
Enhancement of Antioxidant and Anti-Glycation Properties of Beeswax Alcohol in Reconstituted High-Density Lipoprotein: Safeguarding against Carboxymethyllysine Toxicity in Zebrafish


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation of Human Lipoproteins by Ultracentrifugation
2.3. Human apoA-I Purification
2.4. Oxidation of LDL in the Presence of Lipid-Free BWA
2.5. Anti-Glycation Activity of Lipid-Free BWA
2.6. Synthesis of Reconstituted HDL and Electrophoresis
2.7. Transmission Electron Microscopic (TEM) Analysis of Reconstituted HDL
2.8. LDL Oxidation in the Presence of rHDL
2.9. Anti-Glycation Activity of BWA in rHDL
2.10. Protein Determination
2.11. Antioxidant Activities in the rHDL
2.12. Zebrafish Maintenance
2.13. Anti-Inflammatory Activity of Lipid-Free BWA in Zebrafish
2.14. Liver Histology
2.15. Microinjection of CML and rHDL into Zebrafish Embryos
2.16. Visulizing Oxidative Stress and, Apoptosis in the Embryo
2.17. Cutaneous Wound Formation
2.18. Visual Observation of Wound Healing
2.19. Statistical Analysis
3. Results
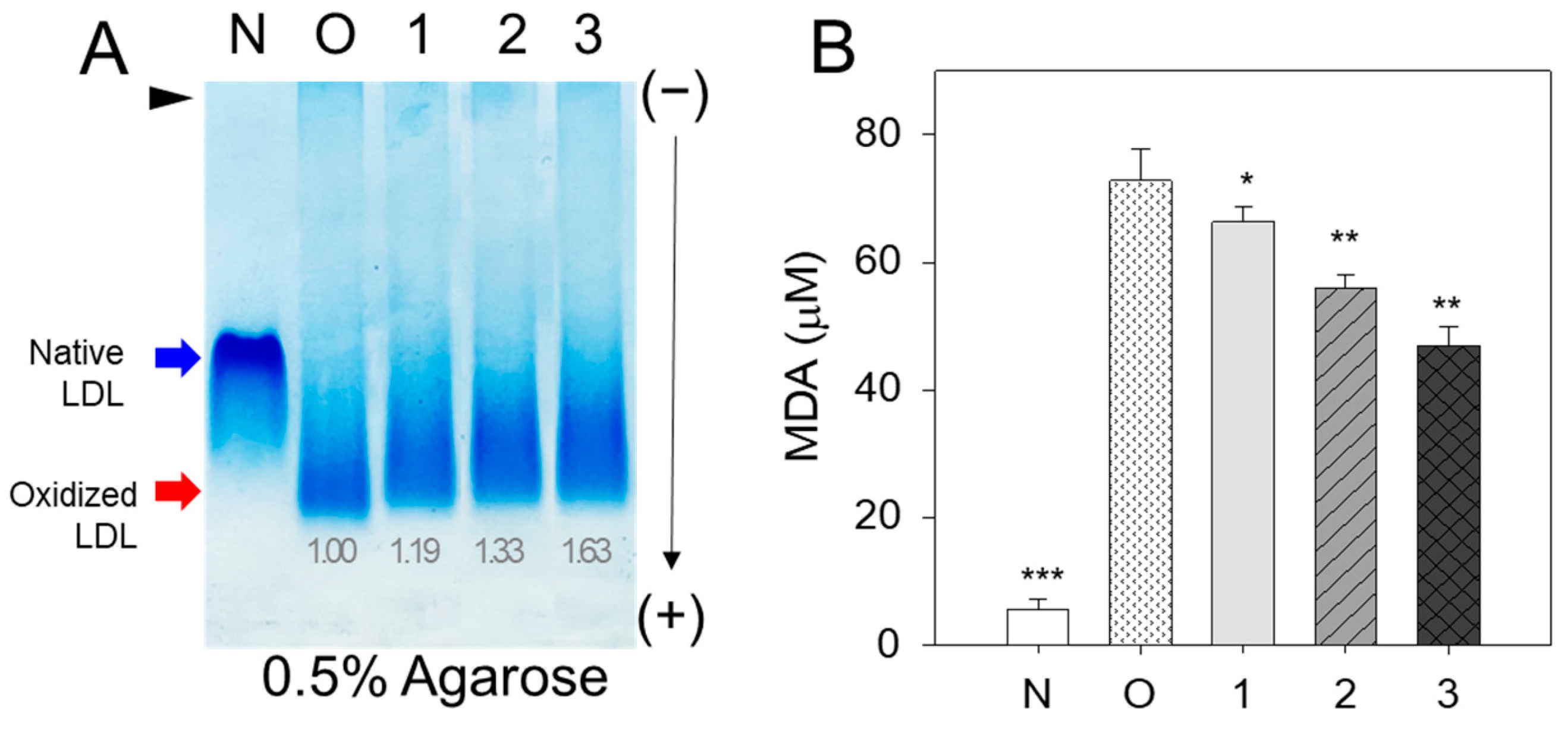
3.1. Antioxidant Ability of BWA against LDL Oxidation
3.2. Anti-Glycation Activity of BWA against CML-Induced Modification of HDL
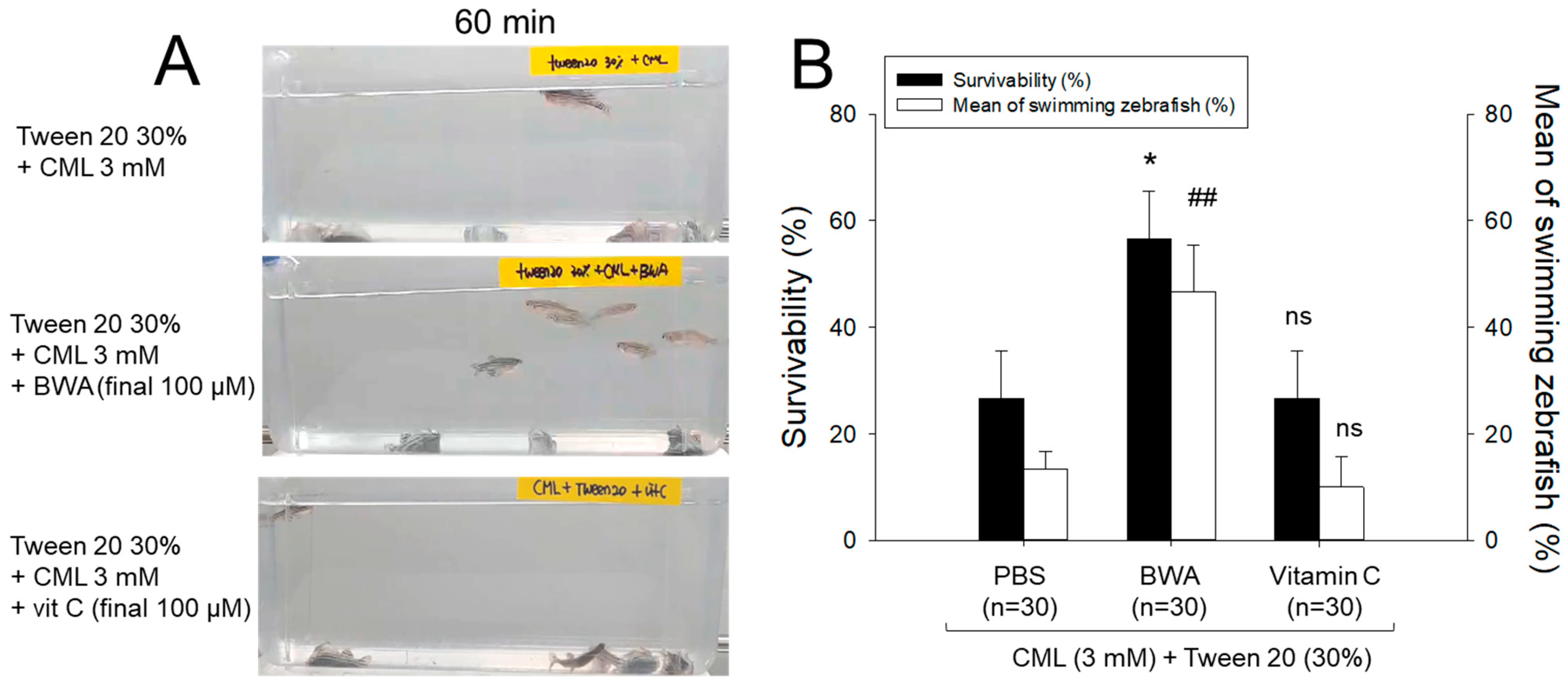
3.3. Anti-Inflammatory Activity of BWA against CML-Induced Acute Paralysis of Zebrafish
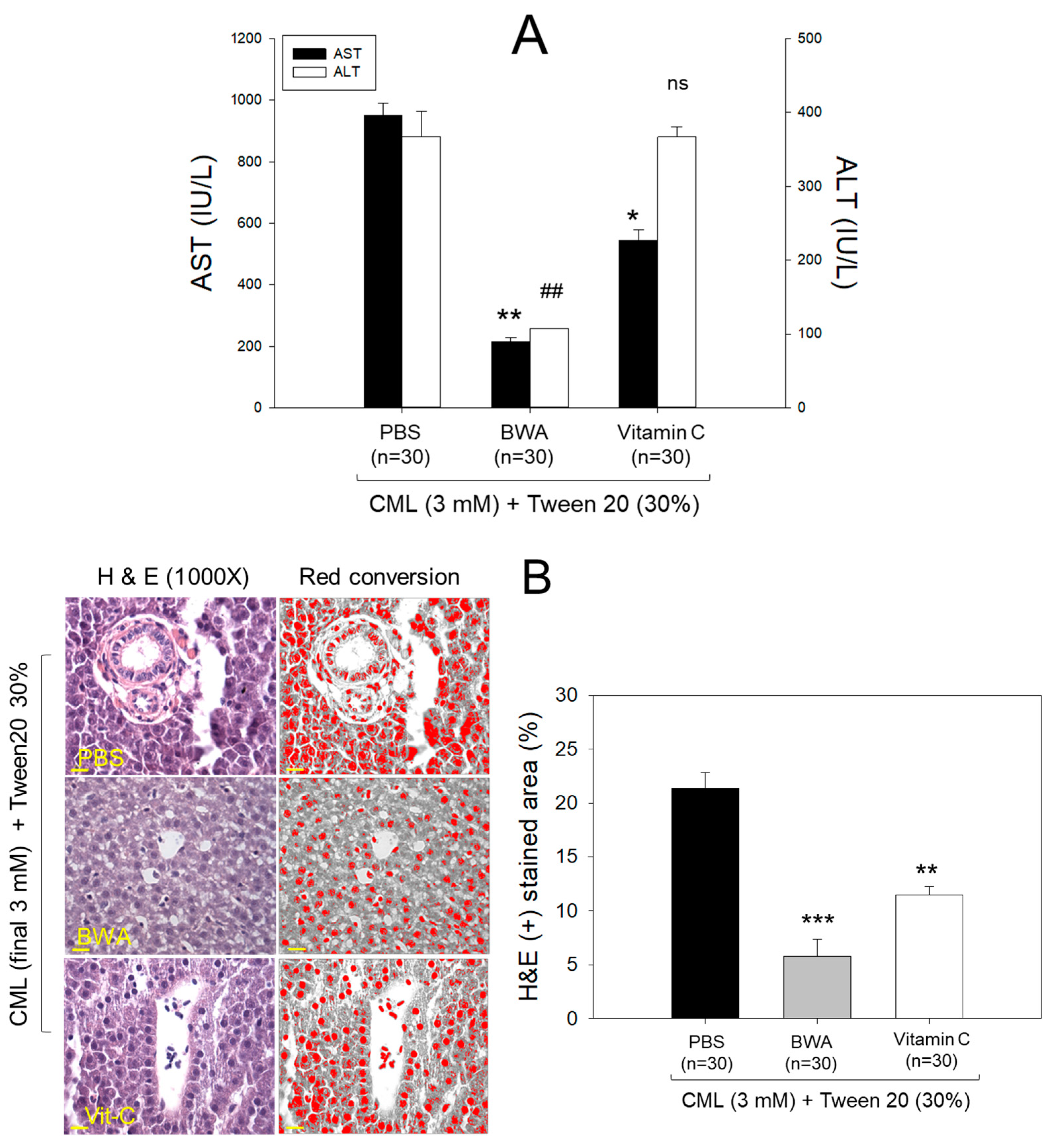
3.4. Amelioration of Hepatic Damage
3.5. Immunohistochemistry for Interleukin (IL)-6
3.6. Synthesis of Reconstituted HDL Containing BWA
3.7. Electrophoretic Profiles of rHDL Containing BWA
3.8. Electron Microscopy Observations
3.9. Anti-Glycation Activity of BWA in rHDL
3.10. Antioxidant Ability of BWA in rHDL against LDL Oxidation
