
Metabolite Profiling, through LC-ESI/LTQOrbitrap/MS Analysis, of Antioxidant Extracts from Physalis alkekengi L.
 , ,
, ,  , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Raw Materials
2.2. Reagents and Solvents
2.3. Sample Preparation
2.4. LC-ESI/LTQOrbitrap/MS and LC-ESI/LTQOrbitrap/MS/MS Analysis
2.5. DPPH· Radical Scavenging Activity
2.6. Trolox Equivalent Antioxidant Capacity (TEAC) Assay
2.7. Cell Culture
2.8. MTT Viability Test
2.9. Determination of Intracellular ROS Production
2.10. Statistical Analyses
3. Results
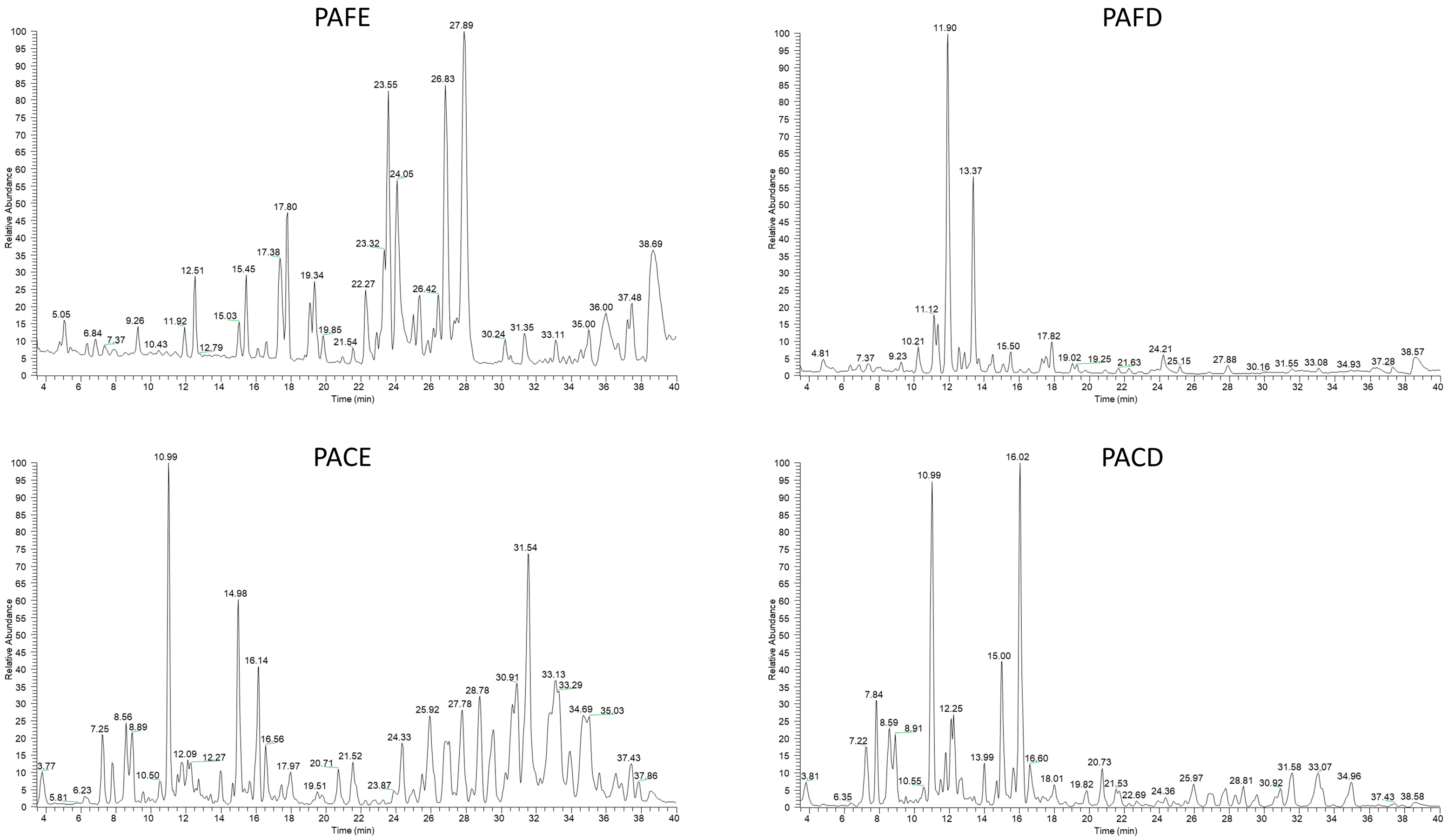
3.1. Identification of Metabolites in Physalis alkekengi L. Fruit and Calyx Extracts by LC-ESI/LTQOrbitrap/MS and LC-ESI-LTQOrbitrap/MS/MS Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Rt | Identity | Molecular Formula | [M + H]+ | Δppm | MS/MS | References | PACE | PAFE | PACD | PAFD |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 7.81 | quercetin 3-O-rutinoside-7-O-glucoside | C33H40O20 | 773.2162 | 3.5 | 611; 449; 303; | Mass Bank | nd | nd | x | x |
| 2 | 8.56 | p-coumaric acid | C9H6O3 | 163.0391 | −2.5 | 165; 147 | [21] | nd | nd | x | x |
| 3 | 8.57 | hydroxycoumarin | C9H6O3 | 163.0395 | 0.9 | 163; 145 | [21] | x | x | x | x |
| 4 | 8.92 | suberic acid | C8H13O4 | 175.0816 | −1.8 | 157; 139 | [21] | nd | nd | x | x |
| 5 | 9.15 | feruloyl dihexoside (2) | C22H30O14 | 519.1713 | 0.7 | 195 | [22] | x | x | x | x |
| 6 | 9.25 | quinic acid | C7H12O6 | 193.0556 | 2.1 | 135; 117 | [21] | x | x | x | x |
| 7 | 10.96 | rutin | C27H30O16 | 611.1534 | −0.6 | 465; 303 | [23] | x | x | x | x |
| 8 | 11.26 | 5,6α-epoxy-physalin C | C28H30O10 | 527.1937 | −1.5 | 509; 499; 483 | [21] | x | nd | x | nd |
| 9 | 11.49 | 1,3-dicaffeoylquinic acid | C25H24O12 | 517.1344 | 0.6 | 193 | [21] | x | x | x | x |
| 10 | 11.68 | 4-hydroxyneophysalin A isomer | C28H30O11 | 543.1866 | −0.5 | 511; 497; 449 | [21] | x | nd | nd | nd |
| 11 | 11.82 | luteolin-rutinoside | C27H30O15 | 595.1584 | −0.7 | 449; 287 | Mass Bank | x | x | x | x |
| 12 | 12.66 | 4,5-dicaffeoylquinic acid | C25H24O12 | 517.1344 | 1 | 284; 269; 141 | [21] | x | x | x | x |
| 13 | 13.13 | L-tryptophan | C11H12N2O2 | 205.0707 | 2.3 | 171; 147 | [22] | x | x | x | x |
| 14 | 14.91 | 4-hydroxyneophysalin A isomer | C28H29O11 | 541.1713 | −1 | 513; 497; 465 | [21] | x | nd | x | nd |
