
Methane Admixture Protects Liver Mitochondria and Improves Graft Function after Static Cold Storage and Reperfusion
 , , , , and
, , , , and 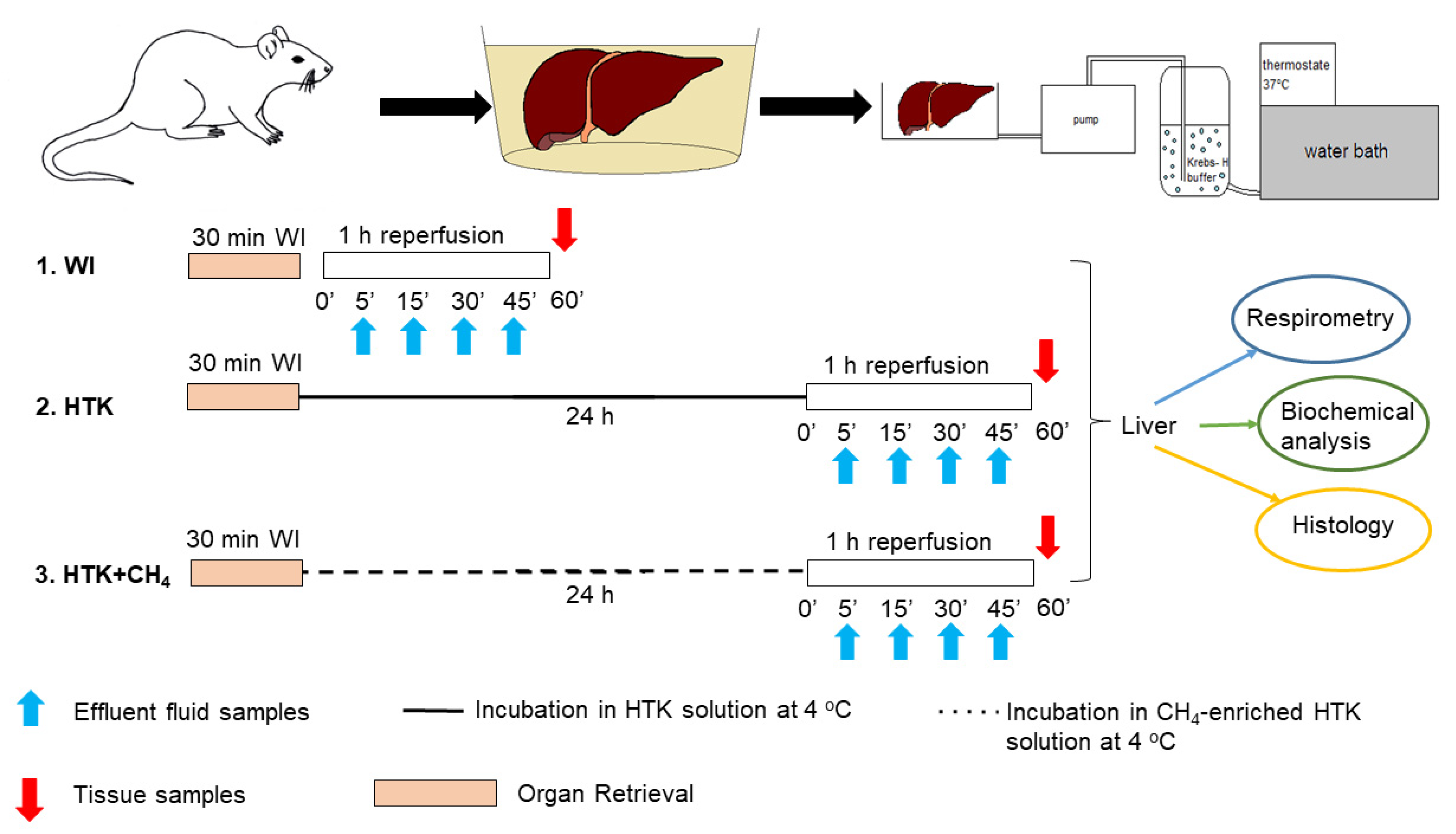{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production of CH4-Enriched HTK
2.2. Experimental Groups
2.3. Organ Perfusion System and Assessment of Hepatic Function and Cell Injury
2.4. High Resolution Respirometry
2.5. Histology and Immunohistochemistry
2.6. Statistics
3. Results
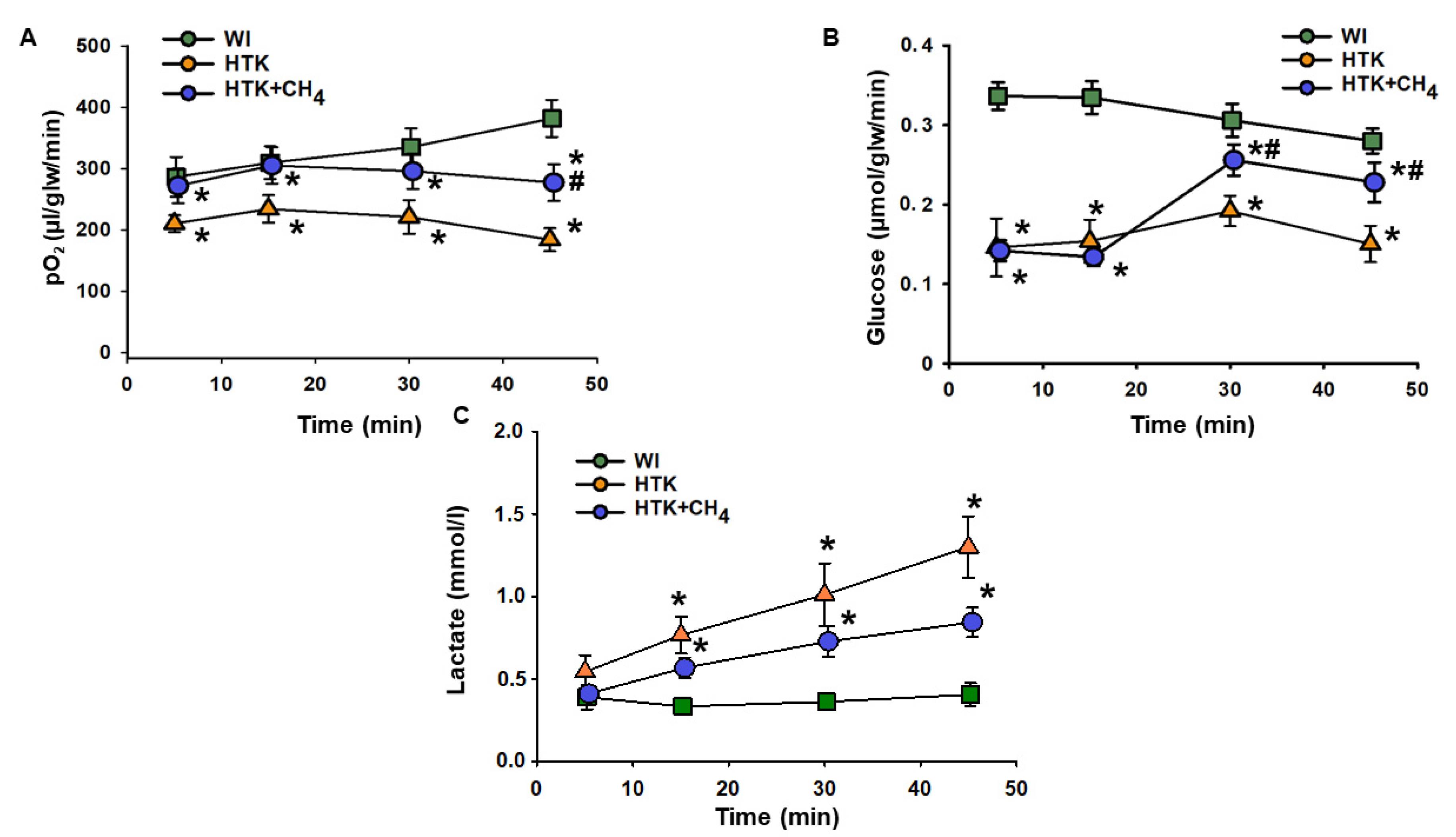
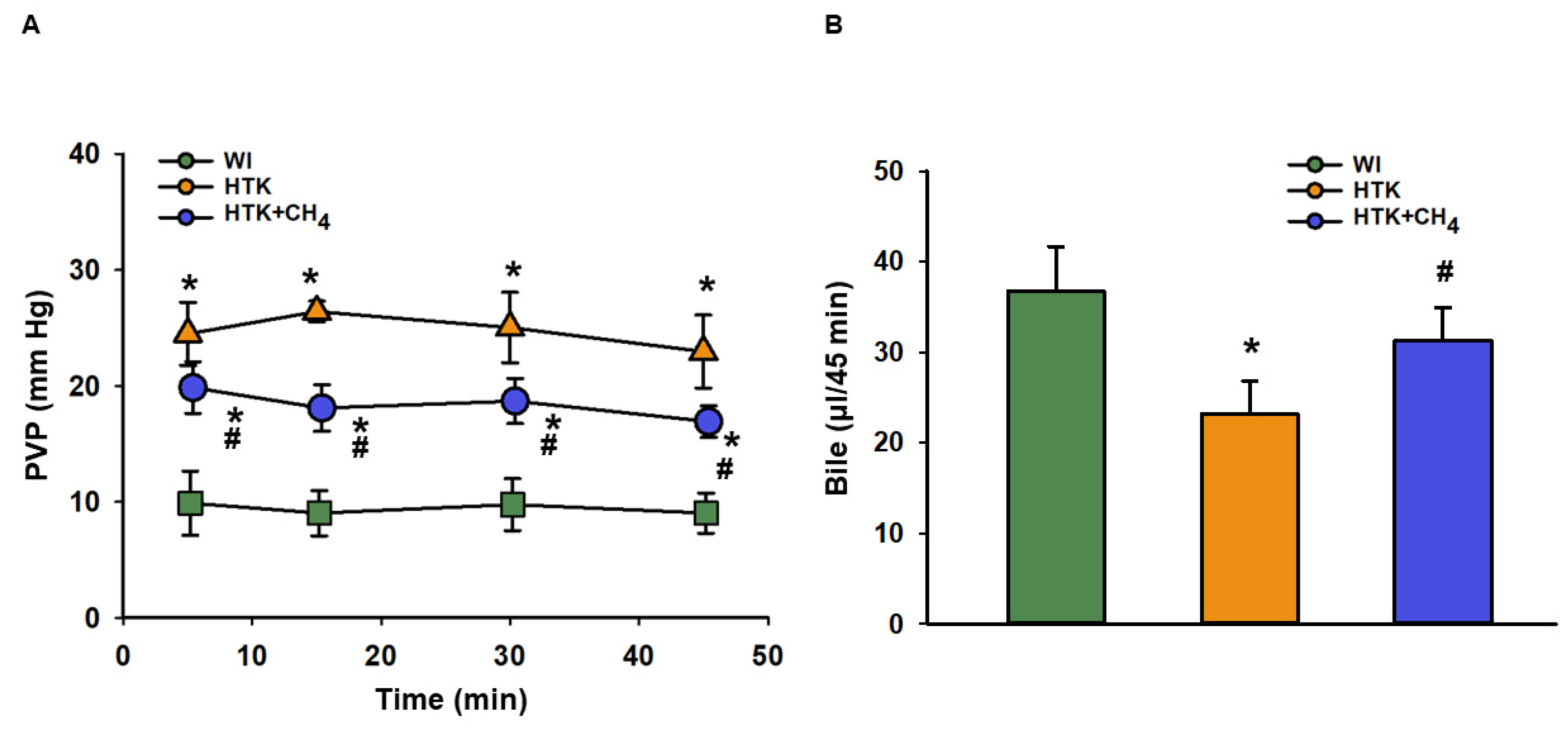
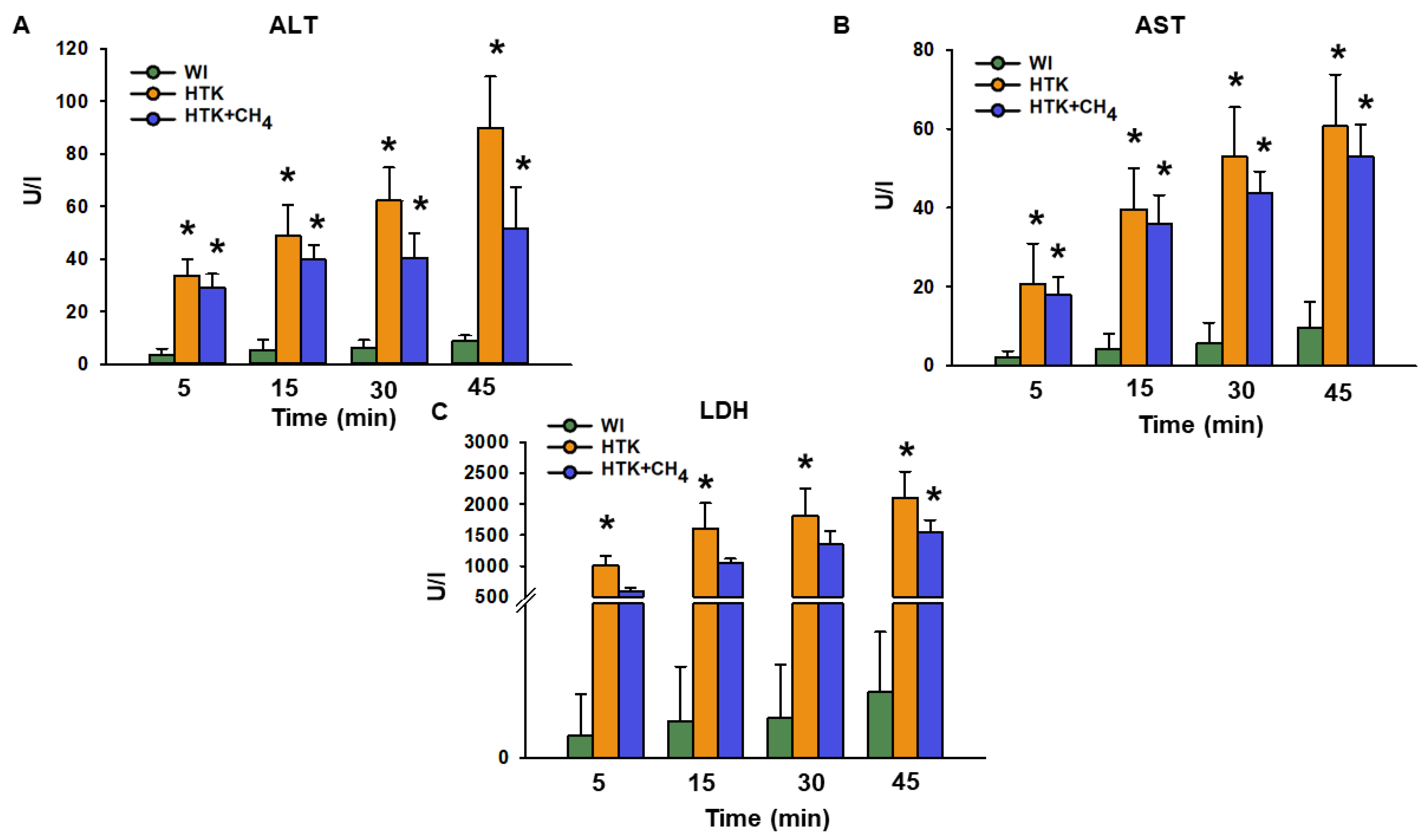
3.1. Graft Function and Cell Injury after Cold Storage
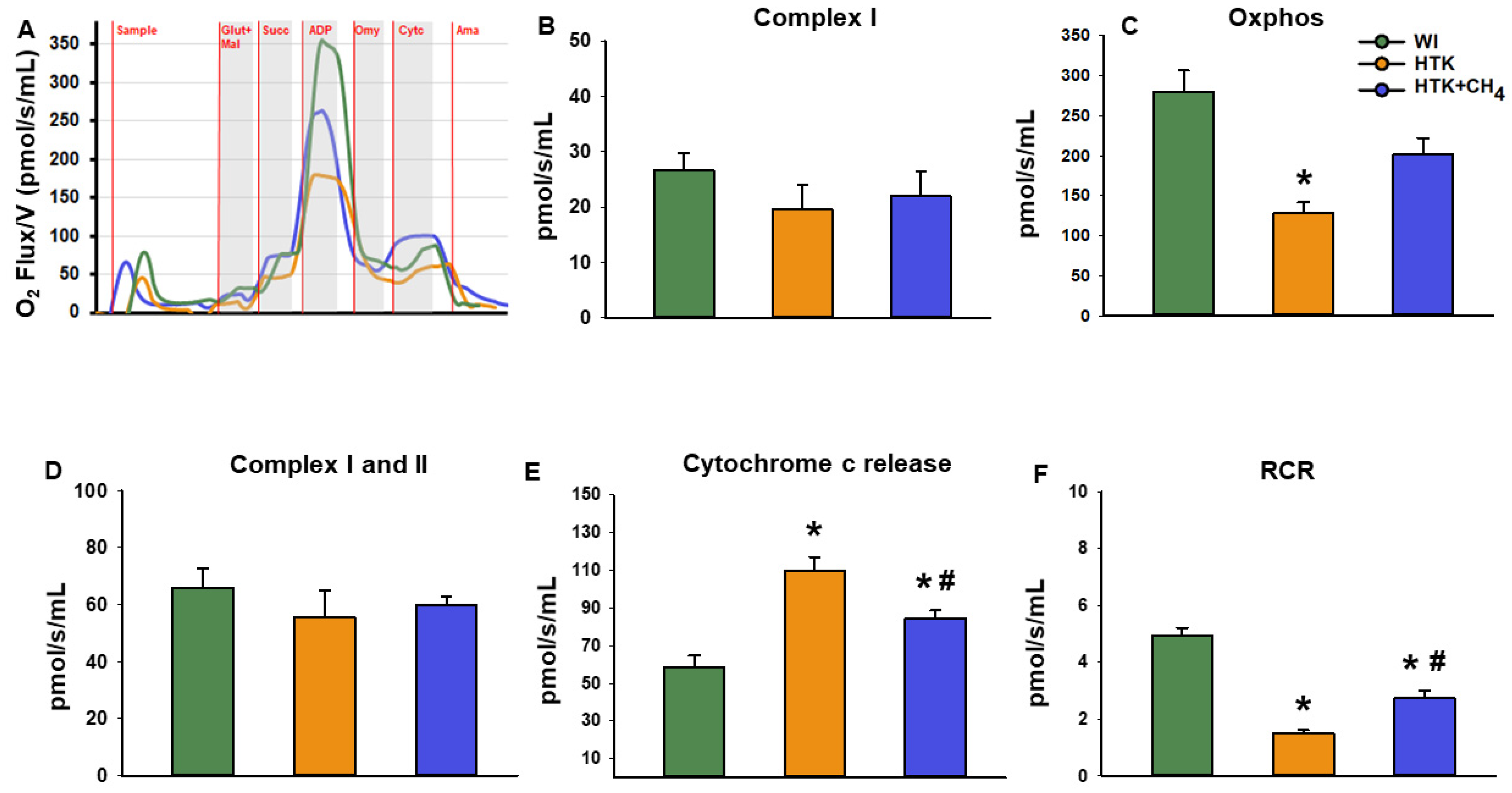
3.2. Mitochondrial Respiration
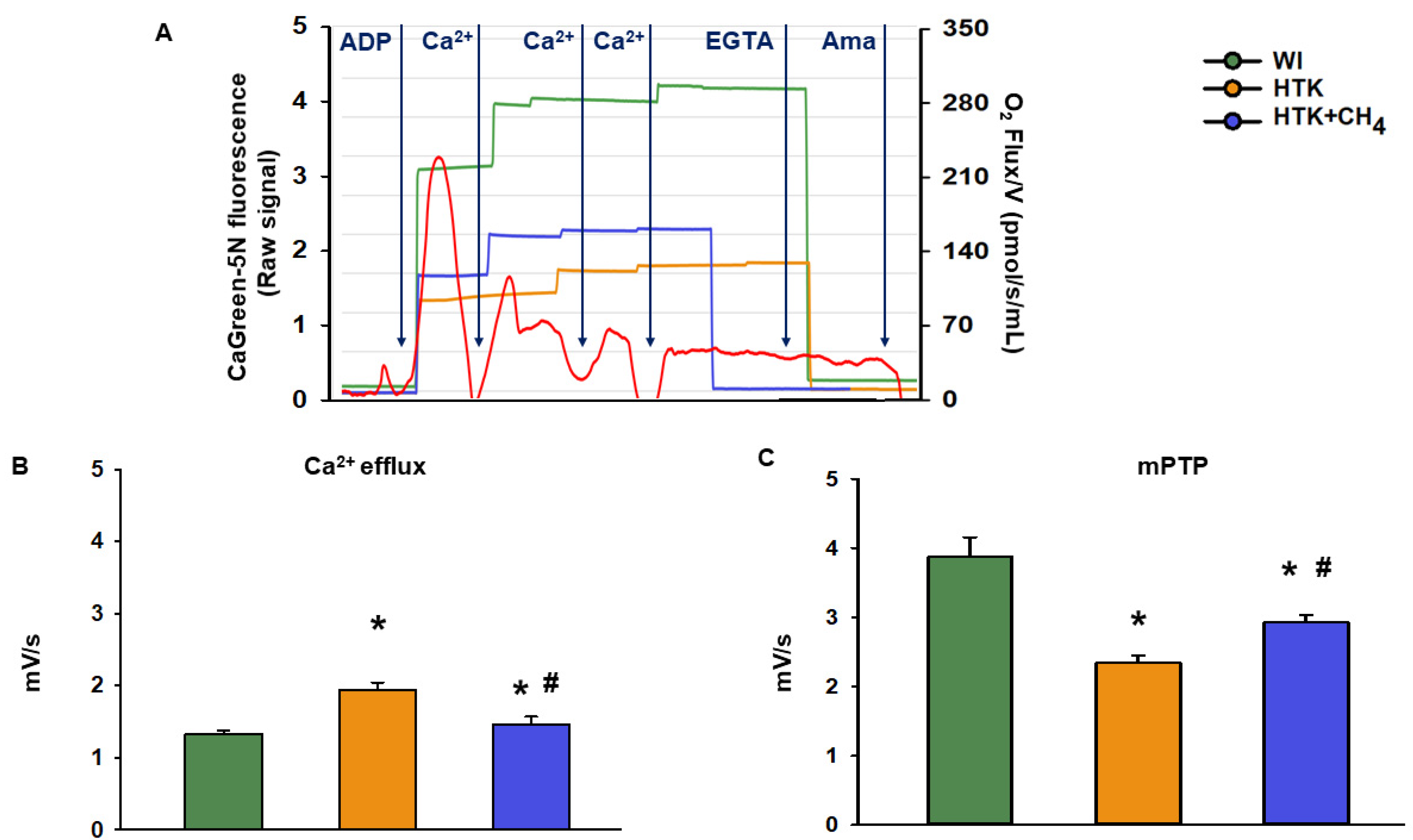
3.3. Extramitochondrial Ca2+ Movement
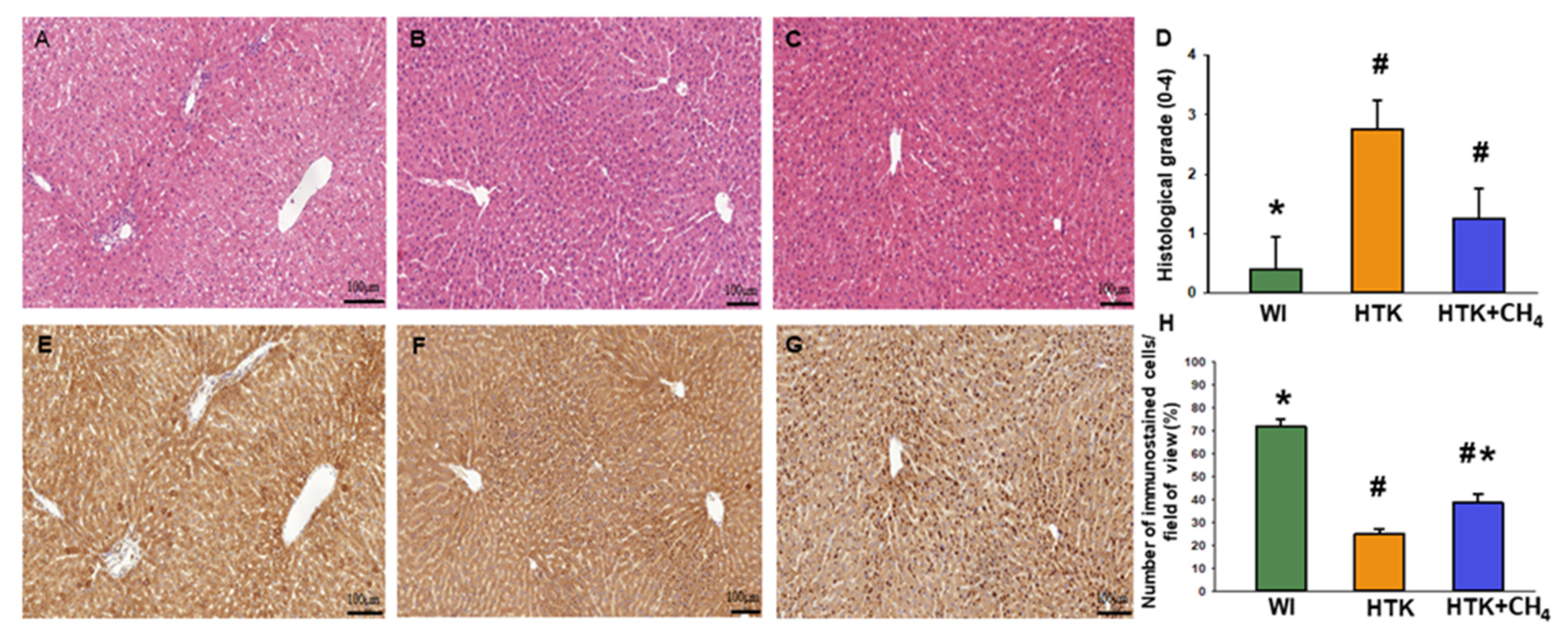
3.4. Histology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ito, T.; Naini, B.V.; Markovic, D.; Aziz, A.; Younan, S.; Lu, M.; Hirao, H.; Kadono, K.; Kojima, H.; DiNorcia, J., 3rd; et al. Ischemia-reperfusion injury and its relationship with early allograft dysfunction in liver transplant patients. Am. J. Transplant. 2021, 21, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Horvath, T.; Jasz, D.K.; Barath, B.; Poles, M.Z.; Boros, M.; Hartmann, P. Mitochondrial Consequences of Organ Preservation Techniques during Liver Transplantation. Int. J. Mol. Sci. 2021, 22, 2816. [Google Scholar] [CrossRef]
- Strifler, G.; Tuboly, E.; Szel, E.; Kaszonyi, E.; Cao, C.; Kaszaki, J.; Meszaros, A.; Boros, M.; Hartmann, P. Inhaled Methane Limits the Mitochondrial Electron Transport Chain Dysfunction during Experimental Liver Ischemia-Reperfusion Injury. PLoS ONE 2016, 11, e0146363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasz, D.K.; Szilagyi, A.L.; Tuboly, E.; Barath, B.; Marton, A.R.; Varga, P.; Varga, G.; Erces, D.; Mohacsi, A.; Szabo, A.; et al. Reduction in hypoxia-reoxygenation-induced myocardial mitochondrial damage with exogenous methane. J. Cell. Mol. Med. 2021, 25, 5113–5123. [Google Scholar] [CrossRef] [PubMed]