3.11. Enhancement of the Antioxidant Ability of HDL by rHDL Containing BWA
3.12. Protection of Embryo Death by BWA in rHDL
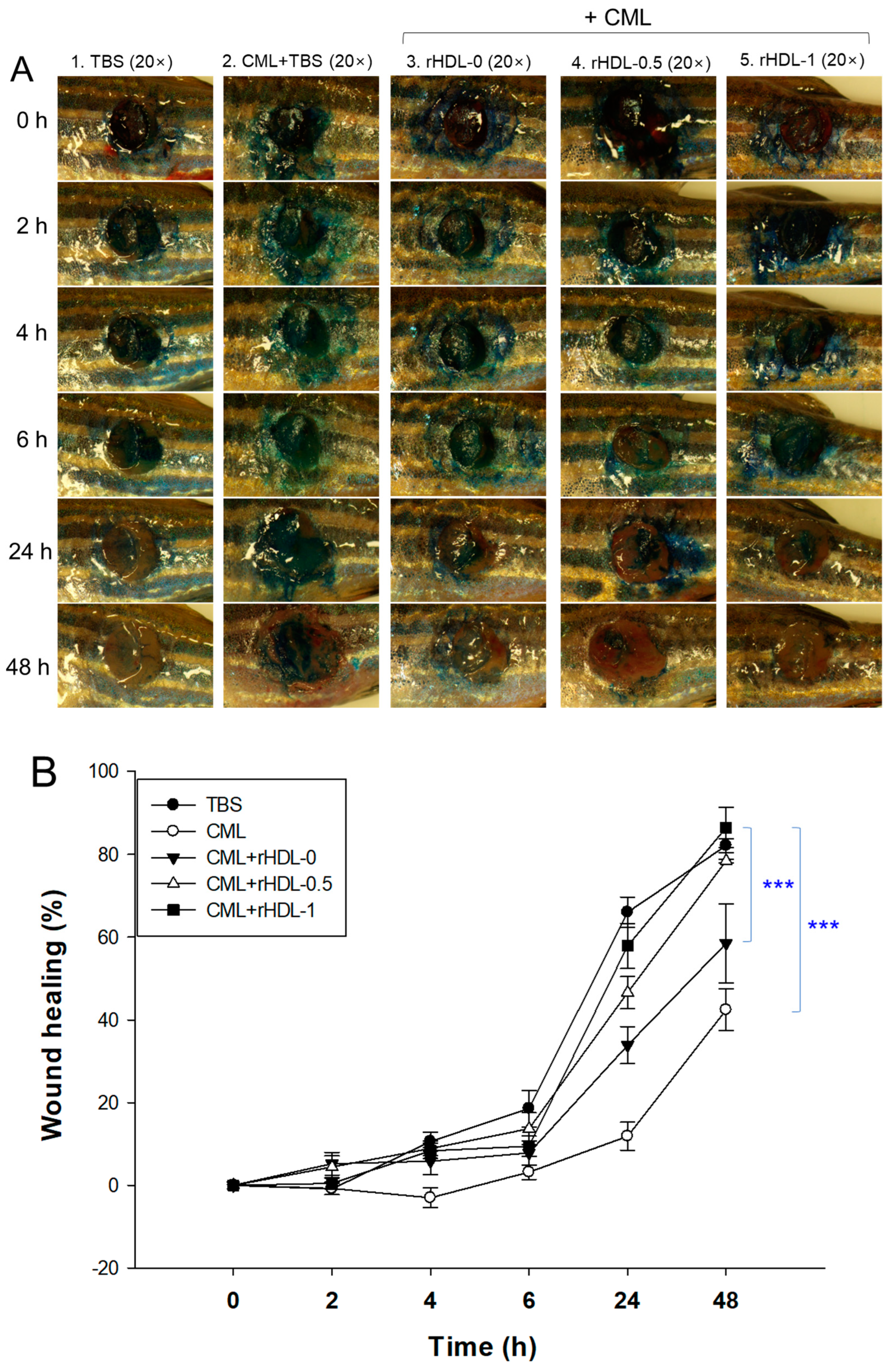
3.13. BWA in rHDL Facilitated the Healing of Cutaneous Wounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yeum, K.-J.; Russell, R.M.; Krinsky, N.I.; Aldini, G. Biomarkers of antioxidant capacity in the hydrophilic and lipophilic compartments of human plasma. Arch. Biochem. Biophys. 2004, 430, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Pinchuk, I.; Shoval, H.; Dotan, Y.; Lichtenberg, D. Evaluation of antioxidants: Scope, limitations and relevance of assays. Chem. Phys. Lipids 2012, 165, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Libardo, M.D.J.; Wang, T.-Y.; Pellois, J.-P.; Angeles-Boza, A.M. How does membrane oxidation affect cell delivery and cell killing? Trends Biotechnol. 2017, 35, 686–690. [Google Scholar] [CrossRef] [PubMed]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef]
