

Inside the Mechanism of Action of Three Pyrazole Derivatives in Human Platelets and Endothelial Cells
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. In Vitro Antioxidant Activity (DPPH Assay)
2.3. Blood Collection and Preparative Procedures
2.4. Superoxide Anion Assay in Human Platelet
2.5. Lipid Peroxidation Measurement in Human Platelet
2.6. NADPH Oxidase Activity Assay in Human Platelet
2.7. Tests to Assay Platelet Viability in Human Platelet
2.8. Antioxidant Activity on Endothelial Cells
2.9. Aerobic Metabolism in Human Platelets and EAhy926 Cell Line
2.10. Anti-Proliferative Activity
2.11. Statistical Analysis
3. Results
3.1. In Vitro Antioxidant Activity (DPPH)
3.2. Human Platelets Oxidative Status
3.3. Tests to Assay Platelet Viability
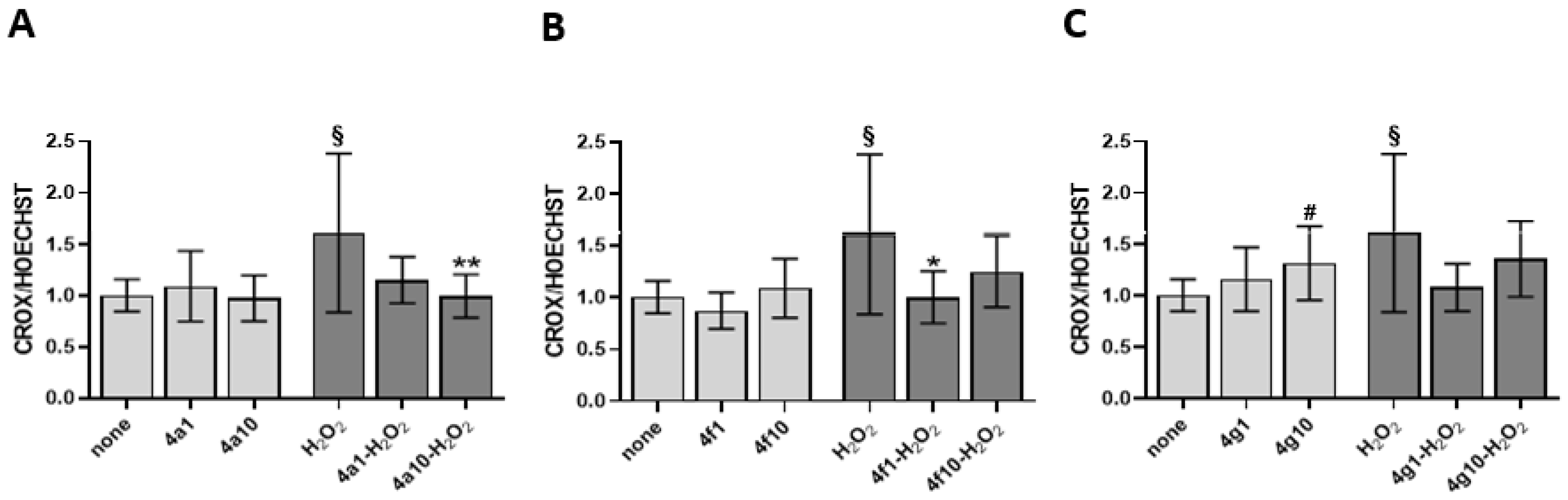
3.4. Antioxidant Activity on Endothelial Cells
3.5. Aerobic Metabolism in Platelets
3.6. Aerobic Metabolism in EA.hy926 Cell Line
3.7. Antiproliferative Activity Evaluation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Finkel, T. Signal Transduction by Reactive Oxygen Species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.I.; Griendling, K.K. Regulation of Signal Transduction by Reactive Oxygen Species in the Cardiovascular System. Circ. Res. 2015, 116, 531–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Q.; Wang, J.Q.; Assaraf, Y.G.; Ren, L.; Gupta, P.; Wei, L.; Ashby, C.R.; Yang, D.H.; Chen, Z.S. Modulating ROS to Overcome Multidrug Resistance in Cancer. Drug Resist. Updates 2018, 41, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Boveris, A.; Chance, B. The Mitochondrial Generation of Hydrogen Peroxide. General Properties and Effect of Hyperbaric Oxygen. Biochem. J. 1973, 134, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Oswald, M.C.W.; Garnham, N.; Sweeney, S.T.; Landgraf, M. Regulation of Neuronal Development and Function by ROS. FEBS Lett. 2018, 592, 679–691. [Google Scholar] [CrossRef]
- Hansson, G.K.; Libby, P.; Tabas, I. Inflammation and Plaque Vulnerability. J. Intern. Med. 2015, 278, 483–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B. Diabetes, Oxidative Stress, and Antioxidants: A Review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef]
- Chen, X.; Guo, C.; Kong, J. Oxidative Stress in Neurodegenerative Diseases. Neural Regen. Res. 2012, 7, 376–385. [Google Scholar] [CrossRef]
- Haigis, M.C.; Yankner, B.A. The Aging Stress Response. Mol. Cell 2010, 40, 333–344. [Google Scholar] [CrossRef]