| 15 | 15.59 | 4-hydroxyneophysalin A | C28H30O11 | 541.1866 | 0.5 | 513; 497; 465 | [21] | x | x | x | x |
| 16 | 15.95 | isolariciresinol O-glucoside | C26H34O11 | 523.2179 | 1 | 505; 184 | [21] | x | x | x | x |
| 17 | 15.96 | 5,6-O-epoxy-physalin C isomer | C28H30O10 | 527.1937 | −1.5 | 509; 499; 483 | [21] | x | nd | x | x |
| 18 | 16.14 | quercetin 3-O-glucoside | C21H20O12 | 465.1033 | −0.9 | 303; 285 | [21] | x | x | x | x |
| 19 | 16.67 | luteolin 7-O-glucoside | C21H20O11 | 449.1083 | −0.9 | 287; 285 | [21] | x | x | x | x |
| 20 | 16.91 | 5,6-O-epoxy-physalin C | C28H30O10 | 527.1917 | 0.8 | 509; 499; 483 | [21] | x | nd | x | nd |
| 21 | 16.99 | quercetin O-rhamnoside | C21H20O11 | 449.1083 | −0.9 | 303; 287 | [21] | x | x | x | x |
| 22 | 17.33 | 3-caffeoylquinic acid | C16H18O9 | 355.0872 | 0.5 | 337; 284 | [21] | x | x | x | x |
| 23 | 17.92 | vitexin | C21H20O10 | 433.1134 | 1.2 | 433; 417; 379 | [21] | x | x | x | x |
| 24 | 17.92 | apigenin O-glucoside | C21H20O10 | 433.1134 | 1.2 | 271 | [21] | x | x | x | x |
| 25 | 17.94 | caffeic acid | C9H8O4 | 181.0500 | −2.2 | 163 | [21] | x | x | x | x |
| 26 | 18.21 | 4-hydroxyneophysalin A isomer | C28H30O11 | 541.1866 | 0.8 | 284; 198; 141 | [21] | x | x | x | x |
| 27 | 19.82 | 4,7-didehydro-7-deoxyneophysalin A | C28H28O9 | 509.1811 | 0.7 | 497; 465 | [21] | x | x | x | nd |
| 28 | 19.82 | N,N′-dicaffeoylspermidine (1) | C25H31N3O6 | 470.2211 | 0.5 | 333; 289 | [22] | x | x | x | x |
| 29 | 19.86 | 5-ethoxy-6-hydroxy-5,6-dihydrophysalin B | C30H36O11 | 573.2335 | 0.7 | 555; 523 | [21] | x | nd | x | nd |
| 30 | 19.97 | physalin Z | C28H30O10 | 527.1917 | 0.6 | 509; 499; 483 | [21] | x | x | x | x |
| 31 | 19.97 | physalin A | C28H30O10 | 527.1917 | 0.8 | 509; 499; 483 | [21] | x | x | x | x |
| 32 | 19.97 | feruloyl hexoside | C16H20O9 | 355.1040 | 1.8 | 195; 179 | [22] | x | x | x | x |
| 33 | 20.71 | physalin C | C28H30O9 | 511.1968 | 0.6 | 493; 481; 423 | [21] | x | x | x | x |
| 34 | 20.73 | physalin O | C28H32O10 | 529.1917 | 0.4 | 511; 483 | [21] | nd | nd | x | nd |
| 35 | 21.85 | physalin N | C28H30O10 | 527.1917 | 0.6 | 509; 499; 483 | [21] | x | x | x | x |
| 36 | 21.87 | caffeoylglycerol | C12H14O6 | 255.0712 | 0.8 | 214 | [21] | nd | x | nd | x |