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Oxidative stress in ageing and chronic degenerative pathologies: Molecular mechanisms involved in counteracting oxidative stress and chronic inflammation. Int. J. Mol. Sci. 2022, 23, 7273. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118. [Google Scholar] [CrossRef]
- Ravelo, Y.; Molina, V.; Carbajal, D.; Fernández, L.; Fernández, J.C.; Arruzazabala, M.L.; Más, R. Evaluation of anti-inflammatory and antinociceptive effects of D-002 (beeswax alcohols). J. Nat. Med. 2011, 65, 330–335. [Google Scholar] [CrossRef]
- Carbajal, D.; Molina, V.; Valdés, S.; Arruzazabala, M.d.L.; Más, R.; Magraner, J. Anti-inflammatory activity of D-002: An active product isolated from beeswax. Prostaglandins Leukot. Essent. Fat. Acids 1998, 59, 235–238. [Google Scholar] [CrossRef]
- Pérez, Y.; Oyárzabal, A.; Mas, R.; Molina, V.; Jiménez, S. Protective effect of D-002, a mixture of beeswax alcohols, against indomethacin-induced gastric ulcers and mechanism of action. J. Nat. Med. 2013, 67, 182–189. [Google Scholar] [CrossRef]
- Puig, M.N.; Castaño, S.M.; Ferreiro, R.M.; Clara, M.V.; Hernansez, N.M. Effects of Oral Administration of D-002 (Beeswax Alcohols) on Histological and Functional Outcomes in a Rat Model of Antigen-Induced Arthritis: Preliminary Study. Int. J. Pharmacol. Phytochem. Ethnomed. 2016, 5, 60–68. [Google Scholar] [CrossRef]
- Information on Each Health Functional Food Ingredient. Available online: https://www.foodsafetykorea.go.kr/portal/board/boardDetail.do?menu_grp=MENU_NEW01&menu_no=2660&bbs_no=bbs987&nticmatr_yn=N&bbs_typecd=01&ans_yn=N&ntctxt_no=21590 (accessed on 16 August 2023).