- Hoffmann, M.H.; Griffiths, H.R. The Dual Role of Reactive Oxygen Species in Autoimmune and Inflammatory Diseases: Evidence from Preclinical Models. Free Radic. Biol. Med. 2018, 125, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Herb, M.; Schramm, M. Functions of ROS in Macrophages and Antimicrobial Immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in Cancer Therapy: The Bright Side of the Moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Sierko, E.; Wojtukiewicz, M.Z. Platelets and Angiogenesis in Malignancy. Semin. Thromb. Hemost. 2004, 30, 95–108. [Google Scholar] [CrossRef]
- Iuliano, L.; Colavita, A.R.; Leo, R.; Praticò, D.; Violi, F. Oxygen Free Radicals and Platelet Activation. Free Radic. Biol. Med. 1997, 22, 999–1006. [Google Scholar] [CrossRef]
- Leoncini, G.; Maresca, M.; Colao, C. Oxidative Metabolism of Human Platelets. Biochem. Int. 1991, 25, 647–655. [Google Scholar]
- Maresca, M.; Colao, C.; Leoncini, G. Generation of Hydrogen Peroxide in Resting and Activated Platelets. Cell Biochem. Funct. 1992, 10, 79–85. [Google Scholar] [CrossRef]
- Pratico, D.; Iuliano, L.; Pulcinelli, F.M.; Bonavita, M.S.; Gazzaniga, P.P.; Violi, F. Hydrogen Peroxide Triggers Activation of Human Platelets Selectively Exposed to Nonaggregating Concentrations of Arachidonic Acid and Collagen. J. Lab. Clin. Med. 1992, 119, 364–370. [Google Scholar]
- Leo, R.; Praticò, D.; Iuliano, L.; Pulcinelli, F.M.; Ghiselli, A.; Pignatelli, P.; Colavita, A.R.; FitzGerald, G.A.; Violi, F. Platelet Activation by Superoxide Anion and Hydroxyl Radicals Intrinsically Generated by Platelets That Had Undergone Anoxia and Then Reoxygenated. Circulation 1997, 95, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Krötz, F.; Sohn, H.Y.; Gloe, T.; Zahler, S.; Riexinger, T.; Schiele, T.M.; Becker, B.F.; Theisen, K.; Klauss, V.; Pohl, U. NAD(P)H Oxidase-Dependent Platelet Superoxide Anion Release Increases Platelet Recruitment. Blood 2002, 100, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Brullo, C.; Massa, M.; Rapetti, F.; Alfei, S.; Bertolotto, M.B.; Montecucco, F.; Signorello, M.G.; Bruno, O. New Hybrid Pyrazole and Imidazopyrazole Antinflammatory Agents Able to Reduce ROS Production in Different Biological Targets. Molecules 2020, 25, 899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielnik, M.B.; Olsen, E.; Vogt, G.; Adeline, D.; Skrede, G. Grape Seed Extract as Antioxidant in Cooked, Cold Stored Turkey Meat. LWT—Food Sci. Technol. 2006, 39, 191–198. [Google Scholar] [CrossRef]
- Russo, E.; Spallarossa, A.; Comite, A.; Pagliero, M.; Guida, P.; Belotti, V.; Caviglia, D.; Schito, A.M. Valorization and Potential Antimicrobial Use of Olive Mill Wastewater (OMW) from Italian Olive Oil Production. Antioxidants 2022, 11, 903. [Google Scholar] [CrossRef]
- Leoncini, G.; Maresca, M.; Colao, C.; Piana, A.; Armani, U. Increased Hydrogen Peroxide Formation in Platelets of Patients Affected with Essential Thrombocythaemia (ET). Blood Coagul. Fibrinolysis 1992, 3, 271–277. [Google Scholar] [CrossRef]