| 37 | 21.9 | quercetin | C15H10O7 | 302.2303 | 0.7 | 257; 229 | Mass Bank | ||||
| 38 | 21.99 | isophysalin B | C28H30O9 | 511.1968 | 0.6 | 493; 467; 423 | [21] | x | x | x | nd |
| 39 | 23.28 | N,N′-dicaffeoylspermidine (2) | C25H31N3O6 | 470.2211 | 0.5 | 333; 289 | [22] | x | x | x | x |
| 40 | 23.55 | sonchuionoside C | C19H30O8 | 387.2018 | −0.7 | 331; 198 | [21] | x | x | x | x |
| 41 | 23.9 | luteolin | C15H10O6 | 285.0383 | 1.3 | 153; 135 | [21] | x | x | x | x |
| 42 | 23.91 | 4,7-didehydro-physalin B | C28H28O9 | 509.1811 | 0.7 | 497; 465 | [21] | x | x | x | nd |
| 43 | 24.02 | 5-caffeoylquinic acid | C16H18O9 | 355.0871 | 1.5 | 198; 141; 109 | [21] | x | x | x | x |
| 44 | 24.33 | 7β-hydroxy-25,27-didehydrophysalin L | C28H30O10 | 527.1917 | 0.6 | 509; 499; 483 | [21] | x | x | x | x |
| 45 | 24.81 | coumaroyl dihexoside | C21H28O13 | 489.1608 | 1.7 | 327 | [22] | x | x | x | x |
| 46 | 25.46 | coumaroyl hexoside | C15H18O8 | 327.1079 | 2.1 | 188; 165 | [22] | x | x | x | x |
| 47 | 26.14 | 4,7-didehydrophysalin B | C28H28O9 | 509.1811 | 0.7 | 509; 499; 483 | [21] | x | x | x | nd |
| 48 | 26.41 | physalin M | C28H32O9 | 513.2124 | 0.8 | 495; 451; 135 | [21] | x | nd | x | x |
| 49 | 26.82 | 4,7-didehydroneophysalin B | C28H28O9 | 509.1811 | 0.7 | 497; 465 | [21] | x | x | x | nd |
| 50 | 28.41 | apigenin | C15H10O5 | 271.0606 | 1.6 | 153 | [21] | x | nd | x | nd |
| 51 | 28.45 | physalin B | C28H30O9 | 511.1968 | 0.6 | 493; 467; 423 | [21] | x | x | x | nd |
| 52 | 30.63 | (all-E)-antheraxanthin | C40H56O3 | 585.4307 | 1.3 | 567; 549; 493 | [20] | x | x | nd | nd |
| 53 | 30.91 | zeaxanthin myristate | C54H82O3 | 779.6342 | 0.6 | 761; 687; 669; 551; 533 | [20] | x | x | nd | nd |
| 54 | 31.18 | (13Z)-β-carotene | C40H56 | 537.4460 | 1.3 | 445; 347; 281; 255 | [20] | x | x | nd | nd |
| 55 | 31.18 | (all-E)-α-carotene | C40H56 | 537.4460 | 1.3 | 481; 399 | [20] | x | x | nd | nd |
| 56 | 31.46 | (all-E)-lutein | C40H56O2 | 569.4358 | 1.4 | 551 | [20] | x | x | nd | nd |
| 57 | 32.49 | (15Z)-zeaxanthin | C40H56O2 | 569.4358 | 1.4 | 551 | [20] | x | x | nd | nd |
| 58 | 33.96 | (all-E)-zeaxanthin | C40H56O2 | 569.4358 | 1.4 | 551 | [20] | x | x | nd | nd |
3.2. Antioxidant Activity Evaluated by Spectrophotometric Assays