- Molina, V.; Valdés, S.; Carbajal, D.; Arruzazabala, M.; Menéndez, R.; Más, R. Antioxidant effects of D-002 on gastric mucosa of rats with injury induced experimentally. J Med. Food 2001, 4, 79–84. [Google Scholar] [CrossRef]
- Molina, V.; Mas, R.; Carbajal, D. D-002 (beeswax alcohols): Concurrent joint health benefits and gastroprotection. Indian J. Pharm. Sci. 2015, 77, 127. [Google Scholar] [CrossRef]
- Cho, K.-H.; Baek, S.H.; Nam, H.-S.; Kim, J.-E.; Kang, D.-J.; Na, H.; Zee, S. Cuban Sugar Cane Wax Alcohol Exhibited Enhanced Antioxidant, Anti-Glycation and Anti-Inflammatory Activity in Reconstituted High-Density Lipoprotein (rHDL) with Improved Structural and Functional Correlations: Comparison of Various Policosanols. Int. J. Mol. Sci. 2023, 24, 3186. [Google Scholar] [CrossRef]
- Cho, K.-H.; Kim, J.-E.; Nam, H.-S.; Kang, D.-J.; Baek, S.-H. Comparison of Policosanols via Incorporation into Reconstituted High-Density Lipoproteins: Cuban Policosanol (Raydel®) Exerts the Highest Antioxidant, Anti-Glycation, and Anti-Inflammatory Activity. Molecules 2023, 28, 6715. [Google Scholar] [CrossRef]
- Cho, K.-H.; Kim, J.-E.; Nam, H.-S.; Kang, D.-J.; Na, H.-J. Anti-inflammatory activity of CIGB-258 against acute toxicity of carboxymethyllysine in paralyzed zebrafish via enhancement of high-density lipoproteins stability and functionality. Int. J. Mol. Sci. 2022, 23, 10130. [Google Scholar] [CrossRef]
- Sakata, N.; Uesugi, N.; Takebayashi, S.; Nagai, R.; Jono, T.; Horiuchi, S.; Takeya, M.; Itabe, H.; Takano, T.; Myint, T. Glycoxidation and lipid peroxidation of low-density lipoprotein can synergistically enhance atherogenesis. Cardiovasc. Res. 2001, 49, 466–475. [Google Scholar] [CrossRef]
- Trede, N.S.; Zapata, A.; Zon, L.I. Fishing for lymphoid genes. Trends Immunol. 2001, 22, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Novoa, B.; Bowman, T.; Zon, L.; Figueras, A. LPS response and tolerance in the zebrafish (Danio rerio). Fish Shellfish. Immunol. 2009, 26, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-H.; Kim, J.-E.; Bahuguna, A.; Kang, D.-J. Ozonated Sunflower Oil Exerted Potent Anti-Inflammatory Activities with Enhanced Wound Healing and Tissue Regeneration Abilities against Acute Toxicity of Carboxymethyllysine in Zebrafish with Improved Blood Lipid Profile. Antioxidants 2023, 12, 1625. [Google Scholar] [CrossRef]
- Cho, K.-H.; Kim, J.-H.; Nam, H.-S.; Kang, D.-J. Efficacy Comparison Study of Human Epidermal Growth Factor (EGF) between Heberprot-P® and Easyef® in Adult Zebrafish and Embryo under Presence or Absence Combination of Diabetic Condition and Hyperlipidemia to Mimic Elderly Patients. Geriatrics 2022, 7, 45. [Google Scholar] [CrossRef]
- Mas, R. D-002. Drugs Future 2001, 26, 0731. [Google Scholar] [CrossRef]
- Havel, R.J.; Eder, H.A.; Bragdon, J.H. The distribution and chemical composition of ultracentrifugally separated lipoproteins in human serum. J. Clin. Investig. 1955, 34, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Brewer, H.B.; Ronan, R.; Meng, M.; Bishop, C. Isolation and Characterization of Apolipoproteins AI, A-II, and A-IV. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1986; Volume 128, pp. 223–246. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- McPherson, J.D.; Shilton, B.H.; Walton, D.J. Role of fructose in glycation and cross-linking of proteins. Biochemistry 1988, 27, 1901–1907. [Google Scholar] [CrossRef] [PubMed]
- Matz, C.E.; Jonas, A. Micellar complexes of human apolipoprotein AI with phosphatidylcholines and cholesterol prepared from cholate-lipid dispersions. J. Biol. Chem. 1982, 257, 4535–4540. [Google Scholar] [CrossRef]
- Noble, R.P. Electrophoretic separation of plasma lipoproteins in agarose gel. J. Lipid Res. 1968, 9, 693–700. [Google Scholar] [CrossRef]