- Halestrap, A.P.; Clarke, S.J.; Khaliulin, I. The role of mitochondria in protection of the heart by preconditioning. Biochim. Biophys. Acta 2007, 1767, 1007–1031. [Google Scholar] [CrossRef] [Green Version]
- Hassanpour, S.H.; Dehghani, M.A.; Karami, S.Z. Study of respiratory chain dysfunction in heart disease. J. Cardiovasc. Thorac. Res. 2018, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Drose, S.; Brandt, U.; Wittig, I. Mitochondrial respiratory chain complexes as sources and targets of thiol-based redox-regulation. Biochim. Biophys. Acta 2014, 1844, 1344–1354. [Google Scholar] [CrossRef]
- Benke, K.; Jasz, D.K.; Szilagyi, A.L.; Barath, B.; Tuboly, E.; Marton, A.R.; Varga, P.; Mohacsi, A.; Szabo, A.; Szell, Z.; et al. Methane supplementation improves graft function in experimental heart transplantation. J. Heart Lung Transplant. Off. Publ. Int. Soc. Heart Transplant. 2021, 40, 183–192. [Google Scholar] [CrossRef]
- Kalogeris, T.; Baines, C.P.; Krenz, M.; Korthuis, R.J. Cell biology of ischemia/reperfusion injury. Int. Rev. Cell Mol. Biol. 2012, 298, 229–317. [Google Scholar] [CrossRef] [Green Version]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Aksentijevic, D.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; Smith, A.C.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef]
- Martins, R.M.; Teodoro, J.S.; Furtado, E.; Oliveira, R.C.; Tralhao, J.G.; Rolo, A.P.; Palmeira, C.M. Mild hypothermia during the reperfusion phase protects mitochondrial bioenergetics against ischemia-reperfusion injury in an animal model of ex-vivo liver transplantation-an experimental study. Int. J. Med. Sci. 2019, 16, 1304–1312. [Google Scholar] [CrossRef] [Green Version]
