


The Effect of Carnosine on UVA-Induced Changes in Intracellular Signaling of Human Skin Fibroblast Spheroids
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Source and Culture
2.2. VitroScreen ORA® Dermis Spheroid Production
2.3. Nucleic Acid Extractions for Nanostring Analysis and RNA QC
2.4. Gene Expression Analysis Using Nanostring nCounter System
2.5. L-Carnosine Treatment and UVA Exposure
2.6. Protein Extraction and In-Solution Trypsin Digestion
2.7. High-Resolution LC-MS/MS Analysis and Data Elaboration
2.8. Protein Network Analysis
3. Results and Discussion
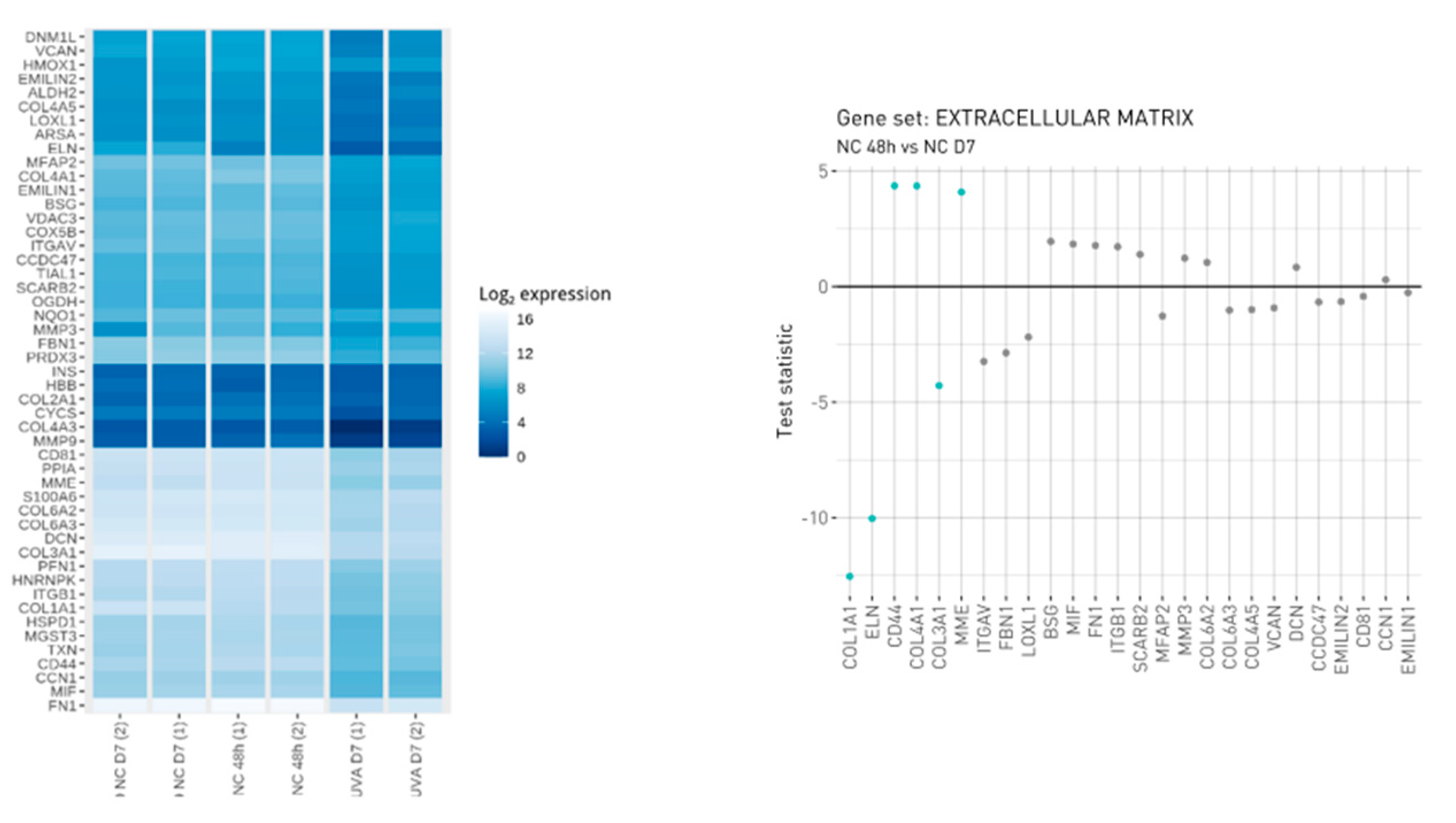
3.1. VitroScreen ORA® Dermis Model to Study the UVA Dermis Damage and Protection Effect of Carnosine
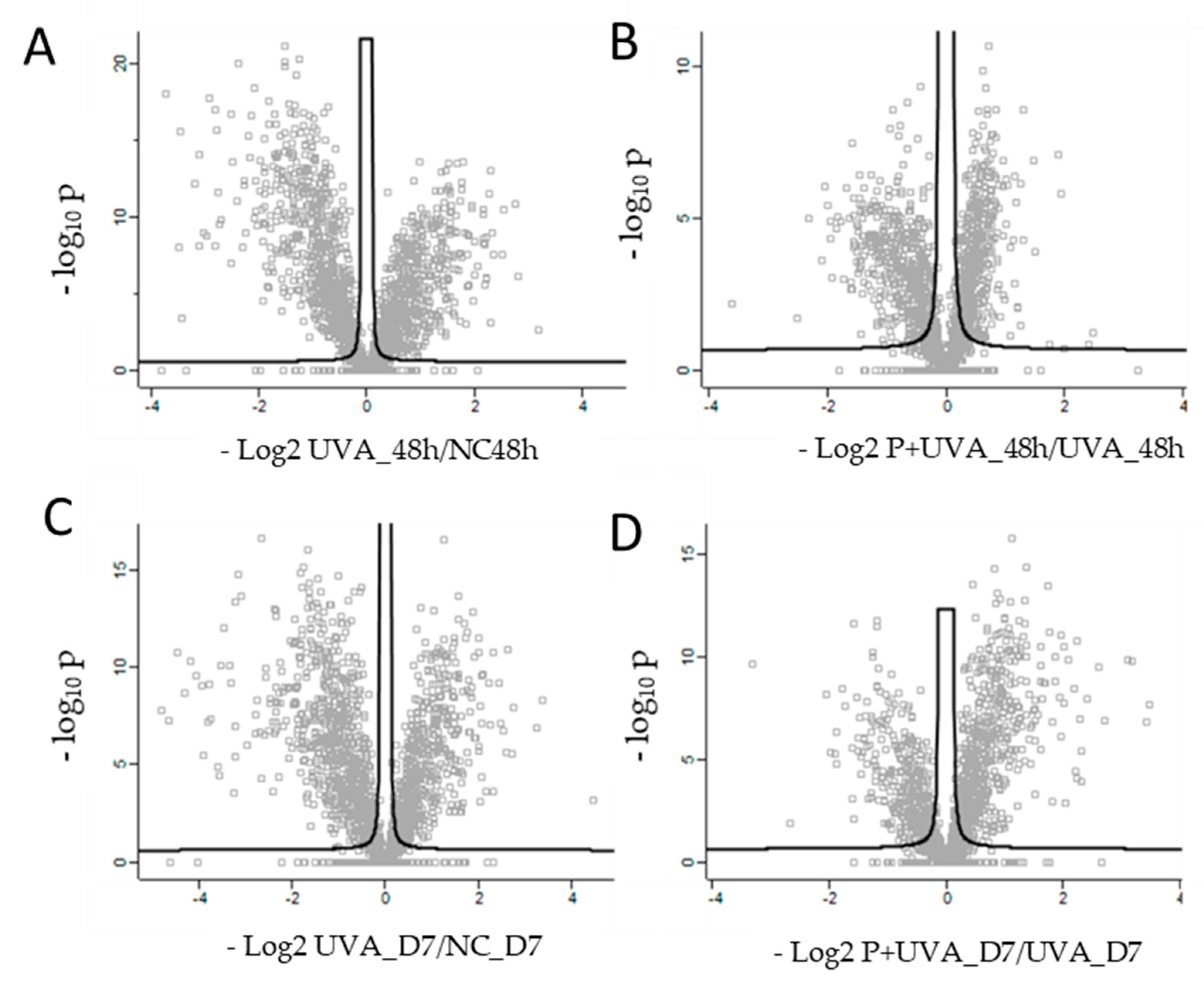
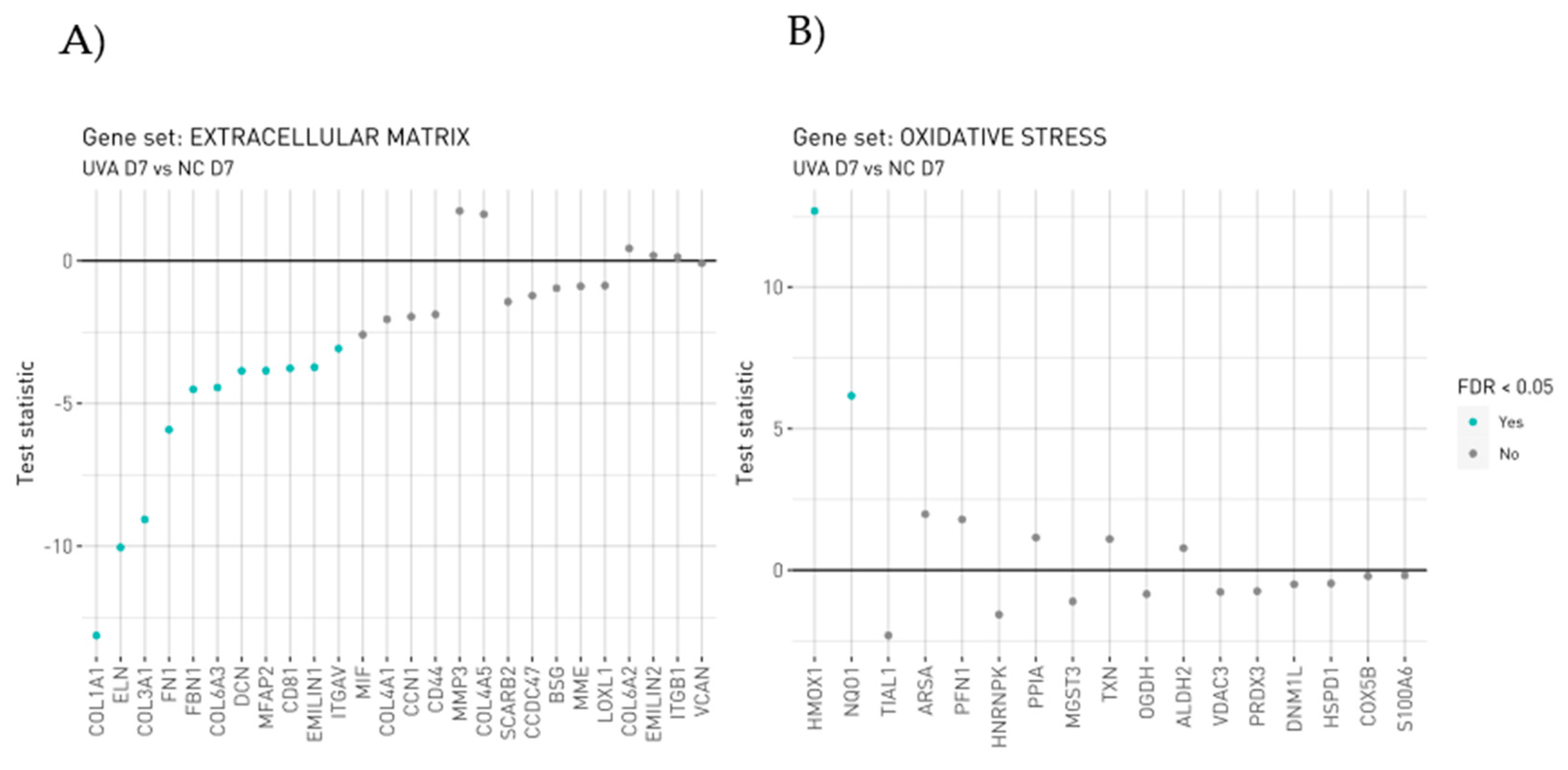
3.2. Protein Profiling of Human Dermis Spheroids Model after Exposure to UVA and Carnosine Treatment
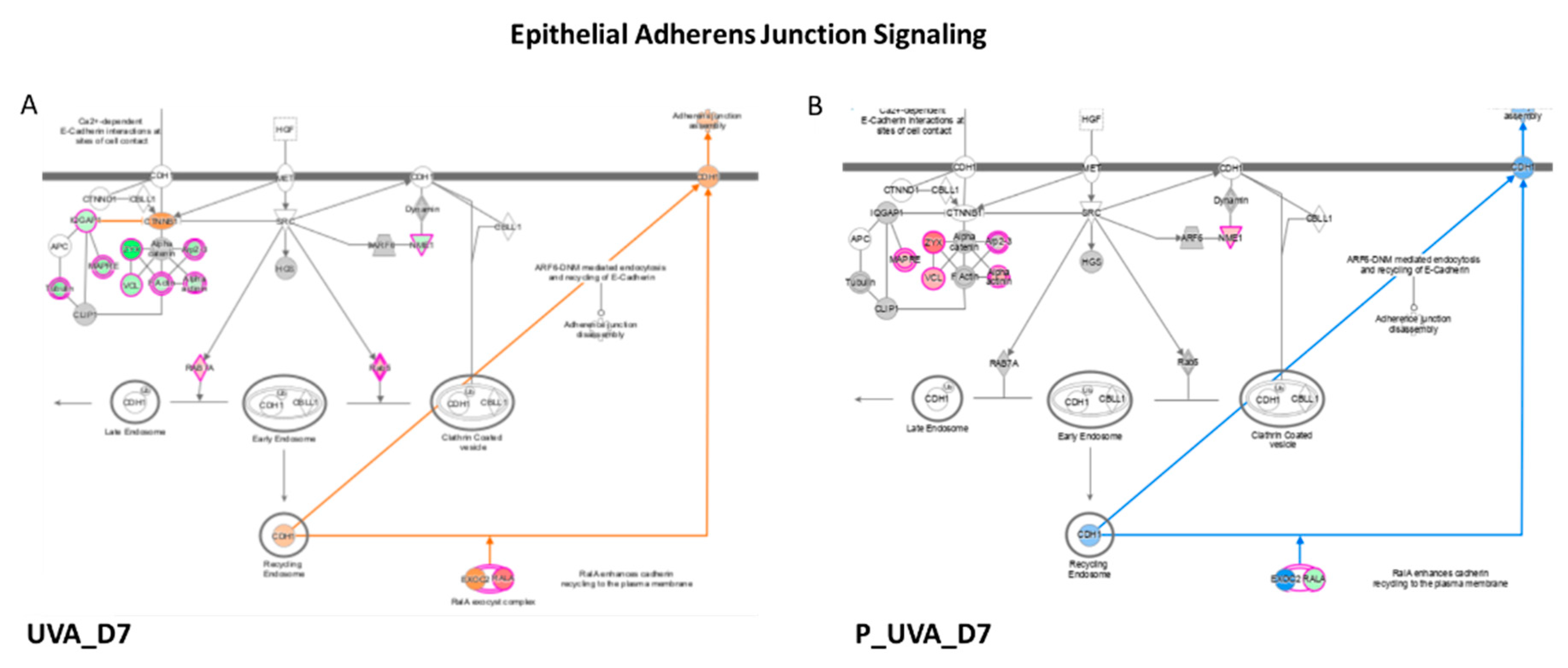
3.3. UVA Induces the Modulation of Epithelial Adherens Junction Signaling
3.4. UVA Induces the Modulation of Wound Healing Signaling Pathway
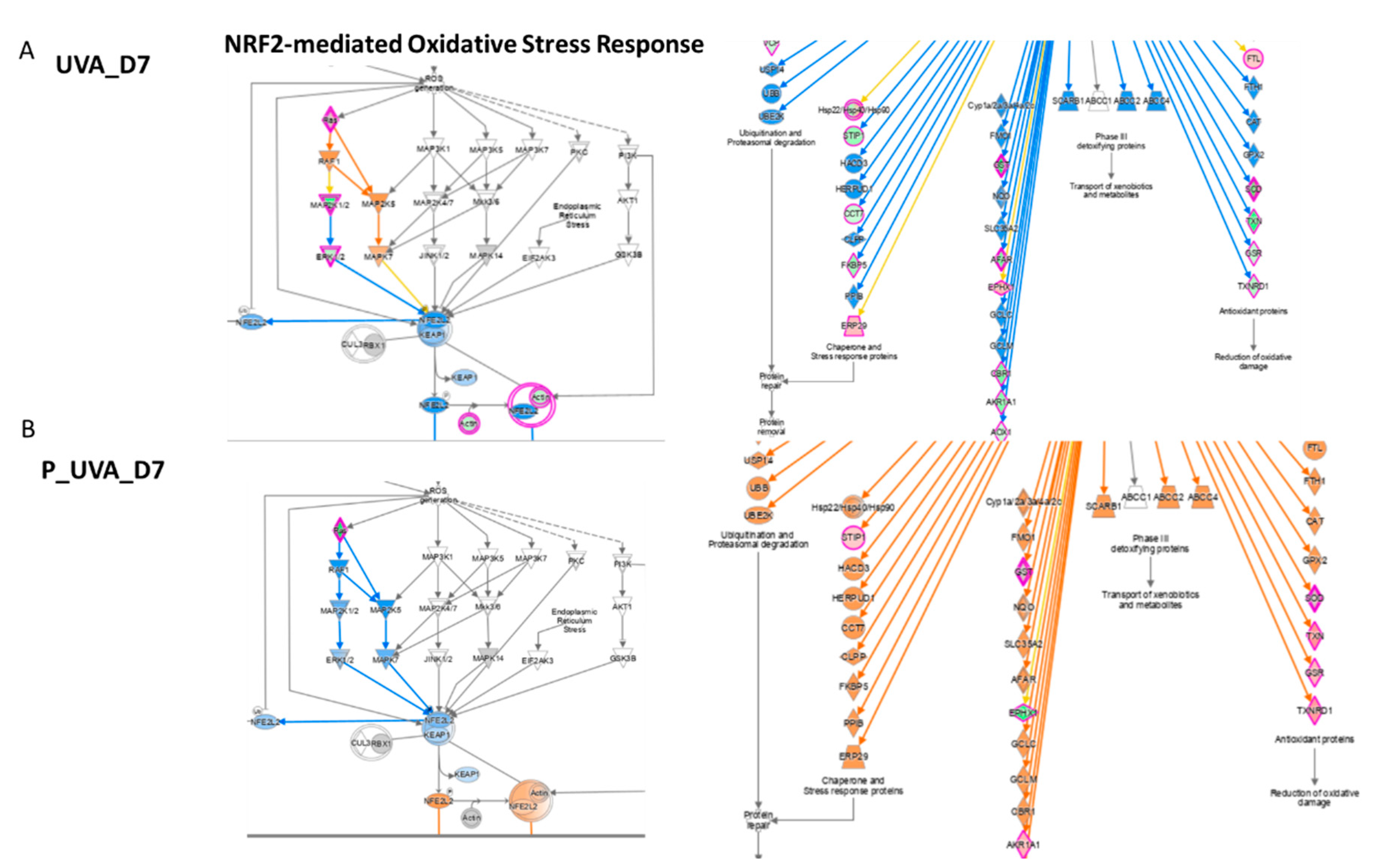
3.5. UVA Induces Nrf2-Mediated Oxidative Stress Response
3.6. UVA Regulates the Glycolysis and the Oxidative Phosphorylation
3.7. UVA Radiations Regulates the Sirtuin Signaling Pathway
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. Uv radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jager, T.L.; Cockrell, A.E.; Du Plessis, S.S. Ultraviolet light induced generation of reactive oxygen species. In Ultraviolet Light in Human Health, Diseases and Environment; Springer: Cham, Switzerland, 2017; Volume 996, pp. 15–23. [Google Scholar] [CrossRef]
- Spickett, C.M.; Pitt, A.R. Modification of proteins by reactive lipid oxidation products and biochemical effects of lipoxidation. Protein Oxid. 2020, 64, 19–31. [Google Scholar] [CrossRef] [PubMed]