- Olas, B.; Kontek, B.; Malinowska, P.; Żuchowski, J.; Stochmal, A. Hippophae rhamnoides L. Fruits Reduce the Oxidative Stress in Human Blood Platelets and Plasma. Oxid. Med. Cell. Longev. 2016, 2016, 4692486. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Hohorst, H.-J. L-(+)-Lactate. In Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 1965; pp. 266–277. [Google Scholar]
- Vassault, A. Lactate Dehydrogenase. In Methods of Enzymatic Analysis Vol III; Bergmeyer, H., Ed.; Verlag Chemie: Weinheim, Germany, 1983; pp. 277–282. [Google Scholar]
- Altieri, P.; Murialdo, R.; Barisione, C.; Lazzarini, E.; Garibaldi, S.; Fabbi, P.; Ruggeri, C.; Borile, S.; Carbone, F.; Armirotti, A.; et al. 5-Fluorouracil Causes Endothelial Cell Senescence: Potential Protective Role of Glucagon-like Peptide 1. Br. J. Pharmacol. 2017, 174, 3713–3726. [Google Scholar] [CrossRef] [Green Version]
- Ravera, S.; Signorello, M.G.; Bartolucci, M.; Ferrando, S.; Manni, L.; Caicci, F.; Calzia, D.; Panfoli, I.; Morelli, A.; Leoncini, G. Extramitochondrial Energy Production in Platelets. Biol. Cell 2018, 110, 97–108. [Google Scholar] [CrossRef]
- Signorello, M.G.; Ravera, S.; Leoncini, G. Lectin-Induced Oxidative Stress in Human Platelets. Redox Biol. 2020, 32, 101456. [Google Scholar] [CrossRef] [PubMed]
- Hinkle, P.C. P/O Ratios of Mitochondrial Oxidative Phosphorylation. Biochim. Biophys. Acta 2005, 1706, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ravera, S.; Vigliarolo, T.; Bruno, S.; Morandi, F.; Marimpietri, D.; Sabatini, F.; Dagnino, M.; Petretto, A.; Bartolucci, M.; Muraca, M.; et al. Identification of Biochemical and Molecular Markers of Early Aging in Childhood Cancer Survivors. Cancers 2021, 13, 5214. [Google Scholar] [CrossRef] [PubMed]
- Coyle, C.H.; Martinez, L.J.; Coleman, M.C.; Spitz, D.R.; Weintraub, N.L.; Kader, K.N. Mechanisms of H2O2-Induced Oxidative Stress in Endothelial Cells. Free Radic. Biol. Med. 2006, 40, 2206–2213. [Google Scholar] [CrossRef] [Green Version]
- Holmström, K.M.; Finkel, T. Cellular Mechanisms and Physiological Consequences of Redox-Dependent Signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, H.; Halliwell, B. Damage to DNA by Reactive Oxygen and Nitrogen Species: Role in Inflammatory Disease and Progression to Cancer. Biochem. J. 1996, 313, 17–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisen, J.A.; Hanawalt, P.C. A Phylogenomic Study of DNA Repair Genes, Proteins, and Processes. Mutat. Res.—DNA Repair 1999, 435, 171–213. [Google Scholar] [CrossRef] [Green Version]