- Markwell, M.A.K.; Haas, S.M.; Bieber, L.; Tolbert, N. A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal. Biochem. 1978, 87, 206–210. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Blatter Garin, M.-C.; Moren, X.; James, R.W. Paraoxonase-1 and serum concentrations of HDL-cholesterol and apoA-I. J. Lipid Res. 2006, 47, 515–520. [Google Scholar] [CrossRef]
- Nusslein-Volhard, C.; Dahm, R. Zebrafish: A Practical Approach, 1st ed.; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- National Research Council of the National Academy of Sciences. Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, DC, USA, 2010. [Google Scholar]
- Cho, K.-H.; Nam, H.-S.; Kim, J.-E.; Na, H.-J.; del Carmen Dominguez-Horta, M.; Martinez-Donato, G. CIGB-258 exerts potent anti-inflammatory activity against carboxymethyllysine-induced acute inflammation in hyperlipidemic zebrafish via the protection of apolipoprotein AI. Int. J. Mol. Sci. 2023, 24, 7044. [Google Scholar] [CrossRef]
- Owusu-Ansah, E.; Yavari, A.; Mandal, S.; Banerjee, U. Distinct mitochondrial retrograde signals control the G1-S cell cycle checkpoint. Nat. Genet. 2008, 40, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Sofuni, T.; Ishidate, M., Jr. An application of acridine orange fluorescent staining to the micronucleus test. Mutat. Res. Lett. 1983, 120, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Hanus, J.; Zhang, H.; Wang, Z.; Liu, Q.; Zhou, Q.; Wang, S. Induction of necrotic cell death by oxidative stress in retinal pigment epithelial cells. Cell Death Dis. 2013, 4, e965. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Mori, Y.; Makino, Y.; Morimitsu, Y.; Hiroi, S.; Ishikawa, T.; Osawa, T. Formation of Nε-(hexanonyl) lysine in protein exposed to lipid hydroperoxide: A plausible marker for lipid hydroperoxide-derived protein modification. J. Biol. Chem. 1999, 274, 20406–20414. [Google Scholar] [CrossRef]
- Pirillo, A.; Norata, G.D.; Catapano, A.L. LOX-1, OxLDL, and atherosclerosis. Mediat. Inflamm. 2013, 2013, 152786. [Google Scholar] [CrossRef]
- Duan, H.; Song, P.; Li, R.; Su, H.; He, L. Attenuating lipid metabolism in atherosclerosis: The potential role of Anti-oxidative effects on low-density lipoprotein of herbal medicines. Front. Pharmacol. 2023, 14, 874. [Google Scholar] [CrossRef]
- Kosmas, C.E.; Martinez, I.; Sourlas, A.; Bouza, K.V.; Campos, F.N.; Torres, V.; Montan, P.D.; Guzman, E. High-density lipoprotein (HDL) functionality and its relevance to atherosclerotic cardiovascular disease. Drugs Context 2018, 7, 212525. [Google Scholar] [CrossRef]
- Cho, K.-H. The current status of research on high-density lipoproteins (HDL): A paradigm shift from HDL quantity to HDL quality and HDL functionality. Int. J. Mol. Sci. 2022, 23, 3967. [Google Scholar] [CrossRef]
- Shariat, S.Z.A.S.; Mostafavi, S.A.; Khakpour, F. Antioxidant effects of vitamins C and E on the low-density lipoprotein oxidation mediated by myeloperoxidase. Iran. Biomed. J. 2013, 17, 22. [Google Scholar] [CrossRef]
- Ojo, O.O.; Leake, D.S. Vitamins E and C do not effectively inhibit low density lipoprotein oxidation by ferritin at lysosomal pH. Free. Radic. Res. 2021, 55, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Amarowicz, R.; B Pegg, R. The potential protective effects of phenolic compounds against low-density lipoprotein oxidation. Curr. Pharm. Des. 2017, 23, 2754–2766. [Google Scholar] [CrossRef] [PubMed]