- Ferng, A.S.; Schipper, D.; Connell, A.M.; Marsh, K.M.; Knapp, S.; Khalpey, Z. Novel vs clinical organ preservation solutions: Improved cardiac mitochondrial protection. J. Cardiothorac. Surg. 2017, 12, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeb-Parsy, K.; Martin, J.L.; Summers, D.M.; Watson, C.J.E.; Krieg, T.; Murphy, M.P. Mitochondria as Therapeutic Targets in Transplantation. Trends Mol. Med. 2021, 27, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Bardallo, R.G.; Da Silva, R.T.; Carbonell, T.; Palmeira, C.; Folch-Puy, E.; Rosello-Catafau, J.; Adam, R.; Panisello-Rosello, A. Liver Graft Hypothermic Static and Oxygenated Perfusion (HOPE) Strategies: A Mitochondrial Crossroads. Int. J. Mol. Sci. 2022, 23, 5742. [Google Scholar] [CrossRef] [PubMed]
- Dugbartey, G.J. Carbon monoxide as an emerging pharmacological tool to improve lung and liver transplantation protocols. Biochem. Pharm. 2021, 193, 114752. [Google Scholar] [CrossRef]
- Kobayashi, E.; Sano, M. Organ preservation solution containing dissolved hydrogen gas from a hydrogen-absorbing alloy canister improves function of transplanted ischemic kidneys in miniature pigs. PLoS ONE 2019, 14, e0222863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boros, M.; Ghyczy, M.; Erces, D.; Varga, G.; Tokes, T.; Kupai, K.; Torday, C.; Kaszaki, J. The anti-inflammatory effects of methane. Crit. Care Med. 2012, 40, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Chen, O.; Zhang, R.; Nakao, A.; Fan, D.; Zhang, T.; Gu, Z.; Tao, H.; Sun, X. Methane Attenuates Hepatic Ischemia/Reperfusion Injury in Rats Through Antiapoptotic, Anti-Inflammatory, and Antioxidative Actions. Shock 2015, 44, 181–187. [Google Scholar] [CrossRef]
- He, R.; Wang, L.; Zhu, J.; Fei, M.; Bao, S.; Meng, Y.; Wang, Y.; Li, J.; Deng, X. Methane-rich saline protects against concanavalin A-induced autoimmune hepatitis in mice through anti-inflammatory and anti-oxidative pathways. Biochem. Biophys. Res. Commun. 2016, 470, 22–28. [Google Scholar] [CrossRef]