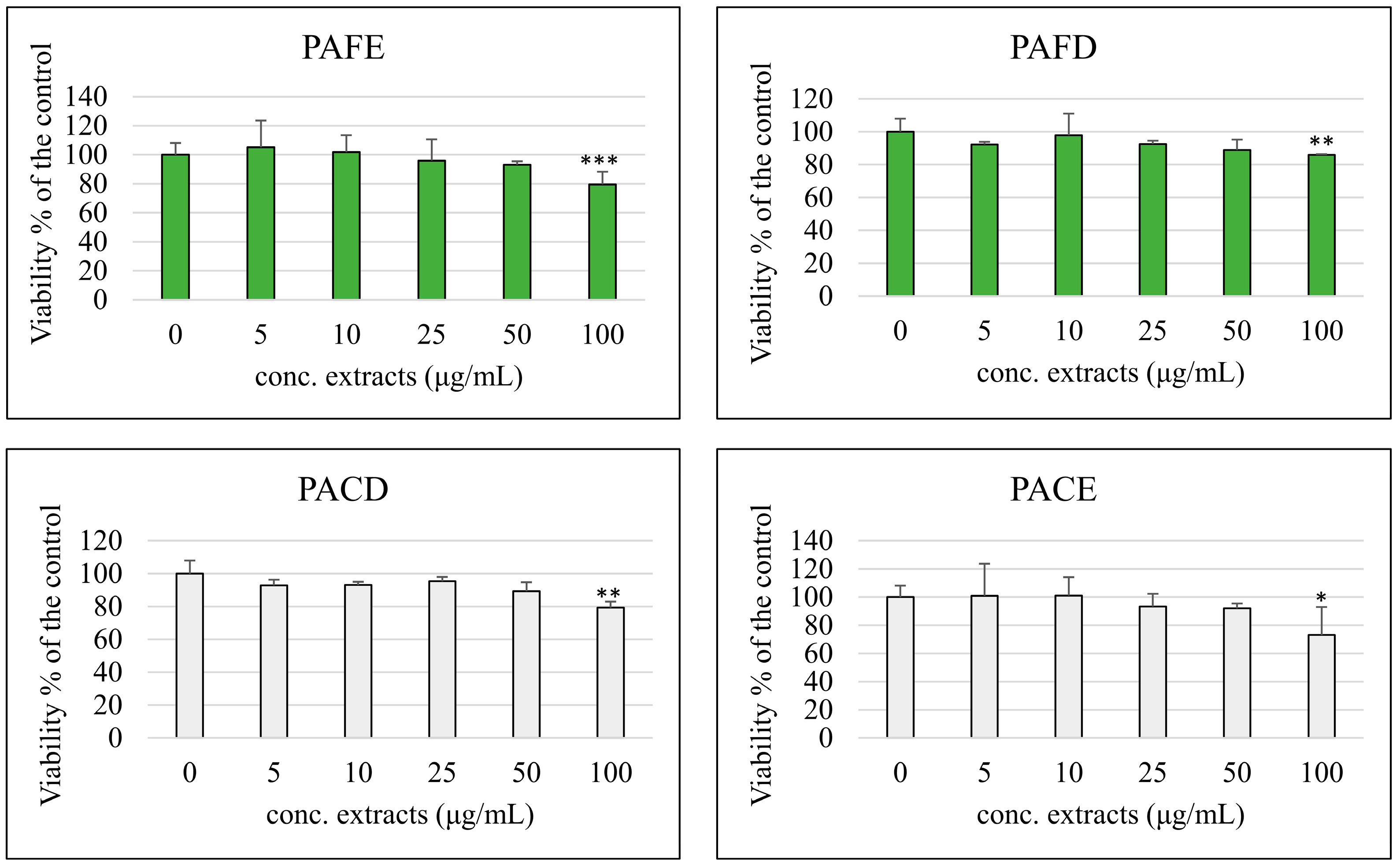
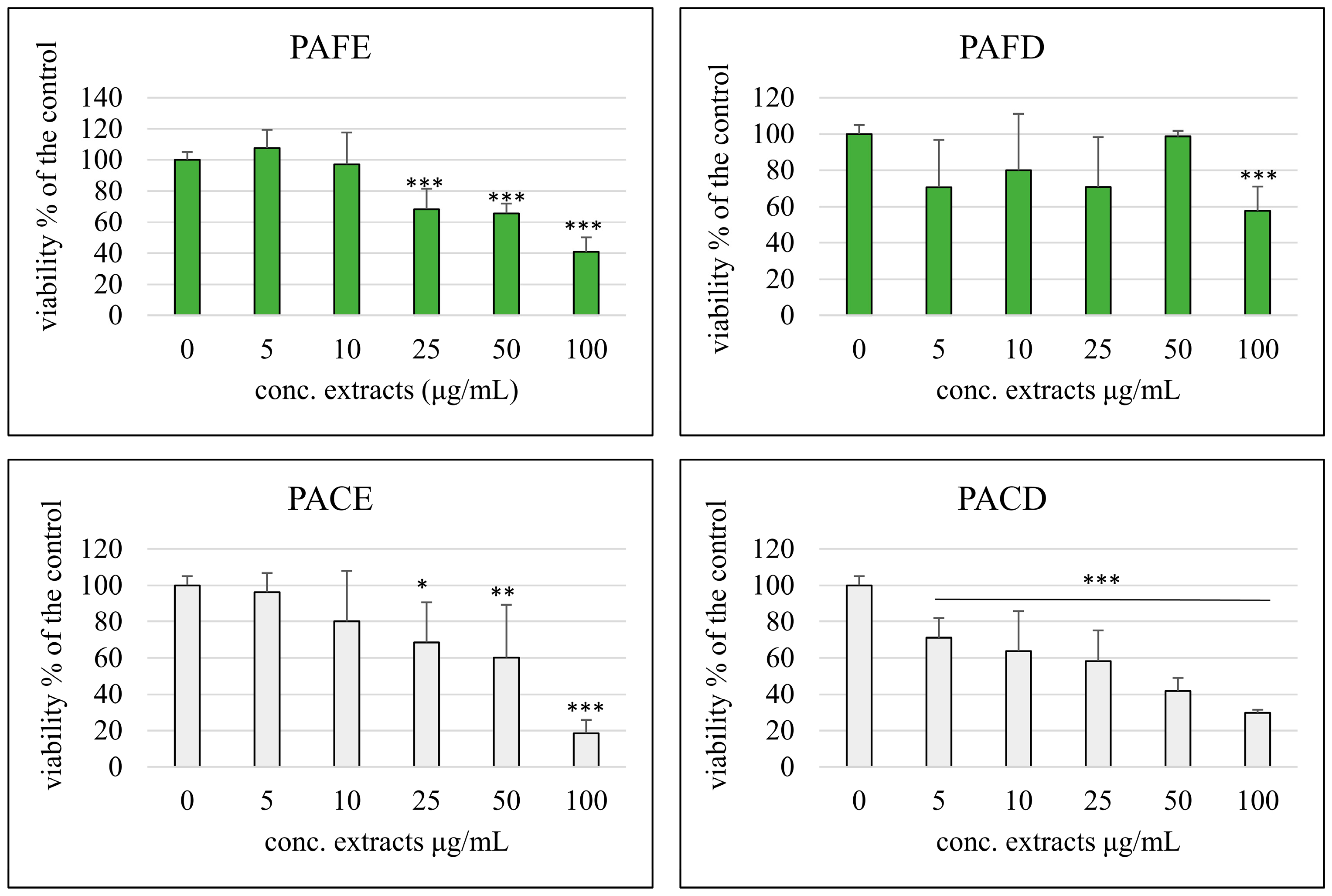
3.3. Cell Viability
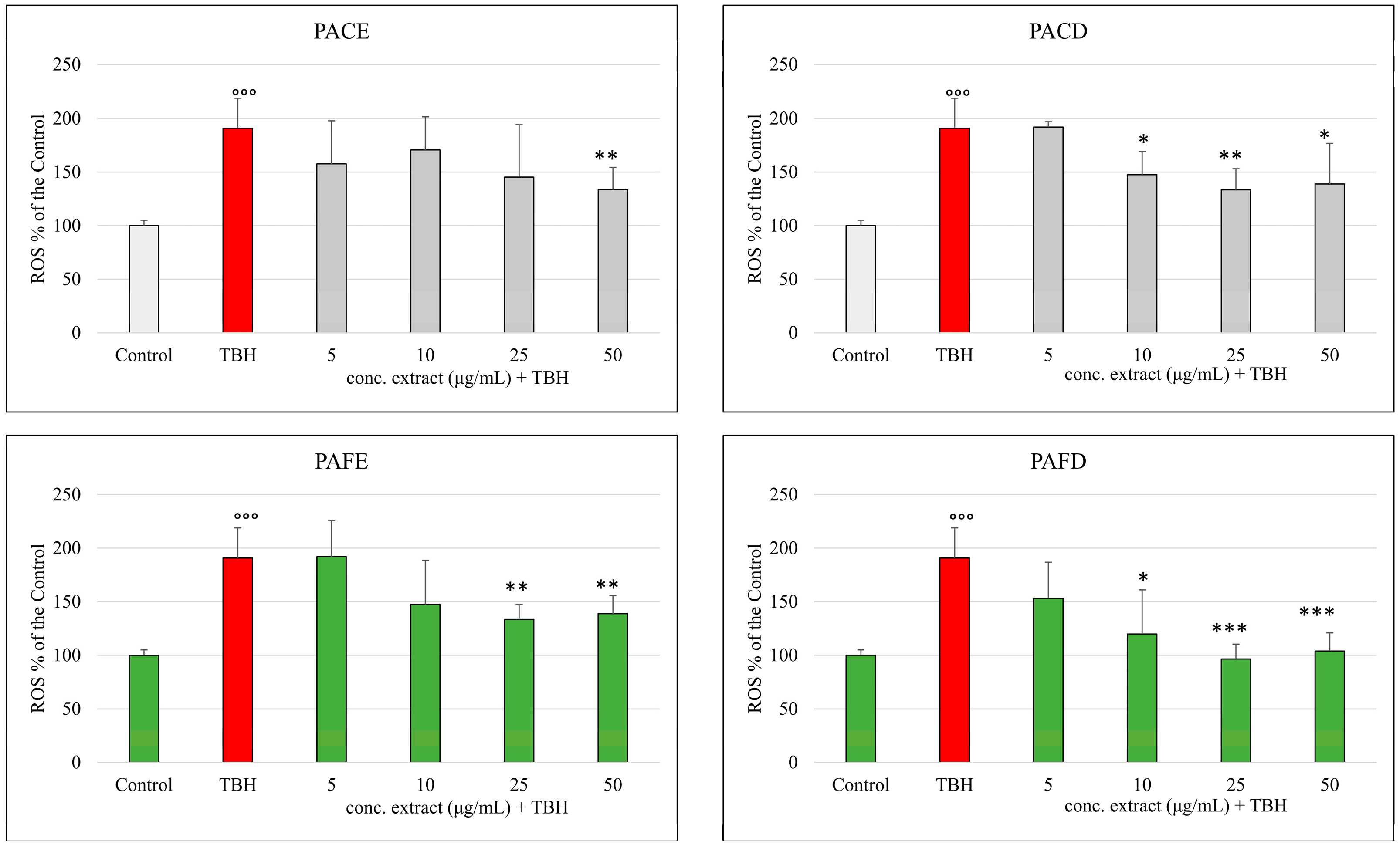
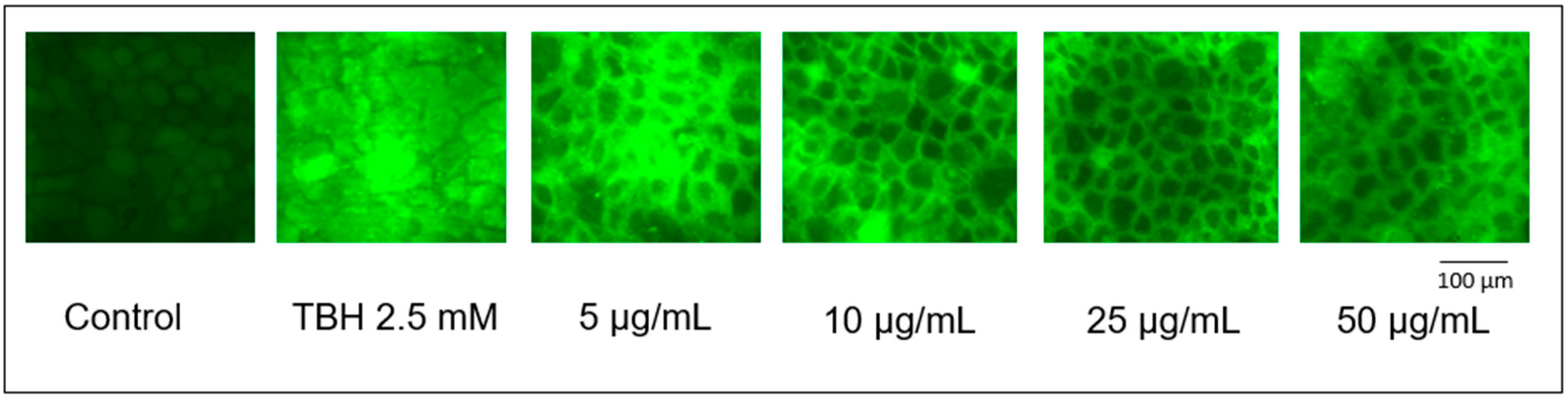
3.4. ROS Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liang, L.; Li, C.; Wang, Y.; Yue, Y.; Zhang, H.; Yang, M.; Cao, X.; Zhao, M.; Du, J.; Peng, M.; et al. Physalis alkekengi L. var. franchetii (Mast.) Makino: A review of the pharmacognosy, chemical constituents, pharmacological effects, quality control, and applications. Phytomedicine 2022, 105, 154328. [Google Scholar] [CrossRef] [PubMed]
- Popova, V.; Petkova, Z.; Mazova, N.; Ivanova, T.; Petkova, N.; Stoyanova, M.; Stoyanova, A.; Ercisli, S.; Okcu, Z.; Skrovankova, S.; et al. Chemical Composition Assessment of Structural Parts (Seeds, Peel, Pulp) of Physalis alkekengi L. Fruits. Molecules 2022, 27, 5787. [Google Scholar] [CrossRef] [PubMed]
- Popova, V.; Ivanova, T.; Stoyanova, M.; Mazova, N.; Dimitrova, I.; Stoyanova, A.; Assouguem, A.; Kara, M.; Topçu, H.; Farah, A.; et al. Phytochemical analysis of leaves and stems of Physalis alkekengi L. (Solanaceae). Open Chem. 2022, 20, 1292–1303. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, Y.; Ren, Y.; Luan, L.; Wu, Y. Quantitative and transformation product analysis of major active physalins from Physalis alkekengi var. franchetii (Chinese lantern) using ultraperformance liquid chromatography with electrospray ionisation tandem mass spectrometry and time-of-flight mass spectrometry. Phytochem. Anal. 2012, 23, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, M.; Rafieian-kopaei, M.; Naghdi, N.; Mozaffari Nejad, A.S.; Afsordeh, O. Physalis alkekengi: A review of Its Therapeutic Effects. J. Chem. Pharm. Sci. 2016, 9, 1472–1485. [Google Scholar]
- Li, A.-L.; Chen, B.-J.; Li, G.-H.; Zhou, M.-X.; Li, Y.-R.; Ren, D.-M.; Lou, H.-X.; Wang, X.-N.; Shen, T. Physalis alkekengi L. var. franchetii (Mast.) Makino: An ethnomedical, phytochemical and pharmacological review. J. Ethnopharmacol. 2018, 210, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Sun, Y.; Cao, F.; Yang, B.; Kuang, H. Natural Products from Physalis alkekengi L. var. franchetii (Mast.) Makino: A Review on Their Structural Analysis, Quality Control, Pharmacology, and Pharmacokinetics. Molecules 2022, 27, 695. [Google Scholar] [CrossRef] [PubMed]