- He, H.L.; Xiong, L.D.; Jian, L.E.; Li, L.M.; Wu, Y.L.; Qiao, S.Y. Role of mitochondria on uv-induced skin damage and molecular mechanisms of active chemical compounds targeting mitochondria. J. Photochem. Photobiol. B-Biol. 2022, 232, 112464. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.H.; Xu, G.Z.; Sayre, L.M. Carnosine inhibits (e)-4-hydroxy-2-nonenal-induced protein cross-linking: Structural characterization of carnosine-hne adducts. Chem. Res. Toxicol. 2003, 16, 1589–1597. [Google Scholar] [CrossRef]
- Davies, S.S.; Brantley, E.J.; Voziyan, P.A.; Amarnath, V.; Zagol-Ikapitte, I.; Boutaud, O.; Hudson, B.G.; Oates, J.A.; Roberts, L.J. Pyridoxamine analogues scavenge lipid-derived gamma-ketoaldehydes and protect against H2O2-mediated cytotoxicity. Biochemistry 2006, 45, 15756–15767. [Google Scholar] [CrossRef] [Green Version]
- Aiello, G.; Rescigno, F.; Meloni, M.; Baron, G.; Aldini, G.; Carini, M.; D’Amato, A. Oxidative stress modulation by carnosine in scaffold free human dermis spheroids model: A proteomic study. Int. J. Mol. Sci. 2022, 23, 1468. [Google Scholar] [CrossRef]
- Silvia, R.; Marina, C.; Baron, G.; Giancarlo, A.; Anne, N.S.; Alfonsina, D. Study of carnosine’s effect on nude mice skin to prevent uv-a damage. Free. Radic. Biol. Med. 2021, 173, 97–103. [Google Scholar] [CrossRef]
- Nakyai, W.; Tissot, M.; Humbert, P.; Grandmottet, F.; Viyoch, J.; Viennet, C. Effects of repeated uva irradiation on human skin fibroblasts embedded in 3d tense collagen matrix. Photochem. Photobiol. 2018, 94, 715–724. [Google Scholar] [CrossRef]
- Meloni, M.; Farina, A.; de Servi, B. Molecular modifications of dermal and epidermal biomarkers following uva exposures on reconstructed full-thickness human skin. Photochem. Photobiol. Sci. 2010, 9, 439–447. [Google Scholar] [CrossRef]
- Fabregat, A.; Jupe, S.; Matthews, L.; Sidiropoulos, K.; Gillespie, M.; Garapati, P.; Haw, R.; Jassal, B.; Korninger, F.; May, B.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2018, 46, D649–D655. [Google Scholar] [CrossRef]
- Beckerle, M.C. Zyxin: Zinc fingers at sites of cell adhesion. Bioessays 1997, 19, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Pylayeva-Gupta, Y.; Grabocka, E.; Bar-Sagi, D. Ras oncogenes: Weaving a tumorigenic web. Nat. Rev. Cancer 2011, 11, 761–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrin, B.J.; Amann, K.J.; Huttenlocher, A. Proteolysis of cortactin by calpain regulates membrane protrusion during cell migration. Mol. Biol. Cell 2006, 17, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Battie, C.; Jitsukawa, S.; Bernerd, F.; Del Bino, S.; Marionnet, C.; Verschoore, M. New insights in photoaging, uva induced damage and skin types. Exp. Dermatol. 