- Barter, P.J.; Nicholls, S.; Rye, K.-A.; Anantharamaiah, G.; Navab, M.; Fogelman, A.M. Antiinflammatory properties of HDL. Circ. Res. 2004, 95, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Nobécourt, E.; Tabet, F.; Lambert, G.; Puranik, R.; Bao, S.; Yan, L.; Davies, M.J.; Brown, B.E.; Jenkins, A.J.; Dusting, G.J. Nonenzymatic glycation impairs the antiinflammatory properties of apolipoprotein AI. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Igarashi, K.; Ogata, N.; Oka, Y.; Ichiyanagi, K.; Yamanouchi, T. Markedly increased serum and urinary fructose concentrations in diabetic patients with ketoacidosis or ketosis. Acta Diabetol. 2012, 49, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.T.; Cruz, S.; Narayanaswami, V. Reconfiguring nature’s cholesterol accepting lipoproteins as nanoparticle platforms for transport and delivery of therapeutic and imaging agents. Nanomaterials 2020, 10, 906. [Google Scholar] [CrossRef] [PubMed]
- Lacko, A.G.; Sabnis, N.A.; Nagarajan, B.; McConathy, W.J. HDL as a drug and nucleic acid delivery vehicle. Front. Pharmacol. 2015, 6, 247. [Google Scholar] [CrossRef]
- Aguirre-Portolés, C.; Feliu, J.; Reglero, G.; Ramírez de Molina, A. ABCA1 overexpression worsens colorectal cancer prognosis by facilitating tumour growth and caveolin-1-dependent invasiveness, and these effects can be ameliorated using the BET inhibitor apabetalone. Mol. Oncol. 2018, 12, 1735–1752. [Google Scholar] [CrossRef]
- Gao, F.; Chattopadhyay, A.; Navab, M.; Grijalva, V.; Su, F.; Fogelman, A.M.; Reddy, S.T.; Farias-Eisner, R. Apolipoprotein AI mimetic peptides inhibit expression and activity of hypoxia-inducible factor-1α in human ovarian cancer cell lines and a mouse ovarian cancer model. J. Pharmacol. Exp. Ther. 2012, 342, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Takagi, T.; Handa, O.; Yoshikawa, T. Lipid Hydroperoxide-Derived Modification of Proteins in Gastrointestinal Tract. In Lipid Hydroperoxide-Derived Modification of Biomolecules; Springer: Berlin/Heidelberg, Germany, 2014; pp. 137–148. [Google Scholar] [CrossRef]
- Naito, Y.; Yoshikawa, T.; Yoshida, N.; Kondo, M. Role of oxygen radical and lipid peroxidation in indomethacin-induced gastric mucosal injury. Dig. Dis. Sci. 1998, 43 (Suppl. 9), 30S–34S. [Google Scholar]
- Puente, R.; Illnait, J.; Mas, R.; Carbajal, D.; Mendoza, S.; Fernández, J.C.; Mesa, M.; Gamez, R.; Reyes, P. Evaluation of the effect of D-002, a mixture of beeswax alcohols, on osteoarthritis symptoms. Korean J. Intern. Med. 2014, 29, 191. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zee, S.; Cho, K.-H. Beeswax Alcohol and Fermented Black Rice Bran Synergistically Ameliorated Hepatic Injury and Dyslipidemia to Exert Antioxidant and Anti-Inflammatory Activity in Ethanol-Supplemented Zebrafish. Biomolecules 2023, 13, 136. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Initial Molar Ratio POPC:FC:apoA-I:BWA 1 | Final Amount (μg) in 0.7 mL (POPC:FC:apoA-I:BWA) | WMF (nm) | Diameter (nm) |
|---|---|---|---|---|
| rHDL-0 | 95:5:1:0 | 2600:70:1000:0 | 330.9 ± 0.1 | 61.2 ± 1.6 |
| rHDL-0.5 | 95:5:1:0.5 | 2600:70:1000:7.5 | 331.0 ± 0.0 | 65.4 ± 2.3 |
| rHDL-1 | 95:5:1:1 | 2600:70:1000:15 | 331.4 ± 0.1 | 70.5 ± 2.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, K.-H.; Baek, S.-H.; Nam, H.-S.; Bahuguna, A. Enhancement of Antioxidant and Anti-Glycation Properties of Beeswax Alcohol in Reconstituted High-Density Lipoprotein: Safeguarding against Carboxymethyllysine Toxicity in Zebrafish. Antioxidants 2023, 12, 2116. https://doi.org/10.3390/antiox12122116
Cho K-H, Baek S-H, Nam H-S, Bahuguna A. Enhancement of Antioxidant and Anti-Glycation Properties of Beeswax Alcohol in Reconstituted High-Density Lipoprotein: Safeguarding against Carboxymethyllysine Toxicity in Zebrafish. Antioxidants. 2023; 12(12):2116. https://doi.org/10.3390/antiox12122116
Chicago/Turabian StyleCho, Kyung-Hyun, Seung-Hee Baek, Hyo-Seon Nam, and Ashutosh Bahuguna. 2023. "Enhancement of Antioxidant and Anti-Glycation Properties of Beeswax Alcohol in Reconstituted High-Density Lipoprotein: Safeguarding against Carboxymethyllysine Toxicity in Zebrafish" Antioxidants 12, no. 12: 2116. https://doi.org/10.3390/antiox12122116