- Popova, V.; Stoyanova, A.; Mazova, N. Phytochemical composition and biological activity of Physalis spp.: A mini-review. Food Sci. Appl. Biotechnol. 2020, 3, 56. [Google Scholar] [CrossRef]
- Teng, Y.A.-O.; Yuan, Q.A.-O.X.; Wu, Y.; Wu, S.; Su, J.; Zhang, P.; Zhang, Y.A.-O. Research on the Chemical Constituents against Alzheimer’s Disease of the Fruits of Physalis alkekengi L. var. franchetii (Mast.) Makino. Chem. Biodivers. 2023, 20, e202301075. [Google Scholar] [CrossRef]
- El-Ghorab, A.; Ramadana, M.; El-Ghoraba, A.; Ghanemb, K. Volatile compounds, antioxidants, and anticancer activities of Cape gooseberry fruit (Physalis peruviana L.): An in-vitro study. J. Arab. Soc. Med. Res. 2015, 10, 56–64. [Google Scholar] [CrossRef]
- Castro, J.; Ocampo, Y.; Franco, L. Cape Gooseberry [Physalis peruviana L.] Calyces Ameliorate TNBS Acid-induced Colitis in Rats. J. Crohns Colitis 2015, 9, 1004–1015. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.C.; Sang-Ngern, M.; Pezzuto, J.M.; Ma, C. The Daniel K. Inouye College of Pharmacy Scripts: Poha Berry (Physalis peruviana) with Potential Anti-inflammatory and Cancer Prevention Activities. Hawai’i J. Med. Public Health 2016, 75, 353. [Google Scholar]
- Etzbach, L.; Pfeiffer, A.; Weber, F.; Schieber, A. Characterization of carotenoid profiles in goldenberry (Physalis peruviana L.) fruits at various ripening stages and in different plant tissues by HPLC-DAD-APCI-MS(n). Food Chem. 2018, 245, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Crescenzi, M.A.; D’Urso, G.; Piacente, S.; Montoro, P. LC-ESI/LTQOrbitrap/MS Metabolomic Analysis of Fennel Waste (Foeniculum vulgare Mill.) as a Byproduct Rich in Bioactive Compounds. Foods 2021, 10, 1893. [Google Scholar] [CrossRef] [PubMed]
- Santoru, M.L.; Piras, C.; Murgia, F.; Spada, M.; Tronci, L.; Leoni, V.P.; Serreli, G.; Deiana, M.; Atzori, L. Modulatory Effect of Nicotinic Acid on the Metabolism of Caco-2 Cells Exposed to IL-1β and LPS. Metabolites 2020, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Deiana, M.; Montoro, P.; Jerković, I.; Atzeri, A.; Marijanović, Z.; Serreli, G.; Piacente, S.; Tuberoso, C.I.G. First characterization of Pompia intrea candied fruit: The headspace chemical profile, polar extract composition and its biological activities. Food Res. Int. 2019, 120, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Gil, K.A.; Nowicka, P.; Wojdyło, A.; Serreli, G.; Deiana, M.; Tuberoso, C.I.G. Antioxidant Activity and Inhibition of Digestive Enzymes of New Strawberry Tree Fruit/Apple Smoothies. Antioxidants 2023, 12, 805. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Jiang, Z.H.; Liu, H.X.; Chen, L.X.; Qu, G.X.; Qiu, F. Flavonoid glycosides of the calyx Physalis. J. Shenyang Pharm. Univ. 2007, 24, 744–747. [Google Scholar]
- Kylli, P.; Nousiainen, P.; Biely, P.; Sipilä, J.; Tenkanen, M.; Heinonen, M. Antioxidant potential of hydroxycinnamic acid glycoside esters. J. Agric. Food Chem. 2008, 56, 4797–4805. [Google Scholar] [CrossRef]
- Wen, X.; Hempel, J.; Schweiggert, R.M.; Ni, Y.; Carle, R. Carotenoids and Carotenoid Esters of Red and Yellow Physalis (Physalis alkekengi L. and P. pubescens L.) Fruits and Calyces. J. Agric. Food Chem. 2017, 65, 6140–6151. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, X.F.; Xin, M.M.; Liu, H.B.; Sun, L.J.; Morris-Natschke, S.L.; Chen, Y.; Lee, K.H. Antidiabetic potential of the ethyl acetate extract of Physalis alkekengi and chemical constituents identified by HPLC-ESI-QTOF-MS. J. Ethnopharmacol. 2018, 225, 202–210. [Google Scholar] [CrossRef]
- Wen, X.; Erşan, S.; Li, M.; Wang, K.; Steingass, C.B.; Schweiggert, R.M.; Ni, Y.; Carle, R. Physicochemical characteristics and phytochemical profiles of yellow and red Physalis (Physalis alkekengi L. and P. pubescens L.) fruits cultivated in China. Food Res. Int. 2019, 120, 389–398. [Google Scholar] [CrossRef]
- Liu, M.; Qiu, Z.; Zhao, X.; Sun, L.; Wang, L.; Ren, X.; Deng, Y. Assessing the Quality of Calyx of Physalis alkekengi L. var. franchetii Based on Quantitative Analysis of Q-Marker Combined with Chemometrics and Machine Learning Algorithms. J. Chem. 2021, 2021, 8502929. [Google Scholar] [CrossRef]
- Matsuura, T.; Kawai, M.; Nakashima, R.; Butsugan, Y. Bitter principles of Physalis Alkekengi var Francheti: Structure of physalin a. Tetrahedron Lett. 1969, 10, 1083–1086. [Google Scholar] [CrossRef]