2014, 23, 7–12. [Google Scholar] [CrossRef]
- Basu, P.; Martins-Green, M. Signaling pathways associated with chronic wound progression: A systems biology approach. Antioxidants 2022, 11, 1506. [Google Scholar] [CrossRef]
- Keene, D.R.; Engvall, E.; Glanville, R.W. Ultrastructure of type-vi collagen in human-skin and cartilage suggests an anchoring function for this filamentous network. J. Cell Biol. 1988, 107, 1995–2006. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.-L. The keap1-nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sykiotis, G.P.; Bohmann, D. Stress-activated cap’n’collar transcription factors in aging and human disease. Sci. Signal. 2010, 3, re3. [Google Scholar] [CrossRef] [Green Version]
- Ludtmann, M.H.R.; Angelova, P.R.; Zhang, Y.; Abramov, A.Y.; Dinkova-Kostova, A.T. Nrf2 affects the efficiency of mitochondrial fatty acid oxidation. Biochem. J. 2014, 457, 415–424. [Google Scholar] [CrossRef]
- Hiebert, P.; Wietecha, M.S.; Cangkrama, M.; Haertel, E.; Mavrogonatou, E.; Stumpe, M.; Steenbock, H.; Grossi, S.; Beer, H.D.; Angel, P.; et al. Nrf2-mediated fibroblast reprogramming drives cellular senescence by targeting the matrisome. Dev. Cell 2018, 46, 145–161.e10. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.C.; Kim, C.; Kim, J.Y.; Chung, J.H.; Kim, J.H. UVB Radiation Induces Apoptosis in Keratinocytes by Activating a Pathway Linked to “BLT2-Reactive Oxygen Species”. J. Investig. Dermatol. 2010, 130, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Mavrogonatou, E.; Angelopoulou, M.; Rizou, S.V.; Pratsinis, H.; Gorgoulis, V.G.; Kletsas, D. Activation of the JNKs/ATM-p53 axis is indispensable for the cytoprotection of dermal fibroblasts exposed to UVB radiation. Cell Death Dis. 2022, 13, 647. [Google Scholar] [CrossRef] [PubMed]
