMed]
- Westhoff, S.; Kloosterman, A.M.; van Hoesel, S.F.A.; van Wezel, G.P.; Rozen, D.E. Competition sensing changes antibiotic production in Streptomyces. mBio 2021, 12, e02729-20. [Google Scholar] [CrossRef]
- Lu, T.; Cao, Q.; Pang, X.; Xia, Y.; Xun, L.; Liu, H. Sulfane sulfur-activated actinorhodin production and sporulation is maintained by a natural gene circuit in Streptomyces coelicolor. Microb. Biotechnol. 2020, 13, 1917–1932. [Google Scholar] [CrossRef]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics, 2nd ed.; John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Fink, D.; Weissschuh, N.; Reuther, J.; Wohlleben, W.; Engels, A. Two transcriptional regulators GlnR and GlnRII are involved in regulation of nitrogen metabolism in Streptomyces coelicolor A3(2). Mol. Microbiol. 2002, 46, 331–347. [Google Scholar] [CrossRef]
- Bai, C.; Zhang, Y.; Zhao, X.; Hu, Y.; Xiang, S.; Miao, J.; Lou, C.; Zhang, L. Exploiting a precise design of universal synthetic modular regulatory elements to unlock the microbial natural products in Streptomyces. Proc. Natl. Acad. Sci. USA 2015, 112, 12181–12186. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zhang, P.; Lu, T.; Wang, X.; Li, A.; Lu, Y.; Tao, M.; Pang, X. Impact of MtrA on phosphate metabolism genes and the response to altered phosphate conditions in Streptomyces. Environ. Microbiol. 2021, 23, 6907–6923. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, X.; Li, H.; Liu, H.; Xia, Y.; Xun, L. The complete pathway for thiosulfate utilization in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2018, 84, e01241-18. [Google Scholar] [CrossRef] [Green Version]
- Ran, M.; Wang, T.; Shao, M.; Chen, Z.; Liu, H.; Xia, Y.; Xun, L. Sensitive method for reliable quantification of sulfane sulfur in biological samples. Anal. Chem. 2019, 91, 11981–11986. [Google Scholar] [CrossRef]
- Lu, T.; Zhu, Y.; Zhang, P.; Sheng, D.; Cao, G.; Pang, X. SCO5351 is a pleiotropic factor that impacts secondary metabolism and morphological development in Streptomyces coelicolor. FEMS Microbiol. Lett. 2018, 365, fny150. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zhang, P.; Zhang, J.; Xu, W.; Wang, X.; Wu, L.; Sheng, D.; Ma, W.; Cao, G.; Chen, X.L.; et al. The developmental regulator MtrA binds GlnR boxes and represses nitrogen metabolism genes in Streptomyces coelicolor. Mol. Microbiol. 2019, 112, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Bystrykh, L.V.; Fernandez-Moreno, M.A.; Herrema, J.K.; Malpartida, F.; Hopwood, D.A.; Dijkhuizen, L. Production of actinorhodin-related “blue pigments” by Streptomyces coelicolor A3(2). J. Bacteriol. 1996, 178, 2238–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, H.; Tomono, A.; Ohnishi, Y.; Horinouchi, S. DNA-binding specificity of AdpA, a transcriptional activator in the A-factor regulatory cascade in Streptomyces griseus. Mol. Microbiol. 2004, 53, 555–572. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhu, J.; Li, X.; Wen, Y. Transcriptional regulator DasR represses daptomycin production through both direct and cascade mechanisms in Streptomyces roseosporus. Antibiotics 2022, 11, 1065. [Google Scholar] [CrossRef] [PubMed]
- Grossoehme, N.; Kehl-Fie, T.E.; Ma, Z.; Adams, K.W.; Cowart, D.M.; Scott, R.A.; Skaar, E.P.; Giedroc, D.P. Control of copper resistance and inorganic sulfur metabolism by paralogous regulators in Staphylococcus aureus. J. Biol. Chem. 2011, 286, 13522–13531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T.; Shen, J.; Fang, M.; Zhang, Y.; Hori, K.; Trinidad, J.C.; Bauer, C.E.; Giedroc, D.P.; Masuda, S. Sulfide-responsive transcriptional repressor SqrR functions as a master regulator of sulfide-dependent photosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, 2355–2360. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, J.; Lu, C.; Xia, Y.; Xin, Y.; Liu, H.; Xun, L.; Liu, H. FisR activates σ54-dependent transcription of sulfide-oxidizing genes in Cupriavidus pinatubonensis JMP134. Mol. Microbiol. 2017, 105, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Walsh, B.J.C.; Wang, J.; Edmonds, K.A.; Palmer, L.D.; Zhang, Y.; Trinidad, J.C.; Skaar, E.P.; Giedroc, D.P. The response of Acinetobacter baumannii to hydrogen sulfide reveals two independent persulfide-sensing systems and a connection to biofilm regulation. mBio 2020, 11, e01254-20. [Google Scholar] [CrossRef]
- Peng, H.; Zhang, Y.; Palmer, L.D.; Kehl-Fie, T.E.; Skaar, E.P.; Trinidad, J.C.; Giedroc, D.P. Hydrogen sulfide and reactive sulfur species impact proteome S-sulfhydration and global virulence regulation in Staphylococcus aureus. ACS Infect. Dis. 2017, 3, 744–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, G.; Lu, C.; Xu, H.; Chen, Z.; Li, K.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. Sulfane Sulfur is an intrinsic signal activating MexR-regulated antibiotic resistance in Pseudomonas aeruginosa. Mol. Microbiol. 2020, 114, 1038–1048. [Google Scholar] [CrossRef]