- Srinivasan, P.K.; Yagi, S.; Doorschodt, B.; Nagai, K.; Afify, M.; Uemoto, S.; Tolba, R. Impact of venous systemic oxygen persufflation supplemented with nitric oxide gas on cold-stored, warm ischemia-damaged experimental liver grafts. Liver Transplant. 2012, 18, 219–225. [Google Scholar] [CrossRef]
- Minor, T.; Akbar, S.; Tolba, R.; Dombrowski, F. Cold preservation of fatty liver grafts: Prevention of functional and ultrastructural impairments by venous oxygen persufflation. J. Hepatol. 2000, 32, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Otto-Slusarczyk, D.; Grabon, W.; Mielczarek-Puta, M. Aspartate aminotransferase--key enzyme in the human systemic metabolism. Postep. Hig. Med. Dosw. Online 2016, 70, 219–230. [Google Scholar] [CrossRef]
- Naszai, A.; Terhes, E.; Kaszaki, J.; Boros, M.; Juhasz, L. Ca2+N It Be Measured? Detection of Extramitochondrial Calcium Movement With High-Resolution FluoRespirometry. Sci. Rep. 2019, 9, 19229. [Google Scholar] [CrossRef] [Green Version]
- t Hart, N.A.; van der Plaats, A.; Leuvenink, H.G.; Wiersema-Buist, J.; Olinga, P.; van Luyn, M.J.; Verkerke, G.J.; Rakhorst, G.; Ploeg, R.J. Initial blood washout during organ procurement determines liver injury and function after preservation and reperfusion. Am. J. Transplant. 2004, 4, 1836–1844. [Google Scholar] [CrossRef]
- Lambert, A.J.; Brand, M.D. Inhibitors of the quinone-binding site allow rapid superoxide production from mitochondrial NADH:ubiquinone oxidoreductase (complex I). J. Biol. Chem. 2004, 279, 39414–39420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teoh, N.C. Hepatic ischemia reperfusion injury: Contemporary perspectives on pathogenic mechanisms and basis for hepatoprotection-the good, bad and deadly. J. Gastroenterol. Hepatol. 2011, 26 (Suppl. S1), 180–187. [Google Scholar] [CrossRef]