- Huang, C.; Xu, Q.; Chen, C.; Song, C.; Xu, Y.; Xiang, Y.; Feng, Y.; Ouyang, H.; Zhang, Y.; Jiang, H. The rapid discovery and identification of physalins in the calyx of Physalis alkekengi L. var. franchetii (Mast.) Makino using ultra-high performance liquid chromatography–quadrupole time of flight tandem mass spectrometry together with a novel three-step data mining strategy. J. Chromatogr. A 2014, 1361, 139–152. [Google Scholar]
- Qiu, L.; Zhao, F.; Jiang, Z.H.; Chen, L.X.; Zhao, Q.; Liu, H.X.; Yao, X.S.; Qiu, F. Steroids and flavonoids from Physalis alkekengi var. franchetii and their inhibitory effects on nitric oxide production. J. Nat. Prod. 2008, 71, 642–646. [Google Scholar] [CrossRef]
- Shu, Z.; Xing, N.; Wang, Q.; Li, X.; Xu, B.; Li, Z.; Kuang, H. Antibacterial and Anti-Inflammatory Activities of Physalis Alkekengi var. franchetii and Its Main Constituents. Evid. Based Complement. Altern. Med. 2016, 2016, 4359394. [Google Scholar] [CrossRef]
- Zhang, W.N.; Tong, W.Y. Chemical Constituents and Biological Activities of Plants from the Genus Physalis. Chem. Biodivers. 2016, 13, 48–65. [Google Scholar] [CrossRef]
- He, B.W.; Wang, F.F.; Qu, L.P. Anti-Inflammatory and Antioxidant Properties of Physalis alkekengi L. Extracts In Vitro and In Vivo: Potential Application for Skin Care. Evid. Based Complement. Altern. Med. 2022, 2022, 7579572. [Google Scholar] [CrossRef]
- Vicas, L.G.; Jurca, T.; Baldea, I.; Filip, G.A.; Olteanu, D.; Clichici, S.V.; Pallag, A.; Marian, E.; Micle, O.; Crivii, C.B.; et al. Physalis alkekengi L. Extract Reduces the Oxidative Stress, Inflammation and Apoptosis in Endothelial Vascular Cells Exposed to Hyperglycemia. Molecules 2020, 25, 3747. [Google Scholar] [CrossRef]
- Liu, M.; Liu, G.; Ma, Z.; Wen, J.L.; Liu, Y.; Sun, L.; Ren, X. A comprehensive quality evaluation method of different medicinal parts of Physalis Calyx seu Fructus by fingerprints, chemometrics, antioxidant activity, network pharmacology and molecular docking. Biomed. Chromatogr. 2023, 37, e5701. [Google Scholar] [CrossRef] [PubMed]
- Chedea, V.S.; Pintea, A.; Bunea, A.; Braicu, C.; Stanila, A.; Socaciu, C. Physalis alkekengi carotenoidic extract inhibitor of soybean lipoxygenase-1 activity. Biomed Res. Int. 2014, 2014, 589168. [Google Scholar] [CrossRef] [PubMed]