- Panich, U.; Sittithumcharee, G.; Rathviboon, N.; Jirawatnotai, S. Ultraviolet radiation-induced skin aging: The role of dna damage and oxidative stress in epidermal stem cell damage mediated skin aging. Stem Cells Int. 2016, 2016, 7370642. [Google Scholar] [CrossRef] [Green Version]
- Mullarky, E.; Cantley, L.C.; Nakao, K.; Minato, N.; Uemoto, S. Diverting glycolysis to combat oxidative stress. In Innovative Medicine: Basic Research and Development; Springer: Tokyo, Japan, 2015. [Google Scholar]
- Ying, W.H. NAD+/NADH and NADP+/NADPH in cellular functions and cell death: Regulation and biological consequences. Antioxid. Redox Signal. 2008, 10, 179–206. [Google Scholar] [CrossRef] [Green Version]
- Blacker, T.S.; Mann, Z.F.; Gale, J.E.; Ziegler, M.; Bain, A.J.; Szabadkai, G.; Duchen, M.R. Separating nadh and nadph fluorescence in live cells and tissues using flim. Nat. Commun. 2014, 5, 3936. [Google Scholar] [CrossRef] [Green Version]
- Ung, T.P.L.; Lim, S.; Solinas, X.; Mahou, P.; Chessel, A.; Marionnet, C.; Bornschlögl, T.; Beaurepaire, E.; Bernerd, F.; Pena, A.M.; et al. Simultaneous NAD(P)H and FAD fluorescence lifetime microscopy of long uva-induced metabolic stress in reconstructed human skin. Sci. Rep. 2021, 11, 22171. [Google Scholar] [CrossRef] [PubMed]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef] [PubMed]
- Kalfalah, F.; Sobek, S.; Bornholz, B.; Götz-Rösch, C.; Tigges, J.; Fritsche, E.; Krutmann, J.; Köhrer, K.; Deenen, R.; Ohse, S.; et al. Inadequate mito-biogenesis in primary dermal fibroblasts from old humans is associated with impairment of pgc1a-independent stimulation. Exp. Gerontol. 2014, 56, 59–68. [Google Scholar] [CrossRef]
- Wahedi, H.M.; Lee, T.H.; Moon, E.-Y.; Kim, S.Y. Juglone up-regulates sirt1 in skin cells under normal and uvb irradiated conditions. J. Dermatol. Sci. 2016, 81, 210–212. [Google Scholar] [CrossRef]
- Lang, A.; Grether-Beck, S.; Singh, M.; Kuck, F.; Jakob, S.; Kefalas, A.; Altinoluk-Hambüchen, S.; Graffmann, N.; Schneider, M.; Lindecke, A.; et al. Microrna-15b regulates mitochondrial ros production and the senescence-associated secretory phenotype through sirtuin 4/sirt4. Aging-Us 2016, 8, 484–505. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Z-Score | ||||