- Nardo, B.; Bertelli, R.; Montalti, R.; Beltempo, P.; Puviani, L.; Pacile, V.; Cavallari, A. Preliminary results of a clinical randomized study comparing Celsior and HTK solutions in liver preservation for transplantation. Transplant. Proc. 2005, 37, 320–322. [Google Scholar] [CrossRef]
- Cavallari, A.; Cillo, U.; Nardo, B.; Filipponi, F.; Gringeri, E.; Montalti, R.; Vistoli, F.; D’Amico, F.; Faenza, A.; Mosca, F.; et al. A multicenter pilot prospective study comparing Celsior and University of Wisconsin preserving solutions for use in liver transplantation. Liver Transplant. 2003, 9, 814–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakoyun, R.; Romano, A.; Nordstrom, J.; Ericzon, B.G.; Nowak, G. Type of Preservation Solution, UW or HTK, Has an Impact on the Incidence of Biliary Stricture following Liver Transplantation: A Retrospective Study. J. Transplant. 2019, 2019, 8150736. [Google Scholar] [CrossRef]
- Pokorny, H.; Rasoul-Rockenschaub, S.; Langer, F.; Windhager, T.; Rosenstingl, A.; Lange, R.; Konigsrainer, A.; Ringe, B.; Muhlbacher, F.; Steininger, R. Histidine-tryptophan-ketoglutarate solution for organ preservation in human liver transplantation-a prospective multi-centre observation study. Transpl. Int. 2004, 17, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Erhard, J.; Lange, R.; Scherer, R.; Kox, W.J.; Bretschneider, H.J.; Gebhard, M.M.; Eigler, F.W. Comparison of histidine-tryptophan-ketoglutarate (HTK) solution versus University of Wisconsin (UW) solution for organ preservation in human liver transplantation. A prospective, randomized study. Transpl. Int. 1994, 7, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Gubernatis, G.; Pichlmayr, R.; Lamesch, P.; Grosse, H.; Bornscheuer, A.; Meyer, H.J.; Ringe, B.; Farle, M.; Bretschneider, H.J. HTK-solution (Bretschneider) for human liver transplantation. First clinical experiences. Langenbecks Arch. Chir. 1990, 375, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Gasotransmitters: Growing pains and joys. Trends Biochem. Sci. 2014, 39, 227–232. [Google Scholar] [CrossRef]