- Sak, K. Cytotoxicity of dietary flavonoids on different human cancer types. Pharmacogn. Rev. 2014, 8, 122–146. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Zhu, M.E.I.; Feng, K.A.I.; Sun, L. Purification, characterization and in vitro antioxidant activities of polysaccharide fractions isolated from the fruits of Physalis alkekengi L. J. Food Biochem. 2011, 35, 524–541. [Google Scholar] [CrossRef]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, M.S.; et al. Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects. Molecules 2022, 27, 233. [Google Scholar] [CrossRef]
- Sun, L.; Xu, G.; Dong, Y.; Li, M.; Yang, L.; Lu, W. Quercetin Protects Against Lipopolysaccharide-Induced Intestinal Oxidative Stress in Broiler Chickens through Activation of Nrf2 Pathway. Molecules 2020, 25, 1053. [Google Scholar] [CrossRef]
- Boronat, A.; Rodriguez-Morató, J.; Serreli, G.; Fitó, M.; Tyndale, R.F.; Deiana, M.; de la Torre, R. Contribution of Biotransformations Carried Out by the Microbiota, Drug-Metabolizing Enzymes, and Transport Proteins to the Biological Activities of Phytochemicals Found in the Diet. Adv. Nutr. 2021, 12, 2172–2189. [Google Scholar] [CrossRef]





| Physalis alkekengi L. Extracts | TEAC [mg/mL ± SDa] b | DPPH [IC50 (mg/mL) ± SDa] |
|---|---|---|
| PACD | 2.382 ± 0.020 | 0.017 ± 0.003 |
| PAFD | 0.720 ± 0.004 | 0.502 ± 0.010 |
| PACE | 1.749 ± 0.010 | 0.033 ± 0.003 |
| PAFE | 0.731 ± 0.002 | 0.670 ± 0.020 |
| Quercetin 3-O-glucoside | 2.426 ± 0.010 | / |
| Vitamin C | / | 0.170 ± 0.020 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crescenzi, M.A.; Serreli, G.; Deiana, M.; Tuberoso, C.I.G.; Montoro, P.; Piacente, S. Metabolite Profiling, through LC-ESI/LTQOrbitrap/MS Analysis, of Antioxidant Extracts from Physalis alkekengi L. Antioxidants 2023, 12, 2101. https://doi.org/10.3390/antiox12122101
Crescenzi MA, Serreli G, Deiana M, Tuberoso CIG, Montoro P, Piacente S. Metabolite Profiling, through LC-ESI/LTQOrbitrap/MS Analysis, of Antioxidant Extracts from Physalis alkekengi L. Antioxidants. 2023; 12(12):2101. https://doi.org/10.3390/antiox12122101
Chicago/Turabian StyleCrescenzi, Maria Assunta, Gabriele Serreli, Monica Deiana, Carlo I. G. Tuberoso, Paola Montoro, and Sonia Piacente. 2023. "Metabolite Profiling, through LC-ESI/LTQOrbitrap/MS Analysis, of Antioxidant Extracts from Physalis alkekengi L." Antioxidants 12, no. 12: 2101. https://doi.org/10.3390/antiox12122101