- Hou, N.; Yan, Z.; Fan, K.; Li, H.; Zhao, R.; Xia, Y.; Xun, L.; Liu, H. OxyR senses sulfane sulfur and activates the genes for its removal in Escherichia coli. Redox Biol. 2019, 26, 101293. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.D.; Ohtsuka, J.; Nagata, K.; Miyazono, K.; Zhi, Y.; Ohnishi, Y.; Tanokura, M. Complex structure of the DNA-binding domain of AdpA, the global transcription factor in Streptomyces griseus, and a target duplex DNA reveals the structural basis of its tolerant DNA sequence specificity. J. Biol. Chem. 2013, 288, 31019–31029. [Google Scholar] [CrossRef] [Green Version]
- Zacharia, V.M.; Ra, Y.; Sue, C.; Alcala, E.; Reaso, J.N.; Ruzin, S.E.; Traxler, M.F. Genetic network architecture and environmental cues drive spatial organization of phenotypic division of labor in Streptomyces coelicolor. mBio 2021, 12, e00794-21. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Lu, X.; Sun, D.; Zhuang, S.; Chen, Q.; Chen, Z.; Li, J.; Wen, Y. BldD, a master developmental repressor, activates antibiotic production in two Streptomyces species. Mol. Microbiol. 2020, 113, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, J.; Liu, X.; Li, D.; Li, Y.; Tian, Y.; Tan, H. Identification of a butenolide signaling system that regulates nikkomycin biosynthesis in Streptomyces. J. Biol. Chem. 2018, 293, 20029–20040. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, J.; Zhuo, J.; Li, Y.; Tian, Y.; Tan, H. Activation and mechanism of a cryptic oviedomycin gene cluster via the disruption of a global regulatory gene, adpA, in Streptomyces ansochromogenes. J. Biol. Chem. 2017, 292, 19708–19720. [Google Scholar] [CrossRef] [Green Version]
- Mak, S.; Nodwell, J.R. Actinorhodin is a redox-active antibiotic with a complex mode of action against Gram-positive cells. Mol. Microbiol. 2017, 106, 597–613. [Google Scholar] [CrossRef]
- Nishiyama, T.; Hashimoto, Y.; Kusakabe, H.; Kumano, T.; Kobayashi, M. Natural low-molecular mass organic compounds with oxidase activity as organocatalysts. Proc. Natl. Acad. Sci. USA 2014, 111, 17152–17157. [Google Scholar] [CrossRef]
- Honma, S.; Ito, S.; Yajima, S.; Sasaki, Y. Nitric oxide signaling for actinorhodin production in Streptomyces coelicolor A3(2) via the DevS/R two-component system. Appl. Environ. Microbiol. 2021, 87, e0048021. [Google Scholar] [CrossRef] [PubMed]
- Lejeune, C.; Sago, L.; Cornu, D.; Redeker, V.; Virolle, M.J. A proteomic analysis indicates that oxidative stress is the common feature triggering antibiotic production in Streptomyces coelicolor and in the pptA mutant of Streptomyces lividans. Front. Microbiol. 2021, 12, 813993. [Google Scholar] [CrossRef] [PubMed]
- Esnault, C.; Dulermo, T.; Smirnov, A.; Askora, A.; David, M.; Deniset-Besseau, A.; Holland, I.B.; Virolle, M.J. Strong antibiotic production is correlated with highly active oxidative metabolism in Streptomyces coelicolor M145. Sci. Rep. 2017, 7, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikura, N.; Darbon, E.; Esnault, C.; Deniset-Besseau, A.; Xu, D.; Lejeune, C.; Jacquet, E.; Nhiri, N.; Sago, L.; Cornu, D.; et al. The phosin PptA plays a negative role in the regulation of antibiotic production in Streptomyces lividans. Antibiotics 2021, 10, 325. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Singh, A.K.; Cheon, D.J.; Roe, J.H. Activation of the SoxR regulon in Streptomyces coelicolor by the extracellular form of the pigmented antibiotic actinorhodin. J. Bacteriol. 2011, 193, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Dela Cruz, R.; Gao, Y.; Penumetcha, S.; Sheplock, R.; Weng, K.; Chander, M. Expression of the Streptomyces coelicolor SoxR regulon is intimately linked with actinorhodin production. J. Bacteriol. 2010, 192, 6428–6438. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Lu, X.; Yang, H.; Yan, H.; Wen, Y. Redox-sensitive transcriptional regulator SoxR directly controls antibiotic production, development and thiol-oxidative stress response in Streptomyces avermitilis. Microb. Biotechnol. 2022, 15, 561–576. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, T.; Wang, Q.; Cao, Q.; Xia, Y.; Xun, L.; Liu, H. The Pleiotropic Regulator AdpA Regulates the Removal of Excessive Sulfane Sulfur in Streptomyces coelicolor. Antioxidants 2023, 12, 312. https://doi.org/10.3390/antiox12020312
Lu T, Wang Q, Cao Q, Xia Y, Xun L, Liu H. The Pleiotropic Regulator AdpA Regulates the Removal of Excessive Sulfane Sulfur in Streptomyces coelicolor. Antioxidants. 2023; 12(2):312. https://doi.org/10.3390/antiox12020312
Chicago/Turabian StyleLu, Ting, Qingda Wang, Qun Cao, Yongzhen Xia, Luying Xun, and Huaiwei Liu. 2023. "The Pleiotropic Regulator AdpA Regulates the Removal of Excessive Sulfane Sulfur in Streptomyces coelicolor" Antioxidants 12, no. 2: 312. https://doi.org/10.3390/antiox12020312