- Carlisle, S.M.; Burchart, P.A.; McCauley, C.; Surette, R.A. Biokinetics of inhaled radioactive methane in rats: A pilot study. Appl. Radiat. Isot. Incl. Data Instrum. Methods Use Agric. Ind. Med. 2005, 62, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Boros, M.; Keppler, F. Methane Production and Bioactivity-A Link to Oxido-Reductive Stress. Front. Physiol. 2019, 10, 1244. [Google Scholar] [CrossRef] [Green Version]
- Juhasz, L.; Tallosy, S.P.; Naszai, A.; Varga, G.; Erces, D.; Boros, M. Bioactivity of Inhaled Methane and Interactions With Other Biological Gases. Front. Cell Dev. Biol. 2021, 9, 824749. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, A.R.; Andersen, K.J.; Hamilton-Dutoit, S.; Nyengaard, J.R.; Mortensen, F.V. Correlation between liver cell necrosis and circulating alanine aminotransferase after ischaemia/reperfusion injuries in the rat liver. Int. J. Exp. Pathol. 2016, 97, 133–138. [Google Scholar] [CrossRef]
- Marzi, I.; Walcher, F.; Menger, M.; Buhren, V.; Harbauer, G.; Trentz, O. Microcirculatory disturbances and leucocyte adherence in transplanted livers after cold storage in Euro-Collins, UW and HTK solutions. Transpl. Int. 1991, 4, 45–50. [Google Scholar] [CrossRef]
- Ghyczy, M.; Torday, C.; Kaszaki, J.; Szabo, A.; Czobel, M.; Boros, M. Hypoxia-induced generation of methane in mitochondria and eukaryotic cells: An alternative approach to methanogenesis. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2008, 21, 251–258. [Google Scholar] [CrossRef]