- Olinski, R.; Gackowski, D.; Foksinski, M.; Rozalski, R.; Roszkowski, K.; Jaruga, P. Oxidative DNA Damage: Assessment of the Role in Carcinogenesis, Atherosclerosis, and Acquired Immunodeficiency Syndrome. Free Radic. Biol. Med. 2002, 33, 192–200. [Google Scholar] [CrossRef]
- Roos, W.P.; Kaina, B. DNA Damage-Induced Cell Death by Apoptosis. Trends Mol. Med. 2006, 12, 440–450. [Google Scholar] [CrossRef]
- Vara, D.; Campanella, M.; Pula, G. The Novel NOX Inhibitor 2-Acetylphenothiazine Impairs Collagen-Dependent Thrombus Formation in a GPVI-Dependent Manner. Br. J. Pharmacol. 2013, 168, 212. [Google Scholar] [CrossRef]
- Delaney, M.K.; Kim, K.; Estevez, B.; Xu, Z.; Stojanovic-Terpo, A.; Shen, B.; Ushio-Fukai, M.; Cho, J.; Du, X. Differential Roles of the NADPH-Oxidase 1 and 2 in Platelet Activation and Thrombosis. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 846–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehinger, J.K.; Piel, S.; Ford, R.; Karlsson, M.; Sjövall, F.; Frostner, E.Å.; Morota, S.; Taylor, R.W.; Turnbull, D.M.; Cornell, C.; et al. Cell-Permeable Succinate Prodrugs Bypass Mitochondrial Complex I Deficiency. Nat. Commun. 2016, 7, 12317. [Google Scholar] [CrossRef] [PubMed]
- Aree, T.; Jongrungruangchok, S. Structure–Antioxidant Activity Relationship of β-Cyclodextrin Inclusion Complexes with Olive Tyrosol, Hydroxytyrosol and Oleuropein: Deep Insights from X-Ray Analysis, DFT Calculation and DPPH Assay. Carbohydr. Polym. 2018, 199, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Poljsak, B.; Šuput, D.; Milisav, I. Achieving the Balance between ROS and Antioxidants: When to Use the Synthetic Antioxidants. Oxid. Med. Cell. Longev. 2013, 2013, 956792. [Google Scholar] [CrossRef]
- Mattson, M.P. Hormesis Defined. Ageing Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Panfoli, I.; Puddu, A.; Bertola, N.; Ravera, S.; Maggi, D. The Hormetic Effect of Metformin: “Less Is More”? Int. J. Mol. Sci. 2021, 22, 6297. [Google Scholar] [CrossRef]
- Ravera, S.; Cossu, V.; Tappino, B.; Nicchia, E.; Dufour, C.; Cavani, S.; Sciutto, A.; Bolognesi, C.; Columbaro, M.; Degan, P.; et al. Concentration-Dependent Metabolic Effects of Metformin in Healthy and Fanconi Anemia Lymphoblast Cells. J. Cell. Physiol. 2018, 233, 1736–1751. [Google Scholar] [CrossRef] [PubMed]
- Son, T.G.; Camandola, S.; Mattson, M.P. Hormetic Dietary Phytochemicals. Neuromol. Med. 2008, 10, 236–246. [Google Scholar] [CrossRef] [Green Version]
- Brullo, C.; Rapetti, F.; Abbate, S.; Prosdocimi, T.; Torretta, A.; Semrau, M.; Massa, M.; Alfei, S.; Storici, P.; Parisini, E.; et al. Design, Synthesis, Biological Evaluation and Structural Characterization of Novel GEBR Library PDE4D Inhibitors. Eur. J. Med. Chem. 2021, 223, 113638. [Google Scholar] [CrossRef]