|---|---|---|---|---|
| Canonical Pathways | UVA_48h | P_UVA_48h | UVA_D7 | P_UVA_D7 |
| Oxidative Phosphorylation | 5.39 | −1.16 | 4.71 | −1.67 |
| Hepatic Fibrosis Signaling Pathway | 2.41 | −1.60 | 1.57 | −2.31 |
| Wound Healing Signaling Pathway | 1.53 | −2.53 | 1.61 | −3.16 |
| Estrogen Receptor Signaling | 3.14 | −2.11 | 1.89 | −2.89 |
| Macropinocytosis Signaling | 2.45 | −2.45 | 2.24 | −2.45 |
| GP6 Signaling Pathway | 2.14 | −1.63 | 0.91 | −2.12 |
| Pulmonary Fibrosis Idiopathic Signaling Pathway | 1.23 | −2.31 | 1.00 | −2.53 |
| G Beta Gamma Signaling | 2.11 | −2.65 | 0.91 | −2.65 |
| Gαi Signaling | 1.90 | −2.45 | 1.51 | −2.45 |
| Regulation of eIF4 and p70S6K Signaling | 1.67 | −2.45 | 1.73 | −2.45 |
| p70S6K Signaling | 1.90 | −2.45 | 1.39 | −2.24 |
| Thrombin Signaling | 1.51 | −2.83 | 0.91 | −2.65 |
| NF-κB Activation by Viruses | 1.89 | −2.24 | 1.41 | −2.24 |
| Regulation of Cellular Mechanics by Calpain Protease | 2.12 | −1.89 | 0.91 | −2.45 |
| Glioma invasiveness Signaling | 1.89 | −2.45 | 0.63 | −2.24 |
| HGF Signaling | 1.16 | −2.45 | 0.28 | −2.45 |
| UVA-induced MAPK signaling | 1.13 | −2.00 | 1.13 | −2.00 |
| CDK5 Signaling | 1.51 | −2.24 | 0.26 | −2.24 |
| PEDF Signaling | 1.14 | −1.63 | 0.71 | −2.24 |
| Fc Epsilon RI signaling | −1.13 | −2.-24 | 0.38 | −2.-24 |
| Glycolysis I | −3.46 | 1.41 | −3.61 | 2.83 |
| PTEN Signaling | −1.73 | 2.65 | −1.16 | 2.45 |
| Epithelial Adherens Junction Signaling | −2.20 | 1.27 | −2.35 | 2.00 |
| CLEAR Signaling Pathway | −1.67 | 1.70 | −1.41 | 2.45 |
| Gluconeogenesis I | −2.11 | 2.45 | −2.11 | 2.83 |
| Sirtuin Signaling Pathway | −0.85 | −2.32 | −1.26 | −0.83 |
| Apotosis | 1.25 | −0.94 | 2.19 | −2.82 |
| Epithelial Adherens Junction Signaling | UVA_48 h | P_UVA_48 h | UVA_D7 | P_UVA_D7 | ||
|---|---|---|---|---|---|---|
| Gene Name | Protein Name | Accession Number | Log Ratio | Log Ratio | Log Ratio | Log Ratio |
| RALA | RAS-like proto-oncogene A | P11233 | 1.53 | −0.66 | 1.59 | −0.78 |
| RAB7A | RAB7A, member RAS oncogene family | A0A158RFU6 | 0.79 | −0.61 | 0.67 | −0.44 |