- Poles, M.Z.; Bodi, N.; Bagyanszki, M.; Fekete, E.; Meszaros, A.T.; Varga, G.; Szucs, S.; Naszai, A.; Kiss, L.; Kozlov, A.V.; et al. Reduction of nitrosative stress by methane: Neuroprotection through xanthine oxidoreductase inhibition in a rat model of mesenteric ischemia-reperfusion. Free. Radic. Biol. Med. 2018, 120, 160–169. [Google Scholar] [CrossRef]
- Dunne, J.B.; Davenport, M.; Williams, R.; Tredger, J.M. Evidence that S-adenosylmethionine and N-acetylcysteine reduce injury from sequential cold and warm ischemia in the isolated perfused rat liver. Transplantation 1994, 57, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Bowers, B.A.; Branum, G.D.; Rotolo, F.S.; Watters, C.R.; Meyers, W.C. Bile flow—An index of ischemic injury. J. Surg. Res. 1987, 42, 565–569. [Google Scholar] [CrossRef]
- Clavien, P.A.; Harvey, P.R.; Strasberg, S.M. Preservation and reperfusion injuries in liver allografts. An overview and synthesis of current studies. Transplantation 1992, 53, 957–978. [Google Scholar] [CrossRef]
- Parajuli, N.; MacMillan-Crow, L.A. Role of reduced manganese superoxide dismutase in ischemia-reperfusion injury: A possible trigger for autophagy and mitochondrial biogenesis? Am. J. Physiol. Renal. Physiol. 2013, 304, F257–F267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parajuli, N.; Shrum, S.; Tobacyk, J.; Harb, A.; Arthur, J.M.; MacMillan-Crow, L.A. Renal cold storage followed by transplantation impairs expression of key mitochondrial fission and fusion proteins. PLoS ONE 2017, 12, e0185542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Wang, Q.; Wang, H.; Guan, W. Tumor necrosis factor-alpha preconditioning attenuates liver ischemia/reperfusion injury through preserving sarco/endoplasmic reticulum calcium-ATPase function. J. Surg. Res. 2013, 184, 1109–1113. [Google Scholar] [CrossRef]
- Halestrap, A.P. Calcium, mitochondria and reperfusion injury: A pore way to die. Biochem. Soc. Trans. 2006, 34, 232–237. [Google Scholar] [CrossRef]