- Favot, L.; Keravis, T.; Holl, V.; Le Bec, A.; Lugnier, C. VEGF-Induced HUVEC Migration and Proliferation Are Decreased by PDE2 and PDE4 Inhibitors. Thromb. Haemost. 2003, 90, 334–343. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cmpd. | Aggregation Inhibition IC50 (µM) | ROS Production Inhibition IC50 (µM) |
|---|---|---|
| 4a | 0.5 | 2.1 |
| 4f | 1.7 | 2.2 |
| 4g | 1.2 | 2.6 |
| Cmpd. | DPPH% * | AA% |
|---|---|---|
| 4a | 90.3 ± 0.4 | 9.8 ± 0.4 |
| 4f | 90.3 ± 0.3 | 9.7 ± 0.3 |
| 4g | 81.9 ± 0.4 | 18.1 ± 0.4 |
| Cmpd. | Superoxide Anion IC50 (µM) | Lipid Peroxidation IC50 (µM) | NADPH Oxidase Activity IC50 (µM) |
|---|---|---|---|
| 4a | 13.4 ± 0.4 | 12.3 ± 0.6 | 12.6 ± 0.5 |
| 4f | 21.8 ± 0.7 | 20.5 ± 0.5 | 20.7 ± 0.4 |
| 4g | 24.6 ± 0.6 | 23.8 ± 0.6 | 24.4 ± 0.6 |
| Cmpd. | Cancer Cell Lines | Cell Growth Percent (%) | |
|---|---|---|---|
| 4a | Non small cell lung cancer | HOP-92 | 74.0 |
| NCI-H460 | 75.0 | ||
| CNS cancer | SNB-75 | 74.0 | |
| Renal cancer | UO-31 | 68.0 | |
| Leukaemia | RPMI-8226 | 49.0 | |
| Colon Cancer | HCT-15 | 47.0 | |
| 4f | CNS cancer | SNR-75 | 50.5 |
| Melanoma | SKMEL-5 | 37.6 | |
| Renal cancer | UO-31 | 45.6 | |
| Breast cancer | HS578T | 47.3 | |
| Leukaemia | CCRF-CEM | 18.5 | |
| MOLT-4 | 17.5 | ||
| K-562 | 27.3 | ||
| RPMI-8226 | 13.4 | ||
| SR | 40.3 | ||
| Non-Small cell lung cancer | A549/ATCC | 45.8 | |
| Melanoma | LOXIMVI | 21.1 | |
| SK-MEL5 | 22.5 | ||
| UACC-62 | 23.9 | ||
| Ovarian cancer | IGROV-1 | 46.7 | |
| 4g | OVCAR-3 | 34.9 | |
| OVCAR-4 | 28.8 | ||
| OVCAR-8 | 16.6 | ||
| NCI/ADR-RES | 37.3 | ||
| Renal cancer | ACHN | 48.9 | |
| CAKI-1 | 35.0 | ||
| SN12C | 44.3 | ||
| UO-31 | 38.3 | ||
| Prostate cancer | PC-3 | 20.4 | |
| DU-145 | 50.2 | ||
| Breast Cancer | MCF-7 | 30.3 | |
| HST578T | 22.2 | ||
| BT-549 | 33.8 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brullo, C.; Russo, E.; Garibaldi, S.; Altieri, P.; Ameri, P.; Ravera, S.; Signorello, M.G. Inside the Mechanism of Action of Three Pyrazole Derivatives in Human Platelets and Endothelial Cells. Antioxidants 2023, 12, 216. https://doi.org/10.3390/antiox12020216
Brullo C, Russo E, Garibaldi S, Altieri P, Ameri P, Ravera S, Signorello MG. Inside the Mechanism of Action of Three Pyrazole Derivatives in Human Platelets and Endothelial Cells. Antioxidants. 2023; 12(2):216. https://doi.org/10.3390/antiox12020216
Chicago/Turabian StyleBrullo, Chiara, Eleonora Russo, Silvano Garibaldi, Paola Altieri, Pietro Ameri, Silvia Ravera, and Maria Grazia Signorello. 2023. "Inside the Mechanism of Action of Three Pyrazole Derivatives in Human Platelets and Endothelial Cells" Antioxidants 12, no. 2: 216. https://doi.org/10.3390/antiox12020216