| RAB5C | RAB5C, member RAS oncogene family | A0A024R1U4 | 0.72 | −0.38 | 0.82 | −0.30 |
| ARPC4 | Actin-related protein 2/3 complex subunit 4 | P59998 | −0.93 | 0.46 | −0.53 | 0.26 |
| ARPC1A | Actin-related protein 2/3 complex subunit 1A | Q75MY0 | −0.65 | 0.25 | −0.76 | 0.31 |
| ARPC2 | Actin-related protein 2/3 complex subunit 2 | Q53R19 | −1.00 | 0.36 | −0.40 | 0.36 |
| NME1 | NME/NM23 nucleoside diphosphate kinase 1 | P15531 | −1.01 | 0.62 | −0.77 | 0.64 |
| MAPRE1 | Microtubule-associated protein RP/EB family member 1 | Q15691 | −1.29 | 0.62 | −1.18 | 0.70 |
| ACTN4 | Actinin alpha 4 | A0A0S2Z3G9 | −0.68 | 0.54 | −0.69 | 0.79 |
| ACTN1 | Actinin alpha 1 | A0A024R694 | −0.88 | 0.57 | −0.83 | 0.87 |
| VCL | Vinculin | A0A024QZN4 | −0.86 | 0.35 | −1.11 | 0.87 |
| ZYX | Zyxin | Q15942 | −2.13 | 0.38 | −2.42 | 1.52 |
| Wound Healing Signaling Pathway | UVA_48 h | P_UVA_48 h | UVA_D7 | P_UVA_D7 | ||
|---|---|---|---|---|---|---|
| Gene Name | Protein Name | Accession Number | Log Ratio | Log Ratio | Log Ratio | Log Ratio |
| COL1A1 | Collagen type I alpha 1 chain | D3DTX7 | 1.99 | −0.99 | 0.96 | −1.27 |
| COL3A1 | Collagen type III alpha 1 chain | P02461 | 0.79 | 0.53 | 0.45 | 0.00 |
| COL1A2 | Collagen type I alpha 2 chain | A0A087WTA8 | 1.20 | 0.00 | 0.49 | 0.00 |
| COL5A1 | Collagen type V alpha 1 chain | A0A024R8E5 | 0.99 | 0.24 | 0.31 | 0.00 |
| COL15A1 | Collagen type XV alpha 1 chain | P39059 | −1.74 | 0.51 | −2.35 | 0.48 |
| LAMA2 | Laminin subunit alpha 2 | A0A087WYF1 | 0.60 | 0.00 | 0.00 | 0.00 |
| COL6A2 | Collagen type VI alpha 2 chain | P12110 | 2.30 | −1.04 | 2.02 | −1.26 |
| COL6A1 | Collagen type VI alpha 1 chain | A0A087X0S5 | 1.77 | −0.50 | 1.71 | −0.93 |
| RRAS | RAS related | A0A024QZF2 | 0.93 | −0.41 | 1.05 | −0.85 |
| RALA | RAS-like proto-oncogene A | P11233 | 1.53 | −0.66 | 1.59 | −0.78 |
| RAP1B | Rap1b Member of RAS oncogene family | P61224 | 1.28 | −0.84 | 1.04 | −0.75 |
| ITGB1 | Integrin subunit beta 1 | P05556 | 1.36 | −0.74 | 1.30 | −0.71 |
| RAC1 | Rac family small gtpase 1 | A4D2P1 | 0.81 | −0.88 | 0.52 | −0.70 |
| COL4A2 | Collagen type IV alpha 2 chain | P08572 | 0.75 | 0.00 | 1.00 | −0.28 |