- Baumgartner, H.K.; Gerasimenko, J.V.; Thorne, C.; Ferdek, P.; Pozzan, T.; Tepikin, A.V.; Petersen, O.H.; Sutton, R.; Watson, A.J.; Gerasimenko, O.V. Calcium elevation in mitochondria is the main Ca2+ requirement for mitochondrial permeability transition pore (mPTP) opening. J. Biol. Chem. 2009, 284, 20796–20803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzuto, R.; Marchi, S.; Bonora, M.; Aguiari, P.; Bononi, A.; De Stefani, D.; Giorgi, C.; Leo, S.; Rimessi, A.; Siviero, R.; et al. Ca(2+) transfer from the ER to mitochondria: When, how and why. Biochim. Biophys. Acta 2009, 1787, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Palty, R.; Silverman, W.F.; Hershfinkel, M.; Caporale, T.; Sensi, S.L.; Parnis, J.; Nolte, C.; Fishman, D.; Shoshan-Barmatz, V.; Herrmann, S.; et al. NCLX is an essential component of mitochondrial Na+/Ca2+ exchange. Proc. Natl. Acad. Sci. USA 2010, 107, 436–441. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horváth, T.; Sándor, L.; Baráth, B.; Donka, T.; Baráth, B.; Mohácsi, Á.; Jász, K.D.; Hartmann, P.; Boros, M. Methane Admixture Protects Liver Mitochondria and Improves Graft Function after Static Cold Storage and Reperfusion. Antioxidants 2023, 12, 271. https://doi.org/10.3390/antiox12020271
Horváth T, Sándor L, Baráth B, Donka T, Baráth B, Mohácsi Á, Jász KD, Hartmann P, Boros M. Methane Admixture Protects Liver Mitochondria and Improves Graft Function after Static Cold Storage and Reperfusion. Antioxidants. 2023; 12(2):271. https://doi.org/10.3390/antiox12020271
Chicago/Turabian StyleHorváth, Tamara, Lilla Sándor, Bálint Baráth, Tibor Donka, Bence Baráth, Árpád Mohácsi, Kurszán Dávid Jász, Petra Hartmann, and Mihály Boros. 2023. "Methane Admixture Protects Liver Mitochondria and Improves Graft Function after Static Cold Storage and Reperfusion" Antioxidants 12, no. 2: 271. https://doi.org/10.3390/antiox12020271