| MAP2K2 | Mitogen-activated protein kinase kinase 2 | P36507 | −1.49 | 0.00 | −2.00 | 0.00 |
| MAPK1 | Mitogen-activated protein kinase 1 | Q1HBJ4 | −0.73 | 0.00 | −0.94 | 0.00 |
| STAT1 | Signal transducer and activator of transcription 1 | P42224 | −0.95 | 0.00 | −0.60 | 0.00 |
| RRAS2 | RAS related 2 | P62070 | 0.77 | −1.04 | 0.95 | 0.00 |
| FN1 | Fibronectin 1 | A0A024R462 | −1.08 | 0.00 | −1.28 | 0.28 |
| COL18A1 | Collagen type XVIII alpha 1 chain | D3DSM4 | −1.36 | 0.58 | −1.38 | 0.55 |
| COL6A3 | Collagen type VI alpha 3 chain | B7ZW00 | 3.19 | −3.60 | 4.48 | −2.66 |
| Nrf2-Mediated Oxidative Stress Response | UVA_48h | P_UVA_48h | UVA_D7 | P_UVA_D7 | ||
|---|---|---|---|---|---|---|
| Gene Name | Protein Name | Accession Number | Log Ratio | Log Ratio | Log Ratio | Log Ratio |
| AKR1A1 | Aldo-keto reductase family 1 member A1 | V9HWI0 | −0.94 | 0.40 | −1.32 | 0.83 |
| CLPP | Caseinolytic mitochondrial matrix peptidase proteolytic subunit | Q16740 | 0.73 | −0.33 | 0.58 | 0.00 |
| EPHX1 | Epoxide hydrolase 1 | R4SBI6 | 1.30 | −1.18 | 0.97 | −0.89 |
| GSTM1 | Glutathione S-transferase mu 1 | X5DR03 | −1.29 | −0.61 | −2.43 | 1.00 |
| GSTM3 | Glutathione S-transferase mu 3 | Q6FGJ9 | −1.20 | 0.00 | −1.84 | 0.71 |
| GSTO1 | Glutathione S-transferase omega 1 | V9HWG9 | −1.69 | 0.73 | −1.67 | 1.24 |
| GSTP1 | Glutathione S-transferase pi 1 | V9HWE9 | −1.44 | 0.51 | −1.77 | 1.10 |
| GSR | Glutathione-disulphide reductase | V9HW90 | −0.92 | 0.64 | −0.72 | 0.77 |
| RAP1B | RAP1B, member of RAS oncogene family | P61224 | 1.28 | −0.84 | 1.04 | −0.75 |
| RALA | RAS-like proto-oncogene A | P11233 | 1.53 | −0.66 | 1.59 | −0.78 |
| RRAS | RAS related | A0A024QZF2 | 0.93 | −0.41 | 1.05 | −0.85 |
| RRAS2 | RAS related 2 | P62070 | 0.77 | −1.04 | 0.95 | 0.00 |
| SOD1 | Superoxide dismutase 1 | V9HWC9 | −1.91 | 0.62 | −2.19 | 1.30 |
| TXN | Thioredoxin | H9ZYJ2 | −1.22 | 0.00 | −1.87 | 1.19 |
| TXNRD1 | Thioredoxin reductase 1 | Q16881 | −1.15 | 0.74 | −0.93 | 1.09 |
| SOD2 | Superoxide dismutase 2 | P04179 | 1.44 | 0.00 | 1.65 | 0.00 |
| MAPK1 | Mitogen-activated protein kinase 1 | Q1HBJ4 | −0.728 | 0.00 | −0.939 | 0.00 |
| MAP2K2 | Mitogen-activated protein kinase kinase 2 | P36507 | −1.488 | 0.00 | −1.996 | 0.00 |
| Glycolysis | UVA_48 h | P_UVA_48 h | UVA_D7 | P_UVA_D7 | ||
|---|---|---|---|---|---|---|
| Gene Name | Protein Name | Accession Number | Log Ratio | Log Ratio | Log Ratio | Log Ratio |
| ALDOA | Aldolase, fructose-bisphosphate A | P04075 | −1.33 | 0.63 | −1.61 | 1.02 |
| ALDOC | Aldolase, fructose-bisphosphate C | A0A024QZ64 | −1.30 | 0.76 | −1.70 | 0.96 |
| ENO1 | Enolase 1 | A0A024R4F1 | −1.24 | 0.66 | −1.59 | 0.92 |
| ENO2 | Enolase 2 | Q6FHV6 | −1.26 | 0.74 | −2.14 | 1.58 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | V9HVZ4 | −1.14 | 0.34 | −1.24 | 0.63 |
| GPI | Glucose-6-phosphate isomerase | K7EIL4 | −1.66 | 0.71 | −1.85 | 1.28 |
| PFKL | Phosphofructokinase, liver type | P17858 | −1.22 | −1.07 | −1.88 | 0.00 |
| PFKM | Phosphofructokinase, muscle | P08237 | −1.00 | 0.00 | −0.93 | 0.00 |
| PGAM1 | Phosphoglycerate mutase 1 | Q6FHU2 | −0.88 | 0.46 | −1.14 | 0.52 |
| PGK1 | Phosphoglycerate kinase 1 | V9HWF4 | −1.51 | 0.54 | −1.82 | 0.78 |
| PKM | Pyruvate kinase M1/2 | A0A024R5Z9 | −1.00 | −0.23 | −1.64 | 0.00 |
| TPI1 | Triosephosphate isomerase 1 | V9HWK1 | −1.35 | 0.85 | −1.61 | 1.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aiello, G.; Rescigno, F.; Meloni, M.; Zoanni, B.; Aldini, G.; Carini, M.; D’Amato, A. The Effect of Carnosine on UVA-Induced Changes in Intracellular Signaling of Human Skin Fibroblast Spheroids. Antioxidants 2023, 12, 300. https://doi.org/10.3390/antiox12020300
Aiello G, Rescigno F, Meloni M, Zoanni B, Aldini G, Carini M, D’Amato A. The Effect of Carnosine on UVA-Induced Changes in Intracellular Signaling of Human Skin Fibroblast Spheroids. Antioxidants. 2023; 12(2):300. https://doi.org/10.3390/antiox12020300
Chicago/Turabian StyleAiello, Gilda, Francesca Rescigno, Marisa Meloni, Beatrice Zoanni, Giancarlo Aldini, Marina Carini, and Alfonsina D’Amato. 2023. "The Effect of Carnosine on UVA-Induced Changes in Intracellular Signaling of Human Skin Fibroblast Spheroids" Antioxidants 12, no. 2: 300. https://doi.org/10.3390/antiox12020300